
Rhabdoviral Endogenous Sequences Identified in the Leishmaniasis Vector Lutzomyia longipalpis Are Widespread in Sandflies from South America
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. Sandflies
2.2. Lutzomyia Longipalpis Cell Lines
2.3. RNA and DNA Extractions
2.4. cDNA Synthesis
2.5. Polymerase Chain Reactions (PCR)
2.6. Agarose Gel Electrophoresis
2.7. Bioinformatics Tools
2.8. Alignment
2.9. Evolutionary Analysis by Maximum Likelihood Method
2.10. Small RNA Analysis
3. Results
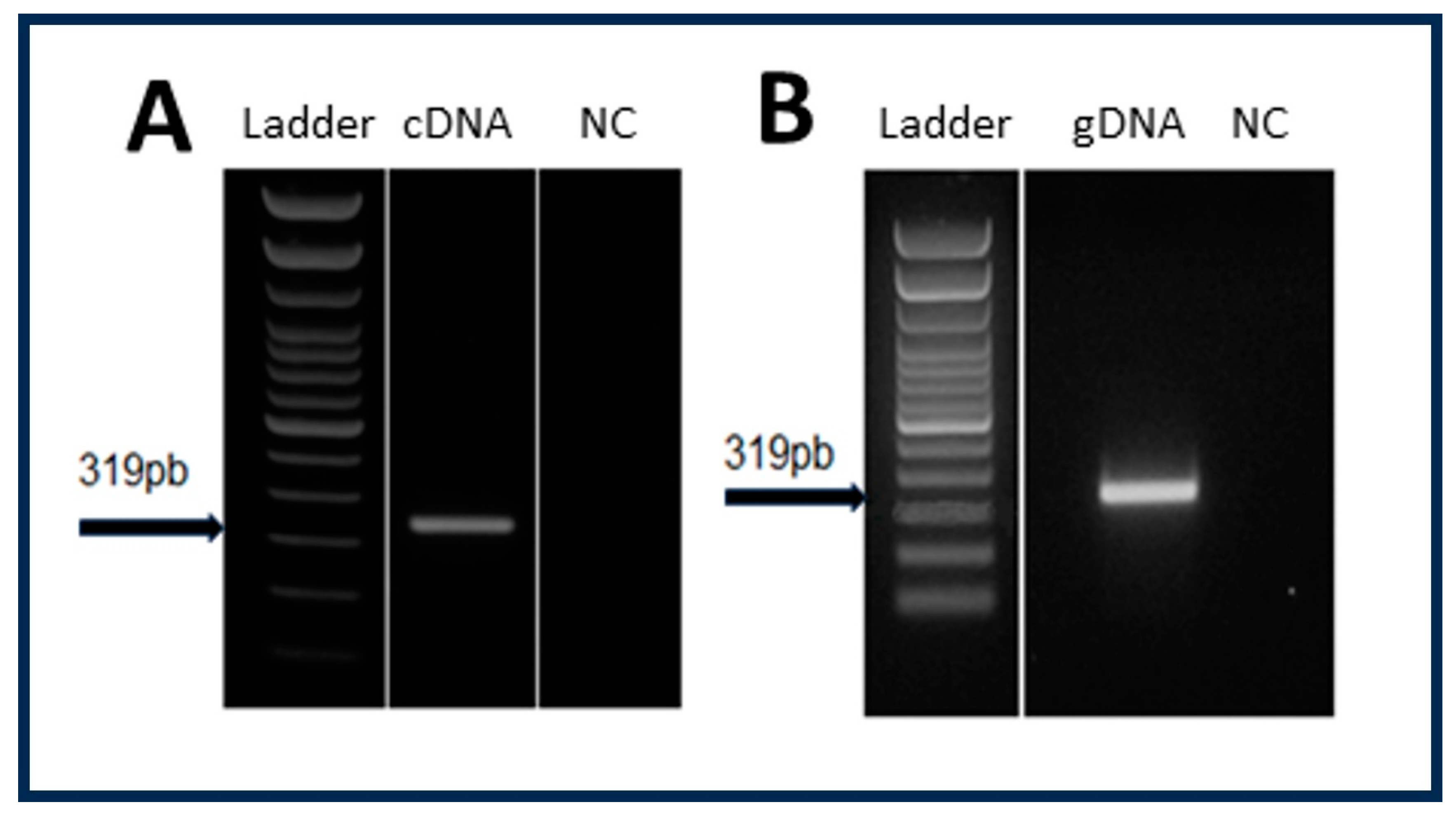
3.1. Determination of the Origin of RNA Sequences Coding for Viral Proteins in Exosomal Fraction of LL5 Cells
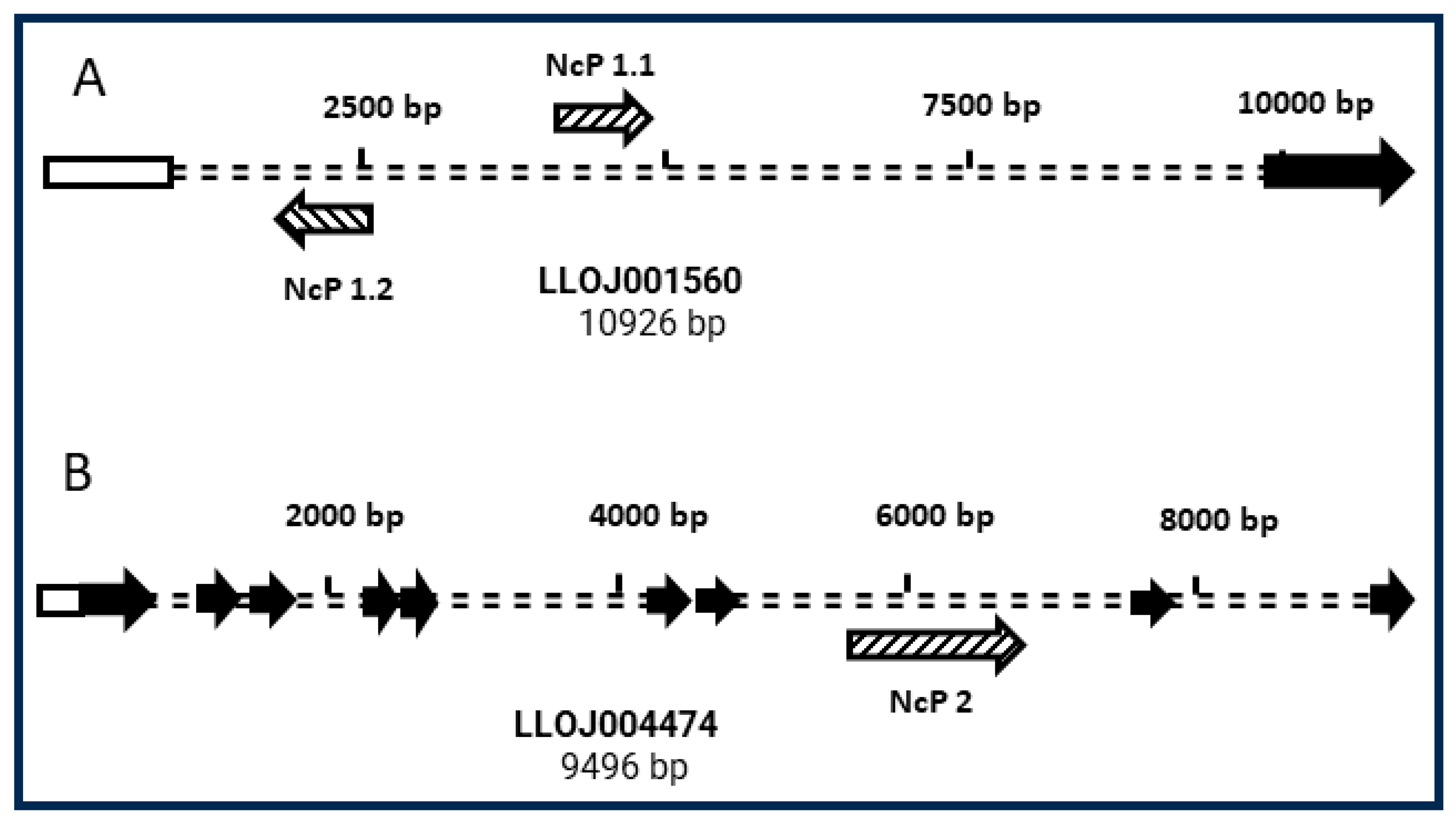
3.2. Genomic Context of NcP1.1 and NcP2 in the L. longipalpis Genome
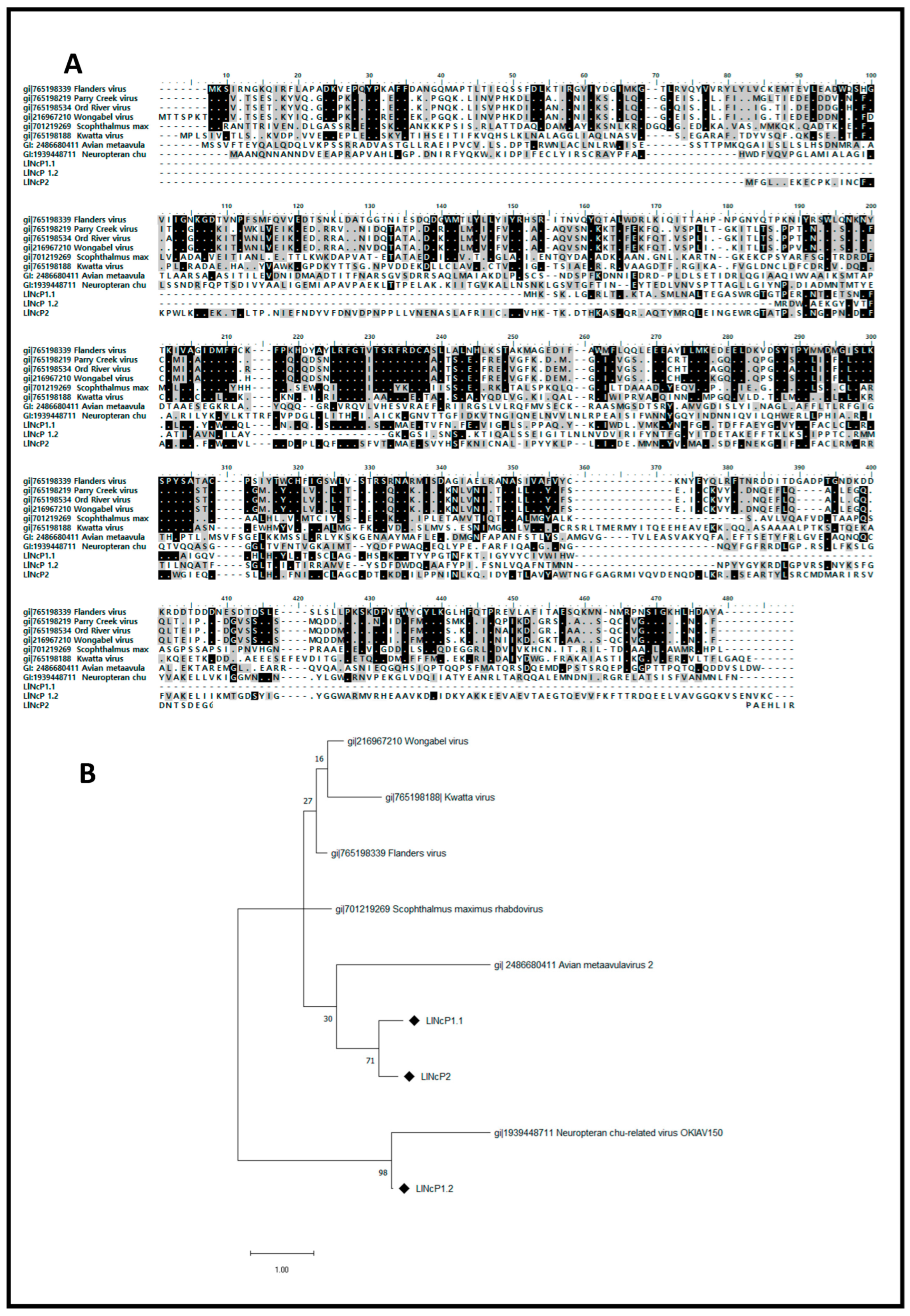
3.3. Multiple Alignment and Phylogenetic Analysis of EVE-Deduced Proteins sequences
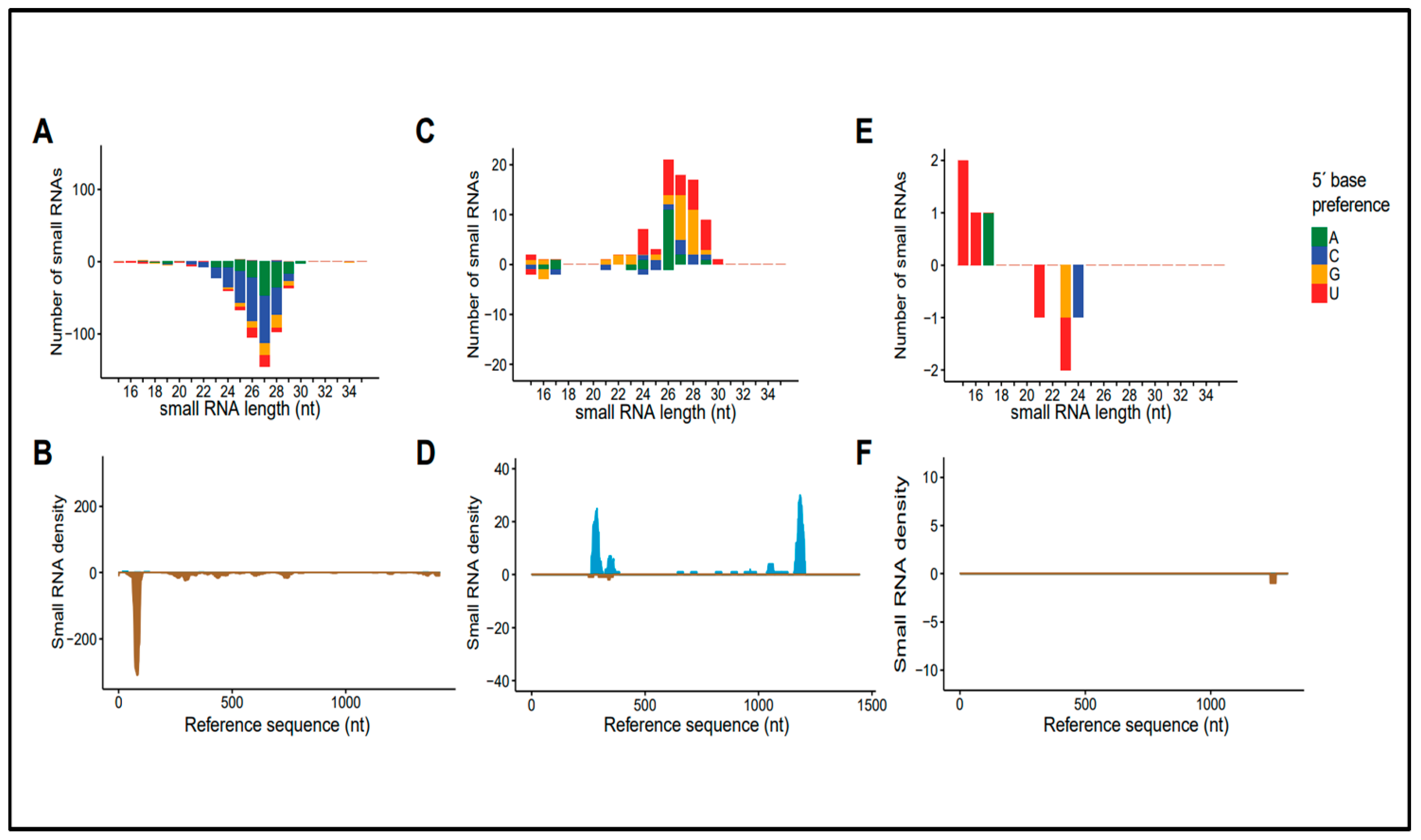
3.4. Molecular Characteristics of Small RNA Sequences Derived from Rhabdoviral Sequences
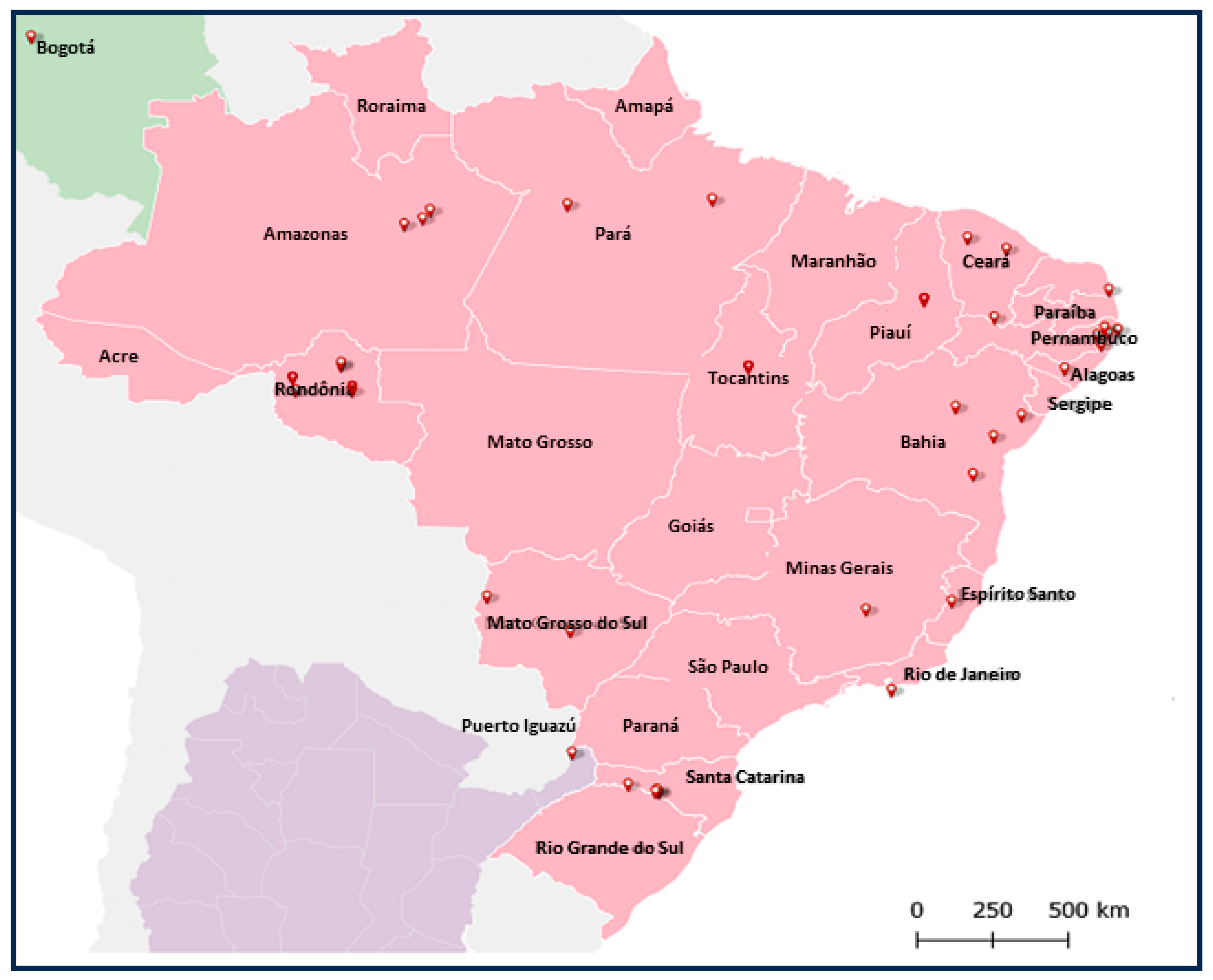
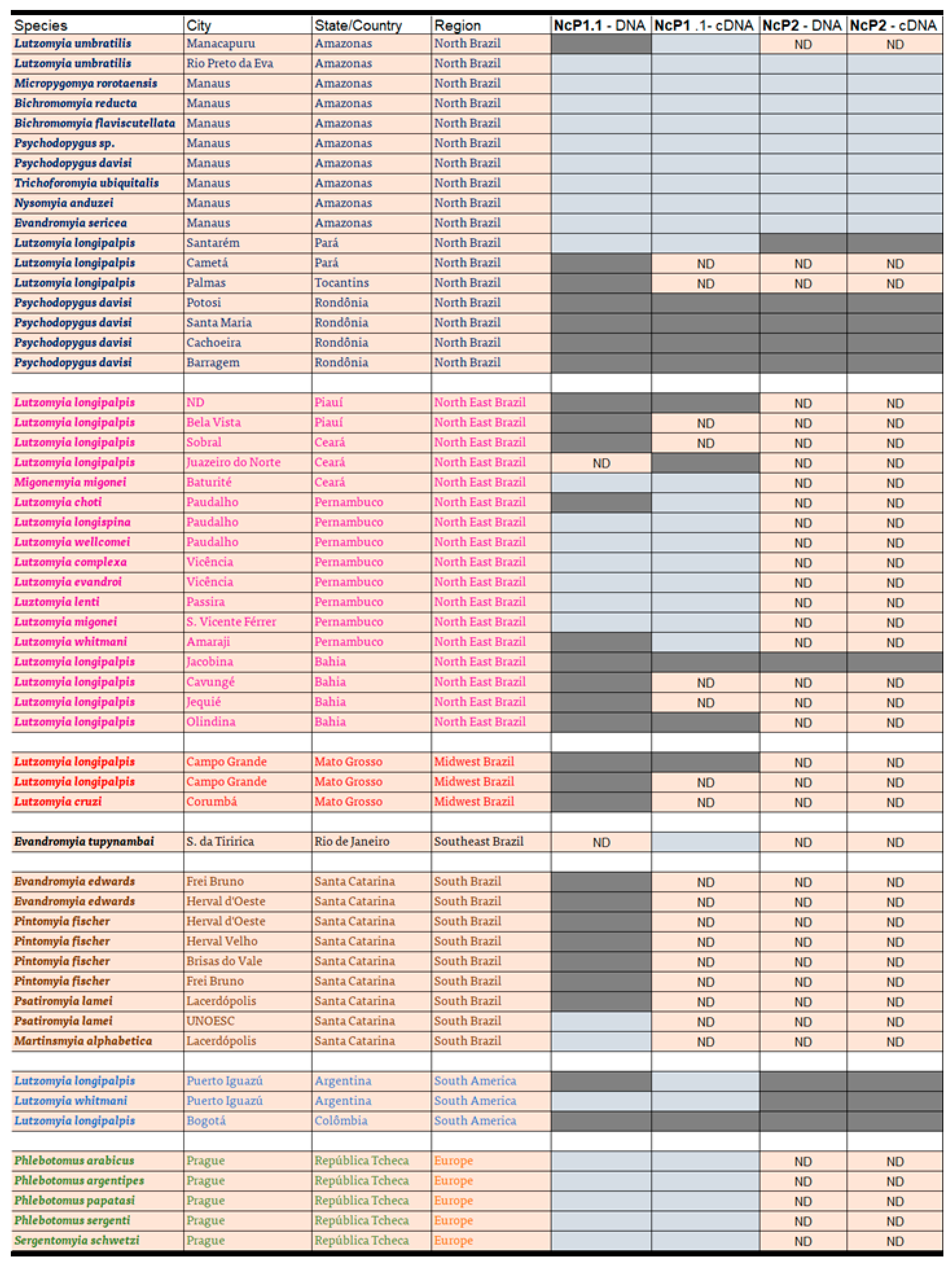
3.5. Distribution of Rhabdoviral Sequences in Sandfly Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; Coates, M.M.; Coggeshall, M.; et al. Global, Regional, and National Life Expectancy, All-Cause Mortality, and Cause-Specific Mortality for 249 Causes of Death, 1980–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef]
- Lainson, R.; Braga, R.R.; De Souza, A.A.; Pôvoa, M.M.; Ishikawa, E.A.; Silveira, F.T. Leishmania (Viannia) shawi sp. n., a parasite of monkeys, sloths and procyonids in Amazonian Brazil. Ann. Parasitol. Hum. Comp. 1989, 64, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Killick-Kendrick, R. Guide to the Identification and Geographic Distribution of Lutzomyia Sand Flies in Mexico, the West Indies, Central and South America (Diptera: Psychodidae). D. G. Young & M. A. Duncan. Memoirs of the American Entomological Institute no. 54. Gainesville, Florida, USA: Associated Publishers, 1994. 881 pp. US$ 85. ISBN 1-5666-054-2. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 125. [Google Scholar] [CrossRef]
- Bruschi, F.; Gradoni, L. (Eds.) The leishmaniases: Old Neglected Tropical Diseases; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Labbé, F.; Abdeladhim, M.; Abrudan, J.; Araki, A.S.; Araujo, R.N.; Arensburger, P.; Benoit, J.B.; Brazil, R.P.; Bruno, R.V.; Bueno da Silva Rivas, G.; et al. Genomic analysis of two phlebotomine sandfly vectors of Leishmania from the New and Old World. PLoS Negl. Trop. Dis. 2023, 17, e0010862. [Google Scholar] [CrossRef] [PubMed]
- Jancarova, M.; Polanska, N.; Volf, P.; Dvorak, V. The role of sandflies as vectors of viruses other than phleboviruses. J. Gen. Virol. 2023, 104, 001837. [Google Scholar] [CrossRef] [PubMed]
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro Surveill. 2010, 15, 19507. [Google Scholar] [CrossRef] [PubMed]
- Letchworth, G.J.; Rodriguez, L.L.; Del Cabarrera, J. Vesicular stomatitis. Vet. J. 1999, 157, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Travassos da Rosa, A.P.A.; Shope, R.E.; Pinheiro, F.P. Arbovirus research in the Brazilian Amazon. In Proceedings of the Fifth Symposium on Arbovirus Research in Australia, Brisbane, Australia, 28 August–1 September 1989; Uren, M.F., Blok, J., Manderson, L.H., Eds.; University of Queensland Medical School: Brisbane, Australia, 1989; pp. 4–8. [Google Scholar]
- Carvalho, M.S.; de Lara Pinto, A.Z.; Pinheiro, A.; Rodrigues, J.S.V.; Melo, F.L.; da Silva, L.A.; Ribeiro, B.M.; Dezengrini-Slhessarenko, R. Viola phlebovirus is a novel Phlebotomus fever serogroup member identified in Lutzomyia (Lutzomyia) longipalpis from Brazilian Pantanal. Parasit. Vectors 2018, 11, 405. [Google Scholar] [CrossRef]
- Tesh, R.B.; Boshell, J.; Young, D.G.; Morales, A.; Ferra de Carrasquilla, C.; Corredor, A.; Modi, G.B.; Travassos da Rosa, A.P.; McLean, R.G.; de Rodriguez, C.; et al. Characterization of five new phleboviruses recently isolated from sandflies in tropical America. Am. J. Trop. Med. Hyg. 1989, 40, 529–533. [Google Scholar] [CrossRef]
- Fonseca, P.; Ferreira, F.; da Silva, F.; Oliveira, L.S.; Marques, J.T.; Goes-Neto, A.; Aguiar, E.; Gruber, A. Characterization of a Novel Mitovirus of the Sandfly Lutzomyia longipalpis Using Genomic and Virus-Host Interaction Signatures. Viruses 2020, 13, 9. [Google Scholar] [CrossRef]
- Jern, P.; Coffin, J.M. Effects of retroviruses on host genome function. Annu. Rev. Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Honda, T.; Suzuki, Y.; Kobayashi, Y.; Daito, T.; Oshida, T.; Ikuta, K.; Jern, P.; Gojobori, T.; Coffin, J.M.; et al. Endogenous non-retroviral RNA virus elements in mammalian genomes. Nature 2010, 463, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Belyi, V.A.; Levine, A.J.; Skalka, A.M. Unexpected inheritance: Multiple integrations of ancient bornavirus and ebolavirus/marburgvirus sequences in vertebrate genomes. PLoS Pathog. 2010, 6, e1001030. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Feschotte, C. Genomic fossils calibrate the long-term evolution of hepadnaviruses. PLoS Biol. 2010, 8, e1000495. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Peng, Y.; Yi, X.; Jiang, D. Widespread endogenization of densoviruses and parvoviruses in animal and human genomes. J. Virol. 2011, 85, 9863–9876. [Google Scholar] [CrossRef]
- Veglia, A.J.; Bistolas, K.S.I.; Voolstra, C.R.; Hume, B.C.C.; Ruscheweyh, H.J.; Planes, S.; Allemand, D.; Boissin, E.; Wincker, P.; Poulain, J.; et al. Endogenous viral elements reveal associations between a non-retroviral RNA virus and symbiotic dinoflagellate genomes. Commun. Biol. 2023, 6, 566. [Google Scholar] [CrossRef]
- Fort, P.; Albertini, A.; Van-Hua, A.; Berthomieu, A.; Roche, S.; Delsuc, F.; Pasteur, N.; Capy, P.; Gaudin, Y.; Weill, M. Fossil rhabdoviral sequences integrated into arthropod genomes: Ontogeny, evolution, and potential functionality. Mol. Biol. Evol. 2012, 29, 381–390. [Google Scholar] [CrossRef]
- Vasilakis, N.; Widen, S.; Mayer, S.V.; Seymour, R.; Wood, T.G.; Popov, V.; Guzman, H.; da Rosa, A.P.; Ghedin, E.; Holmes, E.C. Niakha virus: A novel member of the family Rhabdoviridae isolated from phlebotomine sandflies in Senegal. Virology 2013, 444, 80–89. [Google Scholar] [CrossRef]
- Ter Horst, A.M.; Nigg, J.C.; Dekker, F.M.; Falk, B.W. Endogenous Viral Elements Are Widespread in Arthropod Genomes and Commonly Give Rise to PIWI-Interacting RNAs. J. Virol. 2019, 93, e02124-18. [Google Scholar] [CrossRef]
- Wallau, G.L. RNA virus EVEs in insect genomes. Curr. Opin. Insect Sci. 2022, 49, 42–47. Available online: https://www.sciencedirect.com/science/article/pii/S2214574521001267 (accessed on 1 February 2022). [CrossRef]
- Fujino, K.; Horie, M.; Honda, T.; Merriman, D.K.; Tomonaga, K. Inhibition of Borna disease virus replication by an endogenous bornavirus-like element in the ground squirrel genome. Proc. Natl. Acad. Sci. USA 2014, 111, 13175–13180. [Google Scholar] [CrossRef]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-derived DNA drives mosquito vector tolerance to arboviral infection. Nat. Commun. 2016, 7, 12410. [Google Scholar] [CrossRef]
- Volf, P.; Volfova, V. Establishment and maintenance of sand fly colonies. J. Vector Ecol. 2011, 36, S1–S9. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Aguiar, E.R.; Olmo, R.P.; Marques, J.T. Virus-derived small RNAs: Molecular footprints od host-pahogen interaction. Wiley Interdiscip. Rev. RNA 2016, 7, 824–837. [Google Scholar] [CrossRef]
- Lourenço-de-Oliveira, R.; Marques, J.T.; Sreenu, V.B.; Atyame Nten, C.; Aguiar, E.R.G.R.; Varjak, M.; Kohl, A.; Failloux, A.B. Culex quinquefasciatus mosquitoes do not support replication of Zika virus. J. Gen. Virol. 2018, 99, 258–264. [Google Scholar] [CrossRef]
- Marconcini, M.; Pischedda, E.; Houé, V.; Palatini, U.; Lozada-Chávez, N.; Sogliani, D.; Failloux, A.B.; Bonizzoni, M. Profile of Small RNAs, vDNA Forms and Viral Integrations in Late Chikungunya Virus Infection of Aedes albopictus Mosquitoes. Viruses 2021, 13, 553. [Google Scholar] [CrossRef]
- Olmo, R.P.; Todjro, Y.M.; Aguiar, E.R.; de Almeida, J.P.P.; Ferreira, F.V.; Armache, J.N.; de Faria, I.J.; Ferreira, A.G.; Amadou, S.C.; Silva, A.T.S.; et al. Mosquito vector competence for dengue is modulated by insect-specific viruses. Nat. Microbiol. 2023, 8, 135–149. [Google Scholar] [CrossRef]
- Chiba, S.; Kondo, H.; Tani, A.; Saisho, D.; Sakamoto, W.; Kanematsu, S.; Suzuki, N. Widespread endogenization of genome sequences of non-retroviral RNA viruses into plant genomes. PLoS Pathog. 2011, 7, e1002146. [Google Scholar] [CrossRef]
- Aguiar, E.R.G.R.; de Almeida, J.P.P.; Queiroz, L.R.; Oliveira, L.S.; Olmo, R.P.; de Faria, I.J.D.S.; Imler, J.L.; Gruber, A.; Matthews, B.J.; Marques, J.T. A single unidirectional piRNA cluster similar to the flamenco locus is the major source of EVE-derived transcription and small RNAs in Aedes aegypti mosquitoes. RNA 2020, 26, 581–594. [Google Scholar] [CrossRef]
- Kryukov, K.; Ueda, M.T.; Imanishi, T.; Nakagawa, S. Systematic survey of non-retroviral virus-like elements in eukaryotic genomes. Virus Res. 2019, 262, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Frangeul, L.; Dickson, L.B.; Blanc, H.; Verdier, Y.; Vinh, J.; Lambrechts, L.; Saleh, M.C. Uncovering the Repertoire of Endogenous Flaviviral Elements in Aedes mosquito Genomes. J. Virol. 2017, 91, e00571-17. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.N.; Chao, A.; Baker, J.; Chang, L.; Qiao, D.; Lin, H. A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal. Genes Dev. 1998, 12, 3715–3727. [Google Scholar] [CrossRef] [PubMed]
- Léger, P.; Lara, E.; Jagla, B.; Sismeiro, O.; Mansuroglu, Z.; Coppée, J.Y.; Bonnefoy, E.; Bouloy, M. Dicer-2- and Piwi-mediated RNA interference in Rift Valley fever virus-infected mosquito cells. J. Virol. 2013, 87, 1631–1648. [Google Scholar] [CrossRef] [PubMed]
- Varjak, M.; Maringer, K.; Watson, M.; Sreenu, V.B.; Fredericks, A.C.; Pondeville, E.; Donald, C.L.; Sterk, J.; Kean, J.; Vazeille, M.; et al. Aedes aegypti Piwi4 Is a Noncanonical PIWI Protein Involved in Antiviral Responses. mSphere 2017, 2, e00144-17. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.V.; Aguiar, E.R.G.R.; Olmo, R.P.; de Oliveira, K.P.V.; Silva, E.G.; Sant’Anna, M.R.V.; Gontijo, N.F.; Kroon, E.G.; Imler, J.L.; Marques, J.T. The small non-coding RNA response to virus infection in the Leishmania vector Lutzomyia longipalpis. PLoS Negl. Trop. Dis. 2018, 12, e0006569. [Google Scholar] [CrossRef]
- Martins-da-Silva, A.; Telleria, E.L.; Batista, M.; Marchini, F.K.; Traub-Csekö, Y.M.; Tempone, A.J. Identification of Secreted Proteins Involved in Nonspecific dsRNA-Mediated Lutzomyia longipalpis LL5 Cell Antiviral Response. Viruses 2018, 10, 43. [Google Scholar] [CrossRef]
- Souza Freitas, M.T.; Ríos-Velasquez, C.M.; Costa, C.R., Jr.; Figueirêdo, C.A., Jr.; Aragão, N.C.; da Silva, L.G.; de Aragão Batista, M.V.; Balbino, T.C.; Pessoa, F.A.; de Queiroz Balbino, V. Phenotypic and genotypic variations among three allopatric populations of Lutzomyia umbratilis, main vector of Leishmania guyanensis. Parasit. Vectors 2015, 8, 448. [Google Scholar] [CrossRef] [PubMed]
- Sousa-Paula, L.C.; da Silva, L.G.; da Silva Junior, W.J.; Figueiredo Júnior, C.A.S.; Costa, C.H.N.; Pessoa, F.A.C.; Dantas-Torres, F. Genetic structure of allopatric populations of Lutzomyia longipalpis sensu lato in Brazil. Acta Trop. 2021, 222, 106031. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Amplicon Length (nt) |
|---|---|---|
| Histone forward | 5′ GAAAAGCAGGCAAACACTC 3′ | 500 pb |
| Histone reverse | 5′ GAAGGATGGGTGGAAAGG 3′ | |
| NcP1.1 forward | 5′ GGAACCCCAGAACGATACAA 3′ | 319 pb |
| NcP1.1 reverse | 5′ GACAGAGGCACGCGAAGTAT 3′ | |
| NcP2 forward | 5′ TTCGAAGTGTCGCTTGCAGCC3′ | 415 pb |
| NcP2 reverse | 5′ GCAACCCCAAACTCCTACAA 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tempone, A.J.; Zezza-Ramalho, M.d.S.; Borely, D.; Pitaluga, A.N.; Brazil, R.P.; Brandão-Filho, S.P.; Pessoa, F.A.C.; Bruno, R.V.; Carvalho-Costa, F.A.; Salomón, O.D.; et al. Rhabdoviral Endogenous Sequences Identified in the Leishmaniasis Vector Lutzomyia longipalpis Are Widespread in Sandflies from South America. Viruses 2024, 16, 395. https://doi.org/10.3390/v16030395
Tempone AJ, Zezza-Ramalho MdS, Borely D, Pitaluga AN, Brazil RP, Brandão-Filho SP, Pessoa FAC, Bruno RV, Carvalho-Costa FA, Salomón OD, et al. Rhabdoviral Endogenous Sequences Identified in the Leishmaniasis Vector Lutzomyia longipalpis Are Widespread in Sandflies from South America. Viruses. 2024; 16(3):395. https://doi.org/10.3390/v16030395
Chicago/Turabian StyleTempone, Antonio J., Monique de Souza Zezza-Ramalho, Daniel Borely, André N. Pitaluga, Reginaldo Peçanha Brazil, Sinval P. Brandão-Filho, Felipe A. C. Pessoa, Rafaela V. Bruno, Filipe A. Carvalho-Costa, Oscar D. Salomón, and et al. 2024. "Rhabdoviral Endogenous Sequences Identified in the Leishmaniasis Vector Lutzomyia longipalpis Are Widespread in Sandflies from South America" Viruses 16, no. 3: 395. https://doi.org/10.3390/v16030395