
Protection Induced by Vaccination with Recombinant Baculovirus and Virus-like Particles Expressing Toxoplasma gondii Rhoptry Protein 18
 and
and 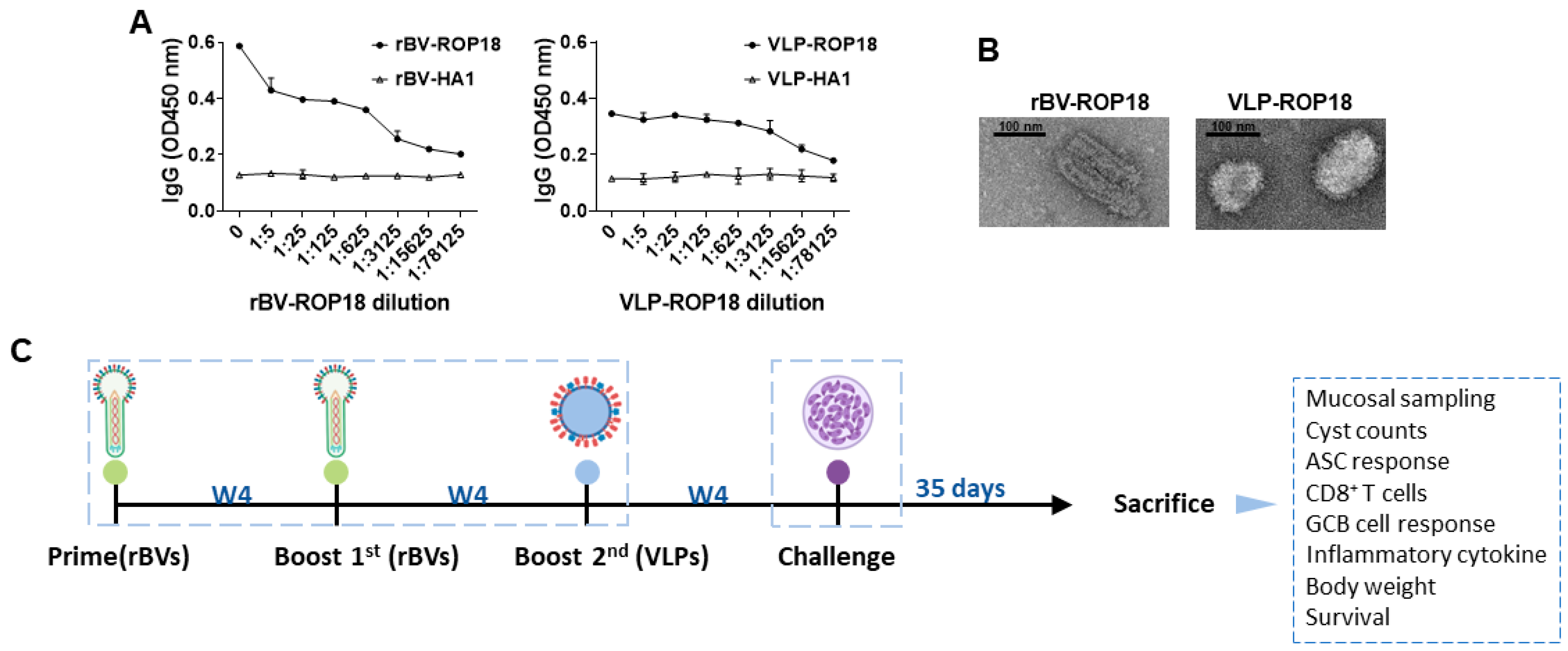{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Cells, Parasites, and Antibodies
2.3. Recombinant Baculovirus (rBV) and VLP Vaccine Generation
2.4. Vaccination and Challenge Infection Schedule
2.5. Sample Preparation
2.6. Antibody Reaction
2.7. Antibody Secreting Cell (ASC) Response
2.8. Flow Cytometry of Immune Cell Populations
2.9. Inflammatory Cytokine Analysis
2.10. Parasite Burden
2.11. Statistical Analysis
3. Results
3.1. Characterization of Vaccines and Animal Experimental Schedule
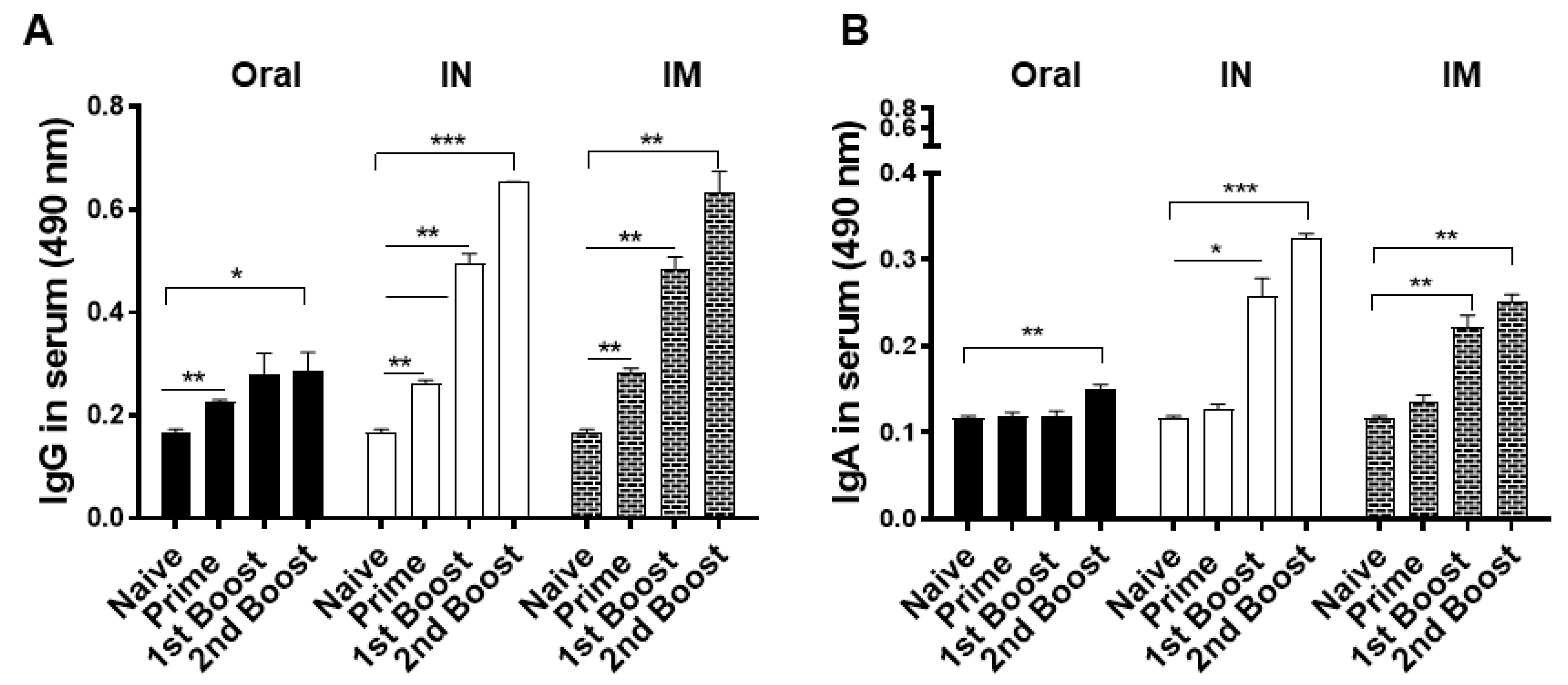
3.2. Antibody Response in Immune Serum
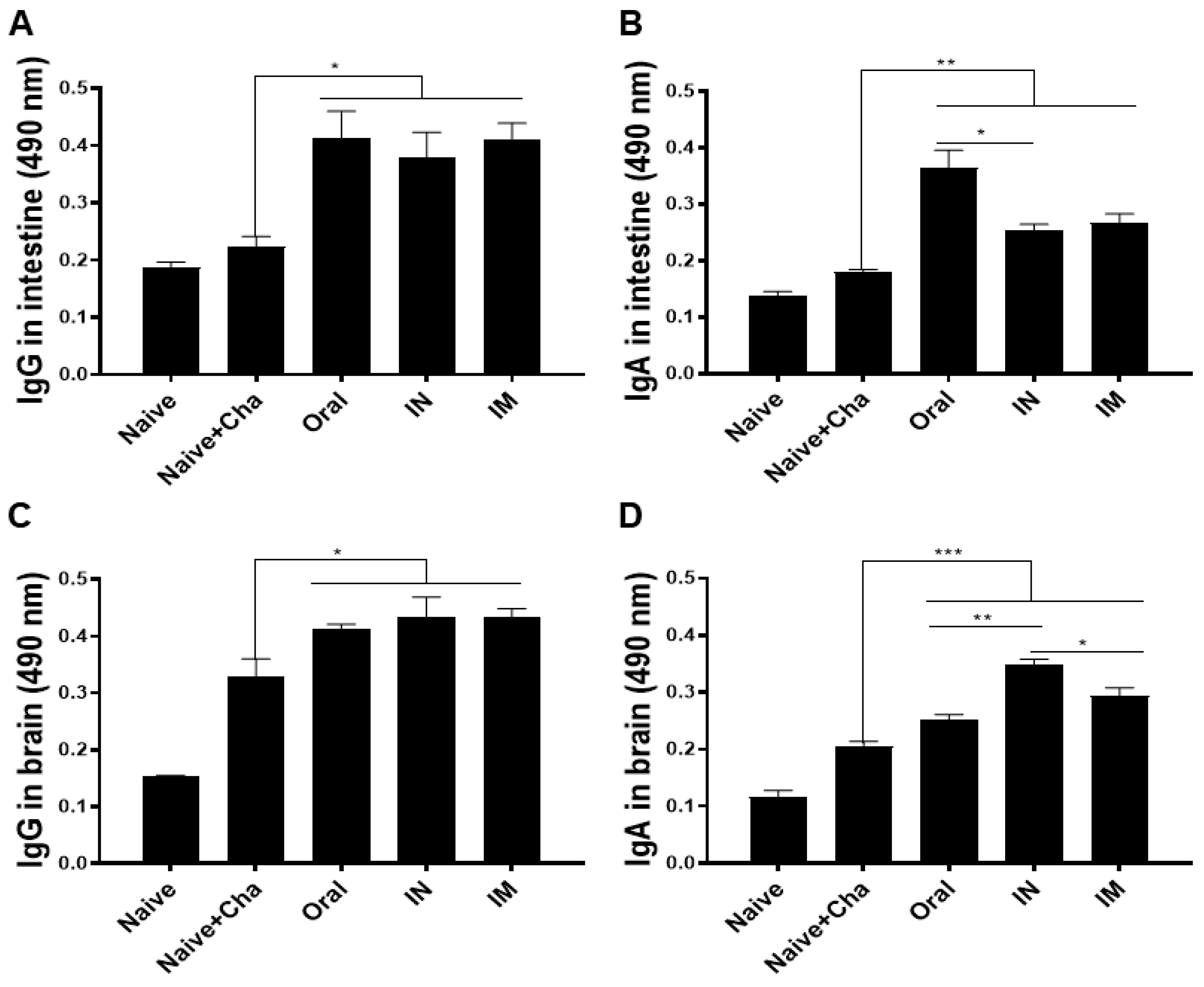
3.3. ROP18-Expressing Vaccines Induce Parasite-Specific Antibody Responses in the Intestines and the Brain
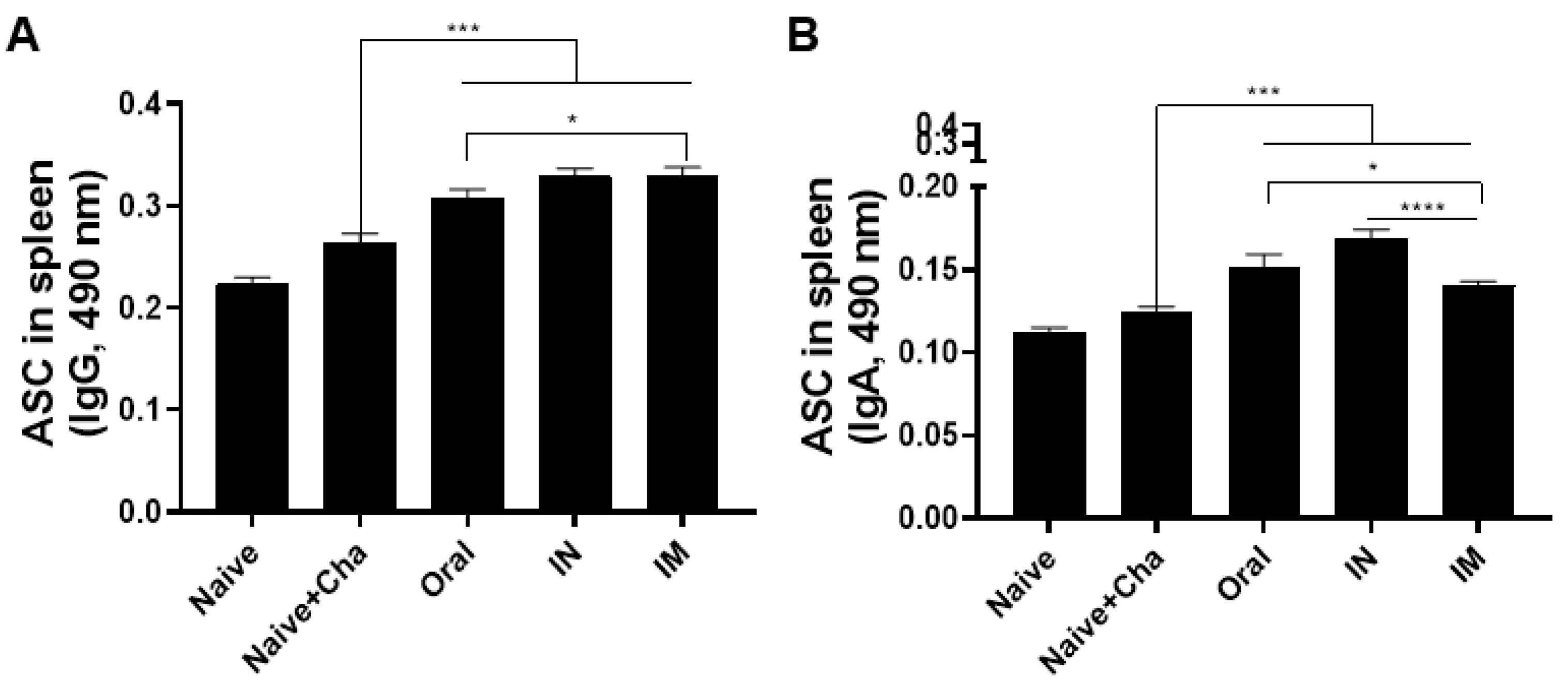
3.4. Vaccination Induces Antibody-Secreting Cell Responses
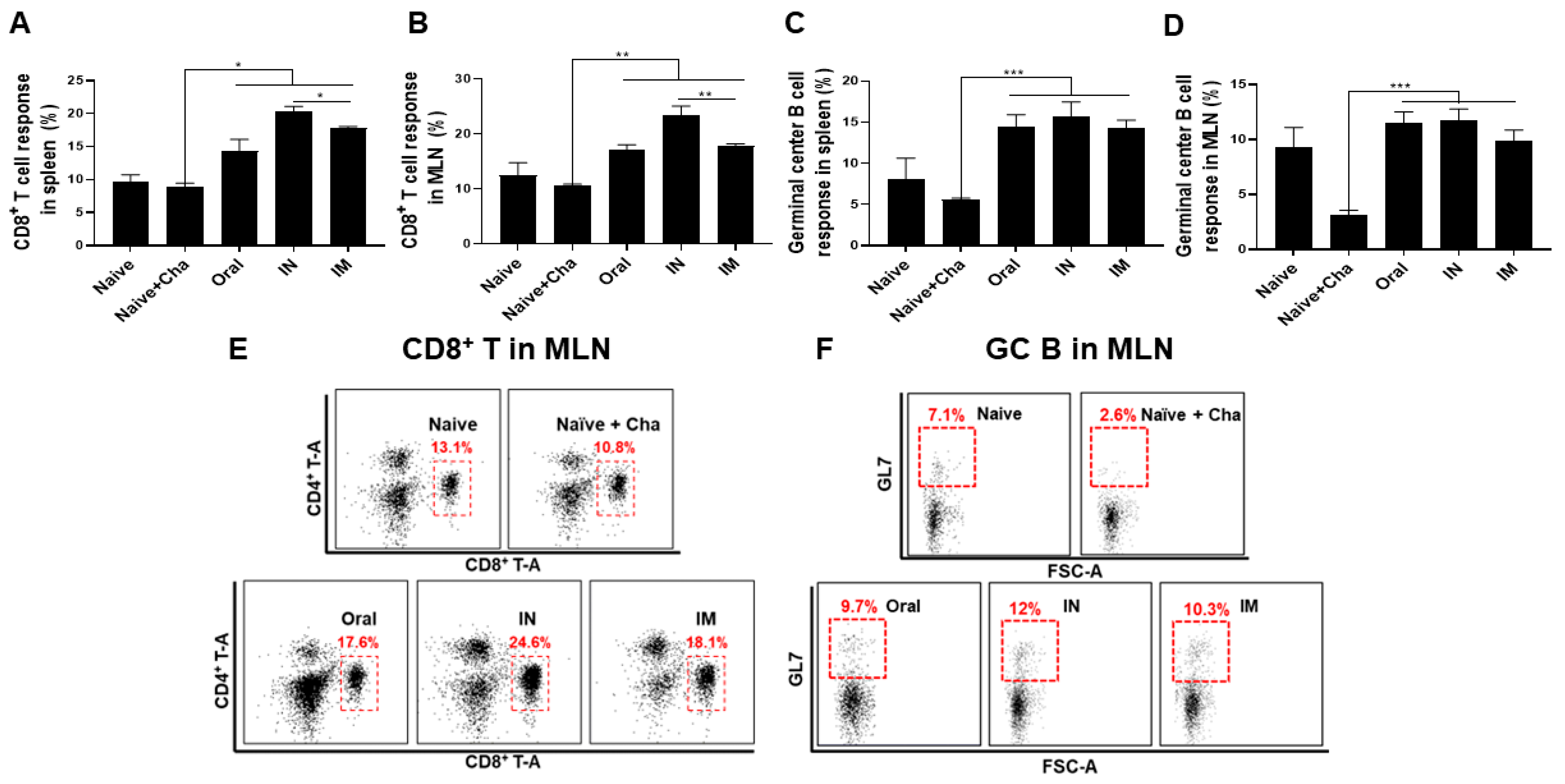
3.5. Activation of CD8+ T Cells and Germinal Center (GC) B Cells in the Spleen and MLN
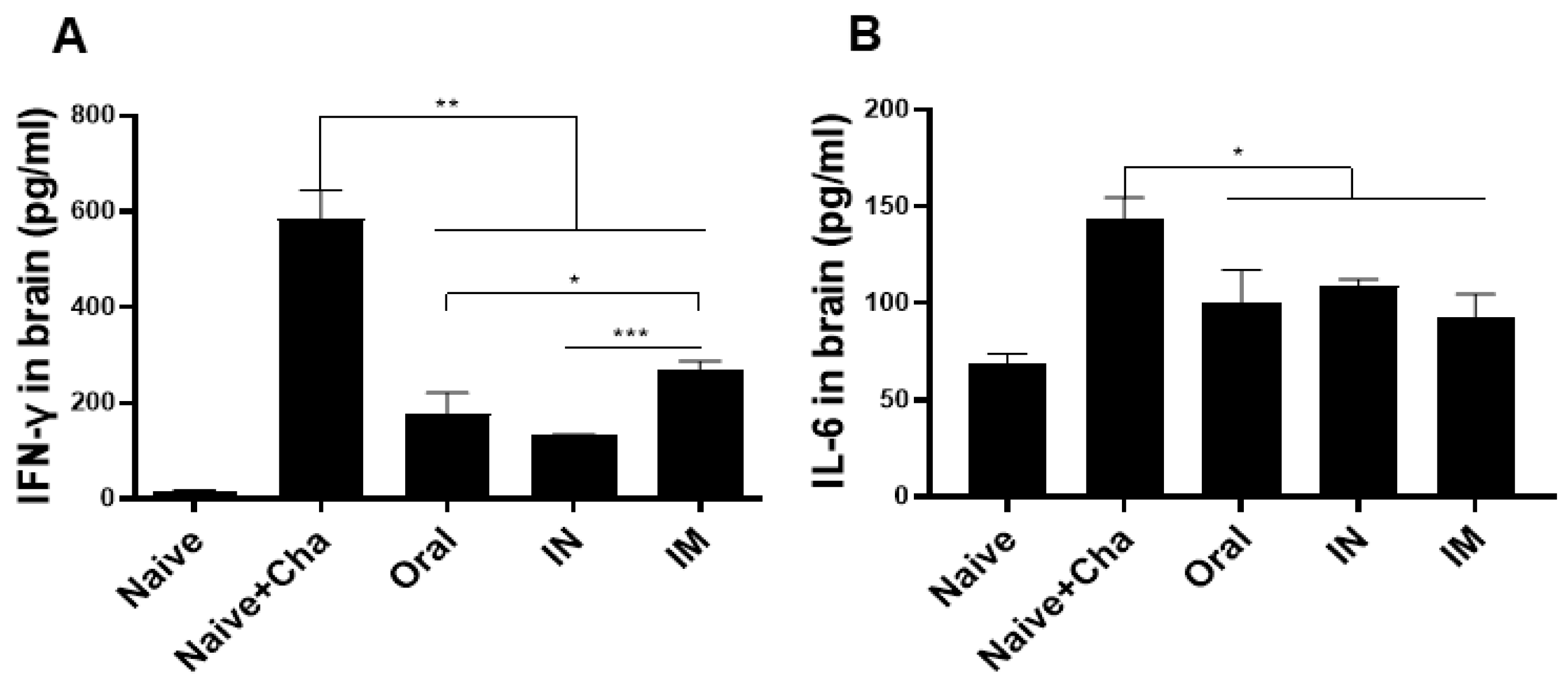
3.6. Pro-Inflammatory Cytokine Response in the Brain
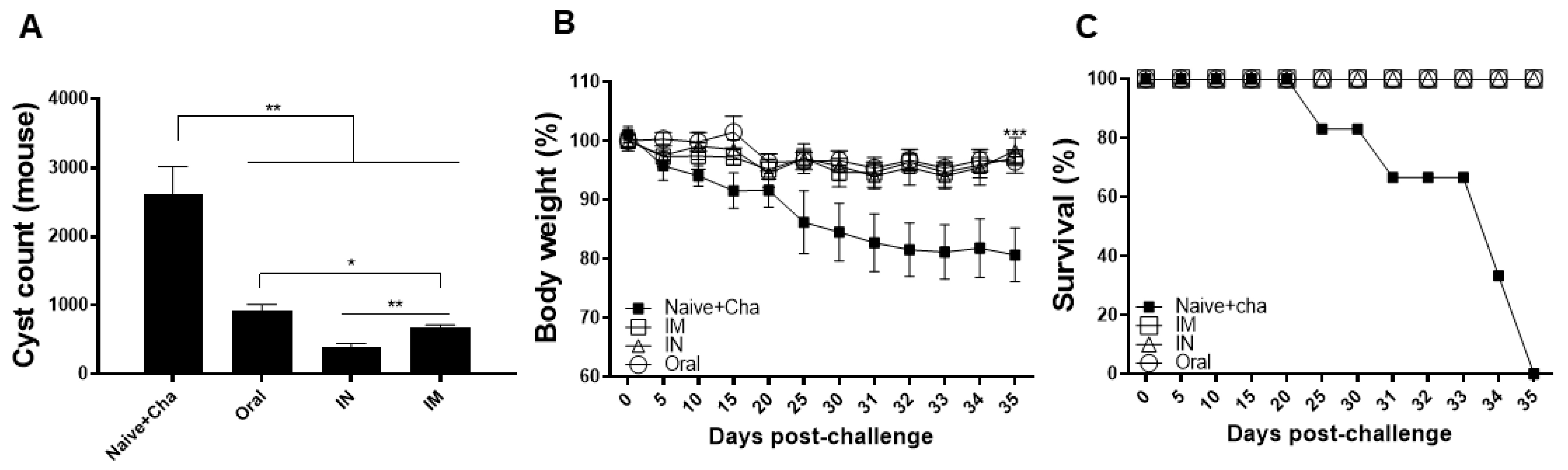
3.7. Protective Efficacy of the Heterologous Immunization Strategy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, L.; Wang, W.; Wang, S. Effect of vaccine administration modality on immunogenicity and efficacy. Vaccines 2015, 14, 1509–1523. [Google Scholar] [CrossRef] [PubMed]
- Vasu, N.; Ghaffari, G.; Craig, E.T.; Craig, T. Adverse events associated with intranasal influenza vaccine in the United States. Ther. Adv. Respir. Dis. 2008, 2, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination strategies for mucosal immune responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef]
- Dabritz, H.; Conrad, P.A. Cats and Toxoplasma: Implications for public health. Zoonoses Public Health 2010, 57, 34–52. [Google Scholar] [CrossRef]
- Innes, E.A.; Hamilton, C.; Garcia, J.L.; Chryssafidis, A.; Smith, D.J.F.; Parasitology, W. A one health approach to vaccines against Toxoplasma gondii. Food Waterborne Parasitol. 2019, 15, e00053. [Google Scholar] [CrossRef]
- Konstantinovic, N.; Guegan, H.; Stäjner, T.; Belaz, S.; Robert-Gangneux, F.J.F. Treatment of toxoplasmosis: Current options and future perspectives. Food Waterborne Parasitol. 2019, 15, e00036. [Google Scholar] [CrossRef]
- Madhan, S.; Prabakaran, M.; Kwang, J. Baculovirus as vaccine vectors. Curr. Gene Ther. 2010, 10, 201–213. [Google Scholar] [CrossRef]
- Fang, R.; Feng, H.; Nie, H.; Wang, L.; Tu, P.; Song, Q.; Zhou, Y.; Zhao, J. Construction and immunogenicity of pseudotype baculovirus expressing Toxoplasma gondii SAG1 protein in BALB/c mice model. Vaccine 2010, 28, 1803–1807. [Google Scholar] [CrossRef]
- Fang, R.; Feng, H.; Hu, M.; Khan, M.K.; Wang, L.; Zhou, Y.; Zhao, J. Evaluation of immune responses induced by SAG1 and MIC3 vaccine cocktails against Toxoplasma gondii. Vet. Parasitol. 2012, 187, 140–146. [Google Scholar] [CrossRef]
- Yoon, K.-W.; Chu, K.-B.; Kang, H.-J.; Kim, M.-J.; Eom, G.-D.; Lee, S.-H.; Moon, E.-K.; Quan, F.-S. Mucosal Administration of Recombinant Baculovirus Displaying Toxoplasma gondii ROP4 Confers Protection Against T. gondii Challenge Infection in Mice. Front. Cell. Infect. Microbiol. 2021, 11, 735191. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-J.; Chu, K.-B.; Kim, M.-J.; Park, H.; Jin, H.; Lee, S.-H.; Moon, E.-K.; Quan, F.-S. Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations. Pharmaceutics 2020, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Mendes, É.A.; Fonseca, F.G.; Casério, B.M.; Colina, J.P.; Gazzinelli, R.T.; Caetano, B.C. Recombinant vaccines against T. gondii: Comparison between homologous and heterologous vaccination protocols using two viral vectors expressing SAG1. PLoS ONE 2013, 8, e63201. [Google Scholar] [CrossRef] [PubMed]
- Niedelman, W.; Gold, D.A.; Rosowski, E.E.; Sprokholt, J.K.; Lim, D.; Farid Arenas, A.; Melo, M.B.; Spooner, E.; Yaffe, M.B.; Saeij, J.P. The rhoptry proteins ROP18 and ROP5 mediate Toxoplasma gondii evasion of the murine, but not the human, interferon-gamma response. PLoS Pathog. 2012, 8, e1002784. [Google Scholar] [CrossRef]
- Behnke, M.S.; Khan, A.; Lauron, E.J.; Jimah, J.R.; Wang, Q.; Tolia, N.H.; Sibley, L.D. Rhoptry proteins ROP5 and ROP18 are major murine virulence factors in genetically divergent South American strains of Toxoplasma gondii. PLoS Genet. 2015, 11, e1005434. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Chu, K.-B.; Kang, H.-J.; Quan, F.-S. Virus-like particles containing multiple antigenic proteins of Toxoplasma gondii induce memory T cell and B cell responses. PLoS ONE 2019, 14, e0220865. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kang, H.-J.; Lee, D.-H.; Kang, S.-M.; Quan, F.-S. Virus-like particle vaccines expressing Toxoplasma gondii rhoptry protein 18 and microneme protein 8 provide enhanced protection. Vaccine 2018, 36, 5692–5700. [Google Scholar] [CrossRef]
- Kang, H.-J.; Lee, S.-H.; Kim, M.-J.; Chu, K.-B.; Lee, D.-H.; Chopra, M.; Choi, H.-J.; Park, H.; Jin, H.; Quan, F.-S. Influenza virus-like particles presenting both Toxoplasma gondii ROP4 and ROP13 Enhance Protection against T. gondii infection. Pharmaceutics 2019, 11, 342. [Google Scholar] [CrossRef]
- Min, J.; Qu, D.; Li, C.; Song, X.; Zhao, Q.; Li, X.-A.; Yang, Y.; Liu, Q.; He, S.; Zhou, H.J.V. Enhancement of protective immune responses induced by Toxoplasma gondii dense granule antigen 7 (GRA7) against toxoplasmosis in mice using a prime-boost vaccination strategy. Vaccine 2012, 30, 5631–5636. [Google Scholar] [CrossRef]
- Yu, L.; Yamagishi, J.; Zhang, S.; Jin, C.; Aboge, G.O.; Zhang, H.; Zhang, G.; Tanaka, T.; Fujisaki, K.; Nishikawa, Y. Protective effect of a prime-boost strategy with plasmid DNA followed by recombinant adenovirus expressing TgAMA1 as vaccines against Toxoplasma gondii infection in mice. Parasitol. Int. 2012, 61, 481–486. [Google Scholar] [CrossRef]
- Zhang, G.; Huong, V.T.; Battur, B.; Zhou, J.; Zhang, H.; Liao, M.; Kawase, O.; Lee, E.G.; Dautu, G.; Igarashi, M.; et al. A heterologous prime-boost vaccination regime using DNA and a vaccinia virus, both expressing GRA4, induced protective immunity against Toxoplasma gondii infection in mice. Parasitology 2007, 134, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Liu, Q.; Liu, W.; Men, J.; Gao, S.; Jiang, L.; Wang, Z.; Zhai, Y.; Jin, H.; Lian, H.; et al. Protection in mice immunized with a heterologous prime-boost regime using DNA and recombinant pseudorabies expressing TgSAG1 against Toxoplasma gondii challenge. Vaccine 2009, 27, 2741–2745. [Google Scholar] [CrossRef]
- Kiyono, H.; Fukuyama, S. NALT-versus Peyer’s-patch-mediated mucosal immunity. Nat. Rev. Immunol. 2004, 4, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Rashid, I.; Moiré, N.; Héraut, B.; Dimier-Poisson, I.; Mévélec, M.-N. Enhancement of the protective efficacy of a ROP18 vaccine against chronic toxoplasmosis by nasal route. Med. Microbiol. Immunol. 2017, 206, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.A.; Hwang, S.; Moretto, M. Toxoplasma gondii: CD8 T cells cry for CD4 help. Front. Cell. Infect. Microbiol. 2019, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.-G.; Zhang, X.-X.; Lin, R.-Q.; Petersen, E.; He, S.; Yu, M.; He, X.-H.; Zhou, D.-H.; He, Y.; Li, H.-X. Protective effect against toxoplasmosis in mice induced by DNA immunization with gene encoding Toxoplasma gondii ROP18. Vaccine 2011, 29, 6614–6619. [Google Scholar] [CrossRef]
- Dimier-Poisson, I.; Aline, F.; Mévélec, M.-N.; Beauvillain, C.; Buzoni-Gatel, D.; Bout, D. Protective mucosal Th2 immune response against Toxoplasma gondii by murine mesenteric lymph node dendritic cells. Infect. Immun. 2003, 71, 5254–5265. [Google Scholar] [CrossRef]
- Mack, D.; McLeod, R. Human Toxoplasma gondii-specific secretory immunoglobulin A reduces T. gondii infection of enterocytes in vitro. J. Clin. Investig. 1992, 90, 2585–2592. [Google Scholar] [CrossRef]
- Parlog, A.; Schlüter, D.; Dunay, I.R. Toxoplasma gondii-induced neuronal alterations. Parasite Immunol. 2015, 37, 159–170. [Google Scholar] [CrossRef]
- Galván-Ramírez, M.d.l.L.; Salas-Lais, A.G.; Dueñas-Jiménez, S.H.; Mendizabal-Ruiz, G.; Franco Topete, R.; Berumen-Solís, S.C.; Rodríguez Pérez, L.R.; Franco Topete, K. Kinematic locomotion changes in C57BL/6 mice infected with toxoplasma strain ME49. Microorganisms 2019, 7, 573. [Google Scholar] [CrossRef] [Green Version]
- Bergersen, K.V.; Barnes, A.; Worth, D.; David, C.; Wilson, E.H. Targeted transcriptomic analysis of C57BL/6 and BALB/c mice during progressive chronic Toxoplasma gondii infection reveals changes in host and parasite gene expression relating to neuropathology and resolution. Front. Cell. Infect. Microbiol. 2021, 11, 645778. [Google Scholar] [CrossRef] [PubMed]
- Herzog, C.J. Influence of parenteral administration routes and additional factors on vaccine safety and immunogenicity: A review of recent literature. Expert Rev. Vaccines 2014, 13, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, R.; Canpolat, F.E.; Yurttutan, S.; Oncel, M.Y.; Erdeve, O.; Dilmen, U. Effect of needle length for response to hepatitis B vaccine in macrosomic neonates: A prospective randomized study. Vaccine 2012, 30, 3155–3158. [Google Scholar] [CrossRef] [PubMed]
- Middleman, A.B.; Anding, R.; Tung, C.J.P. Effect of needle length when immunizing obese adolescents with hepatitis B vaccine. Pediatrics 2010, 125, e508–e512. [Google Scholar] [CrossRef]
- Diggle, L.; Deeks, J.J.B. Effect of needle length on incidence of local reactions to routine immunisation in infants aged 4 months: Randomised controlled trial. BmJ 2000, 321, 931–933. [Google Scholar] [CrossRef]
- Jackson, L.A.; Starkovich, P.; Dunstan, M.; Yu, O.; Nelson, J.; Dunn, J.; Rees, T.; Zavitkovsky, A.; Maus, D.; Froeschle, J.E. Prospective assessment of the effect of needle length and injection site on the risk of local reactions to the fifth diphtheria-tetanus-acellular pertussis vaccination. Pediatrics 2008, 121, e646–e652. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, K.-W.; Chu, K.-B.; Kang, H.-J.; Kim, M.-J.; Eom, G.-D.; Mao, J.; Lee, S.-H.; Ahmed, M.A.; Quan, F.-S. Protection Induced by Vaccination with Recombinant Baculovirus and Virus-like Particles Expressing Toxoplasma gondii Rhoptry Protein 18. Vaccines 2022, 10, 1588. https://doi.org/10.3390/vaccines10101588
Yoon K-W, Chu K-B, Kang H-J, Kim M-J, Eom G-D, Mao J, Lee S-H, Ahmed MA, Quan F-S. Protection Induced by Vaccination with Recombinant Baculovirus and Virus-like Particles Expressing Toxoplasma gondii Rhoptry Protein 18. Vaccines. 2022; 10(10):1588. https://doi.org/10.3390/vaccines10101588
Chicago/Turabian StyleYoon, Keon-Woong, Ki-Back Chu, Hae-Ji Kang, Min-Ju Kim, Gi-Deok Eom, Jie Mao, Su-Hwa Lee, Md Atique Ahmed, and Fu-Shi Quan. 2022. "Protection Induced by Vaccination with Recombinant Baculovirus and Virus-like Particles Expressing Toxoplasma gondii Rhoptry Protein 18" Vaccines 10, no. 10: 1588. https://doi.org/10.3390/vaccines10101588