
Replication and Expression of the Consensus Genome of Hepatitis B Virus Genotype C from the Chinese Population
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Consensus Sequence
2.2. Plasmid Construction
2.3. Cell Culture and Transfection
2.4. Quantitative Analysis of Extracellular HBV DNA and Intracellular HBV pgRNA
2.5. Immunofluorescence Staining
2.6. Quantification Assays for HBsAg and HBeAg
2.7. Hydrodynamic Injection and In Vivo Assays
2.8. Statistical Analysis
3. Results
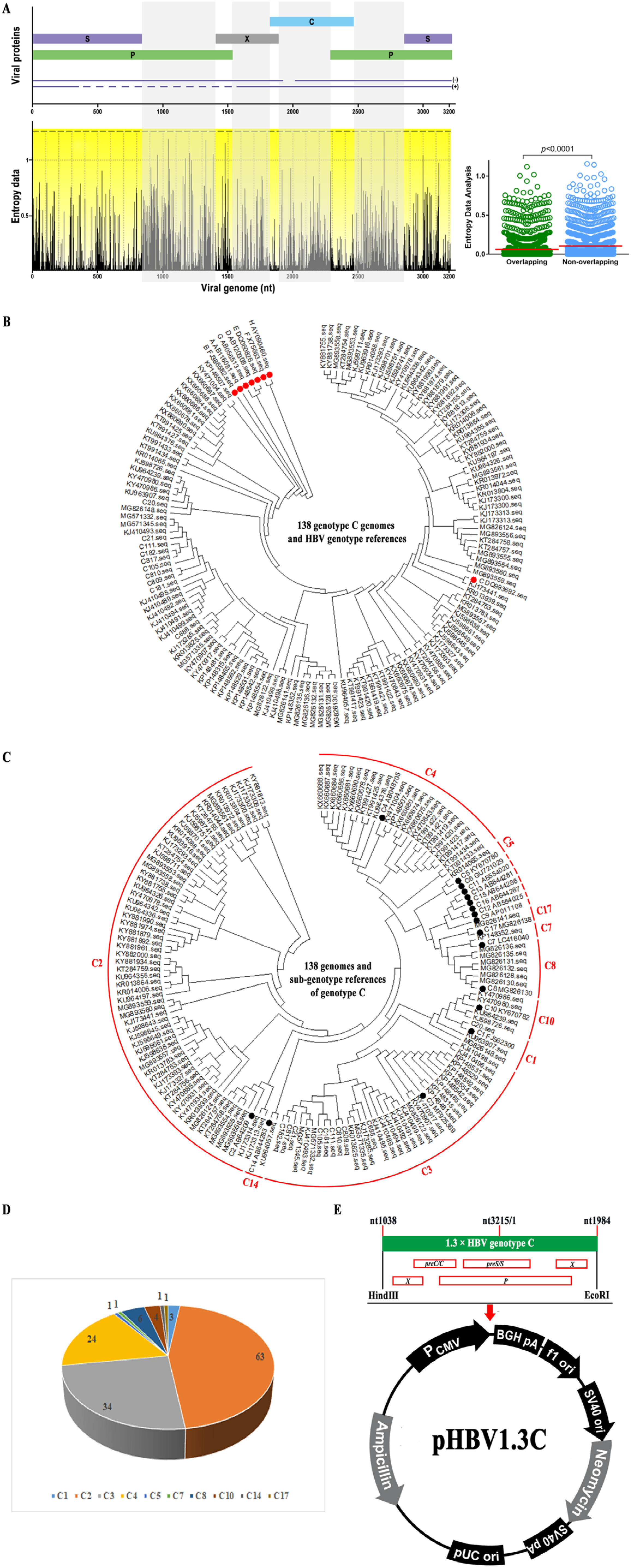
3.1. Consensus Sequence of HBV Genotype C from the Chinese Population
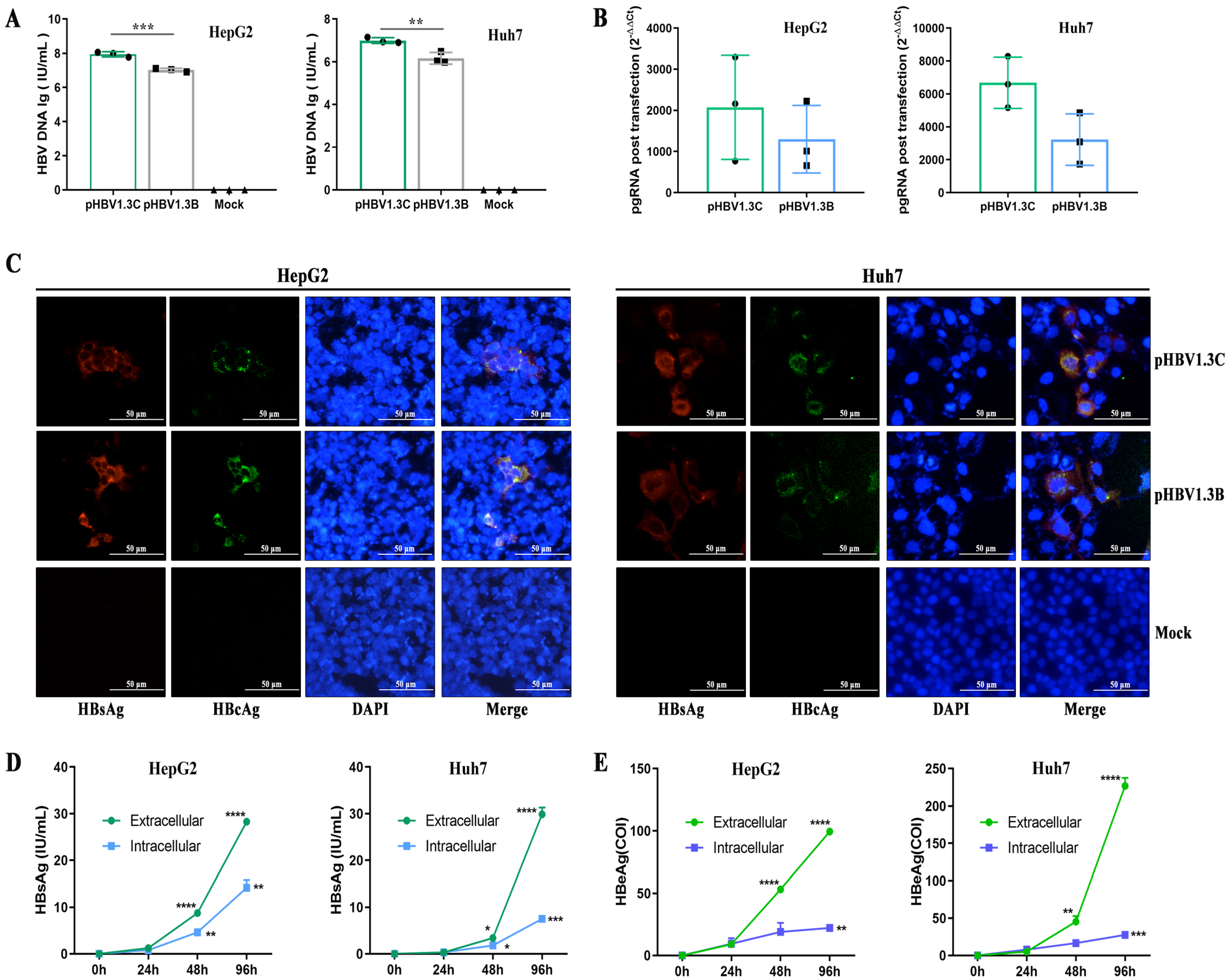
3.2. Replication of pHBV1.3C in HepG2 Cells and Huh7 Cells
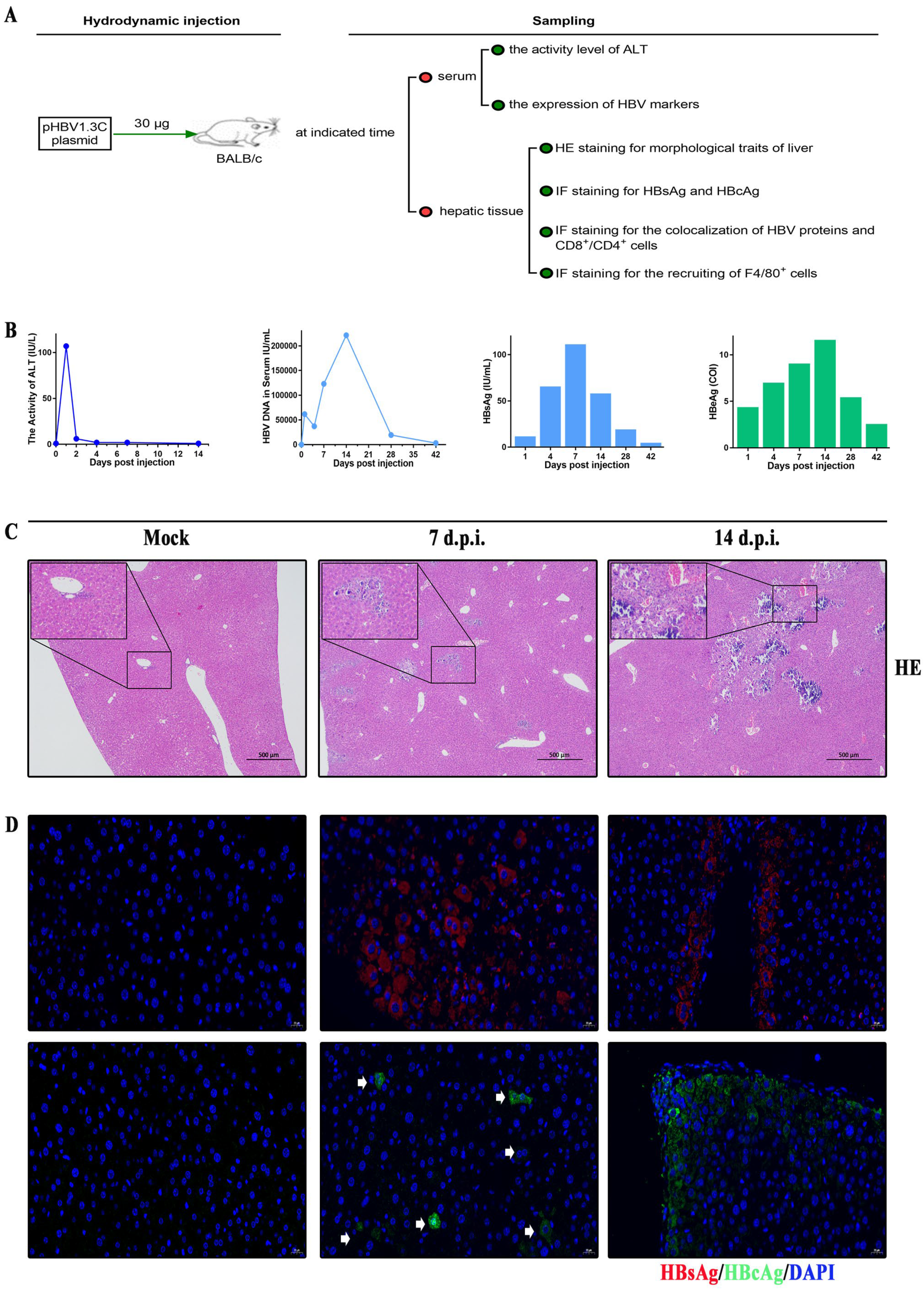
3.3. Kinetic Viremia of the BALB/c Recipients Induced by pHBV1.3C
3.4. Histopathological Characteristics in BALB/c Recipients Injected with pHBV1.3C
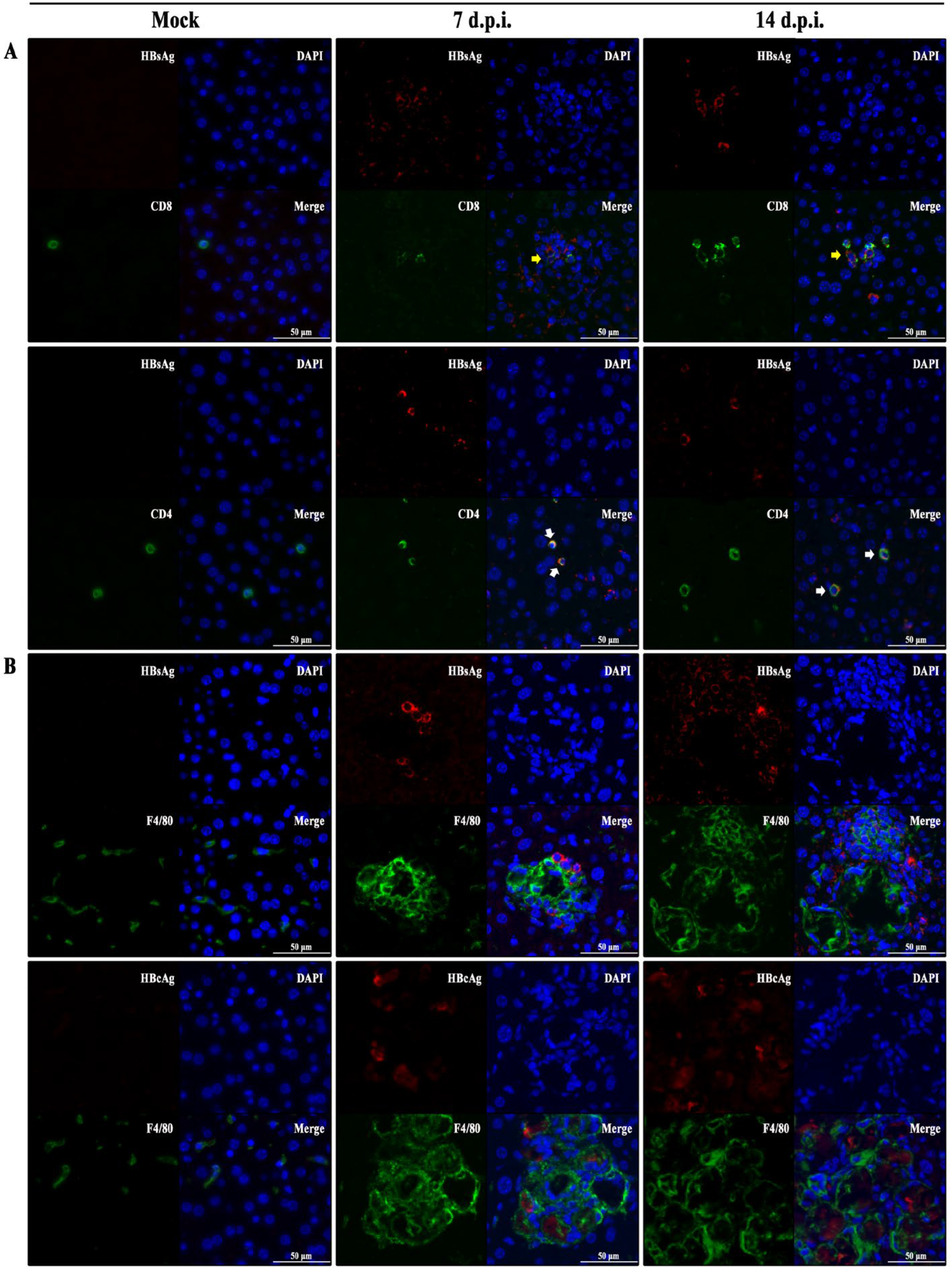
3.5. Hepatic Immunopathological Characteristics in BALB/c Recipients Injected with pHBV1.3C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Hepatitis Report 2017; World Health Organization: Geneva, Switzerland, 2017; pp. 21–24. [Google Scholar]
- Razavi, H. Global Epidemiology of Viral Hepatitis. Gastroenterol. Clin. N. Am. 2020, 49, 179–189. [Google Scholar] [CrossRef] [PubMed]
- El-Serag, H.B. Epidemiology of Viral Hepatitis and Hepatocellular Carcinoma. Gastroenterology 2012, 142, 1264–1273.e1. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne, R.J.; Bagga, S.; Bouchard, M.J. Hepatitis B virus molecular biology and pathogenesis. Hepatoma Res. 2016, 2, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Waheed, Y.; Siddiq, M.; Jamil, Z.; Najmi, M.H. Hepatitis elimination by 2030: Progress and challenges. World J. Gastroenterol. 2018, 24, 4959–4961. [Google Scholar] [CrossRef]
- Razavi-Shearer, D.; Gamkrelidze, I.; Nguyen, M.H.; Chen, D.-S.; Van Damme, P.; Abbas, Z.; Abdulla, M.; Abou Rached, A.; Adda, D.; Aho, I.; et al. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: A modelling study. Lancet Gastroenterol. Hepatol. 2018, 3, 383–403. [Google Scholar] [CrossRef]
- Liu, J.; Liang, W.; Jing, W.; Liu, M. Countdown to 2030: Eliminating hepatitis B disease, China. Bull. World Health Organ. 2019, 97, 230–238. [Google Scholar] [CrossRef]
- Karayiannis, P. Hepatitis B virus: Virology, molecular biology, life cycle and intrahepatic spread. Hepatol. Int. 2017, 11, 500–508. [Google Scholar] [CrossRef]
- Coffin, C.S.; Zhou, K.; Terrault, N.A. New and Old Biomarkers for Diagnosis and Management of Chronic Hepatitis B Virus Infection. Gastroenterology 2019, 156, 355–368. [Google Scholar] [CrossRef]
- Tong, S.; Revill, P. Overview of viral replication and genetic variability. J. Hepatol. 2016, 64, 4–16. [Google Scholar] [CrossRef]
- Hu, J.; Liu, K. Complete and Incomplete Hepatitis B Virus Particles: Formation, Function, and Application. Viruses 2017, 9, 56. [Google Scholar] [CrossRef]
- Tatematsu, K.; Tanaka, Y.; Kurbanov, F.; Sugauchi, F.; Mano, S.; Maeshiro, T.; Nakayoshi, T.; Wakuta, M.; Miyakawa, Y.; Mizokami, M. A genetic variant of hepatitis B virus divergent from known human and ape genotypes isolated from a Japanese patient and provisionally assigned to new genotype J. J. Virol. 2009, 83, 10538–10547. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shan, X.; Liang, Z.; Shan, Y.; Huang, W.; Zhang, D.; Zen, A.; Zhou, X.; Zhao, Y.; Gong, X.; et al. Deep sequencing analysis of HBV genotype shift and correlation with antiviral efficiency during adefovir dipivoxil therapy. PLoS ONE 2015, 10, e0131337. [Google Scholar] [CrossRef] [PubMed]
- Araujo, N.M.; Teles, S.A.; Spitz, N. Comprehensive Analysis of Clinically Significant Hepatitis B Virus Mutations in Relation to Genotype, Subgenotype and Geographic Region. Front. Microbiol. 2020, 11, 616023. [Google Scholar] [CrossRef]
- Rajoriya, N.; Combet, C.; Zoulim, F.; Janssen, H.L.A. How viral genetic variants and genotypes influence disease and treatment outcome of chronic hepatitis B. Time for an individualised approach? J. Hepatol. 2017, 67, 1281–1297. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Wang, J.Q.; Wang, R.; Zhao, Q.; Li, L.; Zhang, J.P.; Shen, T. Hepatitis B virus genotypes and genome characteristics in China. World J. Gastroenterol. 2015, 21, 6684–6697. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Jia, J. Hepatitis B virus genotypes: Epidemiological and clinical relevance in Asia. Hepatol. Int. 2016, 10, 854–860. [Google Scholar] [CrossRef]
- Norder, H.; Courouce, A.M.; Coursaget, P.; Echevarria, J.M.; Lee, S.D.; Mushahwar, I.K.; Robertson, B.H.; Locarnini, S.; Magnius, L.O. Genetic diversity of hepatitis B virus strains derived worldwide: Genotypes, subgenotypes, and HBsAg subtypes. Intervirology 2004, 47, 289–309. [Google Scholar] [CrossRef]
- Bello, K.E.; Jusoh, T.N.A.M.; Irekeola, A.A.; Abu, N.; Amin, N.A.Z.M.; Mustaffa, N.; Shueb, R.H. A Recent Prevalence of Hepatitis B Virus (HBV) Genotypes and Subtypes in Asia: A Systematic Review and Meta-Analysis. Healthcare 2023, 11, 1011. [Google Scholar] [CrossRef]
- Feng, Y.; Ran, J.; Feng, Y.-M.; Miao, J.; Zhao, Y.; Jia, Y.; Li, Z.; Yue, W.; Xia, X. Genetic diversity of hepatitis B virus in Yunnan, China: Identification of novel subgenotype C17, an intergenotypic B/I recombinant, and B/C recombinants. J. Gen. Virol. 2020, 101, 972–981. [Google Scholar] [CrossRef]
- Kramvis, A. Genotypes and genetic variability of hepatitis B virus. Intervirology 2014, 57, 141–150. [Google Scholar] [CrossRef]
- Orito, E. A case-control study for clinical and molecular biological differences between hepatitis B viruses of genotypes B and C. Hepatology 2001, 33, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.L.; Hui, A.Y.; Wong, M.L.; Tse, A.M.; Hung, L.C.; Wong, V.W.; Sung, J.J. Genotype C hepatitis B virus infection is associated with an increased risk of hepatocellular carcinoma. Gut 2004, 53, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Sunbul, M. Hepatitis B virus genotypes: Global distribution and clinical importance. World J. Gastroenterol. 2014, 20, 5427. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, G.; Chen, M.; Yang, Y.; Xie, Y.; Kong, X. A Novel Hydrodynamic Injection Mouse Model of HBV Genotype C for the Study of HBV Biology and the Anti-Viral Activity of Lamivudine. Hepat. Mon. 2016, 16, e34420. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Harrison, T.J.; Sabin, C.A.; Li, G.J.; Huang, G.M.; Yang, J.Y.; Wang, X.Y.; Li, H.; Liu, M.H.; Fang, Z.L. The Effect of HBV Genotype C on the Development of HCC Differs between Wild-Type Viruses and Those with BCP Double Mutations (T(1762)A(1764)). Hepat. Mon. 2014, 14, e16214. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, A.; Liu, Y.; Cen, S.; Zhang, Q. In vitro investigation of HBV clinical isolates from Chinese patients reveals that genotype C isolates possess higher infectivity than genotype B isolates. Virol. Sin. 2022, 37, 398–407. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Huang, J.; Xu, R.; Liao, Q.; Shan, Z.; Zheng, Y.; Rong, X.; Tang, X.; Li, T.; et al. Novel hepatitis B virus surface antigen mutations associated with occult genotype B hepatitis B virus infection affect HBsAg detection. J. Viral Hepat. 2020, 27, 915–921. [Google Scholar] [CrossRef]
- Wang, H. Molecular Biological Characteristics of Hepatitis B Virus S Gene and Its Effect on the Expression of Hepatitis B Surface Antigen in Blood Donors with Occult Hepatitis B Virus Infection in Guangzhou Doctoral Dissertation 2020. Ph.D. Thesis, Southern Medical University, Guangzhou, China, 2021. [Google Scholar]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Hu, J.; Lin, Y.Y.; Chen, P.J.; Watashi, K.; Wakita, T. Cell and Animal Models for Studying Hepatitis B Virus Infection and Drug Development. Gastroenterology 2019, 156, 338–354. [Google Scholar] [CrossRef]
- Austyn, J.; Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur. J. Immunol. 1981, 11, 805–815. [Google Scholar] [CrossRef]
- Pawlotsky, J.M. The concept of hepatitis B virus mutant escape. J. Clin. Virol. 2005, 34 (Suppl. S1), 125–129. [Google Scholar] [CrossRef] [PubMed]
- Valaydon, Z.S.; Locarnini, S.A. The Virological Aspects of Hepatitis B. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, D.M.; Huang, S.Y.; Yu, J.L.; Zhang, D.H.; Gong, Q.M.; Zhang, X.X. Clinical Implications of Evolutionary Patterns of Homologous, Full-Length Hepatitis B Virus Quasispecies in Different Hosts after Perinatal Infection. J. Clin. Microbiol. 2014, 52, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liao, F.; Xie, J.; Gao, W.; Wang, M.; Huang, J.; Xu, R.; Liao, Q.; Shan, Z.; Zheng, Y.; et al. E2 Site Mutations in S Protein Strongly Affect Hepatitis B Surface Antigen Detection in the Occult Hepatitis B Virus. Front. Microbiol. 2021, 12, 664833. [Google Scholar] [CrossRef] [PubMed]
- Sitia, G.; Isogawa, M.; Iannacone, M.; Campbell, I.L.; Chisari, F.V.; Guidotti, L.G. MMPs are required for recruitment of antigen-nonspecific mononuclear cells into the liver by CTLs. J. Clin. Investig. 2004, 113, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.L.; Althage, A.; Chung, J.; Chisari, F.V. Hydrodynamic injection of viral DNA: A mouse model of acute hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2002, 99, 13825–13830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xia, J.; Sun, B.; Dai, Y.; Li, X.; Schlaak, J.F.; Lu, M. In vitro and in vivo replication of a chemically synthesized consensus genome of hepatitis B virus genotype B. J. Virol. Methods 2015, 213, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Nassal, M. Stable HepG2- and Huh7-based human hepatoma cell lines for efficient regulated expression of infectious hepatitis B virus. J. Hepatol. 2006, 45, 636–645. [Google Scholar] [CrossRef]
- Dandri, M.; Petersen, J. Animal models of HBV infection. Best Pr. Res. Clin. Gastroenterol. 2017, 31, 273–279. [Google Scholar] [CrossRef]
- Chen, J.; Liu, B.; Tang, X.; Zheng, X.; Lu, J.; Zhang, L.; Wang, W.; Candotti, D.; Fu, Y.; Allain, J.-P.; et al. Role of core protein mutations in the development of occult HBV infection. J. Hepatol. 2021, 74, 1303–1314. [Google Scholar] [CrossRef]
- Huang, L.-R.; Wu, H.-L.; Chen, P.-J.; Chen, D.-S. An immunocompetent mouse model for the tolerance of human chronic hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2006, 47, 17862–17867. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, L.; Zhu, D.; Peng, H.; Su, L.; Fu, Y.-X.; Zhang, L. A mouse model for HBV immunotolerance and immunotherapy. Cell. Mol. Immunol. 2013, 11, 71–78. [Google Scholar]
- Suresh, M.; Czerwinski, S.; Murreddu, M.G.; Kallakury, B.V.; Ramesh, A.; Gudima, S.O. Innate and adaptive immunity associated with resolution of acute woodchuck hepatitis virus infection in adult woodchucks. PloS Pathog. 2019, 15, e1008248. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cao, M.; Wei, Q.L.; Zhao, Z.H.; Xiang, Q.; Wang, H.J.; Zhang, H.T.; Lai, G.Q. A new model mimicking persistent HBV e antigen-negative infection using covalently closed circular DNA in immunocompetent mice. PLoS ONE 2017, 12, e0175992. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Hwang, L.-Y.; Hsu, H.C.; Lee, C.Y.; Beasley, R.P. Prospective study of asymptomatic HBsAg carrier children infected in the perinatal period clinical and liver histologic studies. Hepatology 1988, 8, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Dusseaux, M.; Masse-Ranson, G.; Darche, S.; Ahodantin, J.; Li, Y.; Fiquet, O.; Beaumont, E.; Moreau, P.; Riviere, L.; Neuveut, C.; et al. Viral Load Affects the Immune Response to HBV in Mice with Humanized Immune System and Liver. Gastroenterology 2017, 153, 1647–1661. [Google Scholar] [CrossRef] [PubMed]
- Shiina, M.; Yamada, N.; Sugiyama, R.; Murayama, A.; Aly, H.H.; Muramatsu, M.; Wakita, T.; Imawari, M.; Kato, T. Hepatitis B Virus Genotype-Dependent Vulnerability of Infected Cells to Immune Reaction in the Early Phase of Infection. Front. Microbiol. 2019, 10, 2427. [Google Scholar] [CrossRef]
- Bertoletti, A.; Ferrari, C. Adaptive immunity in HBV infection. J. Hepatol. 2016, 64, 71–83. [Google Scholar] [CrossRef]
- Song, B.-C.; Cho, Y.-K.; Jwa, H.; Choi, E.K.; Kim, H.U.; Song, H.J.; Na, S.-Y.; Boo, S.-J.; Jeong, S.U. Is it necessary to delay antiviral therapy for 3-6 months to anticipate HBeAg seroconversion in patients with HBeAg-positive chronic hepatitis B in endemic areas of HBV genotype C? Clin. Mol. Hepatol. 2014, 20, 355. [Google Scholar] [CrossRef]
- Zheng, J.R.; Wang, Z.L.; Feng, B. Hepatitis B functional cure and immune response. Front. Immunol. 2022, 13, 1075916. [Google Scholar] [CrossRef]
- Krebs, K.; Bottinger, N.; Huang, L.R.; Chmielewski, M.; Arzberger, S.; Gasteiger, G.; Jager, C.; Schmitt, E.; Bohne, F.; Aichler, M.; et al. T cells expressing a chimeric antigen receptor that binds hepatitis B virus envelope proteins control virus replication in mice. Gastroenterology 2013, 145, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Kah, J.; Koh, S.; Volz, T.; Ceccarello, E.; Allweiss, L.; Lutgehetmann, M.; Bertoletti, A.; Dandri, M. Lymphocytes transiently expressing virus-specific T cell receptors reduce hepatitis B virus infection. J. Clin. Investig. 2017, 127, 3177–3188. [Google Scholar] [CrossRef]
- Xie, Y.; Zhong, K.B.; Hu, Y.; Xi, Y.L.; Guan, S.X.; Xu, M.; Lin, Y.; Liu, F.Y.; Zhou, W.J.; Gao, Y. Liver infiltration of multiple immune cells during the process of acute liver injury and repair. World J. Gastroenterol. 2022, 28, 6537–6550. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.C.; Sung, P.S.; Park, S.H. Immune responses and immunopathology in acute and chronic viral hepatitis. Nat. Rev. Immunol. 2016, 16, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.L.; Zhou, S.N.; Hu, W.; Li, J.; Zhou, M.J.; Li, X.Y.; Wang, Y.Y.; Zhang, P.; Chen, S.Y.; Fan, X.; et al. Metabolic interventions improve HBV envelope-specific T-cell responses in patients with chronic hepatitis B. Hepatol. Int. 2023, 17, 1125–1138. [Google Scholar] [CrossRef]
- Yang, P.L.; Althage, A.; Chung, J.; Maier, H.; Wieland, S.; Isogawa, M.; Chisari, F.V. Immune effectors required for hepatitis B virus clearance. Proc. Natl. Acad. Sci. USA 2010, 107, 798–802. [Google Scholar] [CrossRef]
- Gehring, A.J.; Sun, D.; Kennedy, P.T.F.; Hoen, E.N.; Lim, S.G.; Wasser, S.; Selden, C.; Maini, M.K.; Davis, D.M.; Nassal, M.; et al. The level of viral antigen presented by hepatocytes influences CD8 T-cell function. J. Virol. 2007, 81, 2940–2949. [Google Scholar] [CrossRef]
- Bertoletti, A.; Maini, M.K.; Ferrari, C. The host-pathogen interaction during HBV infection: Immunological controversies. Antivir. Ther. 2010, 15 (Suppl. S3), 15–24. [Google Scholar] [CrossRef]
- Boltjes, A.; Movita, D.; Boonstra, A.; Woltman, A.M. The role of Kupffer cells in hepatitis B and hepatitis C virus infections. J. Hepatol. 2014, 61, 660–671. [Google Scholar] [CrossRef]
- Kinoshita, M.; Uchida, T.; Sato, A.; Nakashima, M.; Nakashima, H.; Shono, S.; Habu, Y.; Miyazaki, H.; Hiroi, S.; Seki, S. Characterization of two F4/80-positive Kupffer cell subsets by their function and phenotype in mice. J. Hepatol. 2010, 53, 903–910. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, F.; Xie, J.; Du, R.; Gao, W.; Lan, L.; Wang, M.; Rong, X.; Fu, Y.; Wang, H. Replication and Expression of the Consensus Genome of Hepatitis B Virus Genotype C from the Chinese Population. Viruses 2023, 15, 2302. https://doi.org/10.3390/v15122302
Liao F, Xie J, Du R, Gao W, Lan L, Wang M, Rong X, Fu Y, Wang H. Replication and Expression of the Consensus Genome of Hepatitis B Virus Genotype C from the Chinese Population. Viruses. 2023; 15(12):2302. https://doi.org/10.3390/v15122302
Chicago/Turabian StyleLiao, Fenfang, Junmou Xie, Rongsong Du, Wenbo Gao, Lanyin Lan, Min Wang, Xia Rong, Yongshui Fu, and Hao Wang. 2023. "Replication and Expression of the Consensus Genome of Hepatitis B Virus Genotype C from the Chinese Population" Viruses 15, no. 12: 2302. https://doi.org/10.3390/v15122302