
Serologic, Virologic and Pathologic Features of Cats with Naturally Occurring Feline Infectious Peritonitis Enrolled in Antiviral Clinical Trials

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Trial Protocol
2.2. Handling and Storage of Feline Samples
2.3. Immunoblotting of Feline Plasma and Ascites Fluid against FIPV Proteins
2.4. Enzyme-Linked Immunosorbent Assays (ELISA)
2.5. ELISA and Western Blots for Anti-FIPV Immunoglobulin Isotypes
2.6. Quantitation of FIPV Replication
2.7. FIPV Serotype I and II Standard PCR
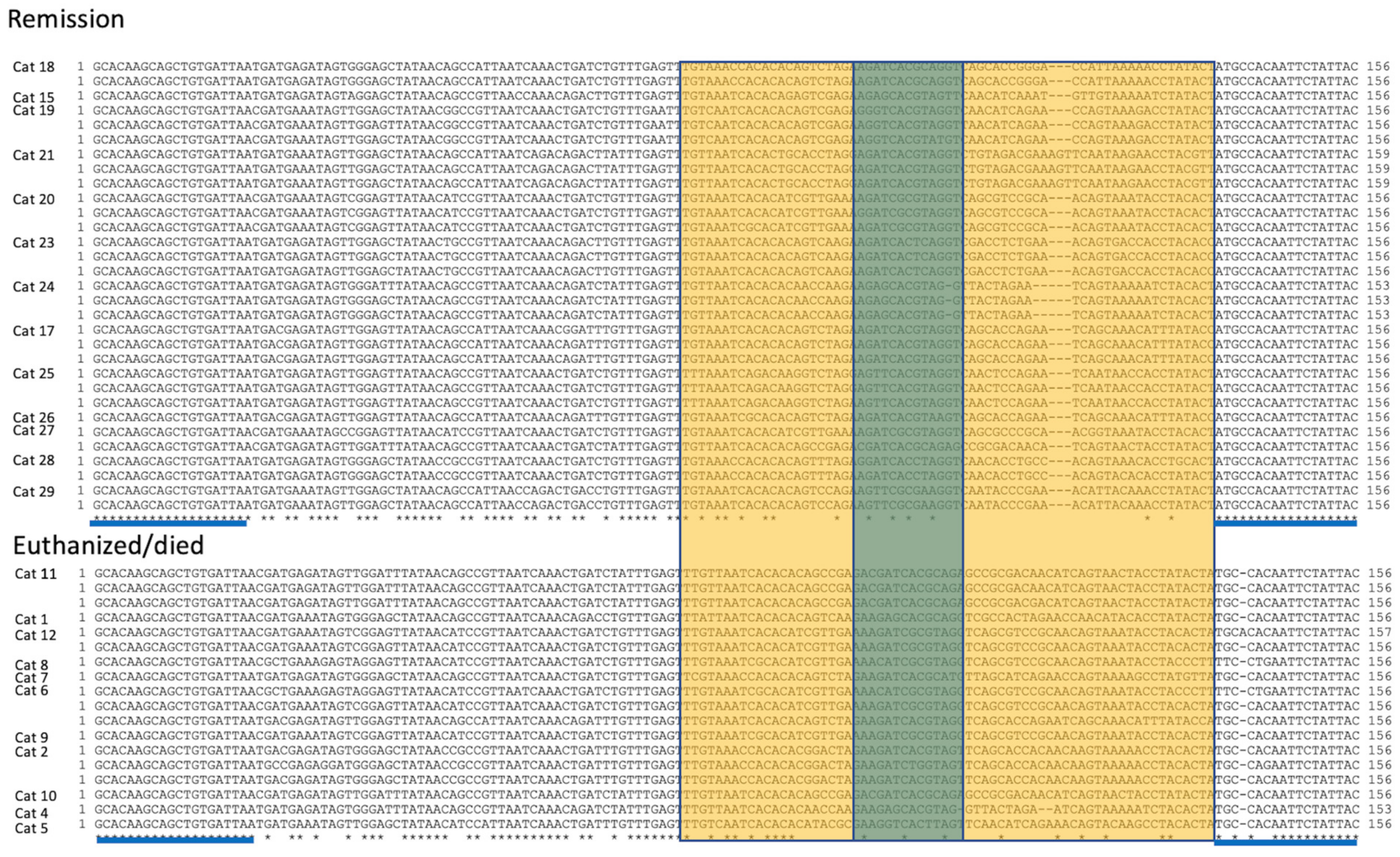
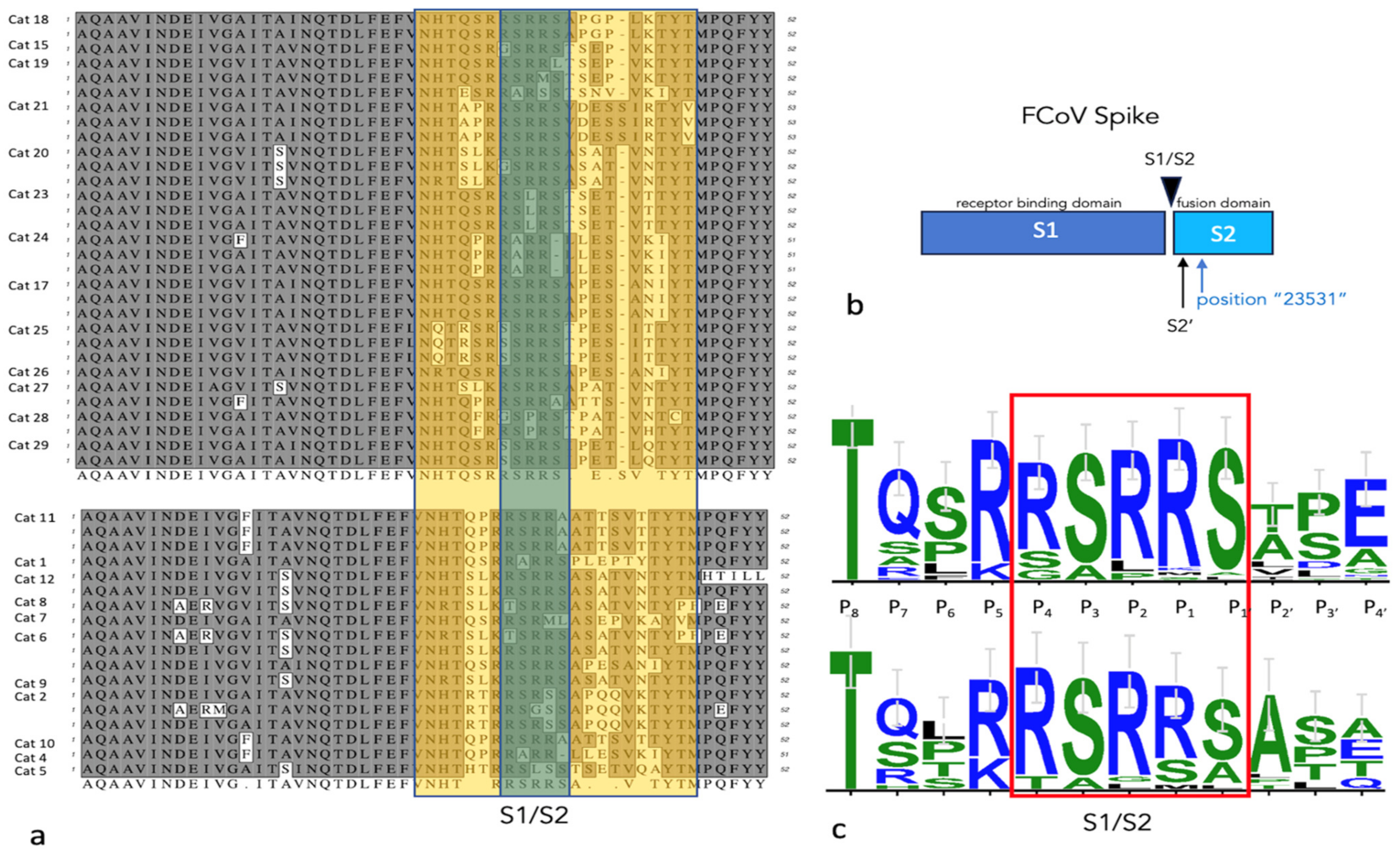
2.8. S1/S2 PCR Amplification, Cloning and Sequencing
2.9. Necropsy and Tissue Analyses
2.10. Statistical Tests
3. Results
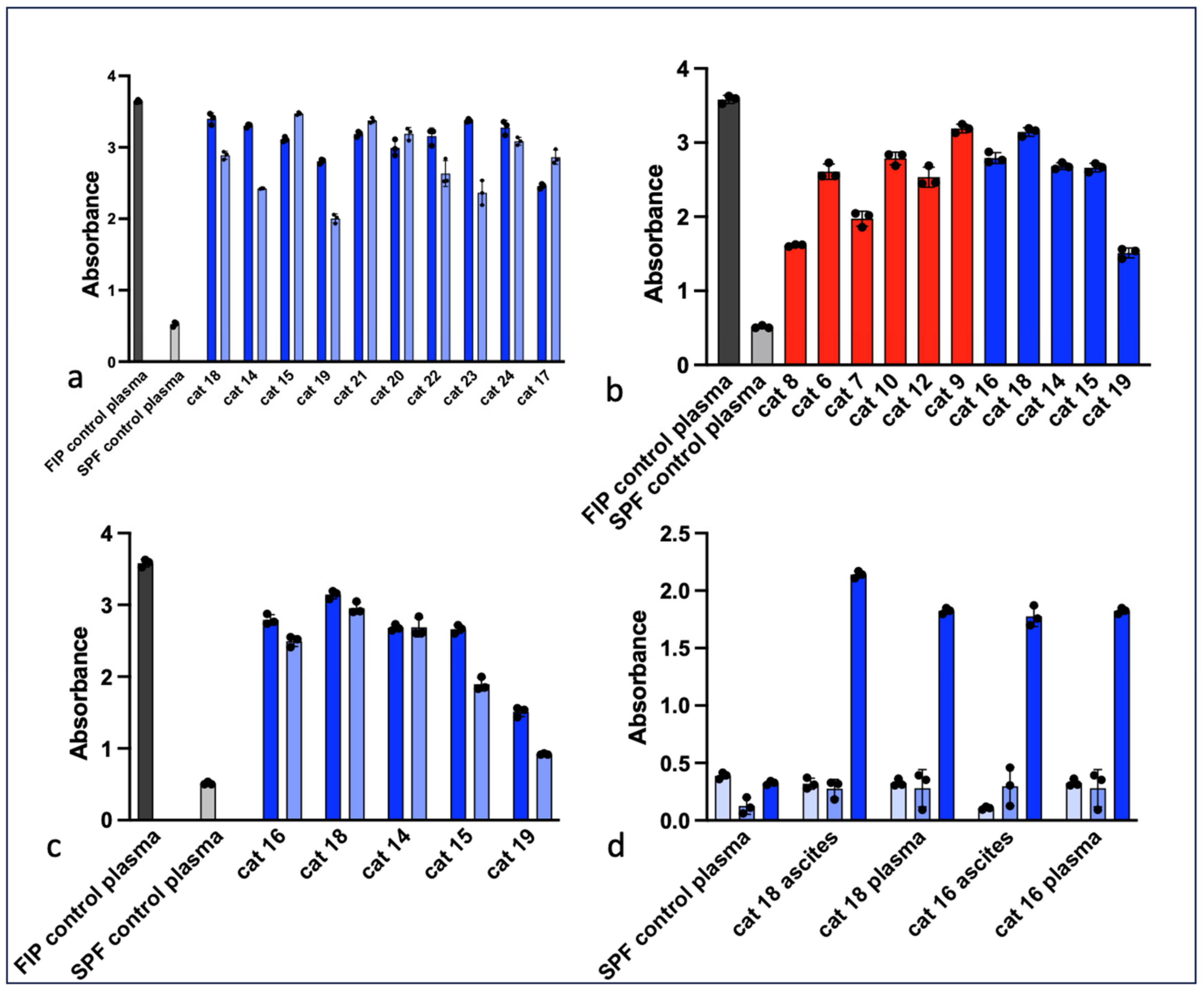
3.1. Serology
3.2. Western Blot (Immunoblot)
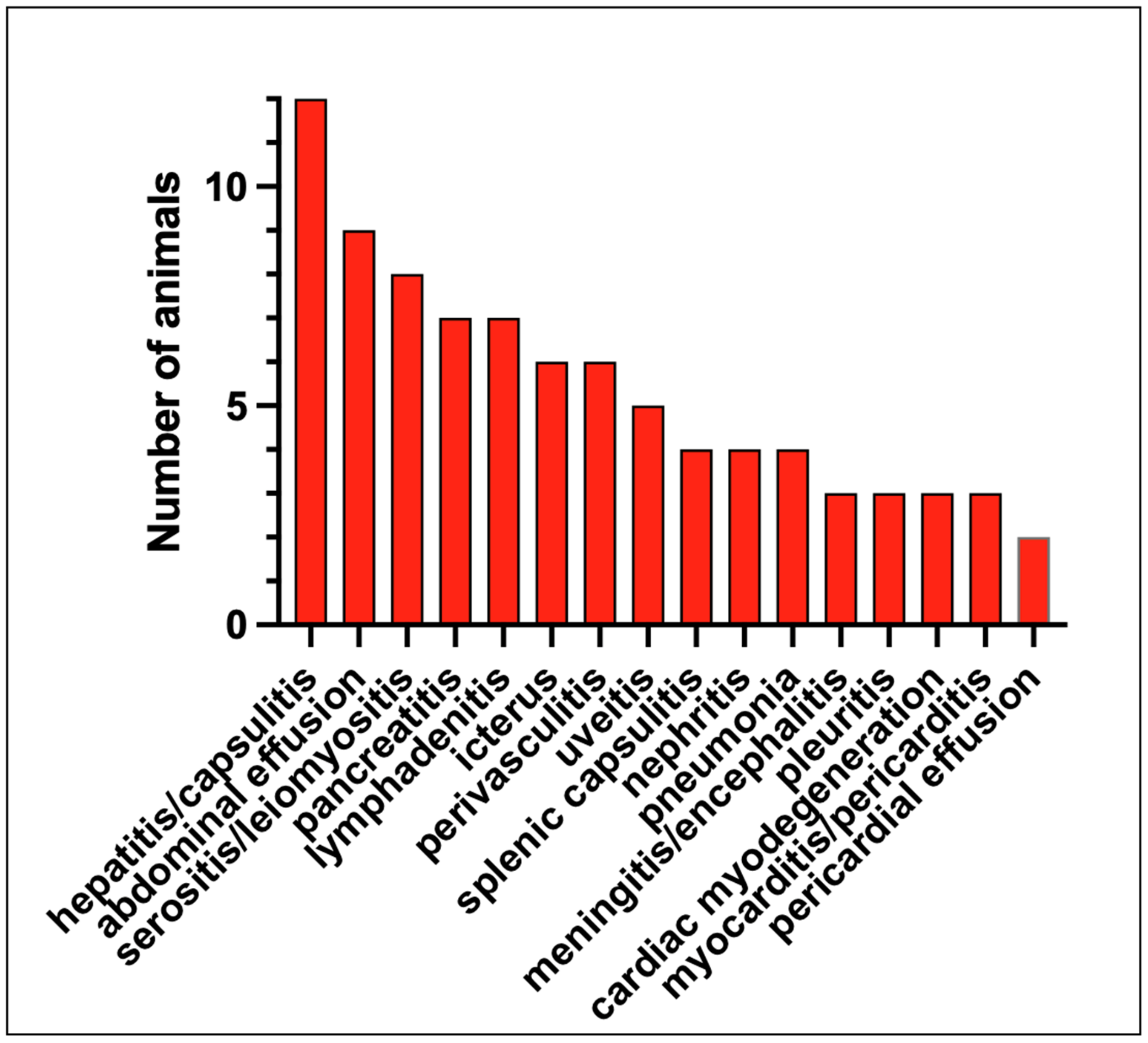
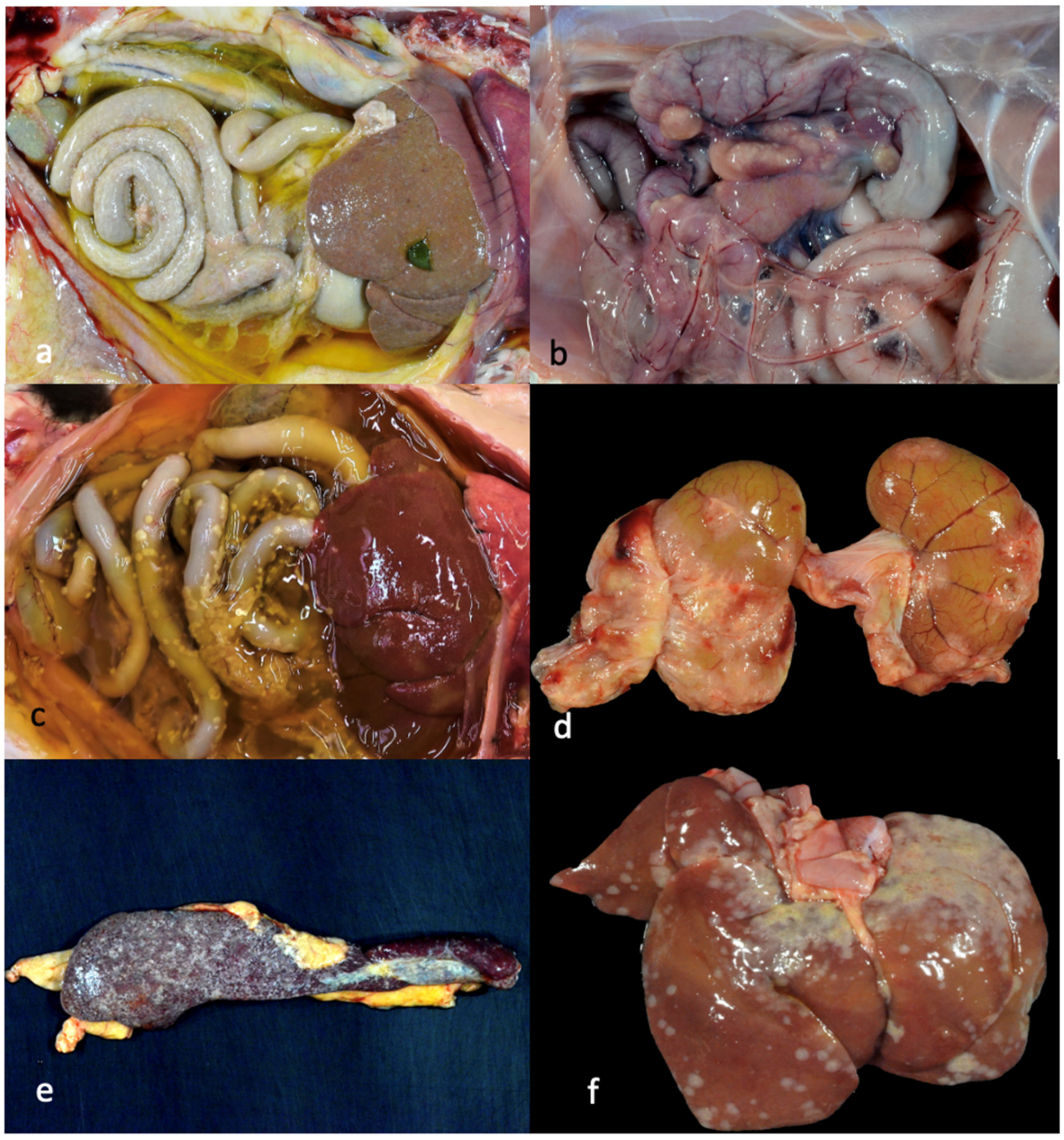
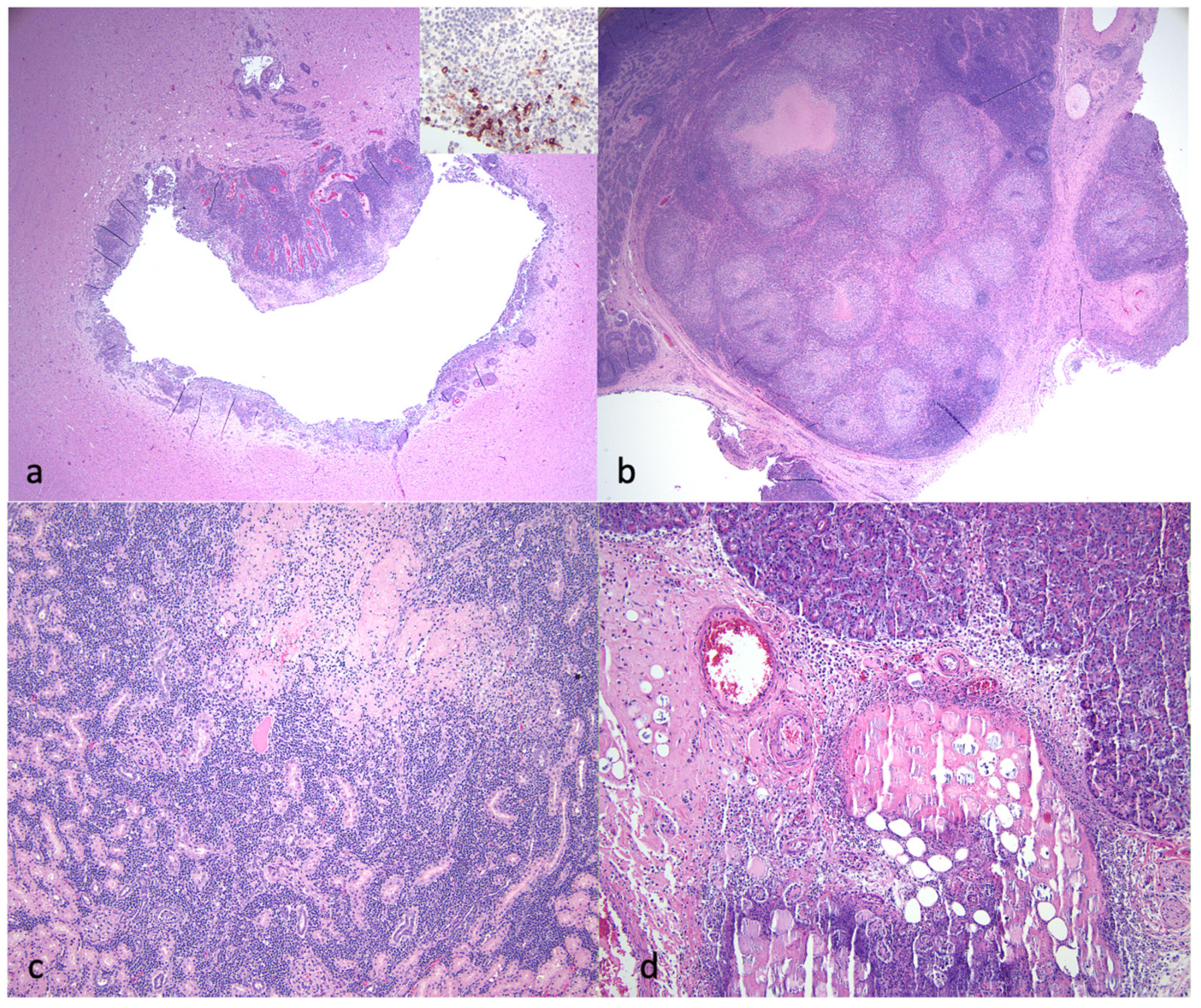
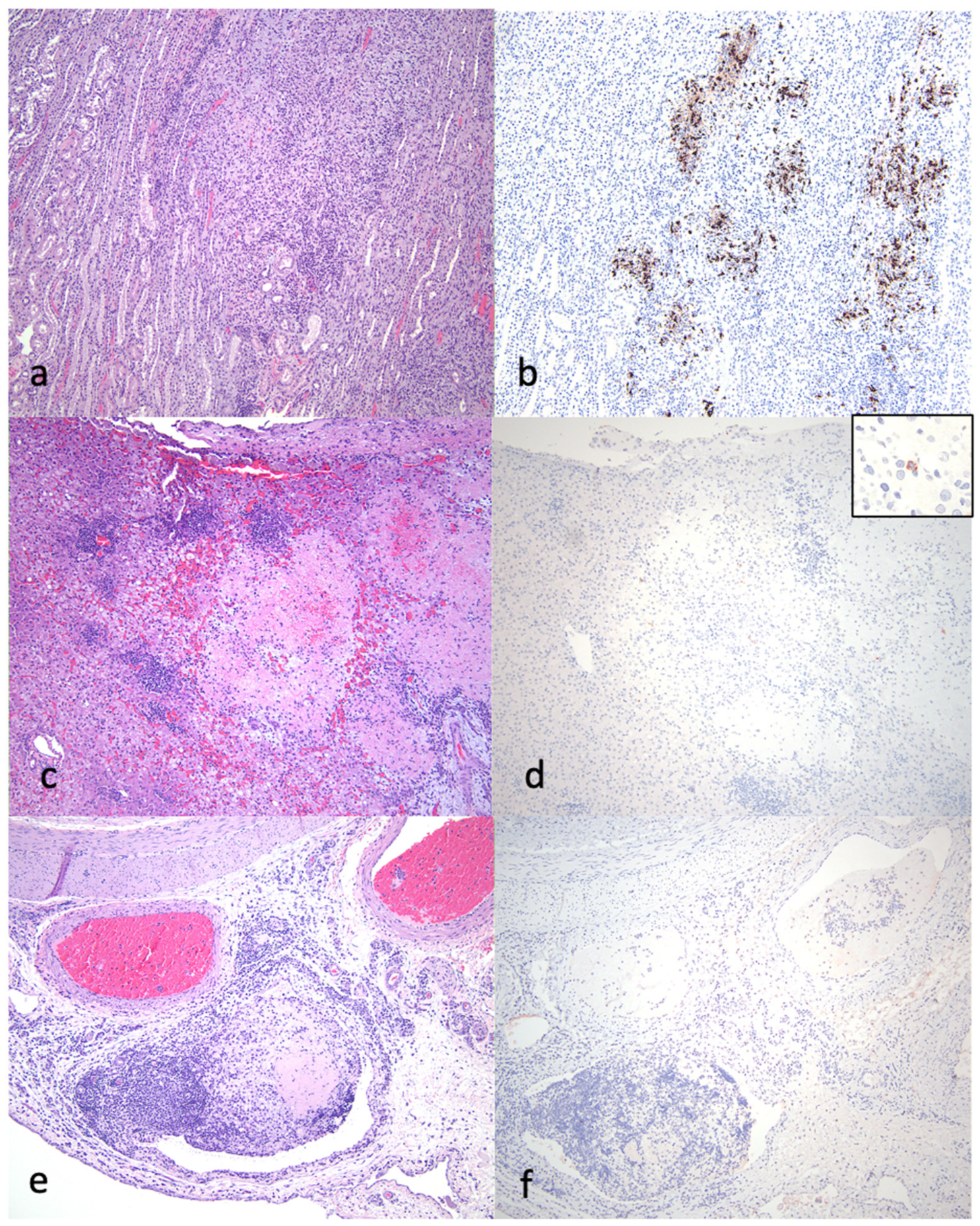
3.3. Pathology
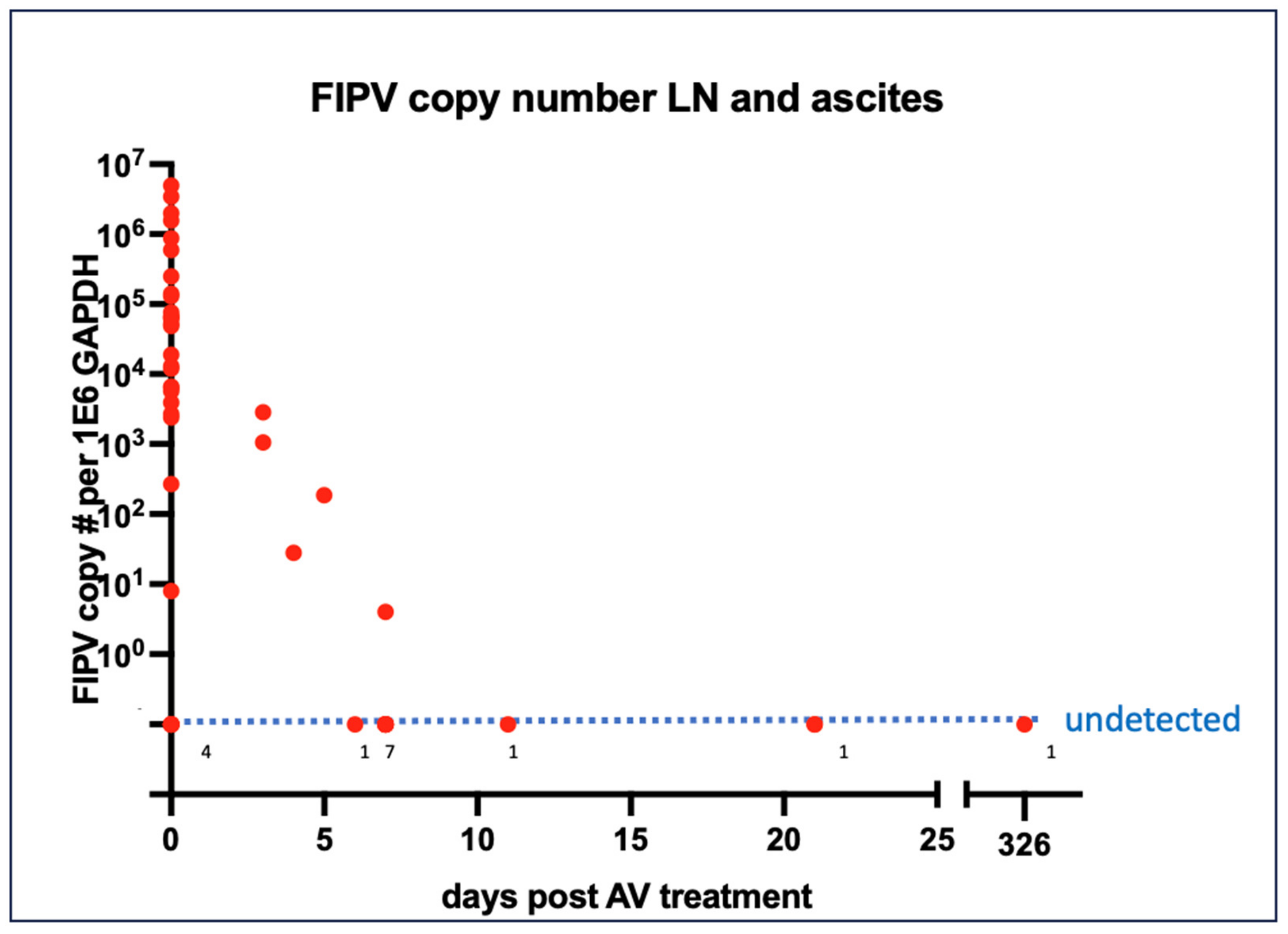
3.4. Virology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J. Spike protein fusion peptide and feline coronavirus virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg. 2009, 11, 225–258. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Allen, C.E.; Lyons, L.A. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Vogel, L.; Van der Lubben, M.; te Lintelo, E.G.; Bekker, C.P.; Geerts, T.; Schuijff, L.S.; Grinwis, G.C.; Egberink, H.F.; Rottier, P.J. Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Vet. Res. 2010, 41, 71. [Google Scholar] [CrossRef] [PubMed]
- Vennema, H.; Poland, A.; Foley, J.; Pedersen, N.C. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 1998, 243, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Liu, H.; Dodd, K.A.; Pesavento, P.A. Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses 2009, 1, 166–184. [Google Scholar] [CrossRef]
- Montali, R.J.; Strandberg, J.D. Extraperitoneal lesions in feline infectious peritonitis. Vet. Pathol. 1972, 9, 109–121. [Google Scholar] [CrossRef]
- Herrewegh, A.A.; Mahler, M.; Hedrich, H.J.; Haagmans, B.L.; Egberink, H.F.; Horzinek, M.C.; Rottier, P.J.; de Groot, R.J. Persistence and evolution of feline coronavirus in a closed cat-breeding colony. Virology 1997, 234, 349–363. [Google Scholar] [CrossRef]
- Terada, Y.; Matsui, N.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Soma, T.; Mochizuki, M.; Maeda, K. Emergence of pathogenic coronaviruses in cats by homologous recombination between feline and canine coronaviruses. PLoS ONE 2014, 9, e106534. [Google Scholar] [CrossRef]
- Herrewegh, A.A.; Smeenk, I.; Horzinek, M.C.; Rottier, P.J.; De Groot, R.J. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998, 72, 4508–4514. [Google Scholar] [CrossRef]
- Jaimes, J.A.; Millet, J.K.; Stout, A.E.; Andre, N.M.; Whittaker, G.R. A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses 2020, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Benetka, V.; Kubber-Heiss, A.; Kolodziejek, J.; Nowotny, N.; Hofmann-Parisot, M.; Mostl, K. Prevalence of feline coronavirus types I and II in cats with histopathologically verified feline infectious peritonitis. Vet. Microbiol. 2004, 99, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Sulejmani, N.; Jafri, S.M.; Gordon, S.C. Pharmacodynamics and pharmacokinetics of elbasvir and grazoprevir in the treatment of hepatitis C. Expert Opin. Drug Metab. Toxicol. 2016, 12, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Kwo, P.; Gane, E.J.; Peng, C.Y.; Pearlman, B.; Vierling, J.M.; Serfaty, L.; Buti, M.; Shafran, S.; Stryszak, P.; Lin, L.; et al. Effectiveness of Elbasvir and Grazoprevir Combination, with or without Ribavirin, for Treatment-Experienced Patients with Chronic Hepatitis C Infection. Gastroenterology 2017, 152, 164–175.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Vedantham, P.; Lu, K.; Agudelo, J.; Carrion, R., Jr.; Nunneley, J.W.; Barnard, D.; Pohlmann, S.; McKerrow, J.H.; Renslo, A.R.; et al. Protease inhibitors targeting coronavirus and filovirus entry. Antivir. Res. 2015, 116, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Dyall, J.; Coleman, C.M.; Hart, B.J.; Venkataraman, T.; Holbrook, M.R.; Kindrachuk, J.; Johnson, R.F.; Olinger, G.G., Jr.; Jahrling, P.B.; Laidlaw, M.; et al. Repurposing of clinically developed drugs for treatment of Middle East respiratory syndrome coronavirus infection. Antimicrob. Agents Chemother. 2014, 58, 4885–4893. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Hu, Y.T.; Wong, S.H.; Lin, L.T. Therapeutic Strategies against Ebola Virus Infection. Viruses 2022, 14, 579. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Perron, M.; Bannasch, M.; Montgomery, E.; Murakami, E.; Liepnieks, M.; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg. 2019, 21, 271–281. [Google Scholar] [CrossRef]
- Murphy, B.G.; Perron, M.; Murakami, E.; Bauer, K.; Park, Y.; Eckstrand, C.; Liepnieks, M.; Pedersen, N.C. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Vet. Microbiol. 2018, 219, 226–233. [Google Scholar] [CrossRef]
- Cosaro, E.; Pires, J.; Castillo, D.; Murphy, B.G.; Reagan, K.L. Efficacy of Oral Remdesivir Compared to GS-441524 for Treatment of Cats with Naturally Occurring Effusive Feline Infectious Peritonitis: A Blinded, Non-Inferiority Study. Viruses 2023, 15, 1680. [Google Scholar] [CrossRef]
- Cook, S.; Wittenburg, L.; Yan, V.C.; Theil, J.H.; Castillo, D.; Reagan, K.L.; Williams, S.; Pham, C.D.; Li, C.; Muller, F.L.; et al. An Optimized Bioassay for Screening Combined Anticoronaviral Compounds for Efficacy against Feline Infectious Peritonitis Virus with Pharmacokinetic Analyses of GS-441524, Remdesivir, and Molnupiravir in Cats. Viruses 2022, 14, 2429. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.E.; Vogel, H.; Castillo, D.; Olsen, M.; Pedersen, N.; Murphy, B.G. Investigation of monotherapy and combined anticoronaviral therapies against feline coronavirus serotype II in vitro. J. Feline Med. Surg. 2022, 24, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Sase, O. Molnupiravir treatment of 18 cats with feline infectious peritonitis: A case series. J. Vet. Intern. Med. 2023, 37, 1876–1880. [Google Scholar] [CrossRef]
- Taechangam, N.; Kol, A.; Arzi, B.; Borjesson, D.L. Multipotent Stromal Cells and Viral Interaction: Current Implications for Therapy. Stem Cell Rev. Rep. 2022, 18, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.G.; Walters-Laird, C.J.; Kol, A.; Santos Rocha, C.; Hirao, L.A.; Mende, A.; Balan, B.; Arredondo, J.; Elizaldi, S.R.; Iyer, S.S.; et al. Gut germinal center regeneration and enhanced antiviral immunity by mesenchymal stem/stromal cells in SIV infection. JCI Insight 2021, 6, e149033. [Google Scholar] [CrossRef] [PubMed]
- Coggins, S.J.; Norris, J.M.; Malik, R.; Govendir, M.; Hall, E.J.; Kimble, B.; Thompson, M.F. Outcomes of treatment of cats with feline infectious peritonitis using parenterally administered remdesivir, with or without transition to orally administered GS-441524. J. Vet. Intern. Med. 2023, 37, 1772–1783. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Novicoff, W.; Nadeau, J.; Evans, S. Unlicensed GS-441524-Like Antiviral Therapy Can Be Effective for at-Home Treatment of Feline Infectious Peritonitis. Animals 2021, 11, 2257. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Kim, Y.; Liu, H.; Galasiti Kankanamalage, A.C.; Eckstrand, C.; Groutas, W.C.; Bannasch, M.; Meadows, J.M.; Chang, K.O. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg. 2018, 20, 378–392. [Google Scholar] [CrossRef]
- Krentz, D.; Zenger, K.; Alberer, M.; Felten, S.; Bergmann, M.; Dorsch, R.; Matiasek, K.; Kolberg, L.; Hofmann-Lehmann, R.; Meli, M.L.; et al. Curing Cats with Feline Infectious Peritonitis with an Oral Multi-Component Drug Containing GS-441524. Viruses 2021, 13, 2228. [Google Scholar] [CrossRef]
- Meli, M.L.; Spiri, A.M.; Zwicklbauer, K.; Krentz, D.; Felten, S.; Bergmann, M.; Dorsch, R.; Matiasek, K.; Alberer, M.; Kolberg, L.; et al. Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524. Viruses 2022, 14, 1069. [Google Scholar] [CrossRef]
- Zwicklbauer, K.; Krentz, D.; Bergmann, M.; Felten, S.; Dorsch, R.; Fischer, A.; Hofmann-Lehmann, R.; Meli, M.L.; Spiri, A.M.; Alberer, M.; et al. Long-term follow-up of cats in complete remission after treatment of feline infectious peritonitis with oral GS-441524. J. Feline Med. Surg. 2023, 25, 1098612X231183250. [Google Scholar] [CrossRef] [PubMed]
- Krentz, D.; Zwicklbauer, K.; Felten, S.; Bergmann, M.; Dorsch, R.; Hofmann-Lehmann, R.; Meli, M.L.; Spiri, A.M.; von Both, U.; Alberer, M.; et al. Clinical Follow-Up and Postmortem Findings in a Cat that Was Cured of Feline Infectious Peritonitis with an Oral Antiviral Drug Containing GS-441524. Viruses 2022, 14, 2040. [Google Scholar] [CrossRef] [PubMed]
- Arzi, B.; Kol, A.; Murphy, B.; Walker, N.J.; Wood, J.A.; Clark, K.; Verstraete, F.J.; Borjesson, D.L. Feline foamy virus adversely affects feline mesenchymal stem cell culture and expansion: Implications for animal model development. Stem Cells Dev. 2015, 24, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Arzi, B.; Mills-Ko, E.; Verstraete, F.J.; Kol, A.; Walker, N.J.; Badgley, M.R.; Fazel, N.; Murphy, W.J.; Vapniarsky, N.; Borjesson, D.L. Therapeutic Efficacy of Fresh, Autologous Mesenchymal Stem Cells for Severe Refractory Gingivostomatitis in Cats. Stem Cells Transl. Med. 2016, 5, 75–86. [Google Scholar] [CrossRef]
- Cook, S.; Castillo, D.; Williams, S.; Haake, C.; Murphy, B. Serotype I and II Feline Coronavirus Replication and Gene Expression Patterns of Feline Cells—Building a Better Understanding of Serotype I FIPV Biology. Viruses 2022, 14, 1356. [Google Scholar] [CrossRef] [PubMed]
- Addie, D.D.; Schaap, I.A.T.; Nicolson, L.; Jarrett, O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003, 84 Pt 10, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.D.; Duhamel, G.E.; Whittaker, G.R. Mutation in spike protein cleavage site and pathogenesis of feline coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Eckstrand, C.; Liu, H.; Leutenegger, C.; Murphy, B. Levels of feline infectious peritonitis virus in blood, effusions, and various tissues and the role of lymphopenia in disease outcome following experimental infection. Vet. Microbiol. 2015, 175, 157–166. [Google Scholar] [CrossRef]
- Jaimes, J.A.; Whittaker, G.R. Feline coronavirus: Insights into viral pathogenesis based on the spike protein structure and function. Virology 2018, 517, 108–121. [Google Scholar] [CrossRef]
- Lemmermeyer, T.; Lamp, B.; Schneider, R.; Ziebuhr, J.; Tekes, G.; Thiel, H.J. Characterization of monoclonal antibodies against feline coronavirus accessory protein 7b. Vet. Microbiol. 2016, 184, 11–19. [Google Scholar] [CrossRef]
- Armstrong, J.; Niemann, H.; Smeekens, S.; Rottier, P.; Warren, G. Sequence and topology of a model intracellular membrane protein, E1 glycoprotein, from a coronavirus. Nature 1984, 308, 751–752. [Google Scholar] [CrossRef]
- Yamamoto, J.K.; Edison, L.K.; Rowe-Haas, D.K.; Takano, T.; Gilor, C.; Crews, C.D.; Tuanyok, A.; Arukha, A.P.; Shiomitsu, S.; Walden, H.D.S.; et al. Both Feline Coronavirus Serotypes 1 and 2 Infected Domestic Cats Develop Cross-Reactive Antibodies to SARS-CoV-2 Receptor Binding Domain: Its Implication to Pan-CoV Vaccine Development. Viruses 2023, 15, 914. [Google Scholar] [CrossRef]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- Gao, Y.Y.; Liang, X.Y.; Wang, Q.; Zhang, S.; Zhao, H.; Wang, K.; Hu, G.X.; Liu, W.J.; Gao, F.S. Mind the feline coronavirus: Comparison with SARS-CoV-2. Gene 2022, 825, 146443. [Google Scholar] [CrossRef]
- Ernandes, M.A.; Cantoni, A.M.; Armando, F.; Corradi, A.; Ressel, L.; Tamborini, A. Feline coronavirus-associated myocarditis in a domestic longhair cat. JFMS Open Rep. 2019, 5, 2055116919879256. [Google Scholar] [CrossRef]
- Shu, H.; Zhao, C.; Wang, D.W. Understanding COVID-19-related myocarditis: Pathophysiology, diagnosis and treatment strategies. Cardiol. Plus 2023, 8, 37539019. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abdominal Effusion | Icterus | Serositis or Leiomyositis | Vasculitis/Uerivasculitis | Hepatitis/Capsulitis | Nephritis | Lymphadenitis | Splenic Capsulitis | Pancreatitis | Meningitis/Encephalitis | Uveitis | Cardiac Myodegeneration | Myocarditis/Pericarditis | Pneumonia | Pericardial Effusion | Pleuritis | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 74-80-41 | √ | √ +++ | √ ++ | √ ++ | √ 0 | √ 0 | √ 0 | |||||||||
| 74-95-21 | √ | √ | √ | √ | √ +++ | √ | √ +++ | √ | ||||||||
| 75-63-32 | √ ++ | √ | √ ++ | √ | √ +++ | |||||||||||
| 75-14-64 1d | √ +++ | √ | √ +++ | √ +++ | √ +++ | √ 0 | √ | |||||||||
| 74-89-23 2d | √ | √ | √ ++ | √ | √ +++ | √ +++ | √ | |||||||||
| 74-90-61 3d | √ | √ | √ | √ | √ | √ | √ | |||||||||
| 74-84-08 3d | √ | √ + | √ 0 | √ 0 | √ + | √ 0 | √ | √ | ||||||||
| 74-83-16 4d | √ | √ | √ 0 | √ 0 | √ 0 | √ 0 | √ 0 | |||||||||
| 74-93-52 5d | √ | √ | √ | √ 0 | √ | √ | √ | √ | √ 0 | √ | ||||||
| 74-98-25 6d | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||
| 74-81-36 7d | √ | √ + | √ 0 | √ + | √ 0 | √ 0 | ||||||||||
| 74-82-32 11d | √ | √ | √ | √ 0 | ||||||||||||
| 74-85-67 132d | √ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murphy, B.G.; Castillo, D.; Neely, N.E.; Kol, A.; Brostoff, T.; Grant, C.K.; Reagan, K.L. Serologic, Virologic and Pathologic Features of Cats with Naturally Occurring Feline Infectious Peritonitis Enrolled in Antiviral Clinical Trials. Viruses 2024, 16, 462. https://doi.org/10.3390/v16030462
Murphy BG, Castillo D, Neely NE, Kol A, Brostoff T, Grant CK, Reagan KL. Serologic, Virologic and Pathologic Features of Cats with Naturally Occurring Feline Infectious Peritonitis Enrolled in Antiviral Clinical Trials. Viruses. 2024; 16(3):462. https://doi.org/10.3390/v16030462
Chicago/Turabian StyleMurphy, Brian G., Diego Castillo, N E Neely, Amir Kol, Terza Brostoff, Chris K. Grant, and Krystle L. Reagan. 2024. "Serologic, Virologic and Pathologic Features of Cats with Naturally Occurring Feline Infectious Peritonitis Enrolled in Antiviral Clinical Trials" Viruses 16, no. 3: 462. https://doi.org/10.3390/v16030462