
The Disruption of a Nuclear Export Signal in the C-Terminus of the Herpes Simplex Virus 1 Determinant of Pathogenicity UL24 Protein Leads to a Syncytial Plaque Phenotype

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Viruses
2.2. Plasmid Construction
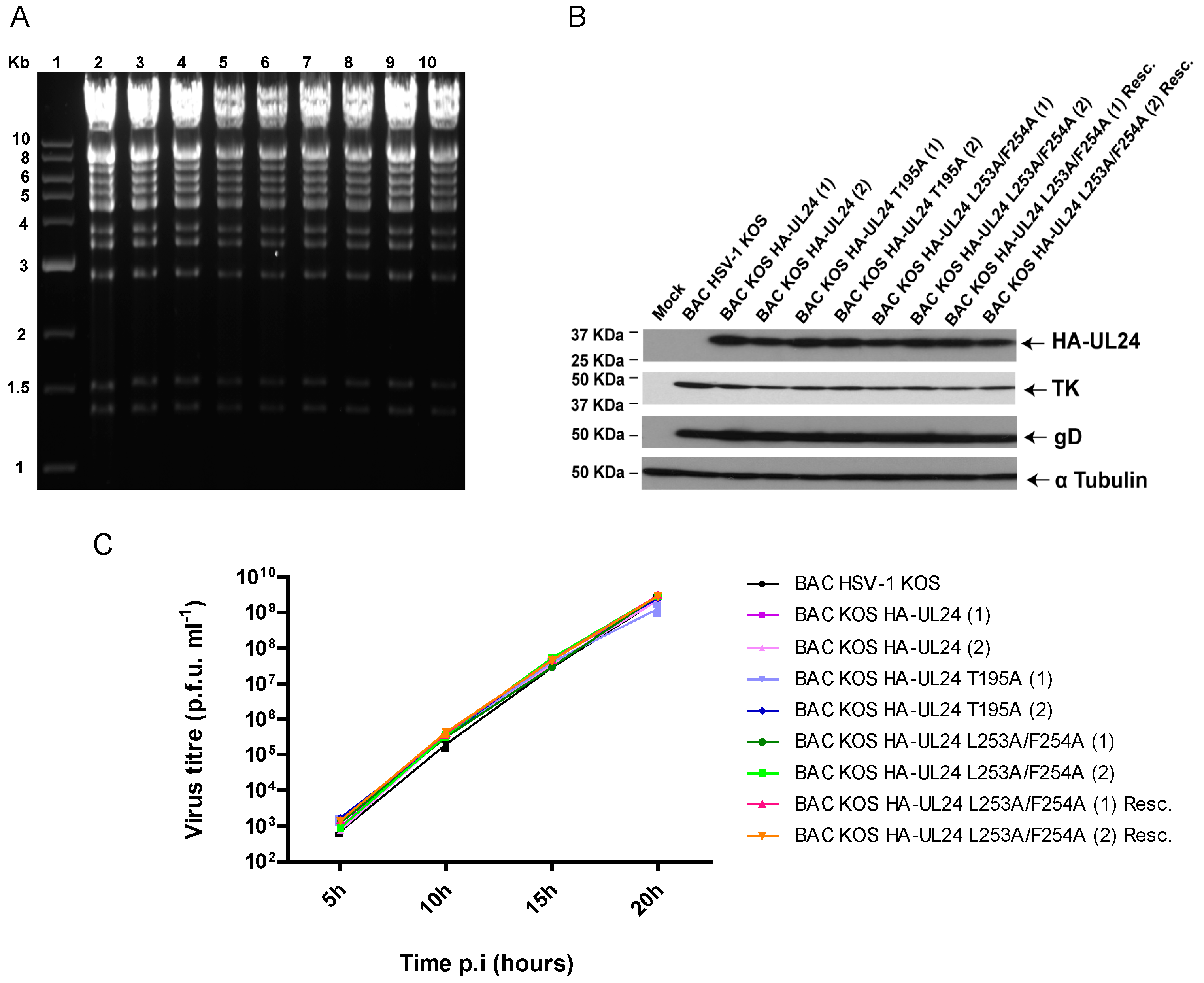
2.3. Generation of BACmids and Recombinant Viruses
2.4. Viral Replication Curves
2.5. Transient Transfections, Virus Infection, and Western Blotting
2.6. Confocal Microscopy
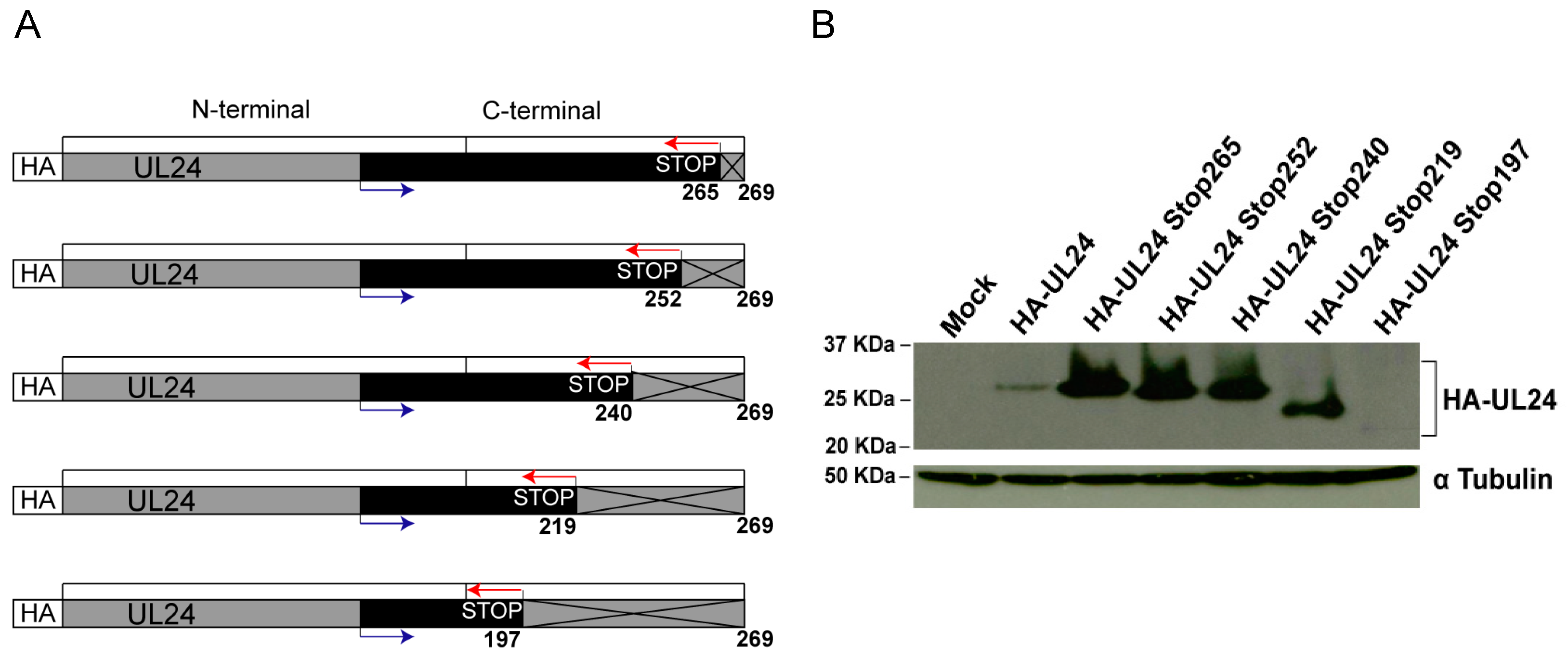
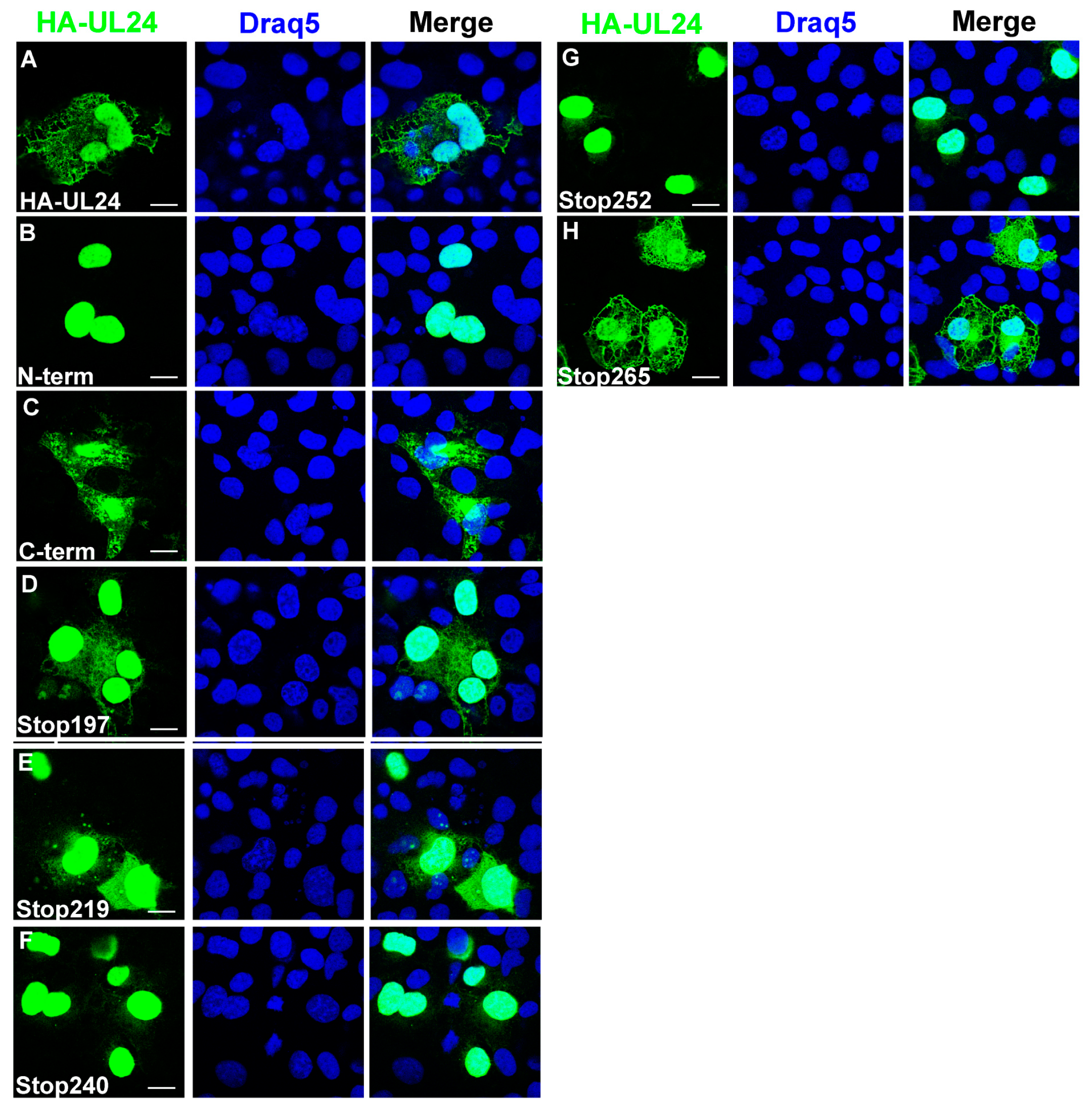
3. Results
3.1. Residues in the C-Terminal Domain of UL24 Are Important for the Cytoplasmic Accumulation of UL24
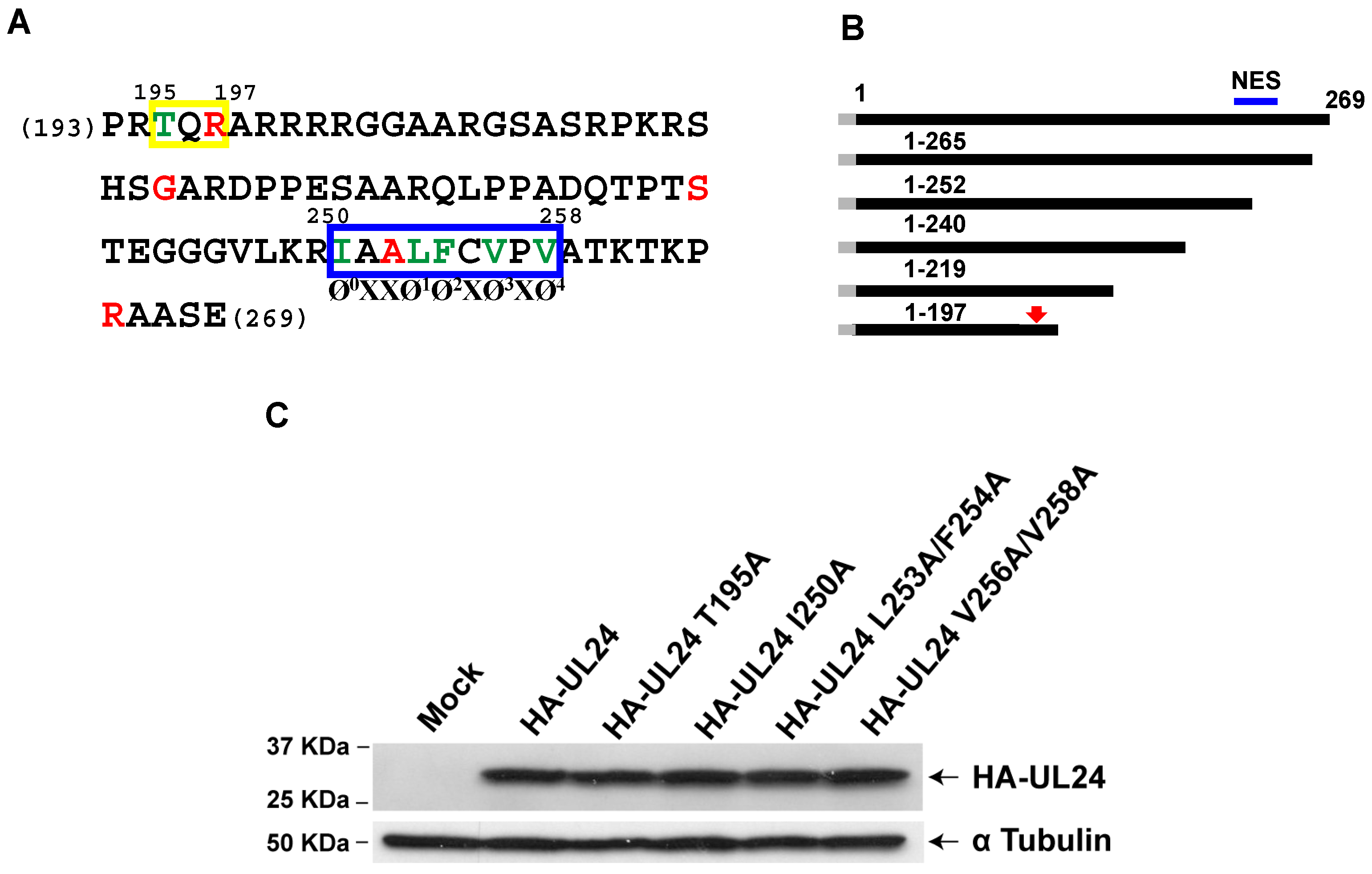
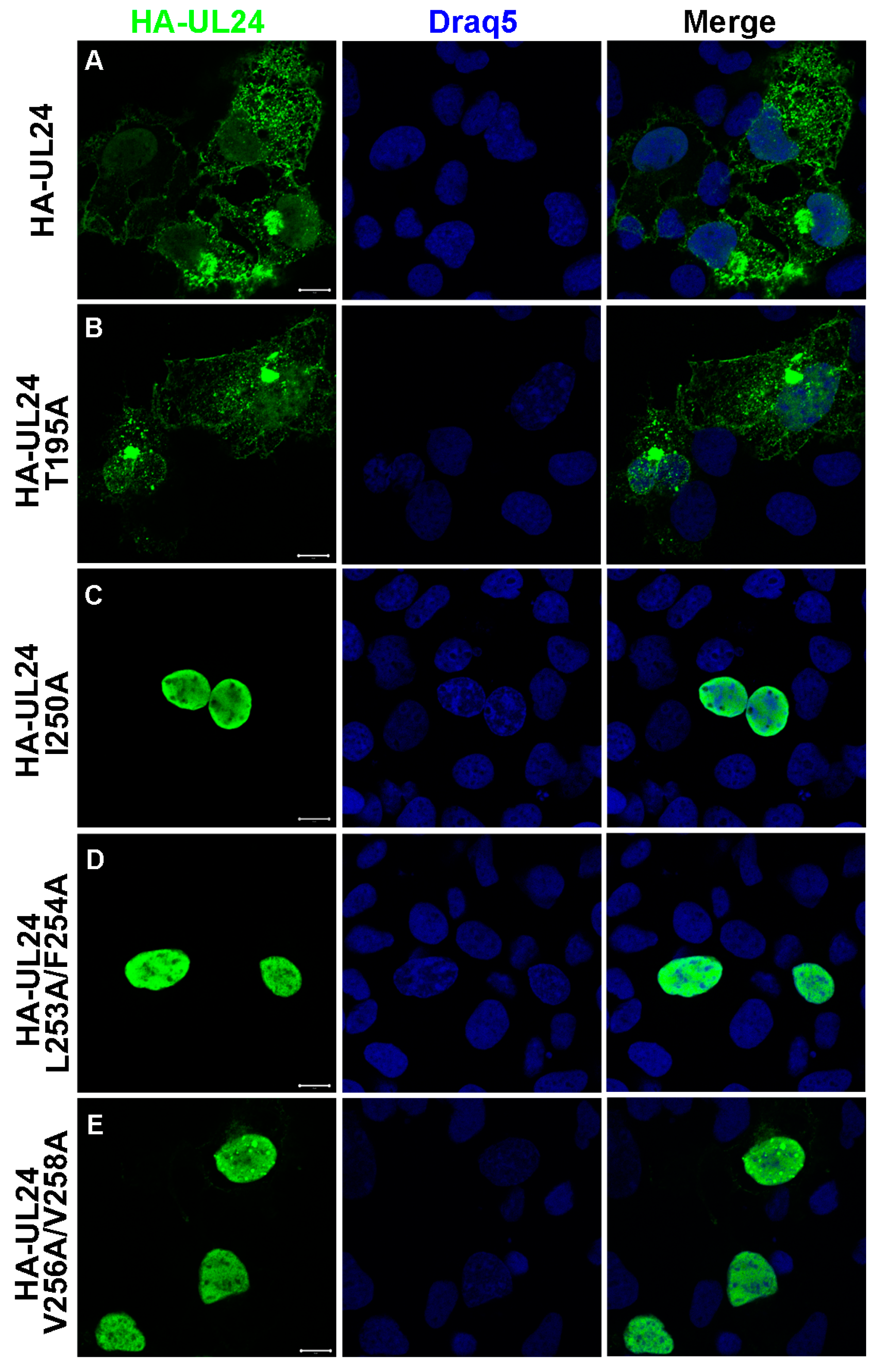
3.2. Identification of an NES in the C-Terminus of HSV-1 UL24
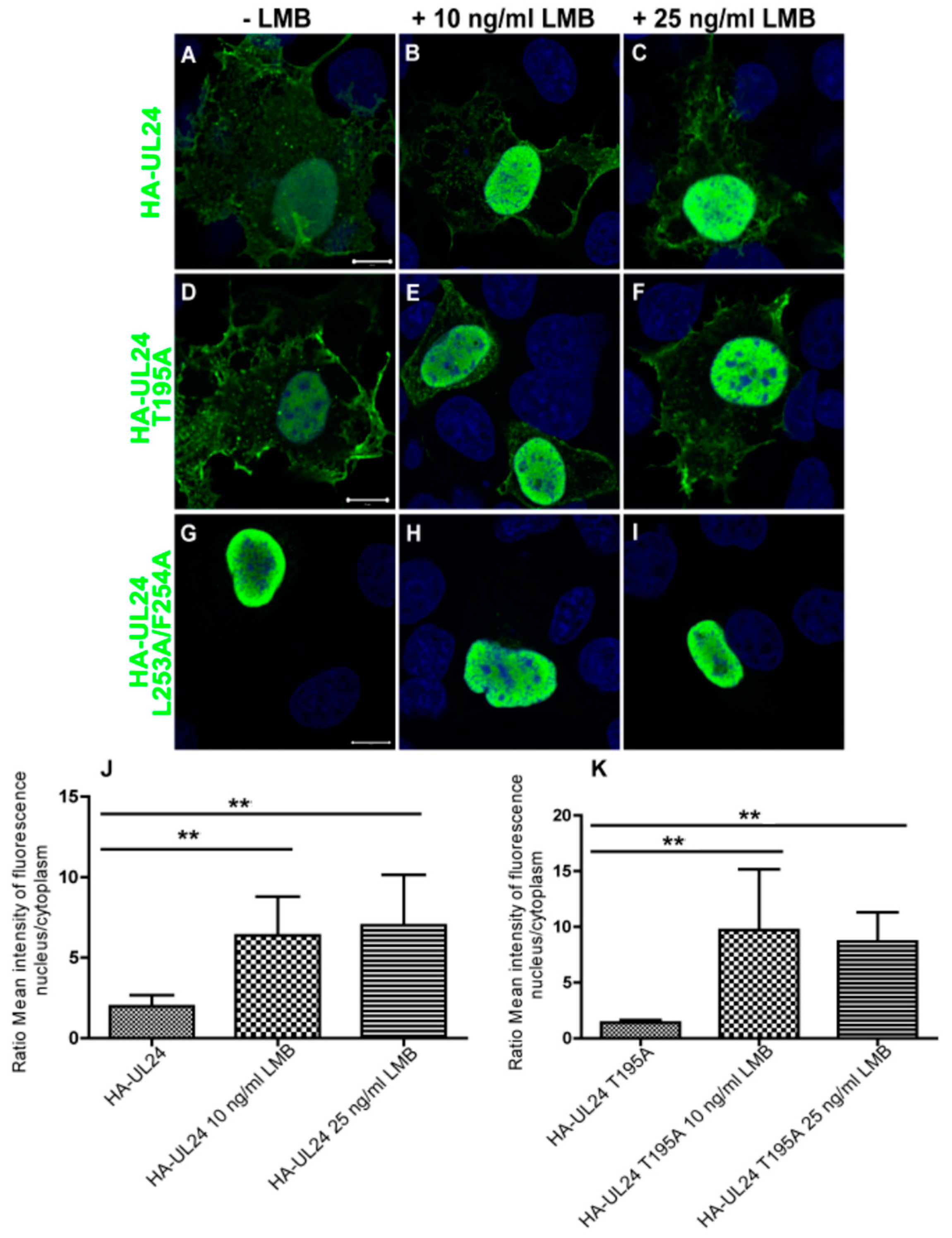
3.3. Nuclear Export Dependent on the UL24 NES Is Mediated by CRM-1 in Transiently Transfected Cells
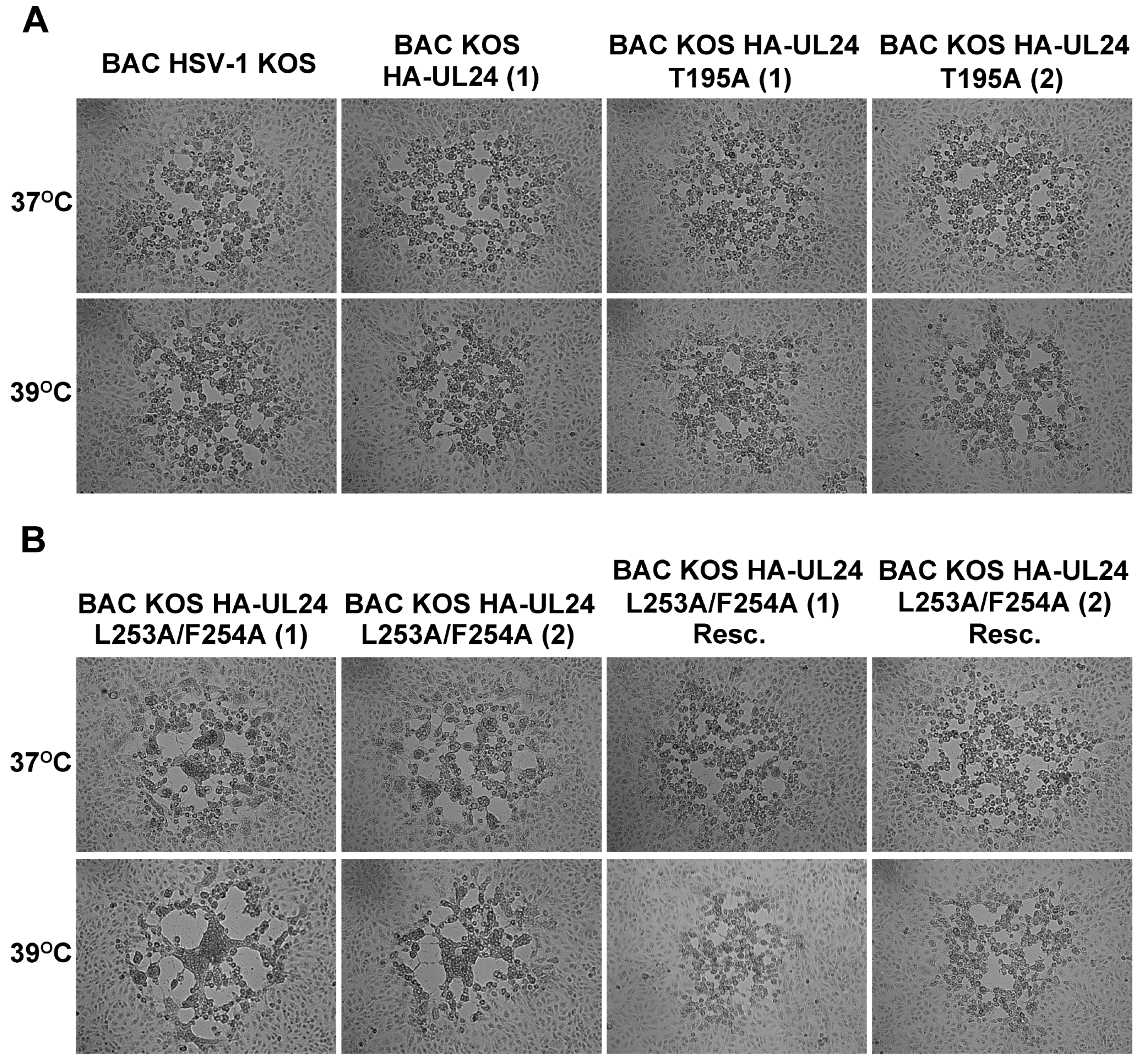
3.4. Mutations Inactivating the UL24 NES Lead to a Syncytial Plaque Phenotype
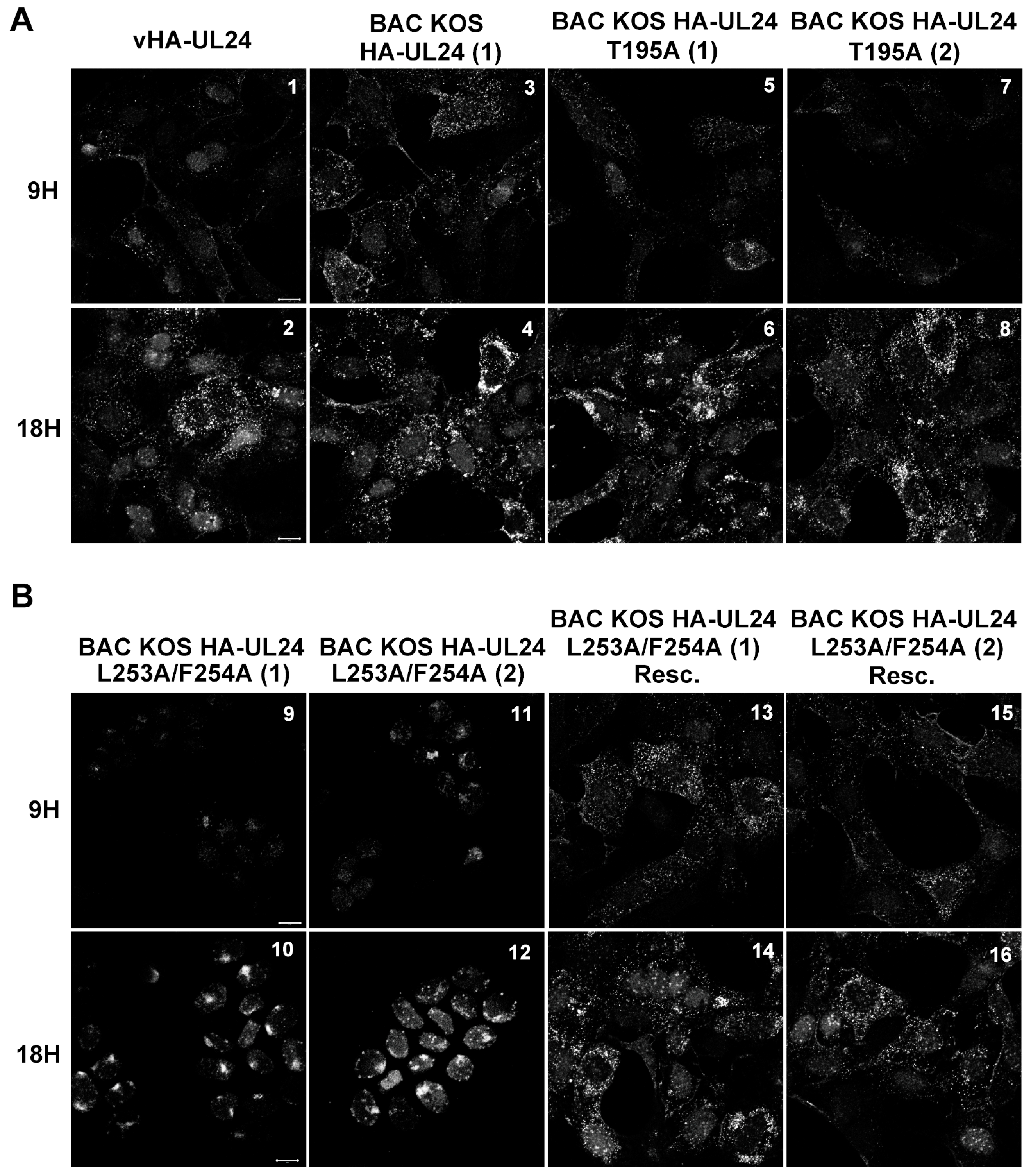
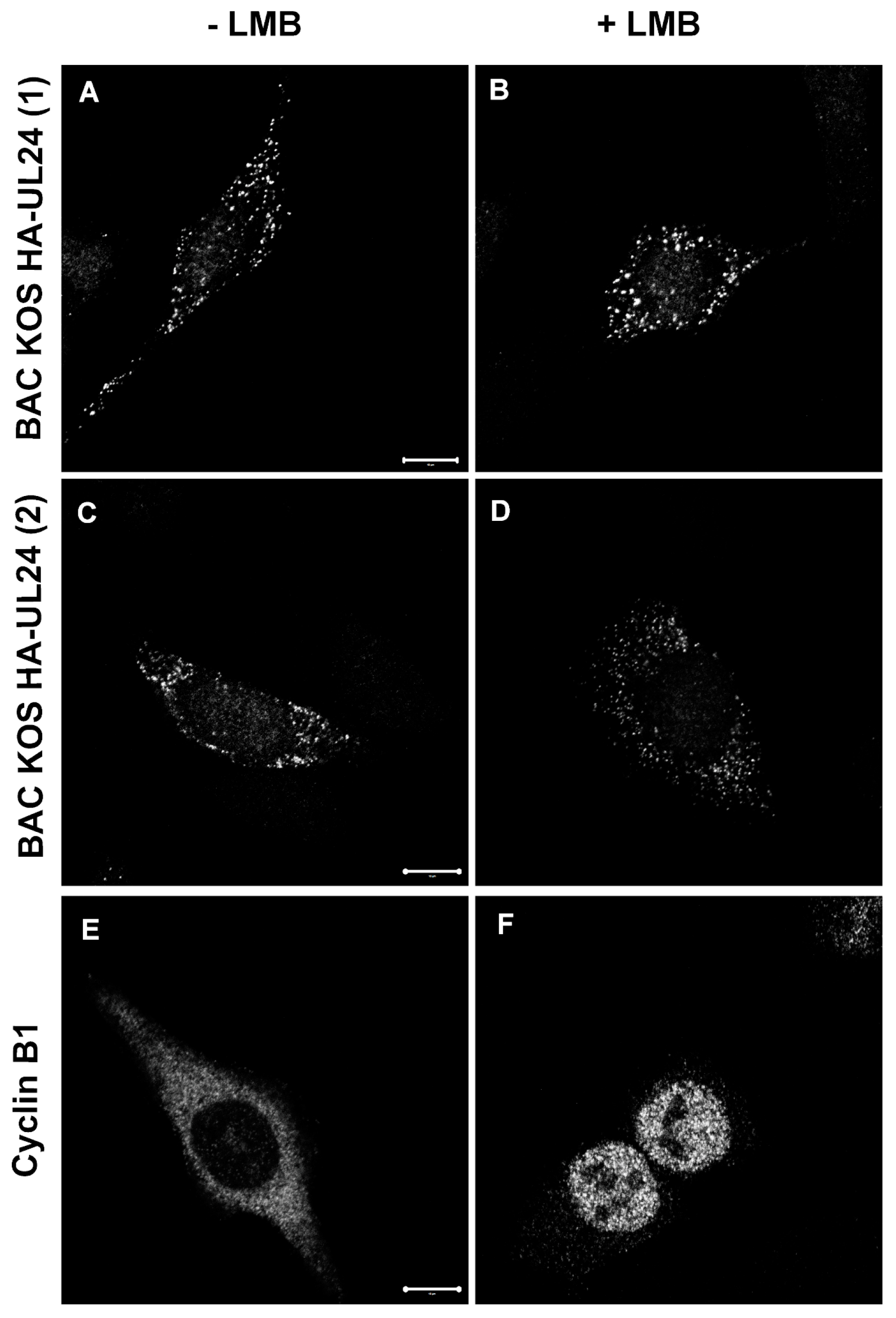
3.5. The HSV-1 UL24 Protein Shuttles between the Nucleus and the Cytoplasm during Infection
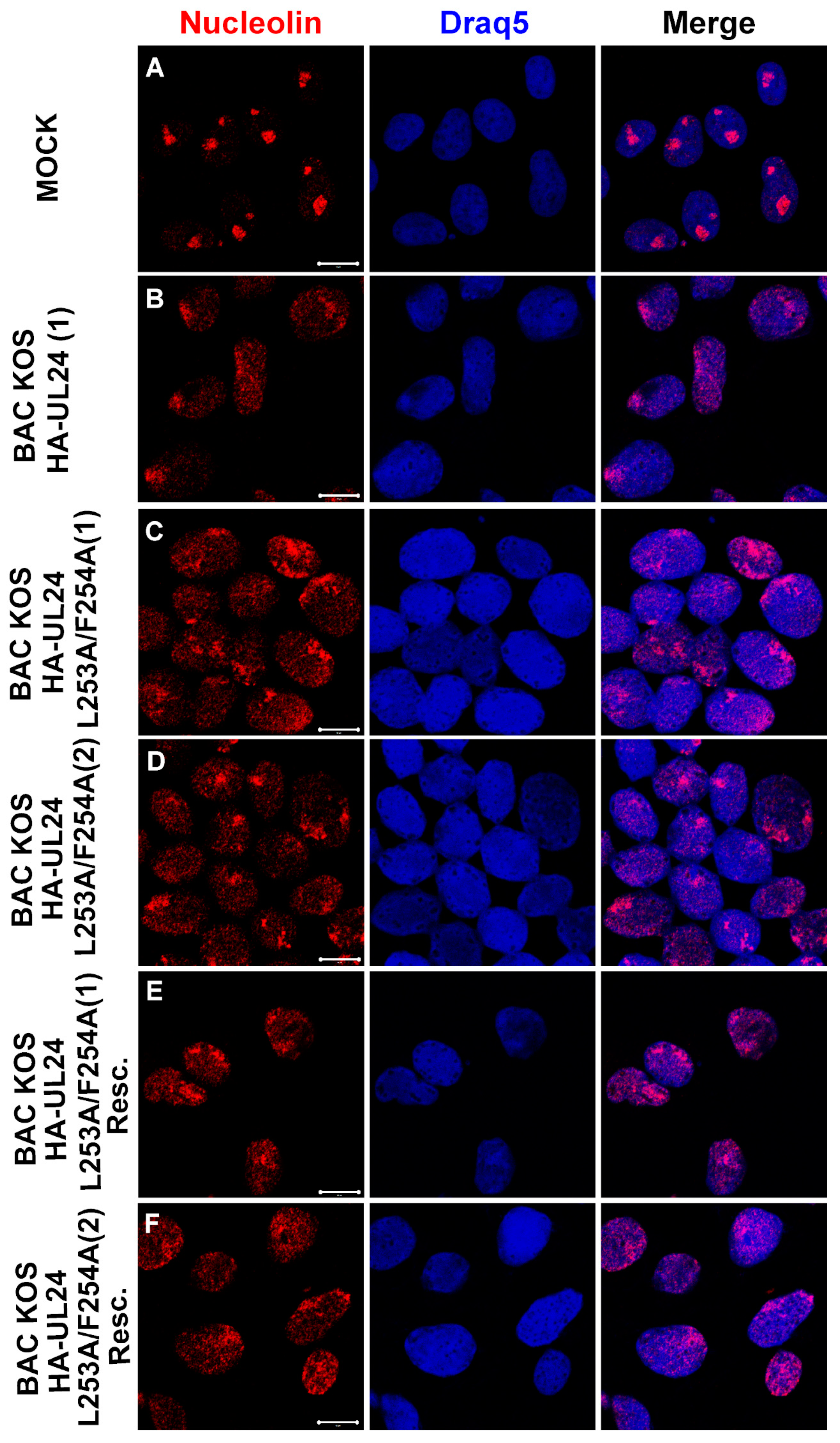
3.6. The UL24-Mediated Redistribution of Nucleolin Does Not Depend on the Ability of UL24 to Shuttle between the Nucleus and the Cytoplasm
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fatahzadeh, M.; Schwartz, R.A. Human herpes simplex virus infections: Epidemiology, pathogenesis, symptomatology, diagnosis, and management. J. Am. Acad. Dermatol. 2007, 57, 737–763. [Google Scholar] [CrossRef] [PubMed]
- McGeoch, D.J.; Cook, S.; Dolan, A.; Jamieson, F.E.; Telford, E.A. Molecular phylogeny and evolutionary timescale for the family of mammalian herpesviruses. J. Mol. Biol. 1995, 247, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.; Coen, D.M. Identification, localization, and regulation of expression of the UL24 protein of herpes simplex virus type 1. J. Virol. 2002, 76, 10821–10828. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.G.; Martin, S.L.; Coen, D.M. A conserved open reading frame that overlaps the herpes simplex virus thymidine kinase gene is important for viral growth in cell culture. J. Virol. 1989, 63, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Kniżewski, Ł.; Kinch, L.; Grishin, N.V.; Rychlewski, L.; Ginalski, K. Human herpesvirus 1 UL24 gene encodes a potential PD-(D/E)XK endonuclease. J. Virol. 2006, 80, 2575–2577. [Google Scholar] [CrossRef]
- Bertrand, L.; Leiva-Torres, G.A.; Hyjazie, H.; Pearson, A. Conserved residues in the UL24 protein of herpes simplex virus 1 are important for dispersal of the nucleolar protein nucleolin. J. Virol. 2010, 84, 109–118. [Google Scholar] [CrossRef]
- Leiva-Torres, G.A.; Rochette, P.-A.; Pearson, A. Differential importance of highly conserved residues in UL24 for herpes simplex virus 1 replication in vivo and reactivation. J. Gen. Virol. 2010, 91, 1109–1116. [Google Scholar] [CrossRef]
- Bertrand, L.; Pearson, A. The conserved N-terminal domain of herpes simplex virus 1 UL24 protein is sufficient to induce the spatial redistribution of nucleolin. J. Gen. Virol. 2008, 89, 1142–1151. [Google Scholar] [CrossRef]
- Sanders, P.G.; Wilkie, N.M.; Davison, A.J. Thymidine kinase deletion mutants of herpes simplex virus type 1. J. Gen. Virol. 1982, 63, 277–295. [Google Scholar] [CrossRef]
- Tognon, M.; Guandalini, R.; Romanelli, M.G.; Manservigi, R.; Trevisani, B. Phenotypic and genotypic characterization of locus Syn 5 in herpes simplex virus 1. Virus Res. 1991, 18, 135–150. [Google Scholar] [CrossRef]
- Rochette, P.-A.; Bourget, A.; Sanabria-Solano, C.; Lahmidi, S.; Lavallée, G.O.; Pearson, A. Mutation of UL24 impedes the dissemination of acute herpes simplex virus 1 infection from the cornea to neurons of trigeminal ganglia. J. Gen. Virol. 2015, 96, 2794–2805. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.G.; Chen, S.-H.; Cook, W.; Kramer, M.F.; Coen, D.M. Importance of the herpes simplex virus UL24 gene for productive ganglionic infection in mice. Virology 1998, 242, 161–169. [Google Scholar] [CrossRef]
- Lymberopoulos, M.H.; Bourget, A.; Ben Abdeljelil, N.; Pearson, A. Involvement of the UL24 protein in herpes simplex virus 1-induced dispersal of B23 and in nuclear egress. Virology 2011, 412, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Lymberopoulos, M.H.; Pearson, A. Involvement of UL24 in herpes-simplex-virus-1-induced dispersal of nucleolin. Virology 2007, 363, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, F.M.; Van Koningsbruggen, S.; Navascués, J.; Lamond, A.I. The multifunctional nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef]
- Xu, H.; Su, C.; Pearson, A.; Mody, C.H.; Zheng, C. Herpes Simplex Virus 1 UL24 Abrogates the DNA Sensing Signal Pathway by Inhibiting NF-kappaB Activation. J. Virol. 2017, 91, e00025-17. [Google Scholar] [CrossRef]
- Ben Abdeljelil, N.; Rochette, P.-A.; Pearson, A. The UL24 protein of herpes simplex virus 1 affects the sub-cellular distribution of viral glycoproteins involved in fusion. Virology 2013, 444, 263–273. [Google Scholar] [CrossRef]
- Gorlich, D.; Mattaj, I.W. Nucleocytoplasmic transport. Science 1996, 271, 1513–1518. [Google Scholar] [CrossRef]
- Nigg, E.A. Nucleocytoplasmic transport: Signals, mechanisms and regulation. Nature 1997, 386, 779–787. [Google Scholar] [CrossRef]
- Ossareh-Nazari, B.; Bachelerie, F.; Dargemont, C. Evidence for a role of CRM1 in signal-mediated nuclear protein export. Science 1997, 278, 141–144. [Google Scholar] [CrossRef]
- Kudo, N.; Matsumori, N.; Taoka, H.; Fujiwara, D.; Schreiner, E.P.; Wolff, B.; Yoshida, M.; Horinouchi, S. Leptomycin B inactivates CRM1/exportin 1 by covalent modification at a cysteine residue in the central conserved region. Proc. Natl. Acad. Sci. USA 1999, 96, 9112–9117. [Google Scholar] [CrossRef] [PubMed]
- Kutay, U.; Güttinger, S. Leucine-rich nuclear-export signals: Born to be weak. Trends Cell Biol. 2005, 15, 121–124. [Google Scholar] [CrossRef]
- Guttler, T.; Madl, T.; Neumann, P.; Deichsel, D.; Corsini, L.; Monecke, T.; Ficner, R.; Sattler, M.; Gorlich, D. NES consensus redefined by structures of PKI-type and Rev-type nuclear export signals bound to CRM1. Nat. Struct. Mol. Biol. 2010, 17, 1367–1376. [Google Scholar] [CrossRef]
- Fischer, U.; Huber, J.; Boelens, W.C.; Mattajt, L.W.; Lührmann, R. The HIV-1 Rev activation domain is a nuclear export signal that accesses an export pathway used by specific cellular RNAs. Cell 1995, 82, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Gomez Corredor, A.; Archambault, D. The bovine immunodeficiency virus Rev protein: Identification of a novel nuclear import pathway and nuclear export signal among retroviral Rev/Rev-like proteins. J. Virol. 2012, 86, 4892–4905. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Collado, M.; Rodriguez, E.; García, M.; Alvarez-Barrientos, A.; Arroyo, J.; Nombela, C.; Rivas, C. Identification of a nuclear export signal in the KSHV latent protein LANA2 mediating its export from the nucleus. Exp. Cell Res. 2005, 311, 96–105. [Google Scholar] [CrossRef]
- Marchand, C.; Lemay, G.; Archambault, D. The Jembrana disease virus Rev protein: Identification of nuclear and novel lentiviral nucleolar localization and nuclear export signals. PLoS ONE 2019, 14, e0221505. [Google Scholar] [CrossRef]
- Williams, P.; Verhagen, J.; Elliott, G. Characterization of a CRM1-dependent nuclear export signal in the C terminus of herpes simplex virus type 1 tegument protein UL47. J. Virol. 2008, 82, 10946–10952. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Lin, F.; Wang, S.; Xing, J. A novel virus-encoded nucleocytoplasmic shuttling protein: The UL3 protein of herpes simplex virus type 1. J. Virol. Methods 2011, 177, 206–210. [Google Scholar] [CrossRef]
- Otero, G.C.; Harris, M.E.; Donello, J.E.; Hope, T.J. Leptomycin B inhibits equine infectious anemia virus Rev and feline immunodeficiency virus rev function but not the function of the hepatitis B virus posttranscriptional regulatory element. J. Virol. 1998, 72, 7593–7597. [Google Scholar] [CrossRef]
- Rowland, R.R.; Yoo, D. Nucleolar-cytoplasmic shuttling of PRRSV nucleocapsid protein: A simple case of molecular mimicry or the complex regulation by nuclear import, nucleolar localization and nuclear export signal sequences. Virus Res. 2003, 95, 23–33. [Google Scholar] [CrossRef] [PubMed]
- De Jesús-González, L.A.; Palacios-Rápalo, S.N.; Reyes-Ruiz, J.M.; Osuna-Ramos, J.F.; Farfán-Morales, C.N.; Cordero-Rivera, C.D.; Cisneros, B.; Gutierrez-Escolano, A.L.; Del Angel, R.M. Nucleo-Cytoplasmic Transport of ZIKV Non-Structural 3 Protein Is Mediated by Importin-alpha/beta and Exportin CRM-1. J. Virol. 2023, 97, e0177322. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Maillard, P.; Minisini, R.; Vidalain, P.-O.; Roohvand, F.; Pecheur, E.-I.; Pirisi, M.; Budkowska, A. Identification of a functional, CRM-1-dependent nuclear export signal in hepatitis C virus core protein. PLoS ONE 2011, 6, e25854. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.J.; Niesman, A.; Chook, Y.M. An update to the CRM1 cargo/NES database NESdb. Mol. Biol. Cell 2021, 32, 467–469. [Google Scholar] [CrossRef]
- Griffiths, A.; Coen, D.M. High-frequency phenotypic reversion and pathogenicity of an acyclovir-resistant herpes simplex virus mutant. J. Virol. 2003, 77, 2282–2286. [Google Scholar] [CrossRef]
- Tischer, B.K.; von Einem, J.; Kaufer, B.; Osterrieder, N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques 2006, 40, 191–197. [Google Scholar] [PubMed]
- Tischer, B.K.; Smith, G.A.; Osterrieder, N. En passant mutagenesis: A two step markerless red recombination system. Methods Mol. Biol. 2010, 634, 421–430. [Google Scholar]
- Jurak, I.; Silverstein, L.B.; Sharma, M.; Coen, D.M. Herpes simplex virus is equipped with RNA- and protein-based mechanisms to repress expression of ATRX, an effector of intrinsic immunity. J. Virol. 2012, 86, 10093–10102. [Google Scholar] [CrossRef] [PubMed]
- Rust, H.L.; Thompson, P.R. Kinase consensus sequences: A breeding ground for crosstalk. ACS Chem. Biol. 2011, 6, 881–892. [Google Scholar] [CrossRef]
- Bell, C.; Desjardins, M.; Thibault, P.; Radtke, K. Proteomics analysis of herpes simplex virus type 1-infected cells reveals dynamic changes of viral protein expression, ubiquitylation, and phosphorylation. J. Proteome Res. 2013, 12, 1820–1829. [Google Scholar] [CrossRef]
- Niu, Y.; Roy, F.; Saltel, F.; Andrieu-Soler, C.; Dong, W.; Chantegrel, A.-L.; Accardi, R.; Thépot, A.; Foiselle, N.; Tommasino, M.; et al. A nuclear export signal and phosphorylation regulate Dok1 subcellular localization and functions. Mol. Cell. Biol. 2006, 26, 4288–4301. [Google Scholar] [CrossRef] [PubMed]
- Lahmidi, S.; Yousefi, M.; Dridi, S.; Duplay, P.; Pearson, A. Dok-1 and Dok-2 Are Required To Maintain Herpes Simplex Virus 1-Specific CD8+ T Cells in a Murine Model of Ocular Infection. J. Virol. 2017, 91, e02297-16. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bardes, E.S.; Moore, J.D.; Brennan, J.; Powers, M.A.; Kornbluth, S. Control of cyclin B1 localization through regulated binding of the nuclear export factor CRM1. Genes Dev. 1998, 12, 2131–2143. [Google Scholar] [CrossRef]
- Chen, X.; Kube, D.M.; Cooper, M.J.; Davis, P.B. Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Mol. Ther. 2008, 16, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mao, M.; Xu, J.C. Cell-surface nucleolin is involved in lipopolysaccharide internalization and signalling in alveolar macrophages. Cell Biol. Int. 2011, 35, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Doron-Mandel, E.; Koppel, I.; Abraham, O.; Rishal, I.; Smith, T.P.; Buchanan, C.N.; Sahoo, P.K.; Kadlec, J.; Oses-Prieto, J.A.; Kawaguchi, R.; et al. The glycine arginine-rich domain of the RNA-binding protein nucleolin regulates its subcellular localization. EMBO J. 2021, 40, e107158. [Google Scholar] [CrossRef]
- Dridi, S.; Richerioux, N.; Suarez, C.E.G.; Vanharen, M.; Sanabria-Solano, C.; Pearson, A. A Mutation in the UL24 Gene Abolishes Expression of the Newly Identified UL24.5 Protein of Herpes Simplex Virus 1 and Leads to an Increase in Pathogenicity in Mice. J. Virol. 2018, 92, e00671-18. [Google Scholar] [CrossRef]
- Lin, S.R.; Jiang, M.J.; Wang, H.H.; Hu, C.H.; Hsu, M.S.; His, E.; Duh, C.Y.; Wang, S.K. Human cytomegalovirus UL76 elicits novel aggresome formation via interaction with S5a of the ubiquitin proteasome system. J. Virol. 2013, 87, 11562–11578. [Google Scholar] [CrossRef]
- Finnen, R.L.; Johnston, S.M.; Neron, C.E.; Banfield, B.W. Nucleocytoplasmic shuttling of the HSV-2 serine/threonine kinase Us3. Virology 2011, 417, 229–237. [Google Scholar] [CrossRef]
- Lengyel, J.; Strain, A.K.; Perkins, K.D.; Rice, S.A. ICP27-dependent resistance of herpes simplex virus type 1 to leptomycin B is associated with enhanced nuclear localization of ICP4 and ICP0. Virology 2006, 352, 368–379. [Google Scholar] [CrossRef]
- Sandri-Goldin, R.M. ICP27 mediates HSV RNA export by shuttling through a leucine-rich nuclear export signal and binding viral intronless RNAs through an RGG motif. Genes Dev. 1998, 12, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Kutay, U.; Bischoff, F.R.; Kostka, S.; Kraft, R.; Görlich, D. Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell 1997, 90, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Lipowsky, G.; Bischoff, F.; Schwarzmaier, P.; Kraft, R.; Kostka, S.; Hartmann, E.; Kutay, U.; Görlich, D. Exportin 4: A mediator of a novel nuclear export pathway in higher eukaryotes. EMBO J. 2000, 19, 4362–4371. [Google Scholar] [CrossRef]
- Brownawell, A.M.; Macara, I.G. Exportin-5, a novel karyopherin, mediates nuclear export of double-stranded RNA binding proteins. J. Cell Biol. 2002, 156, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Li, L.; Sandri-Goldin, R.M. The cellular RNA export receptor TAP/NXF1 is required for ICP27-mediated export of herpes simplex virus 1 RNA, but the TREX complex adaptor protein Aly/REF appears to be dispensable. J. Virol. 2009, 83, 6335–6346. [Google Scholar] [CrossRef] [PubMed]
- Juillard, F.; Hiriart, E.; Sergeant, N.; Vingtdeux-Didier, V.; Drobecq, H.; Sergeant, A.; Manet, E.; Gruffat, H. Epstein-Barr virus protein EB2 contains an N-terminal transferable nuclear export signal that promotes nucleocytoplasmic export by directly binding TAP/NXF1. J. Virol. 2009, 83, 12759–12768. [Google Scholar] [CrossRef]
- Williams, B.J.; Boyne, J.R.; Goodwin, D.J.; Roaden, L.; Hautbergue, G.M.; Wilson, S.A.; Whitehouse, A. The prototype gamma-2 herpesvirus nucleocytoplasmic shuttling protein, ORF 57, transports viral RNA through the cellular mRNA export pathway. Biochem. J. 2005, 387, 295–308. [Google Scholar] [CrossRef]
- Daelemans, D.; Afonina, E.; Nilsson, J.; Werner, G.; Kjems, J.; De Clercq, E.; Pavlakis, G.N.; Vandamme, A.-M. A synthetic HIV-1 Rev inhibitor interfering with the CRM1-mediated nuclear export. Proc. Natl. Acad. Sci. USA 2002, 99, 14440–14445. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Martinez, J.P.; Hinkelmann, B.; Gerth, K.; Washausen, P.; Diez, J.; Frank, R.; Sasse, F.; Meyerhans, A. The myxobacterial metabolite ratjadone A inhibits HIV infection by blocking the Rev/CRM1-mediated nuclear export pathway. Microb. Cell Factories 2014, 13, 17. [Google Scholar] [CrossRef]
- Durut, N.; Sáez-Vásquez, J. Nucleolin: Dual roles in rDNA chromatin transcription. Gene 2015, 556, 7–12. [Google Scholar] [CrossRef]
- Chan, C.M.; Chu, H.; Zhang, A.J.; Leung, L.H.; Sze, K.H.; Kao, R.Y.; Chik, K.K.; To, K.K.; Chan, J.F.; Chen, H.; et al. Hemagglutinin of influenza A virus binds specifically to cell surface nucleolin and plays a role in virus internalization. Virology 2016, 494, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Su, P.Y.; Wang, Y.F.; Huang, S.W.; Lo, Y.C.; Wang, Y.H.; Wu, S.R.; Shieh, D.B.; Chen, S.H.; Wang, J.R.; Lai, M.D.; et al. Cell surface nucleolin facilitates enterovirus 71 binding and infection. J. Virol. 2015, 89, 4527–4538. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 5′ → 3′ a |
|---|---|
| UL24 5′BglIIF | CCCAGATCTGCGGCACGCTGTTG |
| UL24CtermStop795R | CACAGTCGACTCAGTCAGTCATCGGGGTTTGGTCTTGGTGG |
| UL24CtermStop756R | CACAGTCGACTCAGTCAGTCACGCCGCGATCCTCTTAAG |
| UL24CtermStop720R | CACAGTCGACTCAGTCAGTCAGGAGGCGGGGGTTTGGTC |
| UL24CtermStop657R | CACAGTCGACTCAGTCAGTCACCCAGAGTGTGACCTTTTCGG |
| UL24CtermStop591R | CACAGTCGACTCAGTCAGTCATCGCTGGGTCCTAGGCTCC |
| Name | Sequence 5′ → 3′ a | Mutation b |
|---|---|---|
| UL24_T195Atop | CCCATGGAGCCTAGGGCACAGCGAGCCCGTCGC | |
| botUL24_T195A | GCGACGGGCTCGCTGTGCCCTAGGCTCCATGGG | T195A |
| UL24_L253A-F254Atop | AAGAGGATCGCGGCGGCCGCCTGCGTGCCCGTGGC | |
| botUL24_L253A-F254A | GCCACGGGCACGCAGGCGGCCGCCGCGATCCTCTT | L253A–F254A |
| UL24_V256A-V258Atop | GCTCTTCTGCGCGCCCGCGCGCACCAAGA | |
| botUL24_V256A-V258A | TCTTGGTGGCCGCGGGCGCGCAGAAGAGC | V256A–V258A |
| Name | Sequence 5′→ 3′ a | Mutation |
|---|---|---|
| BAC_HA_UL24_F BAC_HA_UL24_R | TCCGTGGCTTCTTGCTGCCGGCGAGGGCGCAACGCCGTA CGTCGGTTGCTATGTACCCATACGATGTTCCAGATTACG CTGCCGCGAGAACGCGCAGCCTGGTCGAGGATGACGAC GATAAGTA CGTACCCCTGCCATCAACACGCGTCTGCGTTCGACCAGG CTGCGCGTTCTCGCGGCAGCGTAATCTGGAACATCGTA TGGGTACATAGCAACCGACGTACGGCGTTCAACCAATT AACCAATTCTGA | Insertion of N-terminal HA tag in UL24 |
BAC_HSV1T195A_F BAC_HSV1T195A_R | CGTGCGGATGCTCCAGAGCCTGTCCACGTATACGGTCCC CATGGAGCCTAGGGCACAGCGAGCCCGTCGCCGCCGAG GATGACGACGATAAGTA CTGCTCGCAGACCCCCGGGCAGCGCCGCCGCGGCGGCG ACGGGCTCGCTGTGCCCTAGGCTCCATGGGGACCGTACA ACCAATTAACCAATTCTGA | UL24 mutation T195AA |
BAC_HSV1L253A-F254A_F BAC_HSV1L253A-F254A_R RescueBAC_HSV1L253A-F254A_F RescueBAC_HSV1L253A-F254A_R | CAAACCCCCGCCTCCACGGAGGGCGGGGGGGTGCTTAA GAGGATCGCGGCGGCCGCCTGCGTGCCCGTGGCCACCA AAGGATGACGACGATAAGTA CTCTCATTCGGAGGCAGCTCGGGGTTTGGTCTTGGTGGCC ACGGGCACGCAGGCGGCCGCCGCGATCCTCTTAAGCAC AACCAATTAACCAATTCTGA CAAACCCCCGCCTCCACGGAGGGCGGGGGGGTGCTTAA GAGGATCGCGGCGCTCTTCTGCGTGCCCGTGGCCACCAA AGGATGACGACGATAAGTA CTCTCATTCGGAGGCAGCTCGGGGTTTGGTCTTGGTGGCC ACGGGCACGCAGAAGAGCGCCGCGATCCTCTTAAGCAC AACCAATTAACCAATTCTGA | UL24 mutation L253AF254A Reinsertion of wild type UL24 sequence |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, C.E.; Ben Abdeljelil, N.; Pearson, A. The Disruption of a Nuclear Export Signal in the C-Terminus of the Herpes Simplex Virus 1 Determinant of Pathogenicity UL24 Protein Leads to a Syncytial Plaque Phenotype. Viruses 2023, 15, 1971. https://doi.org/10.3390/v15091971
Gonzalez CE, Ben Abdeljelil N, Pearson A. The Disruption of a Nuclear Export Signal in the C-Terminus of the Herpes Simplex Virus 1 Determinant of Pathogenicity UL24 Protein Leads to a Syncytial Plaque Phenotype. Viruses. 2023; 15(9):1971. https://doi.org/10.3390/v15091971
Chicago/Turabian StyleGonzalez, Carmen Elena, Nawel Ben Abdeljelil, and Angela Pearson. 2023. "The Disruption of a Nuclear Export Signal in the C-Terminus of the Herpes Simplex Virus 1 Determinant of Pathogenicity UL24 Protein Leads to a Syncytial Plaque Phenotype" Viruses 15, no. 9: 1971. https://doi.org/10.3390/v15091971