
Higher HIV-1 Env gp120-Specific Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity Is Associated with Lower Levels of Defective HIV-1 Provirus
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cohort
2.2. HIV RNA/DNA Measurement
2.3. Rapid and Fluorometric ADCC Assay
2.4. Statistical Methods
2.5. Cell Lines
3. Results
3.1. Study Population and Baseline Characteristics
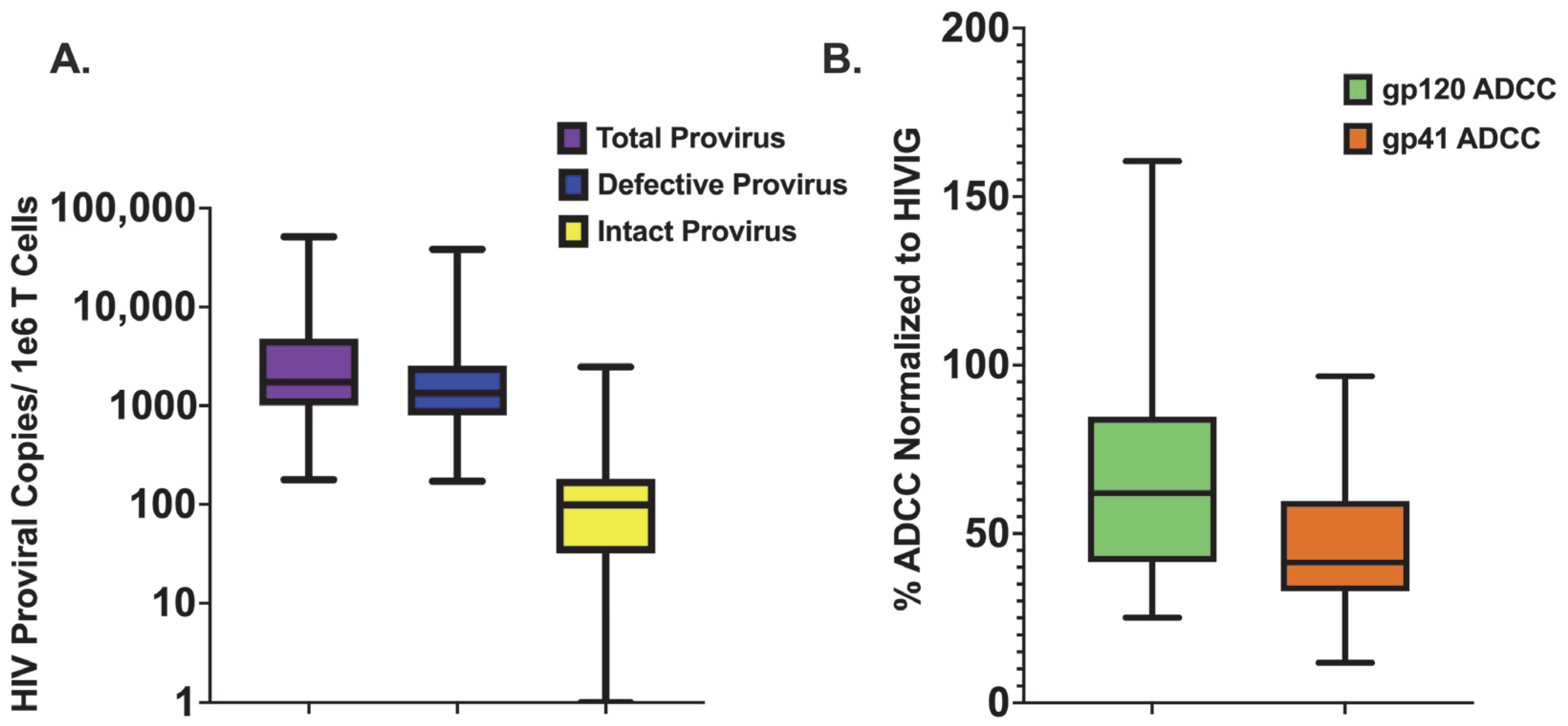
3.2. Quantifying Persistent HIV Provirus
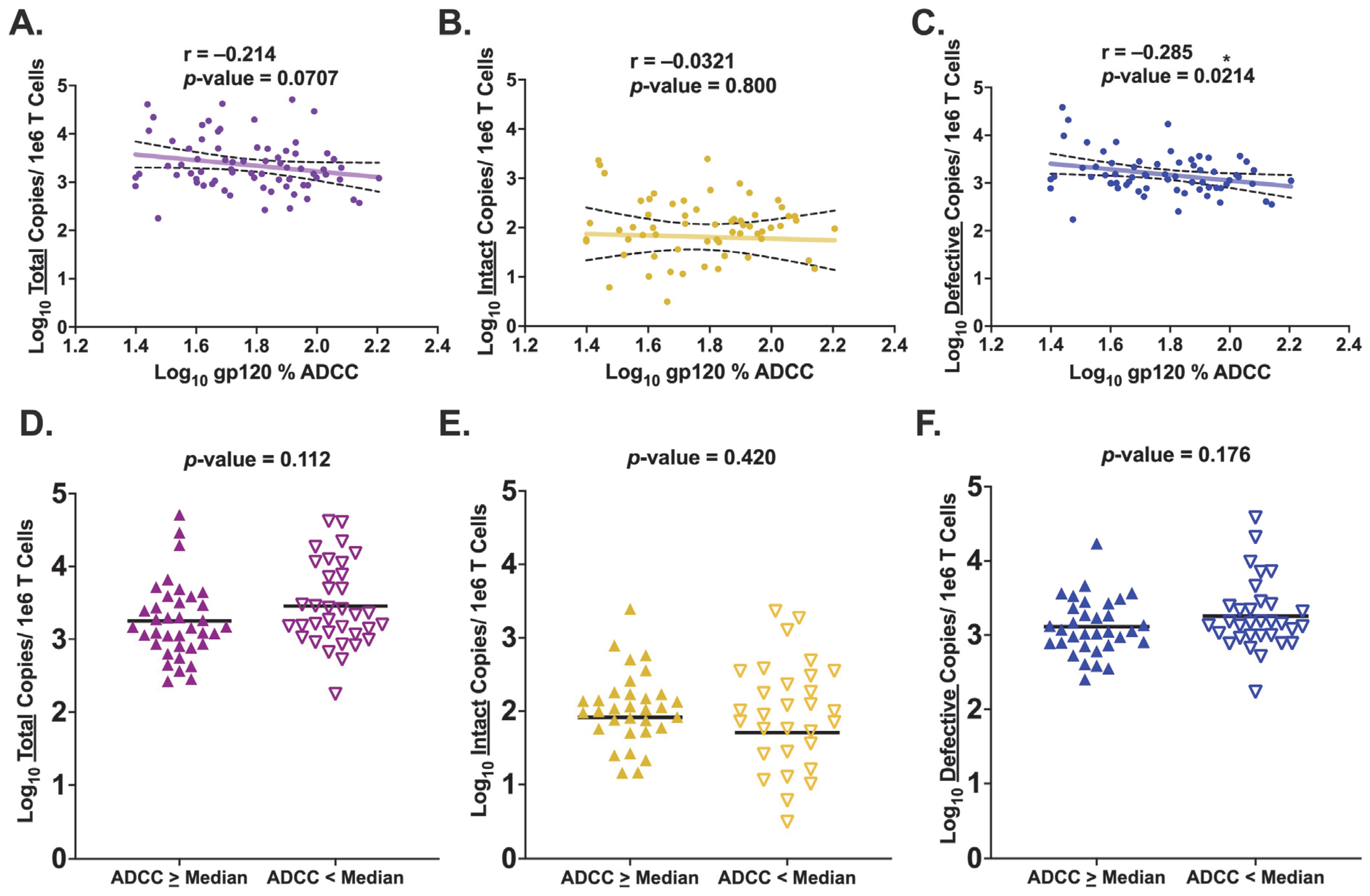
3.3. HIV Env gp120-Specific ADCC Activity Is Inversely Associated with Levels of Defective Provirus
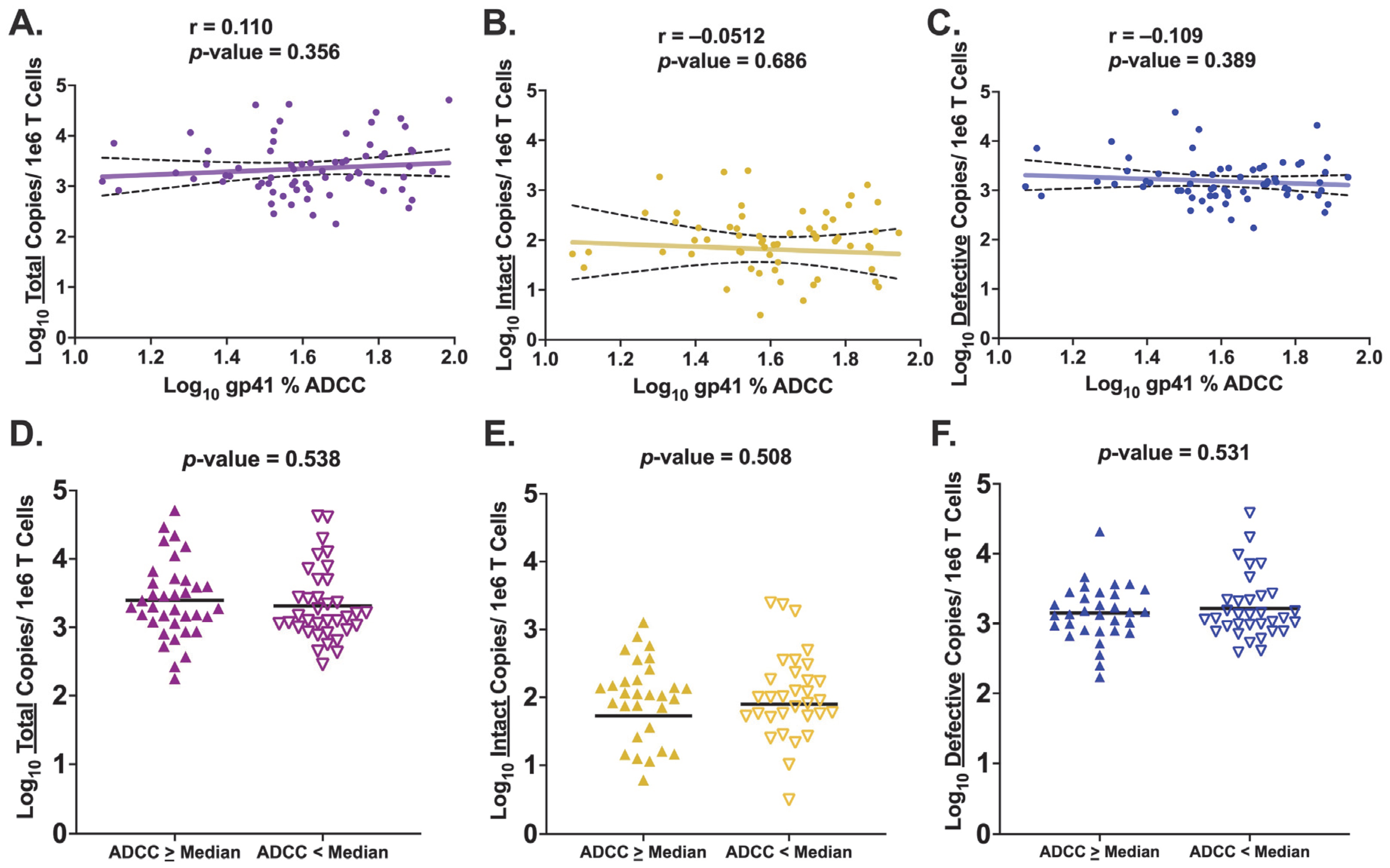
3.4. HIV Env gp41-Specific ADCC Activity Is Not Associated with Levels of Persistent HIV Provirus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control (CDC). Pneumocystis pneumonia—Los Angeles. MMWR Morb. Mortal. Wkly. Rep. 1981, 30, 250–252. [Google Scholar]
- Frank, T.D.; Carter, A.; Jahagirdar, D.; Biehl, M.H.; Douwes-Schultz, D.; Larson, S.L.; Arora, M.; Dwyer-Lindgren, L.; Steuben, K.M.; Abbastabar, H.; et al. Global, regional, and national incidence, prevalence, and mortality of HIV, 1980–2017, and forecasts to 2030, for 195 countries and territories: A systematic analysis for the Global Burden of Diseases, Injuries, and Risk Factors Study 2017. Lancet HIV 2019, 6, e831–e859. [Google Scholar] [CrossRef] [PubMed]
- Samji, H.; Cescon, A.; Hogg, R.S.; Modur, S.P.; Althoff, K.N.; Buchacz, K.; Burchell, A.N.; Cohen, M.; Gebo, K.A.; Gill, M.J.; et al. Closing the Gap: Increases in Life Expectancy among Treated HIV-Positive Individuals in the United States and Canada. PLoS ONE 2013, 8, e81355. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.-W.; Carruth, L.; Finzi, D.; Shen, X.; DiGiuseppe, J.A.; Taylor, H.; Hermankova, M.; Chadwick, K.; Margolick, J.; Quinn, T.C.; et al. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nature 1997, 387, 183–188. [Google Scholar] [CrossRef]
- Chun, T.-W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.M.; Baseler, M.; Lloyd, A.L.; Nowak, M.A.; Fauci, A.S. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 13193–13197. [Google Scholar] [CrossRef]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; et al. Identification of a Reservoir for HIV-1 in Patients on Highly Active Antiretroviral Therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef]
- Wong, J.K.; Hezareh, M.; Günthard, H.F.; Havlir, D.V.; Ignacio, C.C.; Spina, C.A.; Richman, D.D. Recovery of Replication-Competent HIV Despite Prolonged Suppression of Plasma Viremia. Science 1997, 278, 1291–1295. [Google Scholar] [CrossRef]
- Bruner, K.M.; Murray, A.J.; Pollack, R.A.; Soliman, M.G.; Laskey, S.B.; Capoferri, A.A.; Lai, J.; Strain, M.C.; Lada, S.M.; Hoh, R.; et al. Defective proviruses rapidly accumulate during acute HIV-1 infection. Nat. Med. 2016, 22, 1043–1049. [Google Scholar] [CrossRef]
- Ho, Y.-C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-Competent Noninduced Proviruses in the Latent Reservoir Increase Barrier to HIV-1 Cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef]
- Imamichi, H.; Dewar, R.L.; Adelsberger, J.W.; Rehm, C.A.; O’Doherty, U.; Paxinos, E.E.; Fauci, A.S.; Lane, H.C. Defective HIV-1 proviruses produce novel protein-coding RNA species in HIV-infected patients on combination antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2016, 113, 8783–8788. [Google Scholar] [CrossRef]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M.; Perrin, L.; Trono, D. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature 2003, 424, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Piantadosi, A.; Humes, D.; Chohan, B.; McClelland, R.S.; Overbaugh, J. Analysis of the Percentage of Human Immunodeficiency Virus Type 1 Sequences That Are Hypermutated and Markers of Disease Progression in a Longitudinal Cohort, Including One Individual with a Partially Defective Vif. J. Virol. 2009, 83, 7805–7814. [Google Scholar] [CrossRef] [PubMed]
- Davey, R.T.; Bhat, N.; Yoder, C.; Chun, T.-W.; Metcalf, J.A.; Dewar, R.; Natarajan, V.; Lempicki, R.A.; Adelsberger, J.W.; Miller, K.D.; et al. HIV-1 and T cell dynamics after interruption of highly active antiretroviral therapy (HAART) in patients with a history of sustained viral suppression. Proc. Natl. Acad. Sci. USA 1999, 96, 15109–15114. [Google Scholar] [CrossRef]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef]
- Joos, B.; Fischer, M.; Kuster, H.; Pillai, S.K.; Wong, J.K.; Böni, J.; Hirschel, B.; Weber, R.; Trkola, A.; Günthard, H.F.; et al. HIV rebounds from latently infected cells, rather than from continuing low-level replication. Proc. Natl. Acad. Sci. USA 2008, 105, 16725–16730. [Google Scholar] [CrossRef]
- French, M.A.; King, M.S.; Tschampa, J.M.; da Silva, B.A.; Landay, A.L. Serum Immune Activation Markers Are Persistently Increased in Patients with HIV Infection after 6 Years of Antiretroviral Therapy despite Suppression of Viral Replication and Reconstitution of CD4+T Cells. J. Infect. Dis. 2009, 200, 1212–1215. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Jacobs, D.R., Jr.; Baker, J.V.; Calmy, A.; Duprez, D.; La Rosa, A.; Kuller, L.H.; Pett, S.L.; Ristola, M.; Ross, M.J.; et al. Markers of Inflammation, Coagulation, and Renal Function Are Elevated in Adults with HIV Infection. J. Infect. Dis. 2010, 201, 1788–1795. [Google Scholar] [CrossRef]
- Turner, C.T.; Brown, J.; Shaw, E.; Uddin, I.; Tsaliki, E.; Roe, J.K.; Pollara, G.; Sun, Y.; Heather, J.M.; Lipman, M.; et al. Persistent T Cell Repertoire Perturbation and T Cell Activation in HIV After Long Term Treatment. Front. Immunol. 2021, 12, 634489. [Google Scholar] [CrossRef]
- Freiberg, M.S.; Chang, C.C.H.; Kuller, L.H.; Skanderson, M.; Lowy, E.; Kraemer, K.L.; Butt, A.A.; Goetz, M.B.; Leaf, D.; Oursler, K.A.; et al. HIV Infection and the Risk of Acute Myocardial Infarction. JAMA Intern. Med. 2013, 173, 614–622. [Google Scholar] [CrossRef]
- van Zoest, R.A.; Born, B.-J.H.v.D.; Reiss, P. Hypertension in people living with HIV. Curr. Opin. HIV AIDS 2017, 12, 513–522. [Google Scholar] [CrossRef]
- Spudich, S.; Robertson, K.R.; Bosch, R.J.; Gandhi, R.T.; Cyktor, J.C.; Mar, H.; Macatangay, B.J.; Lalama, C.M.; Rinaldo, C.; Collier, A.C.; et al. Persistent HIV-infected cells in cerebrospinal fluid are associated with poorer neurocognitive performance. J. Clin. Investig. 2019, 129, 3339–3346. [Google Scholar] [CrossRef]
- Nansseu, J.R.; Bigna, J.J.; Kaze, A.D.; Noubiap, J.J. Incidence and Risk Factors for Prediabetes and Diabetes Mellitus Among HIV-infected Adults on Antiretroviral Therapy: A Systematic Review and Meta-analysis. Epidemiology 2018, 29, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Grulich, A.E.; van Leeuwen, M.T.; Falster, M.O.; Vajdic, C.M. Incidence of cancers in people with HIV/AIDS compared with immunosuppressed transplant recipients: A meta-analysis. Lancet 2007, 370, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, M.J.; Chao, C.; Leyden, W.A.; Xu, L.; Tang, B.; Horberg, M.A.; Klein, D.; Quesenberry, C.P., Jr.; Towner, W.J.; Abrams, D.I. HIV infection and the risk of cancers with and without a known infectious cause. AIDS 2009, 23, 2337. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Phillips, A.N. HIV infection, antiretroviral treatment, ageing, and non-AIDS related morbidity. BMJ 2009, 338, a3172. [Google Scholar] [CrossRef]
- Desquilbet, L.; Jacobson, L.P.; Fried, L.P.; Phair, J.P.; Jamieson, B.D.; Holloway, M.; Margolick, J.B. HIV-1 Infection Is Associated with an Earlier Occurrence of a Phenotype Related to Frailty. J. Gerontol. Ser. A 2007, 62, 1279–1286. [Google Scholar] [CrossRef]
- Horvath, S.; Levine, A.J. HIV-1 Infection Accelerates Age According to the Epigenetic Clock. J. Infect. Dis. 2015, 212, 1563–1573. [Google Scholar] [CrossRef]
- Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; Neuhaus, J.; Nixon, D.; et al. Inflammatory and Coagulation Biomarkers and Mortality in Patients with HIV Infection. PLOS Med. 2008, 5, e203. [Google Scholar] [CrossRef]
- Imamichi, H.; Smith, M.; Adelsberger, J.W.; Izumi, T.; Scrimieri, F.; Sherman, B.T.; Rehm, C.A.; Imamichi, T.; Pau, A.; Catalfamo, M.; et al. Defective HIV-1 proviruses produce viral proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 3704–3710. [Google Scholar] [CrossRef]
- Kuniholm, J.; Coote, C.; Henderson, A.J. Defective HIV-1 genomes and their potential impact on HIV pathogenesis. Retrovirology 2022, 19, 13. [Google Scholar] [CrossRef]
- Pollack, R.A.; Jones, R.B.; Pertea, M.; Bruner, K.M.; Martin, A.R.; Thomas, A.; Capoferri, A.A.; Beg, S.A.; Huang, S.-H.; Karandish, S.; et al. Defective HIV-1 Proviruses Are Expressed and Can Be Recognized by Cytotoxic T Lymphocytes, which Shape the Proviral Landscape. Cell Host Microbe 2017, 21, 494–506.e4. [Google Scholar] [CrossRef]
- Olson, A.; Coote, C.; Snyder-Cappione, J.E.; Lin, N.; Sagar, M. HIV-1 Transcription but Not Intact Provirus Levels are Associated with Systemic Inflammation. J. Infect. Dis. 2021, 223, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Dhummakupt, A.; Khetan, P.; Nilles, T.; Zhou, W.; Mudvari, P.; Szewczyk, J.; Chen, Y.H.; Boritz, E.; Ji, H.; et al. Immune activation and exhaustion marker expression on T-cell subsets in ART-treated adolescents and young adults with perinatal HIV-1 infection as correlates of viral persistence. Front. Immunol. 2023, 14, 1007626. [Google Scholar] [CrossRef] [PubMed]
- Elahi, S.; Weiss, R.H.; Merani, S. Atorvastatin restricts HIV replication in CD4+ T cells by upregulation of p21. AIDS 2016, 30, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Dalzini, A.; Ballin, G.; Dominguez-Rodriguez, S.; Rojo, P.; Petrara, M.R.; Foster, C.; Cotugno, N.; Ruggiero, A.; Nastouli, E.; Klein, N.; et al. Size of HIV-1 reservoir is associated with telomere shortening and immunosenescence in early-treated European children with perinatally acquired HIV-1. J. Int. AIDS Soc. 2021, 24, e25847. [Google Scholar] [CrossRef]
- Hatano, H.; Jain, V.; Hunt, P.W.; Lee, T.-H.; Sinclair, E.; Do, T.D.; Hoh, R.; Martin, J.N.; McCune, J.M.; Hecht, F.; et al. Cell-Based Measures of Viral Persistence Are Associated with Immune Activation and Programmed Cell Death Protein 1 (PD-1)–Expressing CD4+ T cells. J. Infect. Dis. 2013, 208, 50–56. [Google Scholar] [CrossRef]
- Conway, J.M.; Perelson, A.S. Post-treatment control of HIV infection. Proc. Natl. Acad. Sci. USA 2015, 112, 5467–5472. [Google Scholar] [CrossRef]
- Goujard, C.; Girault, I.; Rouzioux, C.; Lécuroux, C.; Deveau, C.; Chaix, M.-L.; Jacomet, C.; Talamali, A.; Delfraissy, J.-F.; Venet, A.; et al. Hiv-1 Control after Transient Antiretroviral Treatment Initiated in Primary Infection: Role of Patient Characteristics and Effect of Therapy. Antivir. Ther. 2012, 17, 1001–1009. [Google Scholar] [CrossRef]
- Kuhn, L.; Paximadis, M.; Dias, B.D.C.; Loubser, S.; Strehlau, R.; Patel, F.; Shiau, S.; Coovadia, A.; Abrams, E.J.; Tiemessen, C.T. Age at antiretroviral therapy initiation and cell-associated HIV-1 DNA levels in HIV-1-infected children. PLoS ONE 2018, 13, e0195514. [Google Scholar] [CrossRef]
- Williams, J.P.; Hurst, J.; Stöhr, W.; Robinson, N.; Brown, H.; Fisher, M.; Kinloch, S.; Cooper, D.; Schechter, M.; Tambussi, G.; et al. HIV-1 DNA predicts disease progression and post-treatment virological control. eLife 2014, 3, e03821. [Google Scholar] [CrossRef]
- Bachmann, N.; von Siebenthal, C.; Vongrad, V.; Turk, T.; Neumann, K.; Beerenwinkel, N.; Bogojeska, J.; Roth, V.; Kok, Y.L.; Thorball, C.W.; et al. Determinants of HIV-1 reservoir size and long-term dynamics during suppressive ART. Nat. Commun. 2019, 10, 3193. [Google Scholar] [CrossRef] [PubMed]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.; Siliciano, R.F. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat. Med. 2003, 9, 727–728. [Google Scholar] [CrossRef]
- Kreider, E.F.; Bar, K.J. HIV-1 Reservoir Persistence and Decay: Implications for Cure Strategies. Curr. HIV/AIDS Rep. 2022, 19, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Frange, P.; Faye, A.; Avettand-Fenoël, V.; Bellaton, E.; Descamps, D.; Angin, M.; David, A.; Caillat-Zucman, S.; Peytavin, G.; Dollfus, C.; et al. HIV-1 virological remission lasting more than 12 years after interruption of early antiretroviral therapy in a perinatally infected teenager enrolled in the French ANRS EPF-CO10 paediatric cohort: A case report. Lancet HIV 2016, 3, e49–e54. [Google Scholar] [CrossRef] [PubMed]
- Luzuriaga, K. Early Combination Antiretroviral Therapy Limits HIV-1 Persistence in Children. Annu. Rev. Med. 2016, 67, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Persaud, D.; Gay, H.; Ziemniak, C.; Chen, Y.H.; Piatak, M.; Chun, T.-W.; Strain, M.; Richman, D.; Luzuriaga, K. Absence of Detectable HIV-1 Viremia after Treatment Cessation in an Infant. New Engl. J. Med. 2013, 369, 1828–1835. [Google Scholar] [CrossRef]
- Violari, A.; Cotton, M.F.; Kuhn, L.; Schramm, D.B.; Paximadis, M.; Loubser, S.; Shalekoff, S.; Dias, B.D.C.; Otwombe, K.; Liberty, A.; et al. A child with perinatal HIV infection and long-term sustained virological control following antiretroviral treatment cessation. Nat. Commun. 2019, 10, 412. [Google Scholar] [CrossRef]
- Rainwater-Lovett, K.; Ziemniak, C.; Watson, D.; Luzuriaga, K.; Siberry, G.; Petru, A.; Chen, Y.; Uprety, P.; McManus, M.; Ho, Y.-C.; et al. Paucity of Intact Non-Induced Provirus with Early, Long-Term Antiretroviral Therapy of Perinatal HIV Infection. PLoS ONE 2017, 12, e0170548. [Google Scholar] [CrossRef]
- Uprety, P.; Patel, K.; Karalius, B.; Ziemniak, C.; Chen, Y.H.; Brummel, S.S.; Siminski, S.; Van Dyke, R.B.; Seage, G.R.; Persaud, D.; et al. Human Immunodeficiency Virus Type 1 DNA Decay Dynamics with Early, Long-term Virologic Control of Perinatal Infection. Clin. Infect. Dis. 2017, 64, 1471–1478. [Google Scholar] [CrossRef]
- The Collaborative Initiative for Paediatric HIV Education and Research (CIPHER) Global Cohort Collaboration; Slogrove, A.L.; Schomaker, M.; Davies, M.-A.; Williams, P.; Balkan, S.; Ben-Farhat, J.; Calles, N.; Chokephaibulkit, K.; Duff, C.; et al. The epidemiology of adolescents living with perinatally acquired HIV: A cross-region global cohort analysis. PLOS Med. 2018, 15, e1002514. [Google Scholar] [CrossRef]
- Persaud, D.; Pierson, T.; Ruff, C.; Finzi, D.; Chadwick, K.R.; Margolick, J.B.; Ruff, A.; Hutton, N.; Ray, S.; Siliciano, R.F. A stable latent reservoir for HIV-1 in resting CD4+ T lymphocytes in infected children. J. Clin. Investig. 2000, 105, 995–1003. [Google Scholar] [CrossRef]
- Barlow-Mosha, L.; Eckard, A.R.; McComsey, G.A.; Musoke, P.M. Metabolic complications and treatment of perinatally HIV-infected children and adolescents. J. Int. AIDS Soc. 2013, 16, 18600. [Google Scholar] [CrossRef] [PubMed]
- Cotugno, N.; Douagi, I.; Rossi, P.; Palma, P. Suboptimal Immune Reconstitution in Vertically HIV Infected Children: A View on How HIV Replication and Timing of HAART Initiation Can Impact on T and B-cell Compartment. Clin. Dev. Immunol. 2012, 2012 (Suppl. 6), 805151. [Google Scholar] [CrossRef] [PubMed]
- Vreeman, R.C.; Scanlon, M.L.; McHenry, M.S.; Nyandiko, W.M. The physical and psychological effects of HIV infection and its treatment on perinatally HIV-infected children. J. Int. AIDS Soc. 2015, 18, 20258. [Google Scholar] [CrossRef] [PubMed]
- Flynn, P.M.; Abrams, E.J. Growing up with perinatal HIV. AIDS 2019, 33, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D.; on behalf of the WHO–UNAIDS Network for HIV Isolation Characterisation. Global and regional molecular epidemiology of HIV-1, 1990–2015: A systematic review, global survey, and trend analysis. Lancet Infect. Dis. 2019, 19, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Ndung’u, T.; McCune, J.M.; Deeks, S.G. Why and where an HIV cure is needed and how it might be achieved. Nature 2019, 576, 397–405. [Google Scholar] [CrossRef]
- Obimbo, E.M.M.; Mbori-Ngacha, D.A.; Ochieng, J.O.B.; Richardson, B.A.; Otieno, P.A.M.; Bosire, R.M.; Farquhar, C.; Overbaugh, J.; John-Stewart, G.C. Predictors of Early Mortality in a Cohort of Human Immunodeficiency Virus Type 1-Infected African Children. Pediatr. Infect. Dis. J. 2004, 23, 536–543. [Google Scholar] [CrossRef]
- Obimbo, E.M.M.; Wamalwa, D.M.; Richardson, B.; Mbori-Ngacha, D.M.; Overbaugh, J.; Emery, S.B.; Otieno, P.M.; Farquhar, C.; Bosire, R.M.; Payne, B.L.; et al. Pediatric HIV-1 in Kenya: Pattern and Correlates of Viral Load and Association with Mortality. J. Acquir. Immune Defic. Syndr. 2009, 51, 209–215. [Google Scholar] [CrossRef]
- Abrahams, M.-R.; Joseph, S.B.; Garrett, N.; Tyers, L.; Moeser, M.; Archin, N.; Council, O.D.; Matten, D.; Zhou, S.; Doolabh, D.; et al. The replication-competent HIV-1 latent reservoir is primarily established near the time of therapy initiation. Sci. Transl. Med. 2019, 11, eaaw5589. [Google Scholar] [CrossRef]
- Brodin, J.; Zanini, F.; Thebo, L.; Lanz, C.; Bratt, G.; Neher, R.A.; Albert, J. Establishment and stability of the latent HIV-1 DNA reservoir. eLife 2016, 5, e18889. [Google Scholar] [CrossRef] [PubMed]
- Pankau, M.D.; Reeves, D.B.; Harkins, E.; Ronen, K.; Jaoko, W.; Mandaliya, K.; Graham, S.M.; McClelland, R.S.; Iv, F.A.M.; Schiffer, J.T.; et al. Dynamics of HIV DNA reservoir seeding in a cohort of superinfected Kenyan women. PLOS Pathog. 2020, 16, e1008286. [Google Scholar] [CrossRef] [PubMed]
- Borrow, P.; Lewicki, H.; Wei, X.; Horwitz, M.S.; Peffer, N.; Meyers, H.; Nelson, J.A.; Gairin, J.E.; Hahn, B.H.; Oldstone, M.B.; et al. Antiviral pressure exerted by HIV-l-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med. 1997, 3, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.; Pertea, M.; Rongvaux, A.; Wang, L.; Durand, C.M.; Ghiaur, G.; Lai, J.; McHugh, H.L.; Hao, H.; Zhang, H.; et al. Broad CTL response is required to clear latent HIV-1 due to dominance of escape mutations. Nature 2015, 517, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Ren, Y.; Thomas, A.S.; Chan, D.; Mueller, S.; Ward, A.R.; Patel, S.; Bollard, C.M.; Cruz, C.R.; Karandish, S.; et al. Latent HIV reservoirs exhibit inherent resistance to elimination by CD8+ T cells. J. Clin. Investig. 2018, 128, 876–889. [Google Scholar] [CrossRef]
- Statzu, M.; Jin, W.; Fray, E.J.; Wong, A.K.H.; Kumar, M.R.; Ferrer, E.; Docken, S.S.; Pinkevych, M.; McBrien, J.B.; Fennessey, C.M.; et al. CD8+ lymphocytes do not impact SIV reservoir establishment under ART. Nat. Microbiol. 2023, 8, 299–308. [Google Scholar] [CrossRef]
- Halper-Stromberg, A.; Lu, C.-L.; Klein, F.; Horwitz, J.A.; Bournazos, S.; Nogueira, L.; Eisenreich, T.R.; Liu, C.; Gazumyan, A.; Schaefer, U.; et al. Broadly Neutralizing Antibodies and Viral Inducers Decrease Rebound from HIV-1 Latent Reservoirs in Humanized Mice. Cell 2014, 158, 989–999. [Google Scholar] [CrossRef]
- Kim, J.T.; Zhang, T.-H.; Carmona, C.; Lee, B.; Seet, C.S.; Kostelny, M.; Shah, N.; Chen, H.; Farrell, K.; Soliman, M.S.A.; et al. Latency reversal plus natural killer cells diminish HIV reservoir in vivo. Nat. Commun. 2022, 13, 121. [Google Scholar] [CrossRef]
- Barouch, D.H.; Whitney, J.B.; Moldt, B.; Klein, F.; Oliveira, T.Y.; Liu, J.; Stephenson, K.E.; Chang, H.-W.; Shekhar, K.; Gupta, S.; et al. Therapeutic efficacy of potent neutralizing HIV-1-specific monoclonal antibodies in SHIV-infected rhesus monkeys. Nature 2013, 503, 224–228. [Google Scholar] [CrossRef]
- Borducchi, E.N.; Liu, J.; Nkolola, J.P.; Cadena, A.M.; Yu, W.-H.; Fischinger, S.; Broge, T.; Abbink, P.; Mercado, N.B.; Chandrashekar, A.; et al. Antibody and TLR7 agonist delay viral rebound in SHIV-infected monkeys. Nature 2018, 563, 360–364. [Google Scholar] [CrossRef]
- Bar, K.J.; Sneller, M.C.; Harrison, L.J.; Justement, J.S.; Overton, E.T.; Petrone, M.E.; Salantes, D.B.; Seamon, C.A.; Scheinfeld, B.; Kwan, R.W.; et al. Effect of HIV Antibody VRC01 on Viral Rebound after Treatment Interruption. N. Engl. J. Med. 2016, 375, 2037–2050. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Nogueira, L.; Stoffel, E.; Oliveira, T.Y.; Breton, G.; Millard, K.G.; Turroja, M.; Butler, A.; Ramos, V.; Seaman, M.S.; et al. Prolonged viral suppression with anti-HIV-1 antibody therapy. Nature 2022, 606, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-L.; Murakowski, D.K.; Bournazos, S.; Schoofs, T.; Sarkar, D.; Halper-Stromberg, A.; Horwitz, J.A.; Nogueira, L.; Golijanin, J.; Gazumyan, A.; et al. Enhanced clearance of HIV-1–infected cells by broadly neutralizing antibodies against HIV-1 in vivo. Science 2016, 352, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, P.; Gruell, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.Y.; Lorenzi, J.C.C.; et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature 2018, 561, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Scheid, J.F.; Horwitz, J.A.; Bar-On, Y.; Kreider, E.F.; Lu, C.L.; Lorenzi, J.C.; Feldmann, A.; Braunschweig, M.; Nogueira, L.; Oliveira, T.; et al. Faculty Opinions recommendation of HIV-1 antibody 3BNC117 suppresses viral rebound in humans during treatment interruption. Nature 2016, 535, 556–560. [Google Scholar] [CrossRef]
- Bertagnolli, L.N.; Varriale, J.; Sweet, S.; Brockhurst, J.; Simonetti, F.R.; White, J.; Beg, S.; Lynn, K.; Mounzer, K.; Frank, I.; et al. Autologous IgG antibodies block outgrowth of a substantial but variable fraction of viruses in the latent reservoir for HIV-1. Proc. Natl. Acad. Sci. USA 2020, 117, 32066–32077. [Google Scholar] [CrossRef]
- Dhande, J.; Angadi, M.; Murugavel, K.; Poongulali, S.; Nandagopal, P.; Vignesh, R.; Ghate, M.; Kulkarni, S.; Thakar, M. Brief Report: The Anti–HIV-1 ADCC-Mediating Antibodies from Cervicovaginal Secretions of HIV-Infected Women Have an Ability to Mediate Lysing of Autologous CD4+ HIV-Infected Cells. J. Acquir. Immune Defic. Syndr. 2018, 79, 277–282. [Google Scholar] [CrossRef]
- Lee, W.S.; Prévost, J.; Richard, J.; van der Sluis, R.M.; Lewin, S.R.; Pazgier, M.; Finzi, A.; Parsons, M.S.; Kent, S.J. CD4- and Time-Dependent Susceptibility of HIV-1-Infected Cells to Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2019, 93, 10–1128. [Google Scholar] [CrossRef]
- Lee, W.S.; Richard, J.; Lichtfuss, M.; Smith, A.B., III; Park, J.; Courter, J.R.; Melillo, B.N.; Sodroski, J.G.; Kaufmann, D.E.; Finzi, A.; et al. Antibody-Dependent Cellular Cytotoxicity against Reactivated HIV-1-Infected Cells. J. Virol. 2016, 90, 2021–2030. [Google Scholar] [CrossRef]
- Gray, G.E.; Huang, Y.; Grunenberg, N.; Laher, F.; Roux, S.; Andersen-Nissen, E.; De Rosa, S.C.; Flach, B.; Randhawa, A.K.; Jensen, R.; et al. Immune correlates of the Thai RV144 HIV vaccine regimen in South Africa. Sci. Transl. Med. 2019, 11, eaax1880. [Google Scholar] [CrossRef]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-Correlates Analysis of an HIV-1 Vaccine Efficacy Trial. N. Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Mabuka, J.; Nduati, R.; Odem-Davis, K.; Peterson, D.; Overbaugh, J. HIV-Specific Antibodies Capable of ADCC Are Common in Breastmilk and Are Associated with Reduced Risk of Transmission in Women with High Viral Loads. PLOS Pathog. 2012, 8, e1002739. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.S.; Moreau, Y.; Jiang, W.; Isaac, J.E.; Ewing, A.; White, L.F.; Kourtis, A.P.; Sagar, M. Pre-existing infant antibody-dependent cellular cytotoxicity associates with reduced HIV-1 acquisition and lower morbidity. Cell Rep. Med. 2021, 2, 100412. [Google Scholar] [CrossRef] [PubMed]
- Milligan, C.; Richardson, B.A.; John-Stewart, G.; Nduati, R.; Overbaugh, J. Passively Acquired Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity in HIV-Infected Infants Is Associated with Reduced Mortality. Cell Host Microbe 2015, 17, 500–506. [Google Scholar] [CrossRef]
- Yaffe, Z.A.; Naiman, N.E.; Slyker, J.; Wines, B.D.; Richardson, B.A.; Hogarth, P.M.; Bosire, R.; Farquhar, C.; Ngacha, D.M.; Nduati, R.; et al. Improved HIV-positive infant survival is correlated with high levels of HIV-specific ADCC activity in multiple cohorts. Cell Rep. Med. 2021, 2, 100254. [Google Scholar] [CrossRef]
- Ljunggren, K.; Moschese, V.; Broliden, P.-A.; Giaquinto, C.; Quinti, I.; Fenyö, E.-M.; Wahren, B.; Rossi, P.; Jondal, M. Antibodies Mediating Cellular Cytotoxicity and Neutralization Correlate with a Better Clinical Stage in Children Born to Human Immunodeficiency Virus-Infected Mothers. J. Infect. Dis. 1990, 161, 198–202. [Google Scholar] [CrossRef]
- Tranchat, C.; Van de Perre, P.; Simonon-Sorel, A.; Karita, E.; Benchaïb, M.; Lepage, P.; Desgranges, C.; Boyer, V.; Répo, C. Maternal humoral factors associated with perinatal human immunodeficiency virus type-1 transmission in a cohort from Kigali, Rwanda, 1988–1994. J. Infect. 1999, 39, 213–220. [Google Scholar] [CrossRef]
- Kant, S.; Zhang, N.; Barbé, A.; Routy, J.-P.; Tremblay, C.; Thomas, R.; Szabo, J.; Côté, P.; Trottier, B.; LeBlanc, R.; et al. Polyfunctional Fc Dependent Activity of Antibodies to Native Trimeric Envelope in HIV Elite Controllers. Front. Immunol. 2020, 11, 583820. [Google Scholar] [CrossRef]
- Madhavi, V.; Wines, B.D.; Amin, J.; Emery, S.; ENCORE1 Study Group; Lopez, E.; Kelleher, A.; Sydney LTNP Study Group; Center, R.J.; Hogarth, P.M.; et al. HIV-1 Env- and Vpu-Specific Antibody-Dependent Cellular Cytotoxicity Responses Associated with Elite Control of HIV. J. Virol. 2017, 91, 10–1128. [Google Scholar] [CrossRef]
- Lambotte, O.; Ferrari, G.; Moog, C.; Yates, N.L.; Liao, H.-X.; Parks, R.J.; Hicks, C.B.; Owzar, K.; Tomaras, G.D.; Montefiori, D.C.; et al. Heterogeneous neutralizing antibody and antibody-dependent cell cytotoxicity responses in HIV-1 elite controllers. AIDS 2009, 23, 897–906. [Google Scholar] [CrossRef]
- Wren, L.H.; Chung, A.W.; Isitman, G.; Kelleher, A.D.; Parsons, M.S.; Amin, J.; Cooper, D.A.; Stratov, I.; Navis, M.; Kent, S.J.; et al. Specific antibody-dependent cellular cytotoxicity responses associated with slow progression of HIV infection. Immunology 2013, 138, 116–123. [Google Scholar] [CrossRef]
- Wamalwa, D.C.; Farquhar, C.; Obimbo, E.M.; Selig, S.; Mbori-Ngacha, D.A.; A Richardson, B.; Overbaugh, J.; Egondi, T.; Inwani, I.; John-Stewart, G. Medication diaries do not improve outcomes with highly active antiretroviral therapy in Kenyan children: A randomized clinical trial. J. Int. AIDS Soc. 2009, 12, 8. [Google Scholar] [CrossRef]
- Gómez-Román, V.R.; Florese, R.H.; Patterson, L.J.; Peng, B.; Venzon, D.; Aldrich, K.; Robert-Guroff, M. A simplified method for the rapid fluorometric assessment of antibody-dependent cell-mediated cytotoxicity. J. Immunol. Methods 2006, 308, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, N.A.; Fish, C.S.; Levy, C.N.; Roychoudhury, P.; Reeves, D.B.; Hughes, S.M.; Schiffer, J.T.; Benki-Nugent, S.; John-Stewart, G.; Wamalwa, D.; et al. HIV reservoir quantification using cross-subtype multiplex ddPCR. iScience 2022, 25, 103615. [Google Scholar] [CrossRef] [PubMed]
- Wamalwa, D.; Benki-Nugent, S.; Langat, A.; Tapia, K.; Ngugi, E.; Moraa, H.; Maleche-Obimbo, E.; Otieno, V.; Inwani, I.; Richardson, B.A.; et al. Treatment interruption after 2-year antiretroviral treatment initiated during acute/early HIV in infancy. AIDS 2016, 30, 2303–2313. [Google Scholar] [CrossRef]
- Neilson, J.R.; John, G.C.; Carr, J.K.; Lewis, P.; Kreiss, J.K.; Jackson, S.; Nduati, R.W.; Mbori-Ngacha, D.; Panteleeff, D.D.; Bodrug, S.; et al. Subtypes of Human Immunodeficiency Virus Type 1 and Disease Stage among Women in Nairobi, Kenya. J. Virol. 1999, 73, 4393–4403. [Google Scholar] [CrossRef]
- Fish, C.S.; Cassidy, N.A.; Levy, C.N.; Hughes, S.M.; Jerome, K.R.; Overbaugh, J.; Hladik, F.; Lehman, D.A. Protocol for high-throughput reservoir quantification across global HIV subtypes using a cross-subtype intact proviral DNA assay. STAR Protoc. 2022, 3, 101681. [Google Scholar] [CrossRef]
- Levy, C.N.; Hughes, S.M.; Roychoudhury, P.; Reeves, D.B.; Amstuz, C.; Zhu, H.; Huang, M.-L.; Wei, Y.; Bull, M.E.; Cassidy, N.A.; et al. A highly multiplexed droplet digital PCR assay to measure the intact HIV-1 proviral reservoir. Cell Rep. Med. 2021, 2, 100243. [Google Scholar] [CrossRef] [PubMed]
- Schatorjé, E.J.H.; Gemen, E.F.A.; Driessen, G.J.A.; Leuvenink, J.; van Hout, R.W.N.M.; de Vries, E. Paediatric Reference Values for the Peripheral T cell Compartment. Scand. J. Immunol. 2012, 75, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Shearer, W.T.; Rosenblatt, H.M.; Gelman, R.S.; Oyomopito, R.; Plaeger, S.; Stiehm, E.; Wara, D.W.; Douglas, S.D.; Luzuriaga, K.; McFarland, E.J.; et al. Lymphocyte subsets in healthy children from birth through 18 years of age: The pediatric AIDS clinical trials group P1009 study. J. Allergy Clin. Immunol. 2003, 112, 973–980. [Google Scholar] [CrossRef]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.A.R.; Jenike, K.M.; et al. A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nature 2019, 566, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.T.; Cyktor, J.C.; Bosch, R.J.; Mar, H.; Laird, G.M.; Martin, A.; Collier, A.C.; Riddler, S.A.; Macatangay, B.J.; Rinaldo, C.R.; et al. Selective Decay of Intact HIV-1 Proviral DNA on Antiretroviral Therapy. J. Infect. Dis. 2021, 223, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Hartana, C.A.; Garcia-Broncano, P.; Rassadkina, Y.; Lian, X.; Jiang, C.; Einkauf, K.B.; Maswabi, K.; Ajibola, G.; Moyo, S.; Mohammed, T.; et al. Immune correlates of HIV-1 reservoir cell decline in early-treated infants. Cell Rep. 2022, 40, 111126. [Google Scholar] [CrossRef] [PubMed]
- Peluso, M.J.; Bacchetti, P.; Ritter, K.D.; Beg, S.; Lai, J.; Martin, J.N.; Hunt, P.W.; Henrich, T.J.; Siliciano, J.D.; Siliciano, R.F.; et al. Differential decay of intact and defective proviral DNA in HIV-1-infected individuals on suppressive antiretroviral therapy. JCI Insight 2020, 5, e132997. [Google Scholar] [CrossRef]
- White, J.A.; Simonetti, F.R.; Beg, S.; McMyn, N.F.; Dai, W.; Bachmann, N.; Lai, J.; Ford, W.C.; Bunch, C.; Jones, J.L.; et al. Complex decay dynamics of HIV virions, intact and defective proviruses, and 2LTR circles following initiation of antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2022, 119, e2120326119. [Google Scholar] [CrossRef]
- Ananworanich, J.; Puthanakit, T.; Suntarattiwong, P.; Chokephaibulkit, K.; Kerr, S.J.; Fromentin, R.; Bakeman, W.; Intasan, J.; Mahanontharit, A.; Sirivichayakul, S.; et al. Reduced markers of HIV persistence and restricted HIV-specific immune responses after early antiretroviral therapy in children. AIDS 2014, 28, 1015–1020. [Google Scholar] [CrossRef]
- Martínez-Bonet, M.; Puertas, M.C.; Fortuny, C.; Ouchi, D.; Mellado, M.J.; Rojo, P.; Noguera-Julian, A.; Muñoz-Fernández, M.A.; Martinez-Picado, J. Establishment and Replenishment of the Viral Reservoir in Perinatally HIV-1-infected Children Initiating Very Early Antiretroviral Therapy. Clin. Infect. Dis. 2015, 61, 1169–1178. [Google Scholar] [CrossRef]
- Moore, P.L.; Crooks, E.T.; Porter, L.; Zhu, P.; Cayanan, C.S.; Grise, H.; Corcoran, P.; Zwick, M.B.; Franti, M.; Morris, L.; et al. Nature of Nonfunctional Envelope Proteins on the Surface of Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 2515–2528. [Google Scholar] [CrossRef]
- Marchitto, L.; Benlarbi, M.; Prévost, J.; Laumaea, A.; Descôteaux-Dinelle, J.; Medjahed, H.; Bourassa, C.; Gendron-Lepage, G.; Kirchhoff, F.; Sauter, D.; et al. Impact of HIV-1 Vpu-mediated downregulation of CD48 on NK-cell-mediated antibody-dependent cellular cytotoxicity. Mbio 2023, 14, e0078923. [Google Scholar] [CrossRef]
- Veillette, M.; Désormeaux, A.; Medjahed, H.; Gharsallah, N.-E.; Coutu, M.; Baalwa, J.; Guan, Y.; Lewis, G.; Ferrari, G.; Hahn, B.; et al. Interaction with Cellular CD4 Exposes HIV-1 Envelope Epitopes Targeted by Antibody-Dependent Cell-Mediated Cytotoxicity. J. Virol. 2014, 88, 2633–2644. [Google Scholar] [CrossRef]
- Veillette, M.; Coutu, M.; Richard, J.; Batraville, L.-A.; Dagher, O.; Bernard, N.; Tremblay, C.; Kaufmann, D.E.; Roger, M.; Finzi, A. The HIV-1 gp120 CD4-Bound Conformation Is Preferentially Targeted by Antibody-Dependent Cellular Cytotoxicity-Mediating Antibodies in Sera from HIV-1-Infected Individuals. J. Virol. 2015, 89, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.N.; Lukhele, S.; Hajjar, F.; Routy, J.-P.; Cohen, É.A. HIV Nef and Vpu protect HIV-infected CD4+ T cells from antibody-mediated cell lysis through down-modulation of CD4 and BST2. Retrovirology 2014, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Prévost, J.; Richard, J.; Medjahed, H.; Alexander, A.; Jones, J.; Kappes, J.C.; Ochsenbauer, C.; Finzi, A. Incomplete Downregulation of CD4 Expression Affects HIV-1 Env Conformation and Antibody-Dependent Cellular Cytotoxicity Responses. J. Virol. 2018, 92, e00484-18. [Google Scholar] [CrossRef]
- Alsahafi, N.; Ding, S.; Richard, J.; Markle, T.; Brassard, N.; Walker, B.; Lewis, G.K.; Kaufmann, D.E.; Brockman, M.A.; Finzi, A. Nef Proteins from HIV-1 Elite Controllers Are Inefficient at Preventing Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2015, 90, 2993–3002. [Google Scholar] [CrossRef]
- Alsahafi, N.; Richard, J.; Prévost, J.; Coutu, M.; Brassard, N.; Parsons, M.S.; Kaufmann, D.E.; Brockman, M.; Finzi, A. Impaired Downregulation of NKG2D Ligands by Nef Proteins from Elite Controllers Sensitizes HIV-1-Infected Cells to Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2017, 91, 10–128. [Google Scholar] [CrossRef] [PubMed]
- Duette, G.; Hiener, B.; Morgan, H.; Mazur, F.G.; Mathivanan, V.; Horsburgh, B.A.; Fisher, K.; Tong, O.; Lee, E.; Ahn, H.; et al. The HIV-1 proviral landscape reveals that Nef contributes to HIV-1 persistence in effector memory CD4+ T cells. J. Clin. Investig. 2022, 132, e154422. [Google Scholar] [CrossRef]
- Pollara, J.; Bonsignori, M.; Moody, M.A.; Pazgier, M.; Haynes, B.F.; Ferrari, G. Epitope Specificity of Human Immunodeficiency Virus-1 Antibody Dependent Cellular Cytotoxicity [ADCC] Responses. Curr. HIV Res. 2013, 11, 378–387. [Google Scholar] [CrossRef]
- Williams, K.L.; Stumpf, M.; Naiman, N.E.; Ding, S.; Garrett, M.; Gobillot, T.; Vézina, D.; Dusenbury, K.; Ramadoss, N.S.; Basom, R.; et al. Identification of HIV gp41-specific antibodies that mediate killing of infected cells. PLOS Pathog. 2019, 15, e1007572. [Google Scholar] [CrossRef]
- Cook, J.D.; Khondker, A.; Lee, J.E. Conformational plasticity of the HIV-1 gp41 immunodominant region is recognized by multiple non-neutralizing antibodies. Commun. Biol. 2022, 5, 291. [Google Scholar] [CrossRef]
- Jin, S.; Ji, Y.; Wang, Q.; Wang, H.; Shi, X.; Han, X.; Zhou, T.; Shang, H.; Zhang, L. Spatiotemporal hierarchy in antibody recognition against transmitted HIV-1 envelope glycoprotein during natural infection. Retrovirology 2016, 13, 12. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-Cell Responses to Transmitted Human Immunodeficiency Virus Type 1: Virion-Binding Immunoglobulin M (IgM) and IgG Antibodies Followed by Plasma Anti-gp41 Antibodies with Ineffective Control of Initial Viremia. J. Virol. 2008, 82, 12449–12463. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R.; Mascola, J.R. Antibody responses to envelope glycoproteins in HIV-1 infection. Nat. Immunol. 2015, 16, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Steckbeck, J.D.; Craigo, J.K.; Barnes, C.O.; Montelaro, R.C. Highly Conserved Structural Properties of the C-terminal Tail of HIV-1 gp41 Protein Despite Substantial Sequence Variation among Diverse Clades: Implications for Functions in Viral Replication. J. Biol. Chem. 2011, 286, 27156–27166. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, R.; Sodroski, J. The HIV-1 Envelope Glycoproteins: Fusogens, Antigens, and Immunogens. Science 1998, 280, 1884–1888. [Google Scholar] [CrossRef] [PubMed]
- Llano, A.; Parera, M.; Lopez, M.; Oriol-Tordera, B.; Ruiz-Riol, M.; Coll, J.; Perez, F.; Leselbaum, A.R.; McGowan, I.; Sengupta, D.; et al. Safety, immunogenicity and effect on viral rebound of HTI vaccines in early treated HIV-1 infection: A randomized, placebo-controlled phase 1 trial. Nat. Med. 2022, 28, 2611–2621. [Google Scholar] [CrossRef]
- Landovitz, R.J.; Scott, H.; Deeks, S.G. Prevention, treatment and cure of HIV infection. Nat. Rev. Microbiol. 2023, 21, 657–670. [Google Scholar] [CrossRef]
- Newell, M.-L.; Coovadia, H.; Cortina-Borja, M.; Rollins, N.; Gaillard, P.; Dabis, F. Mortality of infected and uninfected infants born to HIV-infected mothers in Africa: A pooled analysis. Lancet 2004, 364, 1236–1243. [Google Scholar] [CrossRef]
- World Health Organization. Treatment and Care in Children and Adolescents. Available online: https://www.who.int/teams/global-hiv-hepatitis-and-stis-programmes/hiv/treatment/treatment-and-care-in-children-and-adolescents (accessed on 11 September 2023).
- Shelton, E.M.; Reeves, D.B.; Ignacio, R.A.B. Initiation of Antiretroviral Therapy during Primary HIV Infection: Effects on the Latent HIV Reservoir, Including on Analytic Treatment Interruptions. Aids Rev. 2020, 23, 28–39. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N = 72 | Median | [Min, Max] |
|---|---|---|
| Age at Enrollment | ||
| Months | 59 | 15.5, 152 |
| Years | 4.9 | 1.3, 12.7 |
| Viral Load (log10 c/mL) | 5.96 | 4.18, 6.96 |
| CD4 % | 6.3 | 0.70, 73.4 |
| CD4 Count (cells/mm3) | 354 | 15, 2009 |
| N | % | |
| Gender | ||
| Male | 33 | 46% |
| Female | 39 | 54% |
| ART Regimen | ||
| NNRTI, NRTI | 70 | 97.2% |
| NRTI | 1 | 1.4% |
| NNRTI, NRTI, Protease Inhibitor | 1 | 1.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yucha, R.; Litchford, M.L.; Fish, C.S.; Yaffe, Z.A.; Richardson, B.A.; Maleche-Obimbo, E.; John-Stewart, G.; Wamalwa, D.; Overbaugh, J.; Lehman, D.A. Higher HIV-1 Env gp120-Specific Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity Is Associated with Lower Levels of Defective HIV-1 Provirus. Viruses 2023, 15, 2055. https://doi.org/10.3390/v15102055
Yucha R, Litchford ML, Fish CS, Yaffe ZA, Richardson BA, Maleche-Obimbo E, John-Stewart G, Wamalwa D, Overbaugh J, Lehman DA. Higher HIV-1 Env gp120-Specific Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity Is Associated with Lower Levels of Defective HIV-1 Provirus. Viruses. 2023; 15(10):2055. https://doi.org/10.3390/v15102055
Chicago/Turabian StyleYucha, Ryan, Morgan L. Litchford, Carolyn S. Fish, Zak A. Yaffe, Barbra A. Richardson, Elizabeth Maleche-Obimbo, Grace John-Stewart, Dalton Wamalwa, Julie Overbaugh, and Dara A. Lehman. 2023. "Higher HIV-1 Env gp120-Specific Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity Is Associated with Lower Levels of Defective HIV-1 Provirus" Viruses 15, no. 10: 2055. https://doi.org/10.3390/v15102055