
Current Knowledge on the Interaction of Human Cytomegalovirus Infection, Encoded miRNAs, and Acute Aortic Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Pathophysiology
3. HCMV, RNAs, and Aortic Dissection
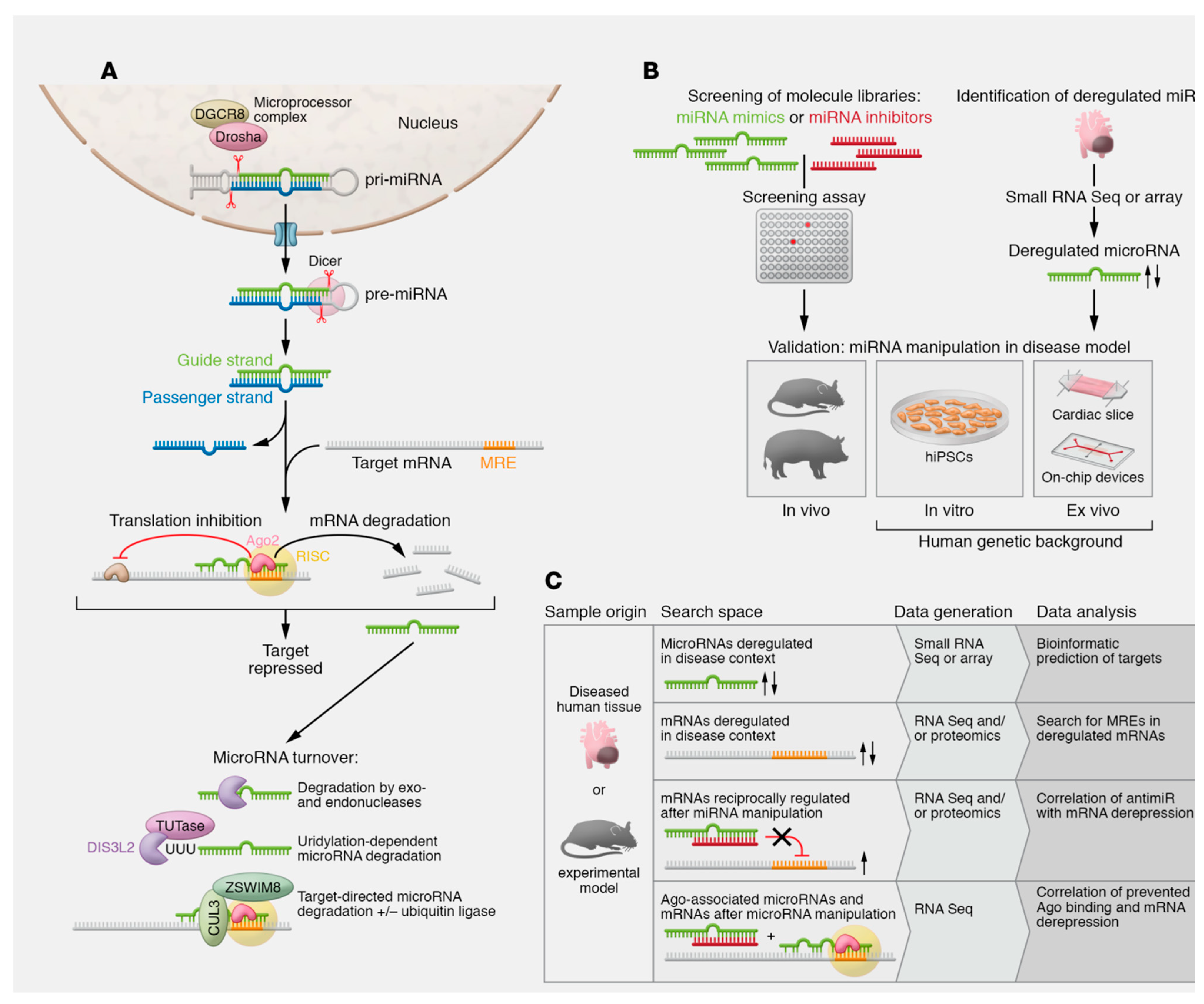
3.1. Human-Cytomegalovirus-Encoded MicroRNAs in Acute Aortic Syndrome
3.2. To Deploy HCMV Latent Infection and Promote Specific Pathological Process via miRNA
3.3. HCMV-Encoded miRNAs Inhibit Viral DNA Replication

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MiRNA Type | Target Gene | Annotated Function | Reference Type Cells | Authors |
|---|---|---|---|---|
| UL112 UL112 | IE72 IE1 | Regulation of viral DNA synthesis Regulation of DNA synthesis | HFs HEK239 | Grey et al. [85] Murphy et al. [88] |
| US25-2 | IE72 and pp65 | Regulation of viral DNA synthesis | HFF | Stern-Ginossar et al. [81] |
| US25-1 | Regulation of viral DNA synthesis; V-ATPase induces acidification of endosome; regulation of cellular bioactivity | HFF HFs MRC-5 | Stern-Ginossar et al. [81] Pavelin et al. [89] Jiang et al. [80] | |
| US33-5p | STX3 | Regulation of cell growth and cytokinesis | MRC-5 | Guo et al. [94] |
| US25-2-5p | eIF4A1 | Regulation of gene expression | MRC-5 | Qi et al. [90] |
| US5-1 | Geminin | host DNA replication inhibitor | U373 | Jiang et al. [92] |
| ULD148D | IER5 | Regulation of cell division | Kasum-3 | Pan et al. [96] |
4. HCMV-Encoded miRNAs Modulate Natural Killer and Cytotoxic T Cell Responses
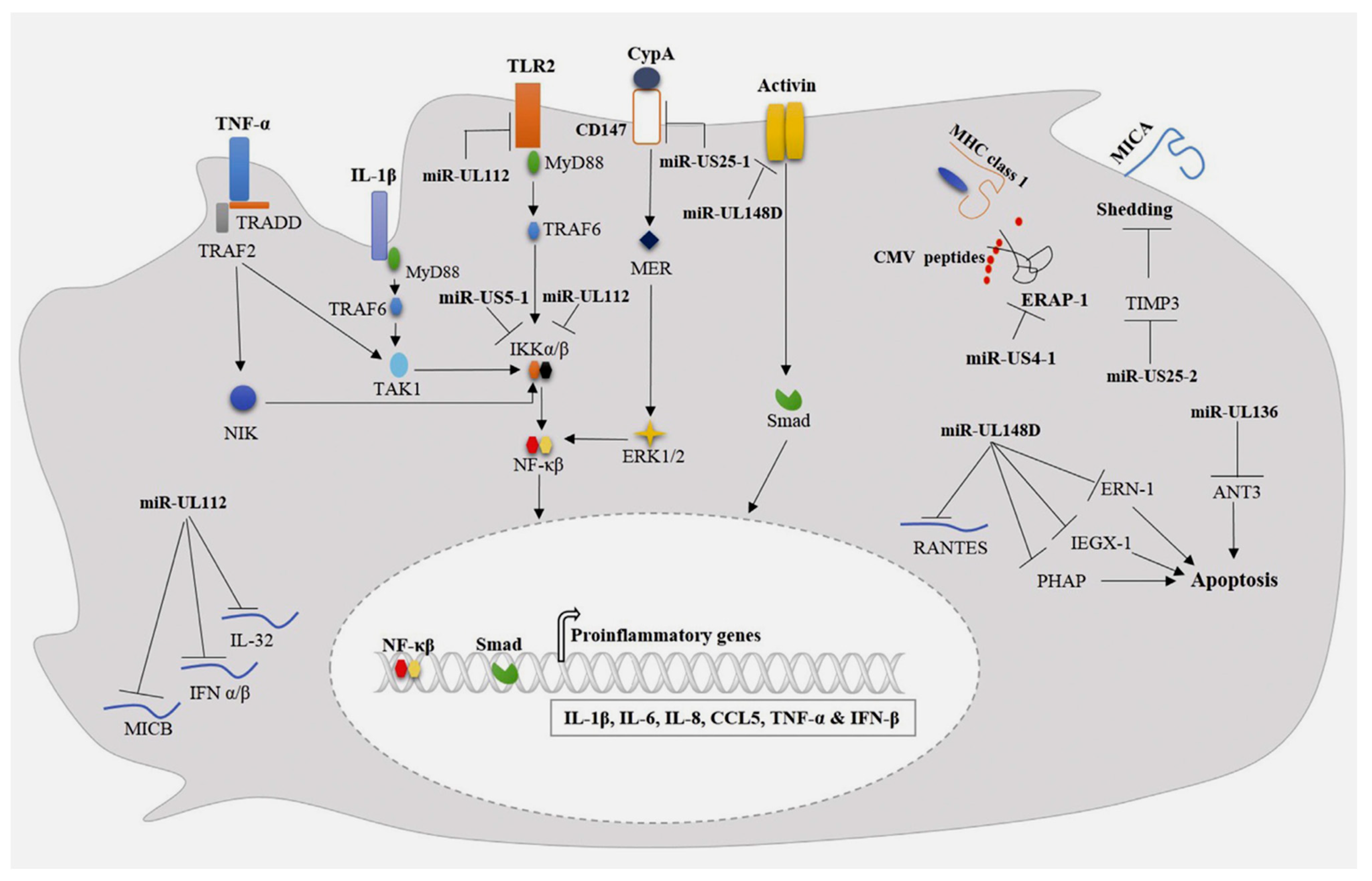
5. Aortic Acute Syndrome and Inflammatory Response: The Role of HCMV-Encoded miRNAs in Downregulation of Inflammatory Responses

6. Future Direction
6.1. Implementing Knowledge on HCMV, Long Noncoding RNAs, and Aortic Dissection
6.2. Implementing Knowledge on HCMV, Circular RNAs, and Aortic Dissection
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mussa, F.F.; Horton, J.D.; Moridzadeh, R.; Nicholson, J.; Trimarchi, S.; Eagle, K.A. Acute Aortic Dissection and Intramural Hematoma: A Systematic Review. JAMA 2016, 316, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Juraszek, A.; Czerny, M.; Rylski, B. Update in aortic dissection. Trends Cardiovasc. Med. 2022, 32, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Half of patients with acute aortic dissection in England die before reaching a specialist centre. BMJ 2020, 368, m304. [Google Scholar] [CrossRef] [PubMed]
- Braverman, A.C. Acute aortic dissection: Clinician update. Circulation 2010, 122, 184–188. [Google Scholar] [CrossRef]
- Howard, D.P.J.; Banerjee, A.; Fairhead, J.F.; Perkins, J.; Silver, L.E.; Rothwell, P.M.; Study, O.V. Population-based study of incidence and outcome of acute aortic dissection and premorbid risk factor control: 10-year results from the Oxford Vascular Study. Circulation 2013, 127, 2031–2037. [Google Scholar] [CrossRef]
- Bossone, E.; Gorla, R.; LaBounty, T.M.; Suzuki, T.; Gilon, D.; Strauss, C.; Ballotta, A.; Patel, H.J.; Evangelista, A.; Ehrlich, M.P.; et al. Presenting systolic blood pressure and outcomes in patients with acute aortic dissection. J. Am. Coll. Cardiol. 2018, 71, 1432–1440. [Google Scholar] [CrossRef]
- Biancari, F.; Juvonen, T.; Fiore, A.; Perrotti, A.; Hervé, A.; Touma, J.; Pettinari, M.; Peterss, S.; Buech, J.; Dell’Aquila, A.M.; et al. Current Outcome after Surgery for Type A Aortic Dissection. Ann. Surg. 2023, 278, e885–e892. [Google Scholar] [CrossRef]
- Harris, K.M.; Nienaber, C.A.; Peterson, M.D.; Woznicki, E.M.; Braverman, A.C.; Trimarchi, S.; Myrmel, T.; Pyeritz, R.; Hutchison, S.; Strauss, C.; et al. Early mortality in type A acute aortic dissection: Insights from the International Registry of Acute Aortic Dissection. JAMA Cardiol. 2022, 7, e222718. [Google Scholar] [CrossRef]
- Benedetto, U.; Dimagli, A.; Kaura, A.; Sinha, S.; Mariscalco, G.; Krasopoulos, G.; Moorjani, N.; Field, M.; Uday, T.; Kendal, S.; et al. Determinants of outcomes following surgery for type A acute aortic dissection: The UK National Adult Cardiac Surgical Audit. Eur. Heart J. 2021, 43, 44–52. [Google Scholar] [CrossRef]
- O’Hara, D.; McLarty, A.; Sun, E.; Itagaki, S.; Tannous, H.; Chu, D.; Egorova, N.; Chikwe, J. Type-A aortic dissection and cerebral perfusion: The Society of Thoracic Surgeons Database Analysis. Ann. Thorac. Surg. 2020, 110, 1461–1467. [Google Scholar] [CrossRef]
- Geirsson, A.; Shioda, K.; Olsson, C.; Ahlsson, A.; Gunn, J.; Hansson, E.C.; Hjortdal, V.; Jeppsson, A.; Mennander, A.; Wickbom, A.; et al. Differential outcomes of open and clamp-on distal anastomosis techniques in acute type A aortic dissection. J. Thorac. Cardiovasc. Surg. 2019, 157, 1750. [Google Scholar] [CrossRef] [PubMed]
- Czerny, M.; Schoenhoff, F.; Etz, C.; Englberger, L.; Khaladj, N.; Zierer, A.; Weigang, E.; Hoffmann, I.; Blettner, M.; Carrel, T.P. The impact of pre-operative malperfusion on outcome in acute type A aortic dissection: Results from the GERAADA Registry. J. Am. Coll. Cardiol. 2015, 65, 2628–2635. [Google Scholar] [CrossRef] [PubMed]
- Flachskampf, F.A.; Daniel, W.G. Aortic dissection. Cardiol. Clin. 2000, 18, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Akutsu, K. Etiology of aortic dissection. Gen. Thorac. Cardiovasc. Surg. 2019, 67, 271–276. [Google Scholar] [CrossRef]
- Khan, I.A.; Nair, C.K. Clinical, diagnostic, and management perspectives of aortic dissection. Chest 2002, 122, 311–328. [Google Scholar] [CrossRef]
- Mukherjee, D.; Eagle, K.A. Aortic dissection-an update. Curr. Probl. Cardiol. 2005, 30, 287–325. [Google Scholar] [CrossRef]
- Laggerbauer, B.; Engelhardt, S. MicroRNAs as therapeutic targets in cardiovascular disease. J. Clin. Investig. 2022, 132, e159179. [Google Scholar] [CrossRef]
- Hook, L.; Hancock, M.; Landais, I.; Grabski, R.; Britt, W.; Nelson, J.A. Cytomegalovirus microRNAs. Curr. Opin. Virol. 2014, 7, 40–46. [Google Scholar] [CrossRef]
- Sinzger, C.; Grefte, A.; Plachter, B.; Gouw, A.S.; The, T.H.; Jahn, G. Fibroblasts, epithelial cells, endothelial cells and smooth muscle cells are major targets of human cytomegalovirus infection in lung and gastrointestinal tissues. J. Gen. Virol. 1995, 76 Pt 4, 741–750. [Google Scholar] [CrossRef]
- Fedak, P.W.; de Sa, M.P.; Verma, S.; Nili, N.; Kazemian, P.; Butany, J.; Strauss, B.H.; Weisel, R.D.; David, T.E. Vascular matrix remodeling in patients with bicuspid aortic valve malformations: Implications for aortic dilatation. J. Thorac. Cardiovasc. Surg. 2003, 126, 797–806. [Google Scholar] [CrossRef]
- Shen, Y.H.; LeMaire, S.A. Molecular pathogenesis of genetic and sporadic aortic aneurysms and dissections. Curr. Probl. Surg. 2017, 54, 95–155. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.D.; Schwartz, M.A.; Tellides, G.; Milewicz, D.M. Role of mechanotransduction in vascular biology focus on thoracic aortic aneurysms and dissections. Circ. Res. 2015, 116, 1448–1461. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, K.B.; van Merrienboer, T.; Ket, J.; Bogunovic, N.; van der Velden, J.; Yeung, K.K. The role of vascular smooth muscle cells in the development of aortic aneurysms and dissections. Eur. J. Clin. Investig. 2022, 52, e13697. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Lin, Z.; Cao, H.; Chen, Y.; Li, J.; Zhuang, X.; Ma, D.; Ji, L.; Li, W.; Xu, S.; et al. Anxa1 in smooth muscle cells protects against acute aortic dissection. Cardiovasc. Res. 2022, 118, 1564–1582. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shi, E.; Gu, T.; Tang, R.; Gao, S.; Wang, Y.; Liu, H. Overexpression of microRNA-30a contributes to the development of aortic dissection by targeting lysyl oxidase. J. Thorac. Cardiovasc. Surg. 2017, 154, 1862–1869. [Google Scholar] [CrossRef]
- Sun, Y.; Xiao, Y.; Sun, H.; Zhao, Z.; Zhu, J.; Zhang, L.; Dong, J.; Han, T.; Jing, Q.; Zhou, J.; et al. miR-27a regulates vascular remodeling by targeting endothelial cells’ apoptosis and interaction with vascular smooth muscle cells in aortic dissection. Theranostics 2019, 9, 7961–7975. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, C.Q.; Peng, G.Y.; Huang, H.Y.; Yu, Y.S.; Ji, Z.C.; Shen, Z.Y. MicroRNA-134-5p regulates media degeneration through inhibiting VSMC phenotypic switch and migration in thoracic aortic dissection. Mol. Ther.-Nucleic Acids 2019, 16, 284–294. [Google Scholar] [CrossRef]
- Duan, H.; Zhang, X.; Song, R.; Liu, T.; Zhang, Y.; Yu, A. Upregulation of miR-133a by adiponectin inhibits pyroptosis pathway and rescues acute aortic dissection. Acta Biochim. Biophys. Sin. 2020, 52, 988–997. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, B.; Li, Y.; Yang, F.; Liu, Y.; Xu, Z.; Wang, G. Dysregulated long non-coding RNAs involved in regulation of matrix degradation during type-B aortic dissection pathogenesis. Gen. Thorac. Cardiovasc. Surg. 2021, 69, 238–245. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ying, S.Y.; Chang, D.C.; Miller, J.D.; Lin, S.L. The microRNA: Overview of the RNA gene that modulates gene functions. Methods Mol. Biol. 2006, 342, 1–18. [Google Scholar] [PubMed]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Lucas, T.; Bonauer, A.; Dimmeler, S. RNA Therapeutics in Cardiovascular Disease. Circ. Res. 2018, 123, 205–220. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Chen, Z.; Liang, H.; Chen, X.; Ke, Y.; Zhou, Z.; Yang, M.; Zen, K.; Yang, R.; Liu, C.; Zhang, C.Y. An Ebola virus-encoded microRNA-like fragment serves as a biomarker for early diagnosis of Ebola virus disease. Cell Res. 2016, 26, 380–383. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, J.; Wang, Z.; Sun, Y.; Wu, J.; Zhang, Y.; Liu, X.; Zhou, Z.; Zhou, L.; Zhang, C.Y.; et al. A virus-derived microRNA-like small RNA serves as a serum biomarker to prioritize the COVID-19 patients at high risk of developing severe disease. Cell Discov. 2021, 7, 48. [Google Scholar] [CrossRef]
- Söderberg-Nauclér, C. HCMV microinfections in inflammatory diseases and cancer. J. Clin. Virol. 2008, 41, 218–223. [Google Scholar] [CrossRef]
- Collins-McMillen, D.; Buehler, J.; Peppenelli, M.; Goodrum, F. Molecular determinants and the regulation of human cytomegalovirus latency and reactivation. Viruses 2018, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, T.; Hò, G.T.; Pump, W.C.; Blasczyk, R.; Bade-Doeding, C. Battle between host immune cellular responses and HCMV immune evasion. Int. J. Mol. Sci. 2019, 20, 3626. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.E.; Mahjoob, M.O.; Abosalif, K.O.A.; Ejaz, H.; Alameen, A.A.M.; Elsaman, T. Human cytomegalovirus-encoded MicroRNAs: A master regulator of latent infection. Infect. Genet. Evol. 2020, 78, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, J.; Liu, Z. MicroRNAs expressed by human cytomegalovirus. Virol. J. 2020, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Goldberger, T.; Mandelboim, O. The use of microRNA by human viruses: Lessons from NK cells and HCMV infection. Semin. Immunopathol. 2014, 36, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, J.; Zhang, W.; Chen, Y.; Zhang, K.; Popescu, L.M.; Ma, X.; Lau, W.B.; Rong, R.; Yu, X.; et al. Signature microRNA expression profile of essential hypertension and its novel link to human cytomegalovirus infection. Circulation 2011, 36, 659–674. [Google Scholar] [CrossRef]
- Ding, M.; Wang, X.; Wang, C.; Liu, X.; Zen, K.; Wang, W.; Zhang, C.Y.; Zhang, C. Distinct expression profile of HCMV encoded miRNAs in plasma from oral lichen planus patients. J. Transl. Med. 2018, 15, 133. [Google Scholar] [CrossRef]
- Dhuruvasan, K.; Sivasubramanian, G.; Pellett, P.E. Roles of host and viral microRNAs in human cytomegalovirus biology. Virus Res. 2011, 157, 180–192. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, N.; Zhou, Z.; Liang, H.; Pan, C.; Zhu, D.; Liu, F.; Zhang, C.Y.; Zhang, Y.; Zen, K. Circulating human cytomegalovirus-encoded HCMV-miR-US4-1 as an indicator for predicting the efficacy of IFN alpha treatment in chronic hepatitis B patients. Sci. Rep. 2016, 6, 23007. [Google Scholar] [CrossRef]
- Barozai, N.; Barozai, M.Y.; Din, M.; Iqbal, A. Differential expressional analysis of serum microRNAs in hepatitis C patients of Quetta, Pakistan. Pure Appl. Biol. 2018, 7, 853–862. [Google Scholar] [CrossRef]
- Vescovini, R.; Telera, A.R.; Pedrazzoni, M.; Abbate, B.; Rossetti, P.; Verzicco, I.; Arcangeletti, M.C.; Medici, M.C.; Calderaro, A.; Volpi, R.; et al. Impact of persistent cytomegalovirus infection on dynamic changes in human immune system profile. PLoS ONE 2016, 11, e0151965. [Google Scholar] [CrossRef] [PubMed]
- Grey, F. Role of microRNAs in herpesvirus latency and persistence. J. Gen. Virol. 2015, 96 Pt 4, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Razonable, R.R.; Humar, A. Cytomegalovirus in solid organ transplantation. Am. J. Transplant. 2013, 13 (Suppl. 4), 93–106. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.E.; Duan, X.; Deng, W.; Zeng, J.; Xie, J. MicroRNAs play big roles in modulating macrophages response toward mycobacteria infection. Infect. Genet. Evol. 2016, 45, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Rajewsky, N. The evolution of gene regulation by transcription factors and microRNAs. Nat. Rev. Genet. 2007, 8, 93–103. [Google Scholar] [CrossRef]
- Pfeffer, S.; Zavolan, M.; Grässer, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef]
- Gottwein, E.; Cullen, B.R. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe 2008, 3, 375–387. [Google Scholar] [CrossRef]
- Grey, F.; Antoniewicz, A.; Allen, E.; Saugstad, J.; McShea, A.; Carrington, J.C.; Nelson, J. Identification and characterization of human cytomegalovirus-encoded microRNAs. J. Virol. 2005, 79, 12095–12099. [Google Scholar] [CrossRef]
- Pfeffer, S.; Sewer, A.; Lagos-Quintana, M.; Sheridan, R.; Sander, C.; Grässer, F.A.; van Dyk, L.F.; Ho, C.K.; Shuman, S.; Chien, M.; et al. Identification of microRNAs of the herpesvirus family. Nat. Methods 2005, 2, 269–276. [Google Scholar] [CrossRef]
- Stark, T.J.; Arnold, J.D.; Spector, D.H.; Yeo, G.W. High-resolution profiling and analysis of viral and host small RNAs during human cytomegalovirus infection. J. Virol. 2012, 86, 226–235. [Google Scholar] [CrossRef]
- Cheng, Y.; Du, Y.; Wang, Q.; Lv, Q.; Xue, Y.; Zhou, W.; Zhang, C.; Chen, X.; Wang, D. Human cytomegalovirus-encoded microRNAs expression profile in plasma of patients with aortic dissection. J. Cardiothorac. Surg. 2023, 18, 39. [Google Scholar] [CrossRef] [PubMed]
- Scrivano, L.; Sinzger, C.; Nitschko, H.; Koszinowski, U.H.; Adler, B. HCMV spread and cell tropism are determined by distinct virus populations. PLoS Pathog. 2011, 7, e1001256. [Google Scholar] [CrossRef] [PubMed]
- Al Qaffas, A.; Camiolo, S.; Vo, M.; Aguiar, A.; Ourahmane, A.; Sorono, M.; Davison, A.J.; McVoy, M.A.; Hertel, L. Genome sequences of human cytomegalovirus strain TB40/E variants propagated in fibroblasts and epithelial cells. Virol. J. 2021, 18, 112. [Google Scholar] [CrossRef] [PubMed]
- Revello, M.G.; Gerna, G. Human cytomegalovirus tropism for endothelial/epithelial cells: Scientific background and clinical implications. Rev. Med. Virol. 2010, 20, 136–155. [Google Scholar] [CrossRef]
- Aguiar, A.; Galinato, M.; Bradley Silva, M.; Toth, B.; McVoy, M.A.; Hertel, L. Human Cytomegalovirus Replication and Infection-Induced Syncytia Formation in Labial, Foreskin, and Fetal Lung Fibroblasts. Viruses 2021, 13, 2355. [Google Scholar] [CrossRef]
- Fornara, C.; Schultz, E.; Lilleri, D.; Baldanti, F.; Ryckman, B.; Gerna, G. Fibroblast, Epithelial and Endothelial Cell-Derived Human Cytomegalovirus Strains Display Distinct Neutralizing Antibody Responses and Varying Levels of gH/gL Complexes. Int. J. Mol. Sci. 2023, 24, 4417. [Google Scholar] [CrossRef]
- Murrell, I.; Tomasec, P.; Wilkie, G.S.; Dargan, D.J.; Davison, A.J.; Stanton, R.J. Impact of sequence variation in the UL128 locus on production of human cytomegalovirus in fibroblast and epithelial cells. J. Virol. 2013, 87, 10489–10500. [Google Scholar] [CrossRef]
- Goodrum, F.; Caviness, K.; Zagallo, P. Human cytomegalovirus persistence. Cell Microbiol. 2012, 14, 644–655. [Google Scholar] [CrossRef]
- Britt, W.J. Maternal immunity and the natural history of congenital human cyto-megalovirus infection. Viruses 2018, 10, 405. [Google Scholar] [CrossRef]
- Christensen-Quick, A.; Vanpouille, C.; Lisco, A.; Gianella, S. Cytomegalovirus and HIV persistence: Pouring gas on the fire. AIDS Res. Hum. Retroviruses 2017, 33, S23–S30. [Google Scholar] [CrossRef]
- Fulkerson, H.L.; Nogalski, M.T.; Collins-McMillen, D.; Yurochko, A.D. Overview of Human Cytomegalovirus Pathogenesis. Methods Mol. Biol. 2021, 2244, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Reischig, T. Cytomegalovirus-associated renal allograft rejection: New challenges for antiviral preventive strategies. Expert Rev. Anti Infect. Ther. 2010, 8, 903–910. [Google Scholar] [CrossRef]
- Prichard, M.N.; Kern, E.R. The search for new therapies for human cytomegalovirus infections. Virus Res. 2011, 157, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Dupont, L.; Reeves, M.B. Cytomegalovirus latency and reactivation: Recent insights into an age old problem. Rev. Med. Virol. 2016, 26, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Hargett, D.; Shenk, T.E. Experimental human cytomegalovirus latency in CD14+ monocytes. Proc. Natl. Acad. Sci. USA 2010, 107, 20039–20044. [Google Scholar] [CrossRef]
- Poole, E.; Sinclair, J. Sleepless latency of human cytomegalovirus. Med. Microbiol. Immunol. 2015, 204, 421–429. [Google Scholar] [CrossRef]
- Elder, E.; Sinclair, J. HCMV latency: What regulates the regulators? Med. Microbiol. Immunol. 2019, 208, 431–438. [Google Scholar] [CrossRef]
- Sinclair, J.H.; Reeves, M.B. Human cytomegalovirus manipulation of latently infected cells. Viruses 2013, 5, 2803–2824. [Google Scholar] [CrossRef]
- Diggins, N.L.; Hancock, M.H. HCMV miRNA Targets Reveal Important Cellular Pathways for Viral Replication, Latency, and Reactivation. Noncoding RNA 2018, 4, 29. [Google Scholar] [CrossRef]
- Jiang, S.; Qi, Y.; He, R.; Huang, Y.; Liu, Z.; Ma, Y.; Guo, X.; Shao, Y.; Sun, Z.; Ruan, Q. Human cytomegalovirus microRNA miR-US25-1-5p inhibits viral replication by targeting multiple cellular genes during infection. Gene 2015, 570, 108–114. [Google Scholar] [CrossRef]
- Stern-Ginossar, N.; Saleh, N.; Goldberg, M.D.; Prichard, M.; Wolf, D.G.; Mandelboim, O. Analysis of human cytomegalovirus-encoded microRNA activity during infection. J. Virol. 2009, 83, 10684–10693. [Google Scholar] [CrossRef] [PubMed]
- Meshesha, M.K.; Bentwich, Z.; Solomon, S.A.; Avni, Y.S. In vivo expression of human cytomegalovirus (HCMV) microRNAs during latency. Gene 2016, 575, 101–107. [Google Scholar] [CrossRef]
- Fu, M.; Gao, Y.; Zhou, Q.; Zhang, Q.; Peng, Y.; Tian, K.; Wang, J.; Zheng, X. Human cytomegalovirus latent infection alters the expression of cellular and viral microRNA. Gene 2014, 536, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Tuddenham, L.; Pfeffer, S. Roles and regulation of microRNAs in cytomegalovirus infection. Biochim. Biophys. Acta 2011, 809, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Grey, F.; Meyers, H.; White, E.A.; Spector, D.H.; Nelson, J. A human cytomegalovirus-encoded microRNA regulates expression of multiple viral genes involved in replication. PLoS Pathog. 2007, 3, e163. [Google Scholar] [CrossRef]
- Huang, Y.; Qi, Y.; Ruan, Q.; Ma, Y.; He, R.; Ji, Y.; Sun, Z. A rapid method to screen putative mRNA targets of any known microRNA. Virol. J. 2011, 8, 8. [Google Scholar] [CrossRef]
- Lau, B.; Poole, E.; Van Damme, E.; Bunkens, L.; Sowash, M.; King, H.; Murphy, E.; Wills, M.; Van Loock, M.; Sinclair, J. Human cytomegalovirus miR-UL112-1 promotes the down-regulation of viral immediate early-gene expression during latency to prevent T-cell recognition of latently infected cells. J. Gen. Virol. 2016, 97, 2387–2398. [Google Scholar] [CrossRef]
- Murphy, E.; Vanícek, J.; Robins, H.; Shenk, T.; Levine, A.J. Suppression of immediate-early viral gene expression by herpesvirus-coded microRNAs: Implications for latency. Proc. Natl. Acad. Sci. USA 2008, 105, 5453–5458. [Google Scholar] [CrossRef]
- Pavelin, J.; Reynolds, N.; Chiweshe, S.; Wu, G.; Tiribassi, R.; Grey, F. Systematic microRNA analysis identifies ATP6V0C as an essential host factor for human cytomegalovirus replication. PLoS Pathog. 2013, 9, e1003820. [Google Scholar] [CrossRef]
- Qi, M.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. Over-expression of human cytomegalovirus miR-US25-2-3p downregulates eIF4A1 and inhibits HCMV replication. FEBS Lett. 2013, 587, 2266–2271. [Google Scholar] [CrossRef]
- Shen, Z.Z.; Pan, X.; Miao, L.F.; Ye, H.Q.; Chavanas, S.; Davrinche, C.; McVoy, M.; Luo, M.H. Comprehensive analysis of human cytomegalovirus microRNA expression during lytic and quiescent infection. PLoS ONE 2014, 9, e88531. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Huang, Y.; Qi, Y.; He, R.; Liu, Z.; Ma, Y.; Guo, X.; Shao, Y.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US5-1 inhibits viral replication by targeting Geminin mRNA. Virol. Sin. 2017, 32, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Bao, J.; Feng, R.; Zhao, Z.; Lu, Q.; Wang, G.; Li, H.; Su, D.; Zhou, J.; Jing, Q.; et al. Circulating microRNAs: A novel potential biomarker for diagnosing acute aortic dissection. Sci. Rep. 2017, 7, 12784. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Qi, Y.; Huang, Y.; Liu, Z.; Ma, Y.; Shao, Y.; Jiang, S.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US33-5p inhibits viral DNA synthesis and viral replication by down-regulating expression of the host Syntaxin3. FEBS Lett. 2015, 589, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Scheller, R.H. SNAREs--engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Pan, C.; Zhu, D.; Wang, Y.; Li, L.; Li, D.; Liu, F.; Zhang, C.Y.; Zen, K. Human Cytomegalovirus miR-UL148D Facilitates Latent Viral Infection by Targeting Host Cell Immediate Early Response Gene 5. PLoS Pathog. 2016, 12, e1006007. [Google Scholar] [CrossRef]
- Esteso, G.; Luzón, E.; Sarmiento, E.; Gómez-Caro, R.; Steinle, A.; Murphy, G.; Carbone, J.; Valés-Gómez, M.; Reyburn, H.T. Altered microRNA expression after infection with human cytomegalovirus leads to TIMP3 downregulation and increased shedding of metalloprotease substrates, including MICA. J. Immunol. 2014, 193, 1344–1352. [Google Scholar] [CrossRef]
- Kim, S.; Lee, S.; Shin, J.; Kim, Y.; Evnouchidou, I.; Kim, D.; Kim, Y.K.; Kim, Y.E.; Ahn, J.H.; Riddell, S.R.; et al. Human cytomegalovirus microRNA miR-US4-1 inhibits CD8(+) T cell responses by targeting the aminopeptidase ERAP1. Nat. Immunol. 2011, 12, 984–991. [Google Scholar] [CrossRef]
- Ma, Y.; Li, X.; Kuang, E. Viral Evasion of Natural Killer Cell Activation. Viruses 2016, 8, 95. [Google Scholar] [CrossRef]
- Miller-Kittrell, M.; Sparer, T.E. Feeling manipulated: Cytomegalovirus immune manipulation. Virol. J. 2009, 6, 4. [Google Scholar] [CrossRef]
- Molfetta, R.; Quatrini, L.; Santoni, A.; Paolini, R. Regulation of NKG2D-Dependent NK Cell Functions: The Yin and the Yang of Receptor Endocytosis. Int. J. Mol. Sci. 2017, 18, 1677. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, D.; He, J.; Cai, J.; Shen, K.; Liu, X.; Yang, X.; Xu, L. Hcmv-miR-UL112 attenuates NK cell activity by inhibition type I interferon secretion. Immunol. Lett. 2015, 163, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Hu, M.; Fedak, P.W.M.; Oudit, G.Y.; Kassiri, Z. Cell-Specific Functions of ADAM17 Regulate the Progression of Thoracic Aortic Aneurysm. Circ. Res. 2018, 123, 372–388. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Khalil, R.A. ADAM and ADAMTS disintegrin and metalloproteinases as major factors and molecular targets in vascular malfunction and disease. Adv. Pharmacol. 2022, 94, 255–363. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.A.V.; Lino, C.A.; Hang, H.T.; Alves, J.V.; Thang, B.Q.; Shin, S.J.; Sugiyama, K.; Matsunaga, H.; Takeyama, H.; Yamashiro, Y.; et al. Protective Role of Endothelial Fibulin-4 in Valvulo-Arterial Integrity. J. Am. Heart Assoc. 2023, 12, e026942. [Google Scholar] [CrossRef]
- De Cario, R.; Giannini, M.; Cassioli, G.; Kura, A.; Gori, A.M.; Marcucci, R.; Nistri, S.; Pepe, G.; Giusti, B.; Sticchi, E. Tracking an Elusive Killer: State of the Art of Molecular-Genetic Knowledge and Laboratory Role in Diagnosis and Risk Stratification of Thoracic Aortic Aneurysm and Dissection. Diagnostics 2022, 12, 1785. [Google Scholar] [CrossRef]
- Compton, T.; Kurt-Jones, E.A.; Boehme, K.W.; Belko, J.; Latz, E.; Golenbock, D.T.; Finberg, R.W. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2. J. Virol. 2003, 77, 4588–4596. [Google Scholar] [CrossRef]
- Szomolanyi-Tsuda, E.; Liang, X.; Welsh, R.M.; Kurt-Jones, E.A.; Finberg, R.W. Role for TLR2 in NK cell-mediated control of murine cytomegalovirus in vivo. J. Virol. 2006, 80, 4286–4291. [Google Scholar] [CrossRef]
- Landais, I.; Pelton, C.; Streblow, D.; DeFilippis, V.; McWeeney, S.; Nelson, J.A. Human Cytomegalovirus miR-UL112-3p Targets TLR2 and Modulates the TLR2/IRAK1/NFκB Signaling Pathway. PLoS Pathog. 2015, 11, e1004881. [Google Scholar] [CrossRef]
- Hancock, M.H.; Hook, L.M.; Mitchell, J.; Nelson, J.A. Human Cytomegalovirus MicroRNAs miR-US5-1 and miR-UL112-3p Block Proinflammatory Cytokine Production in Response to NF-κB-Activating Factors through Direct Downregulation of IKKα and IKKβ. mBio 2017, 8, e00109-17. [Google Scholar] [CrossRef]
- Dawar, F.U.; Xiong, Y.; Khattak, M.N.K.; Li, J.; Lin, L.; Mei, J. Potential role of cyclophilin A in regulating cytokine secretion. J. Leukoc. Biol. 2017, 102, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xia, S.; Yang, X.; Chen, H.; Li, F.; Liu, F.; Chen, Z. Human Cytomegalovirus Encoded miR-US25-1-5p Attenuates CD147/EMMPRIN-Mediated Early Antiviral Response. Viruses 2017, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.; Poole, E.; Krishna, B.; Sellart, I.; Wills, M.R.; Murphy, E.; Sinclair, J. The Expression of Human Cytomegalovirus MicroRNA MiR-UL148D during Latent Infection in Primary Myeloid Cells Inhibits Activin A-triggered Secretion of IL-6. Sci. Rep. 2016, 6, 31205. [Google Scholar] [CrossRef]
- Jones, K.L.; Mansell, A.; Patella, S.; Scott, B.J.; Hedger, M.P.; de Kretser, D.M.; Phillips, D.J. Activin A is a critical component of the inflammatory response, and its binding protein, follistatin, reduces mortality in endotoxemia. Proc. Natl. Acad. Sci. USA 2007, 104, 16239–16244. [Google Scholar] [CrossRef] [PubMed]
- Frascaroli, G.; Lecher, C.; Varani, S.; Setz, C.; van der Merwe, J.; Brune, W.; Mertens, T. Human Macrophages Escape Inhibition of Major Histocompatibility Complex-Dependent Antigen Presentation by Cytomegalovirus and Drive Proliferation and Activation of Memory CD4+ and CD8+ T Cells. Front. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.; Hong, A.; Song, J.; Lee, S.; Kim, M.; Hwang, S.Y.; Jeong, D.; Kim, J.; Son, A.; et al. Functional and molecular dissection of HCMV long non-coding RNAs. Sci. Rep. 2022, 12, 19303. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, N.; Zhou, X.; Bian, X.; Qiu, G.; Zhang, M.; Lin, H.; Li, D. LncRNA and mRNA interaction study based on transcriptome profiles reveals potential core genes in the pathogenesis of human thoracic aortic dissection. Mol. Med. Rep. 2018, 18, 3167–3176. [Google Scholar] [CrossRef]
- Sun, J.; Chen, G.; Jing, Y.; He, X.; Dong, J.; Zheng, J.; Zou, M.; Li, H.; Wang, S.; Sun, Y.; et al. LncRNA Expression Profile of Human Thoracic Aortic Dissection by High-Throughput Sequencing. Cell Physiol. Biochem. 2018, 46, 1027–1041. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, H.; Mai, C.; Qi, Y. Long Noncoding RNA XIST/miR-17/PTEN Axis Modulates the Proliferation and Apoptosis of Vascular Smooth Muscle Cells to Affect Stanford Type A Aortic Dissection. J. Cardiovasc. Pharmacol. 2020, 76, 53–62. [Google Scholar] [CrossRef]
- Li, S.; Zhao, X.; Cheng, S.; Li, J.; Bai, X.; Meng, X. Downregulating long non-coding RNA PVT1 expression inhibited the viability, migration and phenotypic switch of PDGF-BB-treated human aortic smooth muscle cells via targeting miR-27b-3p. Hum. Cell 2021, 34, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Wang, T.; Wei, X.; Wang, Y.; Ouyang, C.; Xie, Y.; Ye, X.; Han, Z. LncRNA H19 regulates smooth muscle cell functions and participates in the development of aortic dissection through sponging miR-193b-3p. Biosci. Rep. 2021, 41, BSR20202298. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, Q.; Wang, Y.; Piao, H.; Zhu, Z.; Li, D.; Wang, T.; Liu, K. LINC01278 Sponges miR-500b-5p to Regulate the Expression of ACTG2 to Control Phenotypic Switching in Human Vascular Smooth Muscle Cells During Aortic Dissection. J. Am. Heart Assoc. 2021, 10, e018062. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular rnas. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular rnas. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and characterizing circrna-protein interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef]
- Deng, J.; Huang, Y.; Wang, Q.; Li, J.; Ma, Y.; Qi, Y.; Liu, Z.; Li, Y.; Ruan, Q. Human Cytomegalovirus Influences Host circRNA Transcriptions during Productive Infection. Virol. Sin. 2021, 36, 241–253. [Google Scholar] [CrossRef]
- Yang, S.; Liu, X.; Wang, M.; Cao, D.; Jaijyan, D.K.; Enescu, N.; Liu, J.; Wu, S.; Wang, S.; Sun, W.; et al. Circular RNAs Represent a Novel Class of Human Cytomegalovirus Transcripts. Microbiol. Spectr. 2022, 10, e0110622. [Google Scholar] [CrossRef] [PubMed]
- Abere, B.; Zhou, H.; Li, J.; Cao, S.; Toptan, T.; Grundhoff, A.; Fischer, N.; Moore, P.S.; Chang, Y. Merkel Cell Polyomavirus Encodes Circular RNAs (circRNAs) Enabling a Dynamic circRNA/microRNA/mRNA Regulatory Network. mBio 2020, 11, e03059-20. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Wang, Q.; Zhang, J.; Ma, Y.; Qi, Y.; Liu, Z.; Li, Y.; Ruan, Q.; Huang, Y. Identification and characterization of human cytomegalovirus-encoded circular RNAs. Front. Cell. Infect. Microbiol. 2022, 12, 980974. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Tang, X.; Zhu, X.; Zhou, Q.; Guo, Y.; Zhao, R.; Wang, D.; Gong, B. Expression profiles of circrnas and the potential diagnostic value of serum circmark3 in human acute stanford type A aortic dissection. PLoS ONE 2019, 14, e219013. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Huang, C.; Li, X.; He, X.; Chen, Y.; Liao, W.; Liao, Y.; Sun, J.; Liu, Z.; Zhong, L.; et al. Circular RNA expression profile and potential function of hsa_circRNA_101238 in human thoracic aortic dissection. Oncotarget 2017, 8, 81825–81837. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Zhou, Z.; Li, H.; Tao, Q.; Wang, Y.; Lin, A.; Xu, J.; Zhang, B.; Wu, Y.; Min, H.; et al. Identification of pathological-related and diagnostic potential circular RNAs in Stanford type A aortic dissection. Front. Cardiovasc. Med. 2023, 9, 1074835. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, Y. Important Role of the IL-32 Inflammatory Network in the Host Response against Viral Infection. Viruses 2015, 7, 3116–3129. [Google Scholar] [CrossRef]
- Huang, Y.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. The expression of interleukin-32 is activated by human cytomegalovirus infection and down regulated by hcmv-miR-UL112-1. Virol. J. 2013, 10, 51. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nappi, F.; Alzamil, A.; Avtaar Singh, S.S.; Spadaccio, C.; Bonnet, N. Current Knowledge on the Interaction of Human Cytomegalovirus Infection, Encoded miRNAs, and Acute Aortic Syndrome. Viruses 2023, 15, 2027. https://doi.org/10.3390/v15102027
Nappi F, Alzamil A, Avtaar Singh SS, Spadaccio C, Bonnet N. Current Knowledge on the Interaction of Human Cytomegalovirus Infection, Encoded miRNAs, and Acute Aortic Syndrome. Viruses. 2023; 15(10):2027. https://doi.org/10.3390/v15102027
Chicago/Turabian StyleNappi, Francesco, Almothana Alzamil, Sanjeet Singh Avtaar Singh, Cristiano Spadaccio, and Nicolas Bonnet. 2023. "Current Knowledge on the Interaction of Human Cytomegalovirus Infection, Encoded miRNAs, and Acute Aortic Syndrome" Viruses 15, no. 10: 2027. https://doi.org/10.3390/v15102027