
Plant Immunity against Tobamoviruses

1
MOE Key Laboratory of Bioinformatics and Center for Plant Biology, School of Life Sciences, Tsinghua University, Beijing 100084, China
2
Tsinghua-Peking Center for Life Sciences, Beijing 100084, China
*
Authors to whom correspondence should be addressed.
Viruses 2024, 16(4), 530; https://doi.org/10.3390/v16040530
Submission received: 1 March 2024
/
Revised: 26 March 2024
/
Accepted: 27 March 2024
/
Published: 29 March 2024
(This article belongs to the Special Issue Tobamoviruses: Molecular Aspects and Resistance Regulation—a Special Issue Commemorating 125 Years of Research on Tobamoviruses)
Abstract
:Tobamoviruses are a group of plant viruses that pose a significant threat to agricultural crops worldwide. In this review, we focus on plant immunity against tobamoviruses, including pattern-triggered immunity (PTI), effector-triggered immunity (ETI), the RNA-targeting pathway, phytohormones, reactive oxygen species (ROS), and autophagy. Further, we highlight the genetic resources for resistance against tobamoviruses in plant breeding and discuss future directions on plant protection against tobamoviruses.
1. Introduction
Tobamoviruses are a group of plant viruses belonging to the Tobamovirus genus, which is part of the Virgaviridae family. The Tobamovirus genus includes 37 members [1]. The tobamovirus genome is a single-stranded, positive-sense RNA of about 6.4 kb size that is encapsidated in rod-shaped particles. It encodes four viral proteins, including two subunits of the viral RNA-dependent RNA polymerase-a 5′, where there is a proximal one of 122–130 kDa and a translational read through of 178–183 kDa from the first open reading frame (ORF); a movement protein (MP); and a coat protein (CP) [2]. In addition, tobacco mosaic virus (TMV) contains two additional ORFs which potentially encode a 54 kDa and 4.8 kDa protein, respectively [3,4]. Tobamoviruses infect a wide range of plant species and cause significant damage to a wide range of economically important crops, such as tomato, pepper, cucumber, and tobacco.
In this review, we discuss the plant immunity responses to tobamoviruses, including pattern-triggered immunity (PTI), effector-triggered immunity (ETI), RNA-targeting pathway, phytohormones, reactive oxygen species (ROS), and autophagy. Further, we highlight the genetic resources for resistance against tobamoviruses in plant breeding and provide future directions on plant protection against tobamoviruses.
2. Plant Defense against Tobamoviruses
Based on the studies on plant interactions with bacterial and fungal pathogens, plants are thought to employ a two–layer immune system consisting of PTI and ETI. PTI is triggered by pathogen-associated molecular patterns (PAMPs) via cell surface-localized pattern-recognition receptors (PRRs), whereas ETI is activated by pathogen effector proteins via predominantly intracellularly localized receptors called nucleotide-binding, leucine-rich repeat immune receptors (NLRs) [5]. The concept of PTI and ETI could also be applicable to plant-virus interactions.
PTI is triggered by PRRs to detect the conserved microbial cues known as PAMPs. Classic PRRs are localized to the plasma membrane (PM) and play a crucial role in plant immunity against extracellular fungi and bacterial pathogens [6]. As viruses are intracellular parasites, it seems unlikely that there is a classic cell-surface-localized PRR-mediated PTI against intracellular viruses. However, antiviral PTI has been proposed because some viral proteins suppress PTI responses activated by non-viral PAMPs, and some receptor-like kinases are involved in basal antiviral defense [7]. For example, Brassinosteroid-associated kinase 1 (BAK1), the core regulator of PTI, contributes to plant defense against several RNA viruses, including two tobamoviruses, TMV and oilseed rape mosaic virus [8]. Viral double-stranded RNAs (dsRNAs) have been proposed to be the potential PAMPs that induce the antiviral defense response. Indeed, plant treatment with the purified viral dsRNA from virus-infected plants and the dsRNA analog polyinosinic:polycytidylic acid triggers typical PTI responses and antiviral defense, which depend on the PTI co-receptor SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE 1 [8]. Small RNAs (sRNAs) stimulate the production of callose near plasmodesmata (PD), consequently restricting the spread of viruses between plant cells. This defense response relies on various PTI signaling elements and several PD-localized proteins [9]. The expression of TMV MP suppresses dsRNA-induced callose deposition, facilitating intercellular TMV movement [10]. In addition, Ca2+ flux triggered by injuries to plant cells is thought to be the common molecular pattern of different viral infections which primes antiviral RNAi defense [11]. These findings suggest the existence of non-classic rather than classic PTI against plant viruses, including tobamoviruses.
Plant NLRs play an important role in plant antiviral ETI by detecting viral effectors. These NLRs can recognize viral effectors and trigger antiviral ETI, usually including hypersensitive response (HR), a type of programmed cell death at pathogen infection sites [12]. Currently, most plant genetic resources for plant breeding are mainly controlled by the naturally identified dominant resistance genes including NLR genes, Tm-1, and recessive resistance genes (see Figure 1 and Table 1).
3. Plant Genetic Resources for Resistance against Tobamoviruses
3.1. The Tobacco N Gene
The N gene is the first resistance (R) gene identified in Nicotiana glutinosa [13,14]. The N gene confers resistance to all known tobamoviruses except TMV-Ob [15,16]. It is a toll-interleukin-1 receptor homology/nucleotide binding/leucine rich repeat (TIR-NB-LRR, TNL) class of resistance gene [17]. The structure-function analysis shows that all three domains of the N protein, including TIR, NB, and LRR, are necessary for its function [18]. The N gene encodes two transcripts, NS and NL, by alternative splicing, both of which are essential for the full resistance to TMV [19]. TMV induces HR lesions in N-containing plants. In N-containing plants, the burst of reactive oxygen intermediates occurs rapidly upon TMV infection [20]. Similarly, nitric oxide accumulates at the beginning of HR during TMV infection [21].
The N protein recognizes the 50 kDa helicase domain (p50) within the 126 kDa replicase subunit of tobamoviruses through its TIR domain, triggering the HR and immune response in the cytoplasm [22,23,24,25]. Simultaneously, the N protein also functions in the resistance to TMV in the cell nucleus [23,26]. Extensive research has revealed the roles of host regulators in N-mediated resistance primarily through their interaction with the N protein. The transcription factor SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 6 (SPL6) associates with the N protein within distinct nuclear compartments, and it is essential for N-mediated resistance against TMV. In particular, the N-SPL6 interaction is present in the nucleus and is detected only when the p50 is present. It suggests that the association of N with SPL6 only occurs after an active defense response [26]. Similar to other TNLs, the N function is ENHANCED DISEASE SUSCEPTIBILITY (EDS1)-dependent [27,28]. N requirement gene 1 (NRG1) serves as a helper NLR and is required for N-mediated resistance [29]. Rar1 is required for N function. The tobacco Rar1 interacts with SGT1, a novel subunit of the SCF-type (Skp1/Cullin/F box protein) E3 ubiquitin ligase complex involved in protein degradation. SGT1 and Rar1 associate with Heat shock protein 90 (Hsp90), which interacts with the N protein [30]. Moreover, Hsp90 suppression compromises N-mediated resistance to TMV [31]. Similarly, the Hsp40-like Dna-J domain movement protein-interacting proteins (MIP1s) interact with SGT1 and are required for N-mediated resistance [32]. N receptor-interacting protein (NRIP1), a functional rhodanese sulfurtransferase, has been identified to directly interact with both the N TIR domain and TMV p50, which is necessary for a complete resistance to TMV. NRIP1, which is normally localized in chloroplasts, is recruited to the cytoplasm and nucleus by the p50 effector. Consequently, NRIP1 interacts with N only in the presence of p50 [33]. UBR7, a putative E3 ubiquitin ligase, directly interacts with the N protein via its TIR domain, and it also negatively regulates the level of the N protein. The downregulation of UBR7 increases the protein level of N and enhances TMV resistance. Moreover, TMV p50 disrupts the N-UBR7 interaction and relieves the negative regulation on N [34]. In addition, the MEKK1-like mitogen-activated protein kinase kinase kinase NPK1, MEK1 MAPKK, NTF6 MAPK, and WRKY/MYB transcription factors are essential for N-mediated resistance [35,36]. Transcription factor alfin-like 7 (AL7) interacts with N and inhibits the transcription of genes involved in ROS scavenging to positively regulate N-mediated resistance to TMV [37]. Mitogen-activated protein kinases (MAPKs), salicylic acid-induced protein kinase (SIPK), and wound-induced protein kinase (WIPK) interact with and phosphorylate AL7, which inhibits AL7-N interaction and enhances its DNA binding activity, thus promoting ROS accumulation and enabling an immune response to TMV [37].
3.2. The Tobacco N′ Gene
The N′ gene from N. sylvestris encodes a coiled-coil (CC) domain-containing NLR (CNL) immune receptor that confers resistance against tobamoviruses accompanying the HR by recognizing viral CP [38,39,40,41]. N′ is an ortholog of the pepper L genes with a different recognition spectrum. N′ can confer resistance against tomato mosaic virus (ToMV), paprika mild mottle virus, pepper mild mottle virus (PMMoV), including the PMMoV pathotype P1,2,3,4 [41] and tomato brown rugose fruit virus (ToBRFV) [16].
3.3. The Tomato Tm-1 Gene
The Tm-1 gene is introgressed into the cultivated tomato species Solanum lycopersicum from the wild tomato species Solanum habrochaites S. Knapp & D.M. Spooner [42,43]. However, Tm-1-mediated ToMV resistance could be easily overcome by ToMV resistance breaking isolates [44]. In addition, Tm-1 suppresses the visible viral mosaic symptoms, but detectable virus multiplication still occurs. The inhibition of TMV multiplication is Tm-1 gene dose-dependent, whereas the suppression of visible symptoms is not [45,46].
The Tm-1 gene encodes a protein that binds to the 130 kDa subunit of the ToMV replicase and inhibits the RNA-dependent RNA replication of ToMV [47,48], suggesting that Tm-1 functions as a viral inhibitor. Moreover, structure analyses have indicated that Tm-1 shows no sequence homology to functionally characterized proteins [47,49]. These results indicate that Tm-1 differs from the previously identified resistance genes in plants.
Tm-1 binds ToMV replication proteins to inhibit the key events in replication complex formation on membranes, preceding negative-strand RNA synthesis. Three host proteins, Tobamovirus multiplication 1 (TOM1), TOM2A, and ADP-ribosylation factor-like 8 (ARL8), are required for ToMV RNA replication, and they are also suggested to be the components of the ToMV replication complex [50]. Upon ToMV infection, Tm-1 inhibits the formation of viral RNA replication complex on membranes by inhibiting the association of TOM1, TOM2A, and ARL8 with the ToMV 130 K replicase component [48].
3.4. The Tomato Tm-2/Tm-22 Gene
Tm-2 and Tm-22 are two alleles of the same gene in tomato that encodes a CNL [51]. They have been introgressed into cultivated tomato from the wild species Solanum peruvianum. Tm-2 and Tm-22 confer resistance to tobamoviruses, including TMV and ToMV, by recognizing the viral MP [42,52,53,54]. Tm-22 also displays resistance to tomato mottle mosaic virus (ToMMV) by recognizing the MP, and the resistance is regulated by the allele combinations and the temperature [55]. The homozygous tomato harboring Tm-22 and the heterozygous tomato containing Tm-22 and Tm-2, but not the heterozygous tomato containing Tm-22 and Tm-2, exhibit resistance to ToMMV. Tm-22-mediated resistance is compromised at 35 °C, but not at 30 °C or lower temperatures [55]. Previous studies showed that the C-terminal 30-amino acid deletion of viral MP compromised Tm-22-mediated resistance [54]. However, we showed that the N-terminus of MP is sufficient for inducing Tm-22-mediated HR [56]. Further, ToBRFV can overcome Tm-22–mediated resistance, and the elements required to evade Tm-22 are located in the N-terminus but not in the C terminus of ToBRFV MP, as proved by virus infection assays [57,58]. These combined data suggest that Tm-22 recognizes the N-terminal but not the C-terminal sequence of the viral MP for Tm-22 recognition, and the C-terminal domain of MP could affect the exposure of protein structures that are recognized by Tm-22. Tm-22 is previously thought to function on PD due to the predominant localization of its avirulence (Avr) protein MP. However, the Tm-22 function is independent of the localization of viral MP to PD. Further, Tm-22 has been found to localize to and function on the PM although it lacks any PM-localization motif [56]. The Tm-22 CC domain is the signaling domain and its self-association triggers a defense response, including HR. In the presence of viral MPs, Tm-22 self-associates and is activated, which requires the nucleotide-binding domain-mediated self-association of a CC domain in an (d)ATP-dependent manner [59]. Tm-22 stability is regulated by SGT1 and Hsp90 [60] and MIP1s [32]. MIP1s function as co-chaperones and are required for both TMV infection and plant immunity, including Tm-22-mediated resistance, by associating with SGT1 and Tm-22 [32]. Further, rubisco small subunit is required for both Tm-22-mediated extreme resistance and tobamovirus movement by interacting with viral MPs [61]. In addition, the Tm-22-mediated resistance response is dependent on its expression level: a high level of expression triggers extreme resistance without visible cell death; an intermediate level of expression triggers complete resistance with HR lesions at virus infection sites; and a low level of expression only confers a partial resistance with systemic viral infection and systemic necrosis throughout the plant [62].
3.5. The Pepper L Gene
The L gene confers resistance against tobamovirus and encodes a CNL, which are introgressed into Capsicum annuum from the wild pepper species [63]. Upon virus infection, the L protein recognizes viral CP, leading to HR [64]. There are four different alleles of L (L1, L2, L3, and L4). All L alleles confer resistance to P0 viruses, including ToMV, yellow pepper mild mottle virus, and chili pepper mild mottle virus. L1 only confers resistance to P0 viruses. L2 confers resistance to all P0 and P1 viruses, including the PMMoV J strain. L3 defends against P0, P1, and P1,2 viruses, including the PMMoV strains which can overcome L2. L4 defends against P0, P1, P1,2, and P1,2,3 PMMoV pathotypes which can overcome L3 [63,64,65]. However, some PMMoV strains can systemically infect all the identified L alleles of pepper plants. Another allele, L1a, is thermosensitive and does not confer resistance against tobamoviruses at elevated temperatures [66].
3.6. Tobamovirus Multiplication (TOM) and ARL8
Host susceptibility proteins help virus infection at different stages of the virus life cycle. Among them, TOBAMOVIRUS MULTIPLICATION (TOM) proteins play a critical role in tobamovirus infection by interacting with viral replication-associated proteins to help the formation of the viral replication complex [67]. TOM genes are critical for infection by tobamoviruses in various plant species [68]. TOM1 and TOM2A encode seven-pass and four-pass transmembrane proteins, respectively [69,70]. TOM1 was first cloned from Arabidopsis with three homologues, TOM1, TOM3, and THH1 [70,71]. In Arabidopsis, the tom1 single mutant partially impairs tobamovirus multiplication, and the double tom1 and tom3 mutant completely inhibits tobamovirus multiplication [72,73]. In double tom1/tom3 mutant lines overexpressing THH1, the level of tobamovirus CP is similar to that of wild-type plants, suggesting that THH1 could weakly contribute to tobamovirus multiplication due to its lower level of expression than that of TOM1 and TOM3 [71].
TOM1 homologs have also been found in tobacco and tomato [74,75]. Knockdown of TOM1 homologs dramatically inhibits TMV/ToMV multiplication without introducing any obvious growth defects [74,75,76]. In addition, tobacco TOM1 mutant lines have been found to be resistant to TMV [77]. In particular, the quadruple knockout of SlTOM1 homologs, which is generated by genome editing, confers resistance to ToBRFV in tomato [78]. Meanwhile, the double knockout of SlTOM1a and SlTOM3 confers resistance to ToBRFV, but not to ToMV and TMV [67].
TOM1 and TOM2A promote tobamovirus multiplication. Upon TMV infection, TOM1 interacts with TMV 126-kDa replicase to promote the assembly of viral replication complex formation on host membranes. TOM2A also facilitates the formation of the viral replication complex by interacting with TOM1 [68].
Arabidopsis ADP-ribosylation factor-like 8 (ARL8) is a small GTP-binding protein that interacts with TOM1. ARL8 also interacts with ToMV 180-kDa replicase and is required for tobamovirus multiplication. Upon tobamovirus infection, ARL8 and TOM1 are components of the replication complex and play crucial roles in the replication activation process, including replicase RNA synthesizing and capping [50,79].
3.7. WPRb
WPRb, a weak chloroplast movement under blue light 1 and plastid movement impaired 2 (WEB1/PMI2)-related protein family gene, is a recessive resistance gene associated with cucumber green mottle mosaic virus (CGMMV) resistance in watermelon. Genome editing of WPRb in N. benthamiana also confers a great tolerance to CGMMV. WPRb targets the PD and interacts with CGMMV MP to facilitate viral cell-to-cell movement by affecting PD permeability [80].

{kind=link}
{kind=link}
Table 1.
Plant genetic resources for resistance against tobamoviruses.
| Gene Name | Alleles | Plant of Origin | Viral Target | Protein Type |
|---|---|---|---|---|
| N | NA | Nicotiana glutinosa [13,14] | p50 (Avr) [24,25,26] | Toll-interleukin-1 receptor homology/nucleotide binding/leucine rich repeat (TNL) [17] |
| N′ | NA | Nicotiana sylvestris [41] | CP (Avr) [41] | coiled-coil domain-containing/nucleotide binding/leucine rich repeat (CNL) [41] |
| Tm-2 | Tm-2 and Tm-22 [51] | Solanum peruvianum [42,52] | MP (Avr) [54,56] | CNL [51] |
| L | L1, L1a, L2, L 3, and L4 [64,66] | Capsicum chinense [63] | CP (Avr) [64] | CNL [64] |
| Tm-1 | Several [49] | Solanum habrochaites S. Knapp & D.M. Spooner [42,43] | Replicase [44,46,49] | Unidentified |
| TOM1; TOM3 | NA | Arabidopsis [72,73] | Replicase [50,68,79] | Seven-pass transmembrane [70] |
| TOM2A | NA | Arabidopsis [69] | NA | Four-pass transmembrane [69] |
| ARL8 | NA | Arabidopsis [79] | Replicase [50,68,79] | N-terminal amphipathic helix [79] |
| WPRb | NA | Citrullus lanatus [80] | MP [80] | coiled-coil [80] |
Abbreviations: NA, not available.
4. RNA-Targeting Mechanisms
RNA silencing plays a key role in antiviral defense against all types of viruses [81,82]. RNA silencing is a sequence-specific process found in both plants and animals, which involves the generation of sRNAs [83,84]. In plants, RNA silencing is orchestrated by 21- to 24-nucleotide sRNA, which is categorized as small interfering RNAs (siRNAs) and microRNAs (miRNAs). These sRNAs are generated as duplexes with 2-nt 3′ overhangs from longer dsRNA precursors or hairpin-like secondary structures, respectively, through the action of Dicer-like (DCL) enzymes [85] (Figure 2). RNA silencing has been used to engineer complete resistance against tobamoviruses [86,87,88,89,90,91]. In the case of tobamoviruses, the small replicase subunit (122–130 kDa) could function as the viral suppressors of RNA silencing (VSR). In particular, the TMV 126 kDa protein is identified as the VSR by disrupting HUA enhancer 1 (HEN1)-mediated methylation of sRNAs to shield viral transcripts from the host RNA silencing pathway [92,93]. In addition, the 122-kDa replicase subunit (p122) of crucifer-infecting TMV (crTMV) is a potent VSR and compromises both siRNA- and miRNA-mediated pathways [94]. p122 is also reported to enhance the levels of miRNA 168 to inhibit the expression of Argonaute 1 (AGO1) [95]. In addition, during oilseed rape mosaic tobamovirus (ORMV) infection, ORMV p125 replicase is required for the inhibition of HEN1 activity to suppress RNA silencing [85]. On the other hand, TMV MP contributes to antiviral silencing during infection by enhancing the spread of RNA silencing signal, and this ability of TMV MP may contribute to the control of virus propagation in the infected host. The TMV 126 kDa replicase-associated protein and MP with contrast roles in RNA silencing may balance viral propagation at different infection stages [96].
Beyond RNA silencing, other RNA-targeting mechanisms have also been shown to be involved in antiviral defense including RNA decay [97]. RNA decay is an essential RNA quality control and gene regulatory mechanism in eukaryotes. It is initiated in the cytoplasm by mRNA deadenylation, followed by exosome complex-mediated exonucleolytic decay in the 3′-5′ direction or by decapping complex and exoribonuclease (XRN)-mediated decay in the 5′-3′direction [98]. A study suggests that TMV proteins (MP and CP) enhance transcriptional levels of RNA decay genes and induce RNA decay to impair antiviral RNA silencing for better virus infection [99]. However, silencing of NbXrn4 facilitates TMV systemic infection in N. benthamiana [100], suggesting that RNA decay may also play a role in antiviral defense against tobamovirus.
5. Phytohormone Interactions with Tobamoviruses
Plant hormone salicylic acid (SA), methyl salicylate (MeSA), jasmonic acid (JA), methyl jasmonate (MeJA), ethylene, and Auxin/indole-3 acetic acid (Aux/IAA) play important roles in plant–virus interactions. Among them, SA plays a critical role in plant defense against a broad spectrum of pathogens, including multiple viruses. SA interferes with different steps of the viral cycle. In tobacco leaves, SA treatment decreases TMV RNA accumulation by disrupting TMV replication in mesophyll cells [101,102]. SA also inhibits TMV cell-to-cell movement [102]. Reducing the early SA accumulation delays HR and promotes TMV dispersal during lesion formation in TMV-infected tobacco Samsun NN, suggesting that early SA accumulation is a key factor in preventing viral escape during N-mediated TMV resistance [103]. In addition, SA could also function its antiviral mechanism by activating RNA silencing. SA induces the expression of host RNA-dependent RNA polymerase1 (RDR1), which contributes to antiviral RNA silencing, thereby promotes the degradation of viral RNA to limit the infection by tobamovirus [104,105,106]. Further, SA upregulates the expression of RDR6 in N. tabacum [107] and RDR2 in tomato [108]. However, it does not affect the expression of RDR2 and RDR6 in Arabidopsis [109], nor does it affect RDR6 in N. glutinosa [110]. Besides SA, JA/MeJA, ABA, ET, and synthetic auxin upregulate RDR1 expression [111,112]. ABA treatment also increases the expression of RDR1, RDR2, and RDR6 when SA synthesis is impaired, suggesting that the antagonism between ABA and SA influences the expression of these genes [112]. N-mediated TMV resistance is compromised in transgenic NahG tobacco plants and NPR1-silenced plants [35,113], suggesting that SA is essential for N-mediated antiviral immunity.
TMV infection induces HR accompanied by the production of phytohormones, including SA and JA. Exogenous MeJA application to plants reduces local TMV resistance and permits systemic TMV movement [114], suggesting a negative role of MeJA in antiviral defense against TMV. However, silencing of JA biosynthetic and signaling genes in N. benthamiana plants increases susceptibility to TMV [115], suggesting a positive role of MeJA/JA in antiviral defense against TMV. The role of COI1 in N-mediated TMV resistance is also controversial. Silencing of COI1, the JA receptor gene, compromises N-mediated TMV resistance in transgenic N. benthamiana [35], suggesting that JA positively regulates N-mediated TMV resistance. However, silencing of COI1 reduces viral accumulation in N. tabacum Samsun NN, which possesses the N gene [114]. The contrast data may be a result of the differences in the experimental plant systems used, i.e., N. tabacum Samsun NN and N. benthamiana. Indeed, N. tabacum Samsun NN is reported to have a novel N gene-associated, temperature-independent resistance [116].
TMV infection triggers the plant release of several airborne compounds including (E)-2-octenal. (E)-2-octenal primes the JA/ET signaling pathway, including the upregulation of NbMYC2, NbERF1, and NbPDF1.2, and then upregulates the pathogenesis-related genes, such as NbPR1a, NbPR1b, NbPR2, and NbNPR1, to activate the antiviral defense against TMV in adjacent N. benthamiana plants [117].
Both MeSA and MeJA contribute to systemic resistance against TMV, possibly acting as the initiating signals for systemic resistance. Silencing of SA or JA biosynthetic and signaling genes in N. benthamiana plants increases susceptibility to TMV [115]. Silencing of either SABP2 or NAC2 compromises antiviral defense, suggesting that SA, but not MeSA, directly activates antiviral defense [118].
Ethylene plays a role in the antiviral defense against tobamoviruses. It increases watermelon resistance to CGMMV infection by inducing the expression of the AGO5 gene [119]. In addition, the ethylene pathway participates in transcription factor MYB4L-mediated resistance against TMV, and ethylene-induced MYB4L is involved in the TMV resistance in N. benthamiana [120]. Moreover, silencing of CTR1, an ethylene receptor, accelerates N-mediated HR [35], suggesting that ethylene signaling negatively regulates the N-mediated HR induced by TMV.
Auxin is a crucial plant hormone and participates in various processes. Aux/IAA proteins are vital components within this regulatory framework, with a primary function of translating auxin levels into gene expression [121]. The interaction between tobamoviruses and Aux/IAA was first reported to involve the helicase domain of TMV replicase and IAA26. The expression of TMV replicase disrupts the nuclear localization of IAA26, inhibiting its putative function as a transcriptional regulator of auxin-responsive genes for better viral symptoms and systemic movement [122,123,124]. TMV can reprogram auxin/IAA protein transcriptional responses and then enhances virus phloem loading [125].
6. ROS in Tobamovirus Infection
Upon pathogen infection, plants rapidly produce ROS to induce local or systemic signaling through the activation of cell surface-localized respiratory burst oxidase homolog (RBOH) proteins. ROS signaling mediates systemic resistance against plant viruses and is often considered as a positive regulator of plant antiviral defense [126,127]. Intact TMV virion and isolated TMV CPs trigger a rapid oxidative burst when added to the apoplast of tobacco epidermal cells. TMV CP stimulates host NAD(P)H oxidase-like activity [128]. Meanwhile, TMV infection increases the expression of ROS-scavenging related genes, including superoxide dismutases (CSD2), ascorbate peroxidase (APX1), and GDP-mannose pyrophosphorylase 1 (GMP1). Furthermore, silencing of GMP1 enhances the ROS level and reduces the TMV accumulation [129,130]. Thus, the ROS-scavenging pathway can also modulate the plant resistance against tobamoviruses. In addition, type-I non-specific lipid transfer protein (LTP1) is reported to enhance antiviral defense against TMV by upregulating SA biosynthesis and its downstream signaling components, while also suppressing ROS accumulation during the later stages of viral pathogenesis [131].
7. Autophagy in Tobamovirus Infection
Autophagy is a conserved vacuole/lysosome-dependent cellular process mediating the degradation of senescent/dysfunctional organelles or cytoplasmic materials into peptides or amino acids for reuse or storage [132]. Autophagy functions as an antiviral mechanism in plants [133,134]. It may also limit tobamovirus infection. Autophagy is activated upon infection by PMMoV or TMV [135,136]. PMMoV infection upregulates the expression of multiple autophagy-related genes (ATGs). Disruption of autophagy by autophagy inhibitor treatment and silencing of ATG genes increases PMMoV RNA accumulation and aggravates viral systemic symptoms [135]. Although increased autophagy by silencing of Cytoplastic Glyceraldehyde-3-Phosphate Dehydrogenase genes has no effect on GFP-tagged TMV infection, it promotes N-mediated HR cell death [137]. Further, autophagy suppression negatively regulates N-mediated cell death and promotes local TMV infection in N-containing plants [138]. Similarly, Bax inhibitor-1 positively regulates the autophagy triggerd by N activation upon TMV infection and negatively regulates N-mediated cell death [139]. In addition, autophagy also regulates the ROS level and PCD progress in TMV-infected tomato plants [140].
8. Conclusions
There are several antiviral defense mechanisms against tobamovirus infection, including the non-classic PTI, ETI, RNA-targeting pathway, phytohormones, ROS, and autophagy. Understanding how they function and interact will help to control plant viral diseases. Additionally, some recessive resistance genes also play a valuable role in this regard.
9. Future Directions
In the last few decades, a number of studies have demonstrated that transgenic approaches, especially by enhancing RNA silencing against viral RNA sequences, can provide effective plant protection against viruses [141]. Due to public concerns and strict regulatory barriers, this approach has been restricted for field use. However, it still holds great potential for generating virus-resistant plants.
Genome editing is another breakthrough for generating efficient virus resistance. Editing of tobamovirus susceptibility genes based on CRISPR/Cas9 systems can be used to generate crops with resistance against tobamoviruses. Numerous host susceptibility genes such as TOM1/3, TOM2, ARL8, and WPRb, have been identified, and their knockouts or mutants do not have any obvious effect on plant growth or morphology. More recently, Kunitz peptidase inhibitor-like protein (KPILP) was identified as a novel proviral factor during TMV or crTMV infection [142]. Editing of these genes may achieve tobamovirus resistance in different crops [143].
Engineering NLRs with new recognition targets is another approach that can be applied for protection against viruses [143]. The engineered NLRs can be generated for recognizing new pathogens by several technologies, including protein engineering, random or site-directed mutagenesis, and structure-based predictions. These strategies may be used to develop new crop resistance against tobamoviruses.
The exogenous application of dsRNA to induce RNA silencing has been perceived as an alternative to transgenesis and can be used for plant protection against viral disease. It has been shown to provide some protection against TMV [144] and can be modified to fight other tobamoviruses. However, the efficiency of this approach is affected by several factors, including the concentration/dose and length/size of dsRNAs, application method, delivery technique, plant organ-specific activities, and stability under open-field conditions [145]. Technical advances in these fields will overcome these restrictions for agricultural application. In addition, cross-protection is another efficient strategy without genetic modification, which is able to fight against severe virus strains. Reverse genetics can be adopted to generate attenuated mutants that have potential in cross-protection against tobamoviruses [141].
Author Contributions
Writing—original draft preparation, X.Z. and Y.L. (Yule Liu); writing—review and editing, X.Z., Y.L. (Yule Liu) and Y.L. (Yiqing Li). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grant numbers: 32130086, 32200118, and U23A20196). And The APC was funded by 32130086.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; Garcia, M.L.; Curtis Hendrickson, R.; et al. Recent changes to virus taxonomy ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2022, 167, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Goelet, P.; Lomonossoff, G.P.; Butler, P.J.; Akam, M.E.; Gait, M.J.; Karn, J. Nucleotide sequence of tobacco mosaic virus RNA. Proc. Natl. Acad. Sci. USA 1982, 79, 5818–5822. [Google Scholar] [CrossRef] [PubMed]
- Canto, T.; MacFarlane, S.A.; Palukaitis, P. ORF6 of Tobacco mosaic virus is a determinant of viral pathogenicity in Nicotiana benthamiana. J. Gen. Virol. 2004, 85, 3123–3133. [Google Scholar] [CrossRef] [PubMed]
- Sulzinski, M.A.; Gabard, K.A.; Palukaitis, P.; Zaitlin, M. Replication of tobacco mosaic virus. VIII. Characterization of a third subgenomic TMV RNA. Virology 1985, 145, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef]
- Fontes, E.P.B.; Teixeira, R.M.; Lozano-Duran, R. Plant virus-interactions: Unraveling novel defense mechanisms under immune-suppressing pressure. Curr. Opin. Biotechnol. 2021, 70, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.P.; Sede, A.R.; Elvira-González, L.; Yan, Y.; Rodriguez, M.E.; Mutterer, J.; Boutant, E.; Shan, L.B.; Heinlein, M. dsRNA-induced immunity targets plasmodesmata and is suppressed by viral movement proteins. Plant Cell 2023, 35, 3845–3869. [Google Scholar] [CrossRef] [PubMed]
- Guenoune-Gelbart, D.; Elbaum, M.; Sagi, G.; Levy, A.; Epel, B.L. Tobacco mosaic virus (TMV) replicase and movement protein function synergistically in facilitating TMV spread by lateral diffusion in the plasmodesmal desmotubule of Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2008, 21, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gong, Q.; Wu, Y.; Huang, F.; Ismayil, A.; Zhang, D.; Li, H.; Gu, H.; Ludman, M.; Fatyol, K.; et al. A calmodulin-binding transcription factor links calcium signaling to antiviral RNAi defense in plants. Cell Host Microbe 2021, 29, 1393–1406.e1397. [Google Scholar] [CrossRef] [PubMed]
- Dangl, J.L. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 1175. [Google Scholar] [CrossRef]
- Holmes, F.O. Inheritance of ability to localize tobacco mosaic virus. Phytopathology 1934, 24, 984–1002. [Google Scholar]
- Holmes, F.O. Inheritance of resistance to tobacco-mosaic disease in tobacco. Phytopathology 1938, 28, 553–561. [Google Scholar]
- Tobias, I.; Rast, A.T.B.; Maat, D.Z. Tobamoviruses of pepper, eggplant and tobacco: Comparative host reactions and serological relationships. Neth. J. Plant Pathol. 1982, 88, 257–268. [Google Scholar] [CrossRef]
- Pelletier, A.; Moffett, P. N and N′-mediated recognition confers resistance to tomato brown rugose fruit virus. MicroPubl. Biol. 2022, 2022. [Google Scholar] [CrossRef]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the tobacco mosaic virus resistance gene N: Similarity to toll and the interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Zambryski, P.; Staskawicz, B.; DineshKumar, S.P. Signaling in plant-microbe interactions. Science 1997, 276, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Dinesh-Kumar, S.P.; Tham, W.H.; Baker, B.J. Structure function analysis of the tobacco mosaic virus resistance gene N. Proc. Natl. Acad. Sci. USA 2000, 97, 14789–14794. [Google Scholar] [CrossRef] [PubMed]
- Doke, N.; Ohashi, Y. Involvement of an O2− generating system in the induction of necrotic lesions on tobacco leaves infected with tobacco mosaic virus. Physiol. Mol. Plant Pathol. 1988, 32, 163–175. [Google Scholar] [CrossRef]
- Durner, J.; Wendehenne, D.; Klessig, D.F. Defense gene induction in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose. Proc. Natl. Acad. Sci. USA 1998, 95, 10328–10333. [Google Scholar] [CrossRef] [PubMed]
- Padgett, H.S.; Watanabe, Y.; Beachy, R.N. Identification of the TMV replicase sequence that activates the N gene-mediated hypersensitive response. Mol. Plant-Microbe Interact. 1997, 10, 709–715. [Google Scholar] [CrossRef]
- Ueda, H.; Yamaguchi, Y.; Sano, H. Direct interaction between the tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor during the hypersensitive response in tobacco plants. Plant Mol. Biol. 2006, 61, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Abbink, T.E.M.; Tjernberg, P.A.; Bol, J.F.; Linthorst, H.J.M. Tobacco mosaic virus helicase domain induces necrosis in gene-carrying tobacco in the absence of virus replication. Mol. Plant-Microbe Interact. 1998, 11, 1242–1246. [Google Scholar] [CrossRef]
- Erickson, F.L.; Holzberg, S.; Calderon-Urrea, A.; Handley, V.; Axtell, M.; Corr, C.; Baker, B. The helicase domain of the TMV replicase proteins induces the N-mediated defence response in tobacco. Plant J. 1999, 18, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, M.S.; Ma, S.S.; Burch-Smith, T.M.; Czymmek, K.; Huijser, P.; Dinesh-Kumar, S.P. Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor-mediated plant innate immunity. PLoS Pathog. 2013, 9, e1003235. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.R.; Cook, G.; Feys, B.J.; Parker, J.E.; Baulcombe, D.C. An EDS1 orthologue is required for N-mediated resistance against tobacco mosaic virus. Plant J. 2002, 29, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Peart, J.R.; Mestre, P.; Lu, R.; Malcuit, I.; Baulcombe, D.C. NRG1, a CC-NB-LRR protein, together with N, a TIR-NB-LRR protein, mediates resistance against tobacco mosaic virus. Curr. Biol. 2005, 15, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Serino, G.; Deng, X.W.; Dinesh-Kumar, S.P. Role of SCF ubiquitin-ligase and the COP9 signalosome in the N gene-mediated resistance response to Tobacco mosaic virus. Plant Cell 2002, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Burch-Smith, T.; Schiff, M.; Feng, S.H.; Dinesh-Kumar, S.P. Molecular chaperone Hsp90 associates with resistance protein n and its signaling proteins SGT1 and Rar1 to modulate an innate immune response in plants. J. Biol. Chem. 2004, 279, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.M.; Zhao, J.P.; Chen, T.Y.; Liu, Q.; Zhang, H.L.; Wang, Y.; Hong, Y.G.; Xiao, F.M.; Zhang, L.; Shen, Q.H.; et al. Type I J-domain NbMIP1 proteins are required for both Tobacco mosaic virus infection and plant innate immunity. PLoS Pathog. 2013, 9, e1003659. [Google Scholar] [CrossRef] [PubMed]
- Caplan, J.L.; Mamillapalli, P.; Burch-Smith, T.M.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplastic protein NRIP1 mediates innate immune receptor recognition of a viral effector. Cell 2008, 132, 449–462. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Song, G.Y.; Lai, N.K.; Nagalakshmi, U.; Li, Y.Y.; Zheng, W.J.; Huang, P.J.; Branon, T.C.; Ting, A.Y.; Walley, J.W.; et al. TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity. Nat. Commun. 2019, 10, 3252. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Involvement of MEK1 MAPKK, NTF6 MAPK, WRKY/MYB transcription factors, COI1 and CTR1 in N-mediated resistance to tobacco mosaic virus. Plant J. 2004, 38, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Axtell, M.J.; Dahlbeck, D.; Ekwenna, O.; Zhang, S.; Staskawicz, B.; Baker, B. NPK1, an MEKK1-like mitogen-activated protein kinase kinase kinase, regulates innate immunity and development in plants. Dev. Cell 2002, 3, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.L.; Gao, Z.Y.; Zhang, H.; Yang, Y.Z.; Yang, X.X.; Zhao, X.F.; Guo, H.L.; Nagalakshmi, U.; Li, D.W.; Dinesh-Kumar, S.P.; et al. The MAPK-Alfin-like 7 module negatively regulates ROS scavenging genes to promote NLR-mediated immunity. Proc. Natl. Acad. Sci. USA 2023, 120, e2214750120. [Google Scholar] [CrossRef] [PubMed]
- Culver, J.N.; Dawson, W.O. Tobacco mosaic virus coat protein: An elicitor of the hypersensitive reaction but not required for the development of mosaic symptoms in Nicotiana sylvestris. Virology 1989, 173, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Hosokawa, D.; Meshi, T.; Okada, Y. Immunocytochemical localization of the 130K and 180K proteins (putative replicase components) of tobacco mosaic virus. Virology 1987, 160, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Yamanaka, K.; Watanabe, Y.; Takamatsu, N.; Meshi, T.; Okada, Y. Mutational analysis of the coat protein gene of tobacco mosaic virus in relation to hypersensitive response in tobacco plants with the N′ gene. Virology 1989, 173, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sekine, K.T.; Tomita, R.; Takeuchi, S.; Atsumi, G.; Saitoh, H.; Mizumoto, H.; Kiba, A.; Yamaoka, N.; Nishiguchi, M.; Hikichi, Y.; et al. Functional differentiation in the leucine-rich repeat domains of closely related plant virus-resistance proteins that recognize common avr proteins. Mol. Plant-Microbe Interact. 2012, 25, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Pelham, J. Resistance in tomato to Tobacco mosaic virus. Euphytica 1966, 15, 258–267. [Google Scholar] [CrossRef]
- Clayberg, C. Relative resistance of Tm-1 and Tm-2 to tobacco mosaic virus. Tomato Genet. Coop. Rep. 1960, 10, 13–14. [Google Scholar]
- Meshi, T.; Motoyoshi, F.; Adachi, A.; Watanabe, Y.; Takamatsu, N.; Okada, Y. Two concomitant base substitutions in the putative replicase genes of tobacco mosaic virus confer the ability to overcome the effects of a tomato resistance gene, Tm-1. Embo J. 1988, 7, 1575–1581. [Google Scholar] [CrossRef]
- Fraser, R.S.S.; Loughlin, S.A.R. Resistance to Tobacco mosaic virus in Tomato—Effects of the Tm-1 gene on virus multiplication. J. Gen. Virol. 1980, 48, 87–96. [Google Scholar] [CrossRef]
- Strasser, M.; Pfitzner, A.J. The double-resistance-breaking Tomato mosaic virus strain ToMV1-2 contains two independent single resistance-breaking domains. Arch. Virol. 2007, 152, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Masuda, K.; Naito, S.; Meshi, T.; Ishikawa, M. An inhibitor of viral RNA replication is encoded by a plant resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 13833–13838. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Ishikawa, M. The resistance protein Tm-1 inhibits formation of a Tomato mosaic virus replication protein-host membrane protein complex. J. Virol. 2013, 87, 7933–7939. [Google Scholar] [CrossRef]
- Ishibashi, K.; Kezuka, Y.; Kobayashi, C.; Kato, M.; Inoue, T.; Nonaka, T.; Ishikawa, M.; Matsumura, H.; Katoh, E. Structural basis for the recognition-evasion arms race between tomato mosaic virus and the resistance gene Tm-1. Proc. Natl. Acad. Sci. USA 2014, 111, E3486–E3495. [Google Scholar] [CrossRef]
- Ishibashi, K.; Miyashita, S.; Katoh, E.; Ishikawa, M. Host membrane proteins involved in the replication of tobamovirus RNA. Curr. Opin. Virol. 2012, 2, 699–704. [Google Scholar] [CrossRef]
- Lanfermeijer, F.C.; Warmink, J.; Hille, J. The products of the broken Tm-2 and the durable Tm-2(2) resistance genes from tomato differ in four amino acids. J. Exp. Bot. 2005, 56, 2925–2933. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.J. Resistance at the Tm-2 locus in the tomato to Tomato mosaic virus. Euphytica 1980, 29, 189–197. [Google Scholar] [CrossRef]
- Lanfermeijer, F.C.; Dijkhuis, J.; Sturre, M.J.G.; de Haan, P.; Hille, J. Cloning and characterization of the durable tomato mosaic virus resistance gene Tm-2 from Lycopersicon esculentum. Plant Mol. Biol. 2003, 52, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Pfitzner, A.J. Tm-2 resistance in tomato requires recognition of the carboxy terminus of the movement protein of tomato mosaic virus. Mol. Plant Microbe Interact. 1998, 11, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Tettey, C.K.; Mu, X.Q.; Ma, H.Y.; Chen, X.Y.; Geng, C.; Tian, Y.P.; Yan, Z.Y.; Li, X.D. The role of different innate and environmental factors in Tm-22-mediated resistance to tomato mottle mosaic virus. Phytopathol. Res. 2023, 5, 8. [Google Scholar] [CrossRef]
- Chen, T.; Liu, D.; Niu, X.; Wang, J.; Qian, L.; Han, L.; Liu, N.; Zhao, J.; Hong, Y.; Liu, Y. Antiviral resistance protein Tm-2(2) functions on the plasma membrane. Plant Physiol. 2017, 173, 2399–2410. [Google Scholar] [CrossRef] [PubMed]
- Hak, H.; Spiegelman, Z. The Tomato brown rugose fruit virus movement protein overcomes Tm-22 resistance in tomato while attenuating viral transport. Mol. Plant-Microbe Interact. 2021, 34, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.Y.; Ma, H.Y.; Wang, L.; Tettey, C.; Zhao, M.S.; Geng, C.; Tian, Y.P.; Li, X.D. Identification of genetic determinants of tomato brown rugose fruit virus that enable infection of plants harbouring the Tm-22 resistance gene. Mol. Plant Pathol. 2021, 22, 1347–1357. [Google Scholar] [CrossRef]
- Wang, J.; Chen, T.; Han, M.; Qian, L.; Li, J.; Wu, M.; Han, T.; Cao, J.; Nagalakshmi, U.; Rathjen, J.P.; et al. Plant NLR immune receptor Tm-22 activation requires NB-ARC domain-mediated self-association of CC domain. PLoS Pathog. 2020, 16, e1008475. [Google Scholar] [CrossRef]
- Qian, L.C.; Zhao, J.P.; Du, Y.M.; Zhao, X.J.; Han, M.; Liu, Y.L. Hsp90 interacts with Tm-2 and is essential for Tm-2-mediated resistance to Tobacco mosaic virus. Front. Plant Sci. 2018, 9, 411. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.P.; Liu, Q.; Zhang, H.L.; Jia, Q.; Hong, Y.G.; Liu, Y.L. The rubisco small subunit is involved in tobamovirus movement and Tm-22-mediated extreme resistance. Plant Physiol. 2013, 161, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, J.; Liu, S.; Zhang, D.P.; Liu, Y. Tm-22 confers different resistance responses against tobacco mosaic virus dependent on its expression level. Mol. Plant 2013, 6, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Boukema, I.W. Allelism of genes controlling resistance to TMV in Capsicum L. Euphytica 1980, 29, 433–439. [Google Scholar] [CrossRef]
- Tomita, R.; Sekine, K.T.; Mizumoto, H.; Sakamoto, M.; Murai, J.; Kiba, A.; Hikichi, Y.; Suzuki, K.; Kobayashi, K. Genetic basis for the hierarchical interaction between Tobamovirus spp. and L resistance gene alleles from different pepper species. Mol. Plant-Microbe Interact. 2011, 24, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Velez-Olmedo, J.B.; Fribourg, C.E.; Melo, F.L.; Nagata, T.; de Oliveira, A.S.; Resende, R.O. Tobamoviruses of two new species trigger resistance in pepper plants harbouring functional L alleles. J. Gen. Virol. 2021, 102, 001524. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, H.; Nakamura, I.; Shimomoto, Y.; Sawada, H.; Tomita, R.; Sekine, K.T.; Kiba, A.; Nishiguchi, M.; Kobayashi, K.; Hikichi, Y. Amino acids in Tobamovirus coat protein controlling pepper L1a gene-mediated resistance. Mol. Plant Pathol. 2012, 13, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Kravchik, M.; Shnaider, Y.; Abebie, B.; Shtarkman, M.; Kumari, R.; Kumar, S.; Leibman, D.; Spiegelman, Z.; Gal-On, A. Knockout of SlTOM1 and SlTOM3 results in differential resistance to tobamovirus in tomato. Mol. Plant Pathol. 2022, 23, 1278–1289. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, Y.; Komoda, K.; Yamanaka, T.; Tamai, A.; Meshi, T.; Funada, R.; Tsuchiya, T.; Naito, S.; Ishikawa, M. Subcellular localization of host and viral proteins associated with tobamovirus RNA replication. EMBO J. 2003, 22, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Numaga, T.; Ohshima, K.; Yano, M.A.; Ohsawa, R.; Goto, D.B.; Naito, S.; Ishikawa, M. Arabidopsis TOBAMOVIRUS MULTIPLICATION (TOM) 2 locus encodes a transmembrane protein that interacts with TOM1. EMBO J. 2003, 22, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Ohta, T.; Takahashi, M.; Meshi, T.; Schmidt, R.; Dean, C.; Naito, S.; Ishikawa, M. TOM1, an Arabidopsis gene required for efficient multiplication of a tobamovirus, encodes a putative transmembrane protein. Proc. Natl. Acad. Sci. USA 2000, 97, 10107–10112. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Ravelo, G.B.; Naito, S.; Ishikawa, M. Involvement of THH1, an Arabidopsis thaliana homologue of the TOM1 gene, in tobamovirus multiplication. J. Gen. Virol. 2006, 87, 2397–2401. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Obata, F.; Kumagai, T.; Ohno, T. Isolation of Mutants of Arabidopsis-Thaliana in Which Accumulation of Tobacco Mosaic-Virus Coat Protein Is Reduced to Low-Levels. Mol. Gen. Genet. 1991, 230, 33–38. [Google Scholar] [CrossRef]
- Yamanaka, T.; Imai, T.; Satoh, R.; Kawashima, A.; Takahashi, M.; Tomita, K.; Kubota, K.; Meshi, T.; Naito, S.; Ishikawa, M. Complete inhibition of tobamovirus multiplication by simultaneous mutations in two homologous host genes. J. Virol. 2002, 76, 2491–2497. [Google Scholar] [CrossRef] [PubMed]
- Asano, M.; Satoh, R.; Mochizuki, A.; Tsuda, S.; Yamanaka, T.; Nishiguchi, M.; Hirai, K.; Meshi, T.; Naito, S.; Ishikawa, M. Tobamovirus-resistant tobacco generated by RNA interference directed against host genes. Febs Lett. 2005, 579, 4479–4484. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Jiang, J.-h.; Zhou, X.-p. A TOM1 homologue is required for multiplication of Tobacco mosaic virus in Nicotiana benthamiana. J. Zhejiang Univ. Sci. B 2007, 8, 256–259. [Google Scholar] [CrossRef]
- Ali, M.E.; Ishii, Y.; Taniguchi, J.; Waliullah, S.; Kobayashi, K.; Yaeno, T.; Yamaoka, N.; Nishiguchi, M. Conferring virus resistance in tomato by independent RNA silencing of three tomato homologs of Arabidopsis TOM1. Arch. Virol. 2018, 163, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Jogam, P.; Sandhya, D.; Alok, A.; Peddaboina, V.; Singh, S.P.; Abbagani, S.; Zhang, B.H.; Allini, V.R. Editing of gene in tobacco using CRISPR/Cas9 confers resistance to Tobacco mosaic virus. Mol. Biol. Rep. 2023, 50, 5165–5176. [Google Scholar] [CrossRef]
- Ishikawa, M.; Yoshida, T.; Matsuyama, M.; Kouzai, Y.; Kano, A.; Ishibashi, K. Tomato brown rugose fruit virus resistance generated by quadruple knockout of homologs of TOBAMOVIRUS MULTIPLICATION1 in tomato. Plant Physiol. 2022, 189, 679–686. [Google Scholar] [CrossRef]
- Nishikiori, M.; Mori, M.; Dohi, K.; Okamura, H.; Katoh, E.; Naito, S.; Meshi, T.; Ishikawa, M. A Host Small GTP-binding Protein ARL8 Plays Crucial Roles in Tobamovirus RNA Replication. PLoS Pathog. 2011, 7, e1002409. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Liu, J.; Wang, S.; Gong, Z.; Yang, S.; Xu, F.; Hu, Z.; Zhang, M.; Yang, J. The coiled-coil protein gene WPRb confers recessive resistance to Cucumber green mottle mosaic virus. Plant Physiol. 2023, 191, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Carrington, J.C.; Kasschau, K.D.; Johansen, L.K. Activation and suppression of RNA silencing by plant viruses. Virology 2001, 281, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Vance, V.; Vaucheret, H. RNA silencing in plants—Defense and counterdefense. Science 2001, 292, 2277–2280. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef]
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell 2000, 101, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Malpica-López, N.; Rajeswaran, R.; Beknazariants, D.; Seguin, J.; Golyaev, V.; Farinelli, L.; Pooggin, M.M. Revisiting the Roles of Tobamovirus Replicase Complex Proteins in Viral Replication and Silencing Suppression. Mol. Plant-Microbe Interact. 2018, 31, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Niu, Y.; Zhang, K.; Liu, Y.; Zhou, X. Virus-derived transgenes expressing hairpin RNA give immunity to Tobacco mosaic virus and Cucumber mosaic virus. Virol. J. 2011, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zheng, Q.; Yi, X.; An, H.; Zhao, Y.; Ma, S.; Zhou, G. Establishing RNA virus resistance in plants by harnessing CRISPR immune system. Plant Biotechnol. J. 2018, 16, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, X.; Zhou, G.; Zhang, T. Engineering plant virus resistance: From RNA silencing to genome editing strategies. Plant Biotechnol. J. 2020, 18, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.H.; Sun, Z.N.; Song, Y.Z.; An, H.L.; Zhu, C.X.; Wen, F.J. Bacterially expressed double-stranded RNAs against hot-spot sequences of tobacco mosaic virus or potato virus Y genome have different ability to protect tobacco from viral infection. Appl. Biochem. Biotechnol. 2010, 162, 1901–1914. [Google Scholar] [CrossRef] [PubMed]
- Akbarimotlagh, M.; Azizi, A.; Shams-Bakhsh, M.; Jafari, M.; Ghasemzadeh, A.; Palukaitis, P. Critical points for the design and application of RNA silencing constructs for plant virus resistance. Adv. Virus Res. 2023, 115, 159–203. [Google Scholar] [CrossRef]
- Taliansky, M.; Samarskaya, V.; Zavriev, S.K.; Fesenko, I.; Kalinina, N.O.; Love, A.J. RNA-based technologies for engineering plant virus resistance. Plants 2021, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.S.; Liu, J.; Cheng, N.H.; Folimonov, A.; Hou, Y.M.; Bao, Y.; Katagi, C.; Carter, S.A.; Nelson, R.S. The Tobacco mosaic virus 126-kDa protein associated with virus replication and movement suppresses RNA silencing. Mol. Plant Microbe Interact. 2004, 17, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Watanabe, Y. A TMV-Cg mutant with a truncated coat protein induces cell death resembling the hypersensitive response in Arabidopsis. Mol. Cells 2004, 17, 334–339. [Google Scholar] [CrossRef]
- Csorba, T.; Bovi, A.; Dalmay, T.; Burgyán, J. The p122 subunit of replicase is a potent silencing suppressor and compromises both small interfering RNA- and MicroRNA-mediated pathways. J. Virol. 2007, 81, 11768–11780. [Google Scholar] [CrossRef] [PubMed]
- Várallyay, É.; Havelda, Z. Unrelated viral suppressors of RNA silencing mediate the control of ARGONAUTE1 level. Mol. Plant Pathol. 2013, 14, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Vogler, H.; Kwon, M.O.; Dang, V.; Sambade, A.; Fasler, M.; Ashby, J.; Heinlein, M. Tobacco mosaic virus movement protein enhances the spread of RNA silencing. PLoS Pathog. 2008, 4, e1000038. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Zhou, X.; Li, F. Plant-virus arms race beyond RNA interference. Trends Plant Sci. 2024, 29, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Cougot, N.; Babajko, S.; Seraphin, B. Cytoplasmic foci are sites of mRNA decay in human cells. J. Cell Biol. 2004, 165, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Conti, G.; Zavallo, D.; Venturuzzi, A.L.; Rodriguez, M.C.; Crespi, M.; Asurmendi, S. TMV induces RNA decay pathways to modulate gene silencing and disease symptoms. Plant J. 2017, 89, 73–84. [Google Scholar] [CrossRef]
- Peng, J.; Yang, J.; Yan, F.; Lu, Y.; Jiang, S.; Lin, L.; Zheng, H.; Chen, H.; Chen, J. Silencing of NbXrn4 facilitates the systemic infection of Tobacco mosaic virus in Nicotiana benthamiana. Virus Res. 2011, 158, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Chivasa, S.; Murphy, A.M.; Naylor, M.; Carr, J.P. Salicylic acid interferes with tobacco mosaic virus replication via a novel salicylhydroxamic acid-sensitive mechanism. Plant Cell 1997, 9, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Carr, J.P. Salicylic acid has cell-specific effects on replication and cell-to-cell movement. Plant Physiol. 2002, 128, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Bi, Y.M.; Darby, R.M.; Firek, S.; Draper, J. Compromising early salicylic acid accumulation delays the hypersensitive response and increases viral dispersal during lesion establishment in TMV-infected tobacco. Plant J. 1997, 12, 1113–1126. [Google Scholar] [CrossRef]
- Xie, Z.X.; Fan, B.F.; Chen, C.H.; Chen, Z.X. An important role of an inducible RNA-dependent RNA polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.Q.; Fan, B.F.; MacFarlane, S.A.; Chen, Z.X. Analysis of the involvement of an inducible RNA-dependent RNA polymerase in antiviral defense. Mol. Plant-Microbe Interact. 2003, 16, 206–216. [Google Scholar] [CrossRef]
- Lee, W.S.; Fu, S.F.; Li, Z.; Murphy, A.M.; Dobson, E.A.; Garland, L.; Chaluvadi, S.R.; Lewsey, M.G.; Nelson, R.S.; Carr, J.P. Salicylic acid treatment and expression of an RNA-dependent RNA polymerase 1 transgene inhibit lethal symptoms and meristem invasion during tobacco mosaic virus infection in Nicotiana benthamiana. BMC Plant Biol. 2016, 16, 1–14. [Google Scholar] [CrossRef]
- Li, S.W.; Yu, F.F.; Wang, M.A.; Guo, X.Q.; Li, H. Molecular characterization of a Nicotiana tabacum NtRDR6 gene. Plant Mol. Biol. Rep. 2012, 30, 1375–1384. [Google Scholar] [CrossRef]
- Campos, L.; Granell, P.; Tarraga, S.; Lopez-Gresa, P.; Conejero, V.; Belles, J.M.; Rodrigo, I.; Lison, P. Salicylic acid and gentisic acid induce RNA silencing-related genes and plant resistance to RNA pathogens. Plant Physiol. Biochem. 2014, 77, 35–43. [Google Scholar] [CrossRef]
- Alazem, M.; Kim, K.H.; Lin, N.S. Effects of abscisic acid and salicylic acid on gene expression in the antiviral RNA silencing pathway in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2538. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, M.; Gao, Z.; Zhu, C.; Guo, X. Isolation of a novel RNA-dependent RNA polymerase 6 from Nicotiana glutinosa, NgRDR6, and analysis of its response to biotic and abiotic stresses. Mol. Biol. Rep. 2011, 38, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, Q.Q.; Wu, B.; Ai, T.B.; Guo, X.Q. NgRDR1, an RNA-dependent RNA polymerase isolated from Nicotiana glutinosa, was involved in biotic and abiotic stresses. Plant Physiol. Biochem. 2009, 47, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Hunter, L.J.; Westwood, J.H.; Heath, G.; Macaulay, K.; Smith, A.G.; Macfarlane, S.A.; Palukaitis, P.; Carr, J.P. Regulation of RNA-dependent RNA polymerase 1 and isochorismate synthase gene expression in Arabidopsis. PLoS ONE 2013, 8, e66530. [Google Scholar] [CrossRef] [PubMed]
- Delaney, T.P.; Uknes, S.; Vernooij, B.; Friedrich, L.; Weymann, K.; Negrotto, D.; Gaffney, T.; Gut-Rella, M.; Kessmann, H.; Ward, E.; et al. A central role of salicylic acid in plant disease resistance. Science 1994, 266, 1247–1250. [Google Scholar] [CrossRef]
- Oka, K.; Kobayashi, M.; Mitsuhara, I.; Seo, S. Jasmonic acid negatively regulates resistance to Tobacco mosaic virus in tobacco. Plant Cell Physiol. 2013, 54, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Xi, D.H.; Yuan, S.; Xu, F.; Zhang, D.W.; Lin, H.H. Salicylic acid and jasmonic acid are essential for systemic resistance against tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant Microbe Interact. 2014, 27, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Canto, T.; Palukaitis, P. Novel N gene-associated, temperature-independent resistance to the movement of tobacco mosaic virus vectors neutralized by a cucumber mosaic virus RNA1 transgene. J. Virol. 2002, 76, 12908–12916. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zheng, Q.; Cheng, L.; Liu, P.; Xu, G.; Zhang, H.; Cao, P.; Zhou, H. Identification and characterization of TMV-induced volatile signals in Nicotiana benthamiana: Evidence for JA/ET defense pathway priming in congeneric neighbors via airborne (E)-2-octenal. Funct. Integr. Genom. 2023, 23, 272. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Wang, Y.; He, L.; Huang, F.; Zhang, D.; Wang, Y.; Wei, X.; Han, M.; Deng, H.; Luo, L.; et al. Molecular basis of methyl-salicylate-mediated plant airborne defence. Nature 2023, 622, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, H.; Hong, N.; Kang, B.; Peng, B.; Liu, L.; Gu, Q. Argonaute 1 and 5 proteins play crucial roles in the defence against cucumber green mottle mosaic virus in watermelon. Mol. Plant Pathol. 2023, 24, 961–972. [Google Scholar] [CrossRef]
- Zhu, T.; Zhou, X.; Zhang, J.L.; Zhang, W.H.; Zhang, L.P.; You, C.X.; Jameson, P.E.; Ma, P.T.; Guo, S.L. Ethylene-induced is involved in resistance against tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant Pathol. 2022, 23, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: Structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, M.S.; Goregaoker, S.P.; Golem, S.; Shiferaw, H.; Culver, J.N. Interaction of the tobacco mosaic virus replicase protein with the Aux/IAA protein PAP1/IAA26 is associated with disease development. J. Virol. 2005, 79, 2549–2558. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, M.S.; Shiferaw, H.; Culver, J.N. The replicase protein disrupts the localization and function of interacting Aux/IAA proteins. Mol. Plant-Microbe Interact. 2006, 19, 864–873. [Google Scholar] [CrossRef]
- Padmanabhan, M.S.; Kramer, S.R.; Wang, X.; Culver, J.N. Tobacco mosaic virus replicase-auxin/indole acetic acid protein interactions: Reprogramming the auxin response pathway to enhance virus infection. J. Virol. 2008, 82, 2477–2485. [Google Scholar] [CrossRef]
- Collum, T.D.; Padmanabhan, M.S.; Hsieh, Y.C.; Culver, J.N. Tobacco mosaic virus-directed reprogramming of auxin/indole acetic acid protein transcriptional responses enhances virus phloem loading. Proc. Natl. Acad. Sci. USA 2016, 113, E2740–E2749. [Google Scholar] [CrossRef]
- Deng, X.G.; Zhu, T.; Peng, X.J.; Xi, D.H.; Guo, H.Q.; Yin, Y.H.; Zhang, D.W.; Lin, H.H. Role of brassinosteroid signaling in modulating Tobacco mosaic virus resistance in Nicotiana benthamiana. Sci. Rep. 2016, 6, 20579. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.G.; Zhu, T.; Zou, L.J.; Han, X.Y.; Zhou, X.; Xi, D.H.; Zhang, D.W.; Lin, H.H. Orchestration of hydrogen peroxide and nitric oxide in brassinosteroid-mediated systemic virus resistance in Nicotiana benthamiana. Plant J. 2016, 85, 478–493. [Google Scholar] [CrossRef]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef]
- Conti, G.; Rodriguez, M.C.; Manacorda, C.A.; Asurmendi, S. Transgenic expression of Tobacco mosaic virus capsid and movement proteins modulate plant basal defense and biotic stress responses in Nicotiana tabacum. Mol. Plant-Microbe Interact. 2012, 25, 1370–1384. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Conti, G.; Zavallo, D.; Manacorda, C.A.; Asurmendi, S. TMV-Cg Coat Protein stabilizes DELLA proteins and in turn negatively modulates salicylic acid-mediated defense pathway during Arabidopsis thaliana viral infection. BMC Plant Biol. 2014, 14, 210. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Cao, M.Y.; Zhu, P.X.; Zhang, Q.P.; Lam, H.M. Non-specific LIPID TRANSFER PROTEIN 1 enhances immunity against tobacco mosaic virus in Nicotiana benthamiana. J. Exp. Bot. 2023, 74, 5236–5254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bassham, D.C. Autophagy: Pathways for self-eating in plant cells. Annu. Rev. Plant Biol. 2012, 63, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Ismayil, A.; Liu, Y. Autophagy in plant-virus interactions. Annu. Rev. Virol. 2020, 7, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.Y.; Chen, T.Y.; Qian, L.C.; Liu, N.; Wang, Y.J.; Han, S.J.; et al. Autophagy functions as an antiviral mechanism against geminiviruses in plants. Elife 2017, 6, e23897. [Google Scholar] [CrossRef]
- Jiao, Y.; An, M.; Li, X.; Yu, M.; Zhao, X.; Xia, Z.; Wu, Y. Transcriptomic and functional analyses reveal an antiviral role of autophagy during pepper mild mottle virus infection. BMC Plant Biol. 2020, 20, 495. [Google Scholar] [CrossRef] [PubMed]
- Macharia, M.W.; Tan, W.Y.Z.; Das, P.P.; Naqvi, N.I.; Wong, S.M. Proximity-dependent biotinylation screening identifies NbHYPK as a novel interacting partner of ATG8 in plants. BMC Plant Biol. 2019, 19, 326. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Wang, Y.; Zheng, X.; Jia, Q.; Zhao, J.; Bai, F.; Hong, Y.; Liu, Y. Cytoplastic glyceraldehyde-3-phosphate dehydrogenases interact with ATG3 to negatively regulate autophagy and immunity in Nicotiana benthamiana. Plant Cell 2015, 27, 1316–1331. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Czymmek, K.; Talloczy, Z.; Levine, B.; Dinesh-Kumar, S.P. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005, 121, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.Y.; Wang, S.S.; Han, S.J.; Xie, K.; Wang, Y.; Li, J.L.; Liu, Y.L. Plant Bax Inhibitor-1 interacts with ATG6 to regulate autophagy and programmed cell death. Autophagy 2017, 13, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.M.; Hong, Q.; Li, Y.; Li, Q.; Wang, M. Autophagy contributes to regulate the ROS levels and PCD progress in TMV-infected tomatoes. Plant Sci. 2018, 269, 12–19. [Google Scholar] [CrossRef]
- Anikina, I.; Kamarova, A.; Issayeva, K.; Issakhanova, S.; Mustafayeva, N.; Insebayeva, M.; Mukhamedzhanova, A.; Khan, S.M.; Ahmad, Z.; Lho, L.H.; et al. Plant protection from virus: A review of different approaches. Front. Plant Sci. 2023, 14, 1163270. [Google Scholar] [CrossRef]
- Ershova, N.; Kamarova, K.; Sheshukova, E.; Antimonova, A.; Komarova, T. A novel cellular factor of Nicotiana benthamiana susceptibility to tobamovirus infection. Front. Plant Sci. 2023, 14, 1224958. [Google Scholar] [CrossRef] [PubMed]
- Marchal, C.; Pai, H.; Kamoun, S.; Kourelis, J. Emerging principles in the design of bioengineered made-to-order plant immune receptors. Curr. Opin. Plant Biol. 2022, 70, 102311. [Google Scholar] [CrossRef] [PubMed]
- Konakalla, N.C.; Kaldis, A.; Berbati, M.; Masarapu, H.; Voloudakis, A.E. Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco. Planta 2016, 244, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Das, P.R.; Sherif, S.M. Application of exogenous dsRNAs-induced RNAi in agriculture: Challenges and triumphs. Front. Plant Sci. 2020, 11, 946. [Google Scholar] [CrossRef] [PubMed]
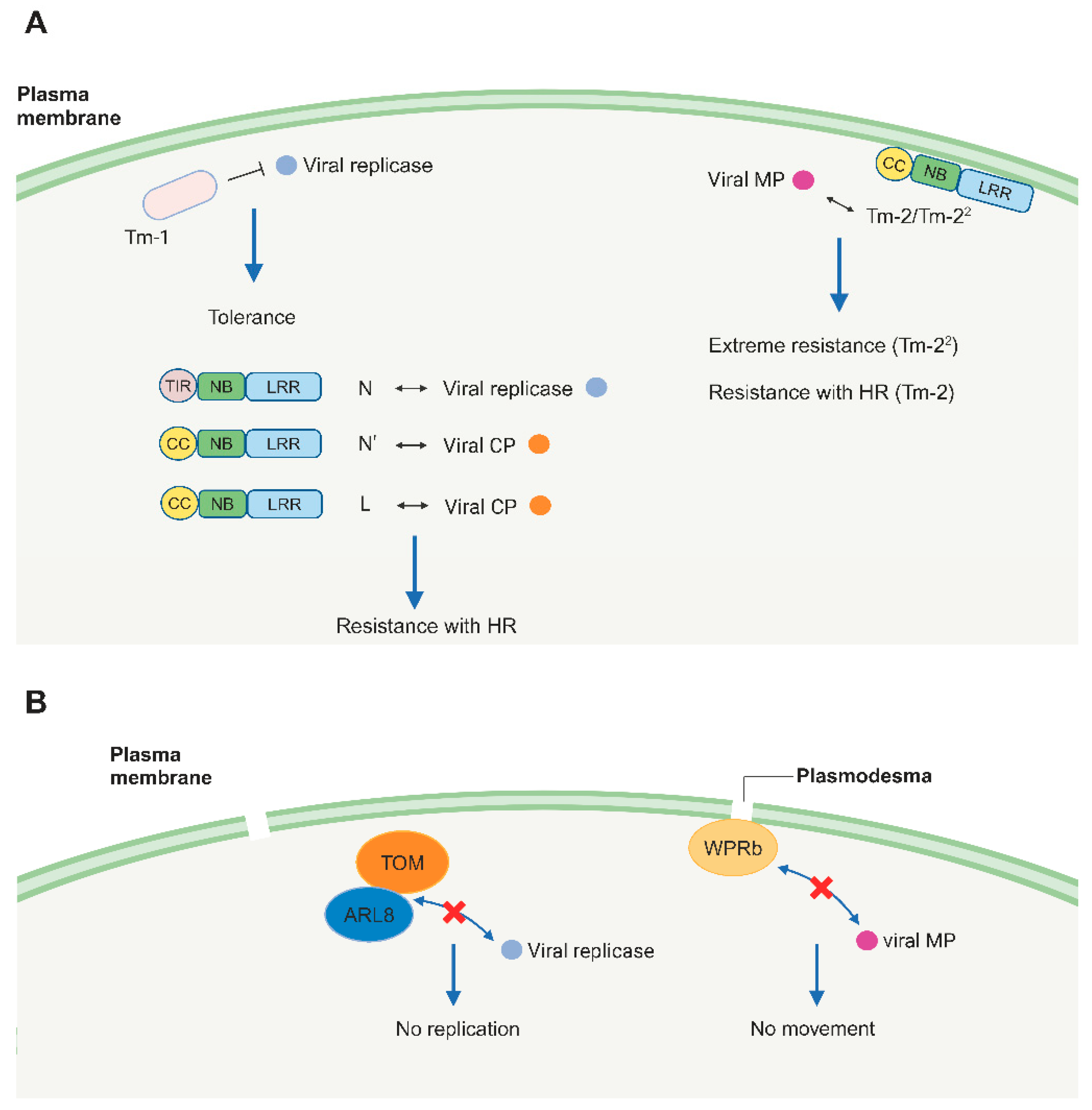
Figure 1.
Dominant (A) and recessive resistance (B) against tobamoviruses. (A). In nucleotide-binding leucine-rich repeat receptor (NLR)-mediated resistance, NLR proteins interact with avirulence (Avr) proteins, leading to the hypersensitive response (HR) or extreme resistance. In Tm-1-mediated tolerance, Tm-1 interacts with viral replicase to inhibit viral replication. (B). Recessive resistance is caused by the absence of a host factor that is essential for tobamovirus infection. Both TOM and ARL8 interact with viral replicase to promote tobamoviruses replication. WPRb interacts with the viral movement protein (MP) to facilitate viral movement. TIR: Toll-interleukin-1 receptor domain, CC: coiled-coil domain, NB: nucleotide-binding domain, LRR: leucine-rich repeat domain, CP: coat protein, TOM: Tobamovirus multiplication, ARL8: Arabidopsis ADP-ribosylation factor-like 8, WPRb: WEB1/PMI2-related protein. Figure adapted from images created with BioRender.com.
Figure 1.
Dominant (A) and recessive resistance (B) against tobamoviruses. (A). In nucleotide-binding leucine-rich repeat receptor (NLR)-mediated resistance, NLR proteins interact with avirulence (Avr) proteins, leading to the hypersensitive response (HR) or extreme resistance. In Tm-1-mediated tolerance, Tm-1 interacts with viral replicase to inhibit viral replication. (B). Recessive resistance is caused by the absence of a host factor that is essential for tobamovirus infection. Both TOM and ARL8 interact with viral replicase to promote tobamoviruses replication. WPRb interacts with the viral movement protein (MP) to facilitate viral movement. TIR: Toll-interleukin-1 receptor domain, CC: coiled-coil domain, NB: nucleotide-binding domain, LRR: leucine-rich repeat domain, CP: coat protein, TOM: Tobamovirus multiplication, ARL8: Arabidopsis ADP-ribosylation factor-like 8, WPRb: WEB1/PMI2-related protein. Figure adapted from images created with BioRender.com.

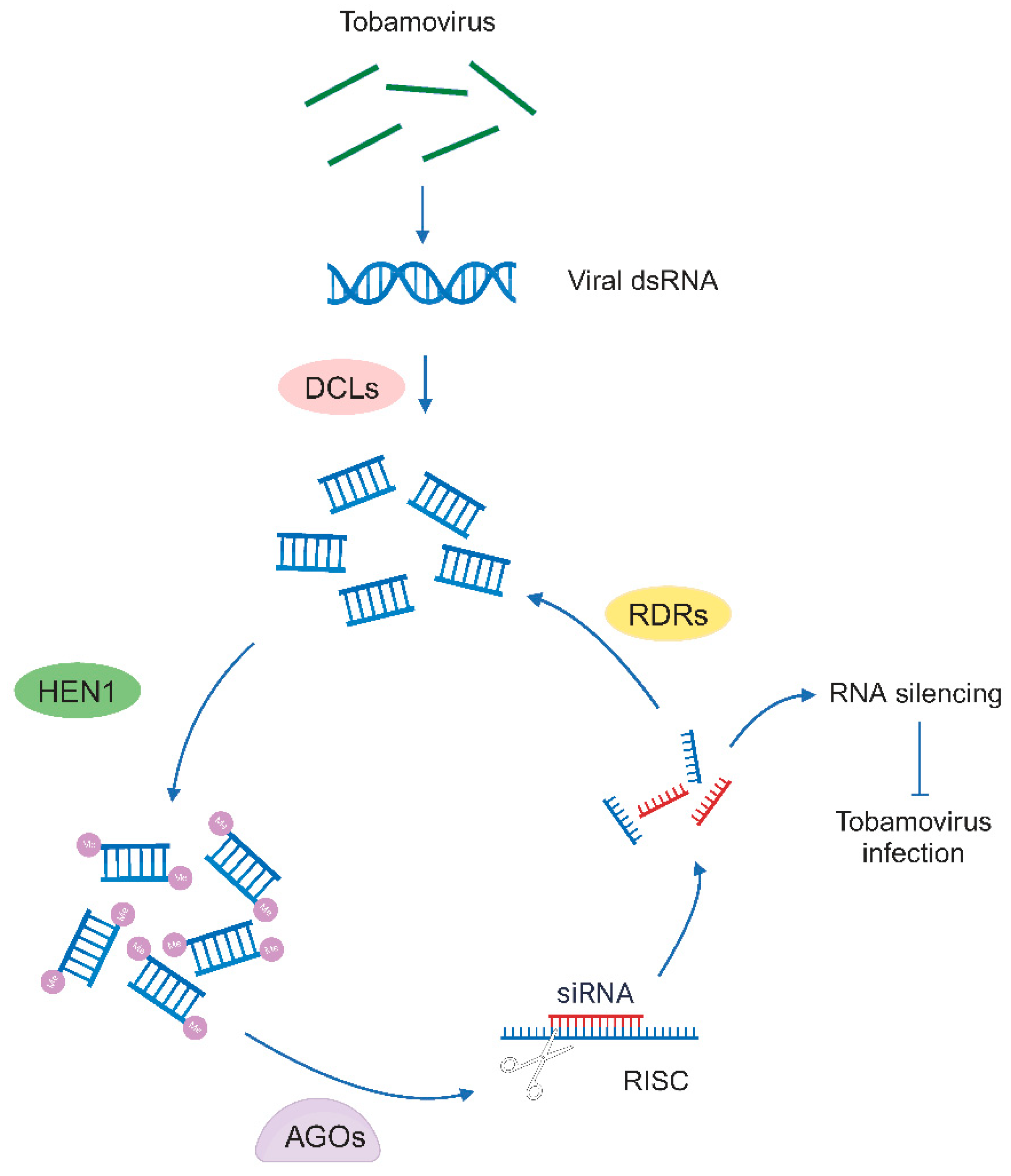
Figure 2.
The RNA silencing pathway against tobamoviruses. Upon tobamovirus infection, viral double-stranded RNA (dsRNA) is generated by viral replicase as an intermediate in genome replication, and it is cleaved by Dicer-like (DCL) enzymes to generate viral small RNAs (sRNAs) duplexes of 21–24 nucleotides. The 3′ terminal nucleotides of the sRNAs duplexes are methylated by HUA ENHANCER1 (HEN1) to protect them from degradation. The resulting sRNAs are incorporated into the Argonaute (AGO)-containing RNA-induced silencing complex (RISC) for silencing, thus inhibit the tobamovirus infection. This process is further enhanced by RNA-dependent RNA polymerases (RDRs), which bind to the product of RISC and produce the secondary sRNAs. Figure adapted from images created with BioRender.com.
Figure 2.
The RNA silencing pathway against tobamoviruses. Upon tobamovirus infection, viral double-stranded RNA (dsRNA) is generated by viral replicase as an intermediate in genome replication, and it is cleaved by Dicer-like (DCL) enzymes to generate viral small RNAs (sRNAs) duplexes of 21–24 nucleotides. The 3′ terminal nucleotides of the sRNAs duplexes are methylated by HUA ENHANCER1 (HEN1) to protect them from degradation. The resulting sRNAs are incorporated into the Argonaute (AGO)-containing RNA-induced silencing complex (RISC) for silencing, thus inhibit the tobamovirus infection. This process is further enhanced by RNA-dependent RNA polymerases (RDRs), which bind to the product of RISC and produce the secondary sRNAs. Figure adapted from images created with BioRender.com.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zheng, X.; Li, Y.; Liu, Y. Plant Immunity against Tobamoviruses. Viruses 2024, 16, 530. https://doi.org/10.3390/v16040530
AMA Style
Zheng X, Li Y, Liu Y. Plant Immunity against Tobamoviruses. Viruses. 2024; 16(4):530. https://doi.org/10.3390/v16040530
Chicago/Turabian StyleZheng, Xiyin, Yiqing Li, and Yule Liu. 2024. "Plant Immunity against Tobamoviruses" Viruses 16, no. 4: 530. https://doi.org/10.3390/v16040530
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.