

Dengue Exposure and Wolbachia wMel Strain Affects the Fertility of Quiescent Eggs of Aedes aegypti
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. Dengue Virus
2.3. Experimental Virus Infection
2.4. Detection and Quantification of DENV-1
2.5. Detection and Quantification of Wolbachia
2.6. Impact of DENV-1 and Wolbachia on Mosquito Survival, Fecundity and Oviposition Success
2.7. Impact of DENV-1 and Wolbachia on the Fertility of Quiescent Eggs
2.8. Statistical Analysis
3. Results
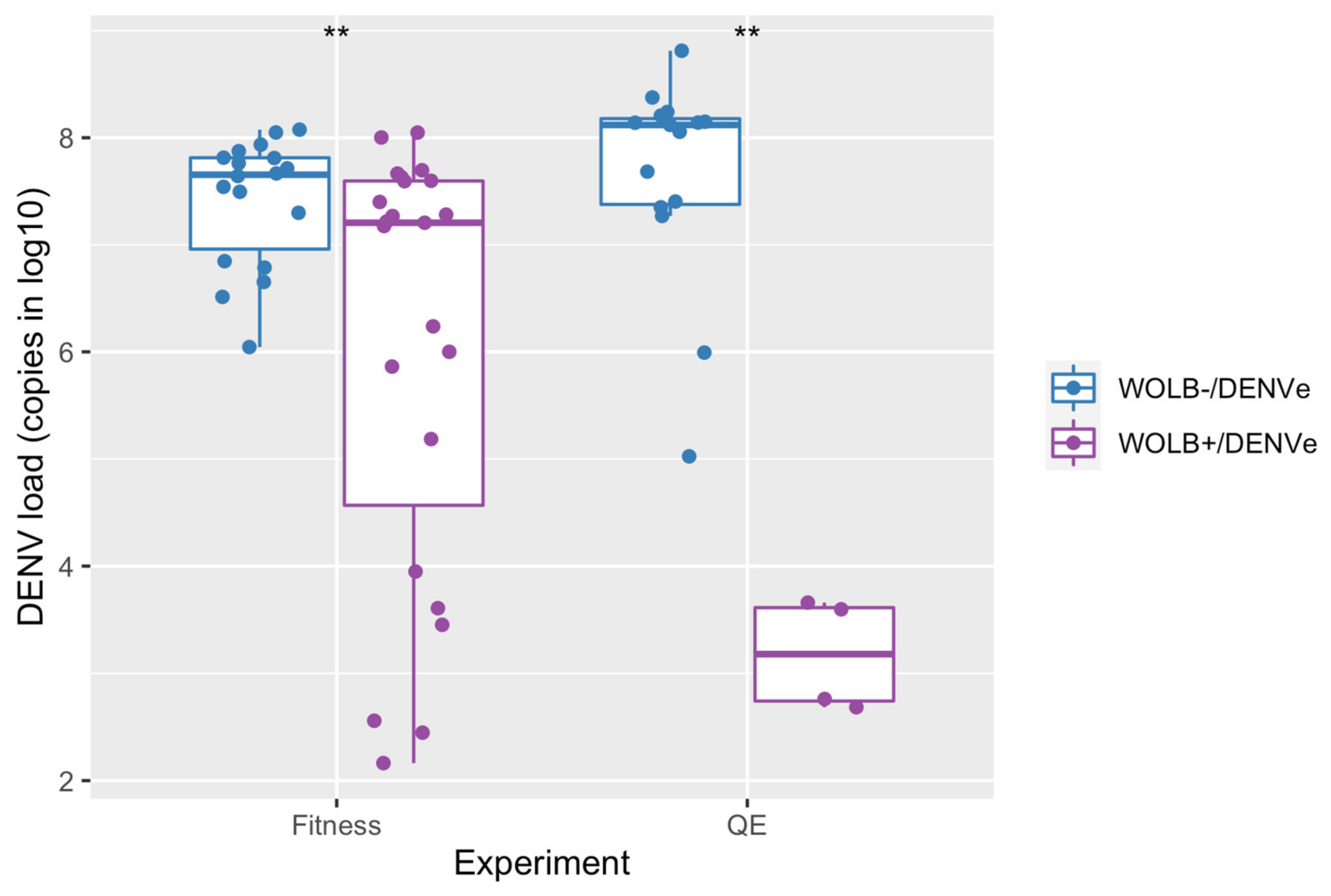
3.1. DENV-1 Load Was Lower in WOLB+ Than in WOLB− Aedes aegypti
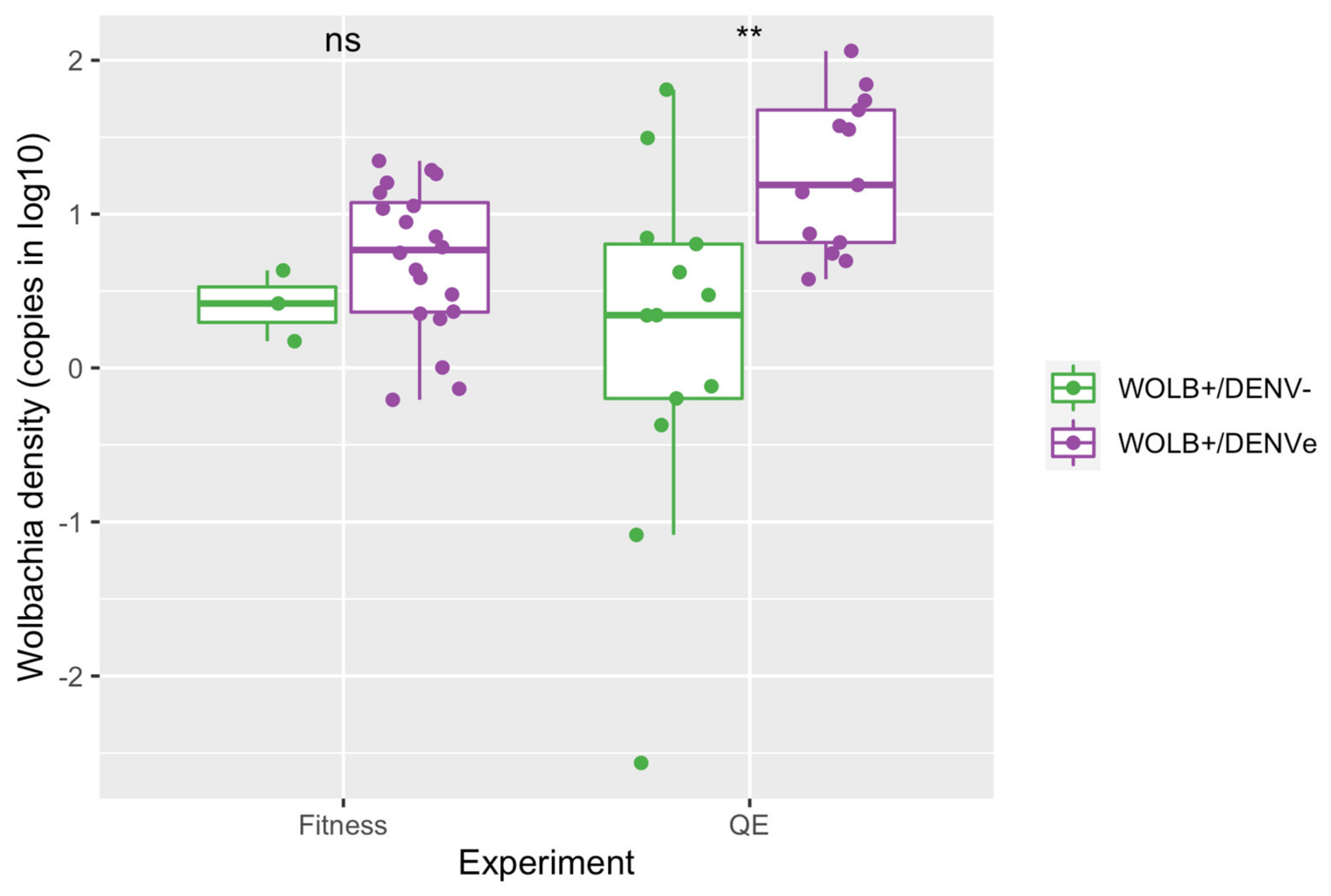
3.2. Wolbachia Density Was Higher in DENVe than in DENV− Mosquitoes from Quiescent Eggs Experiment
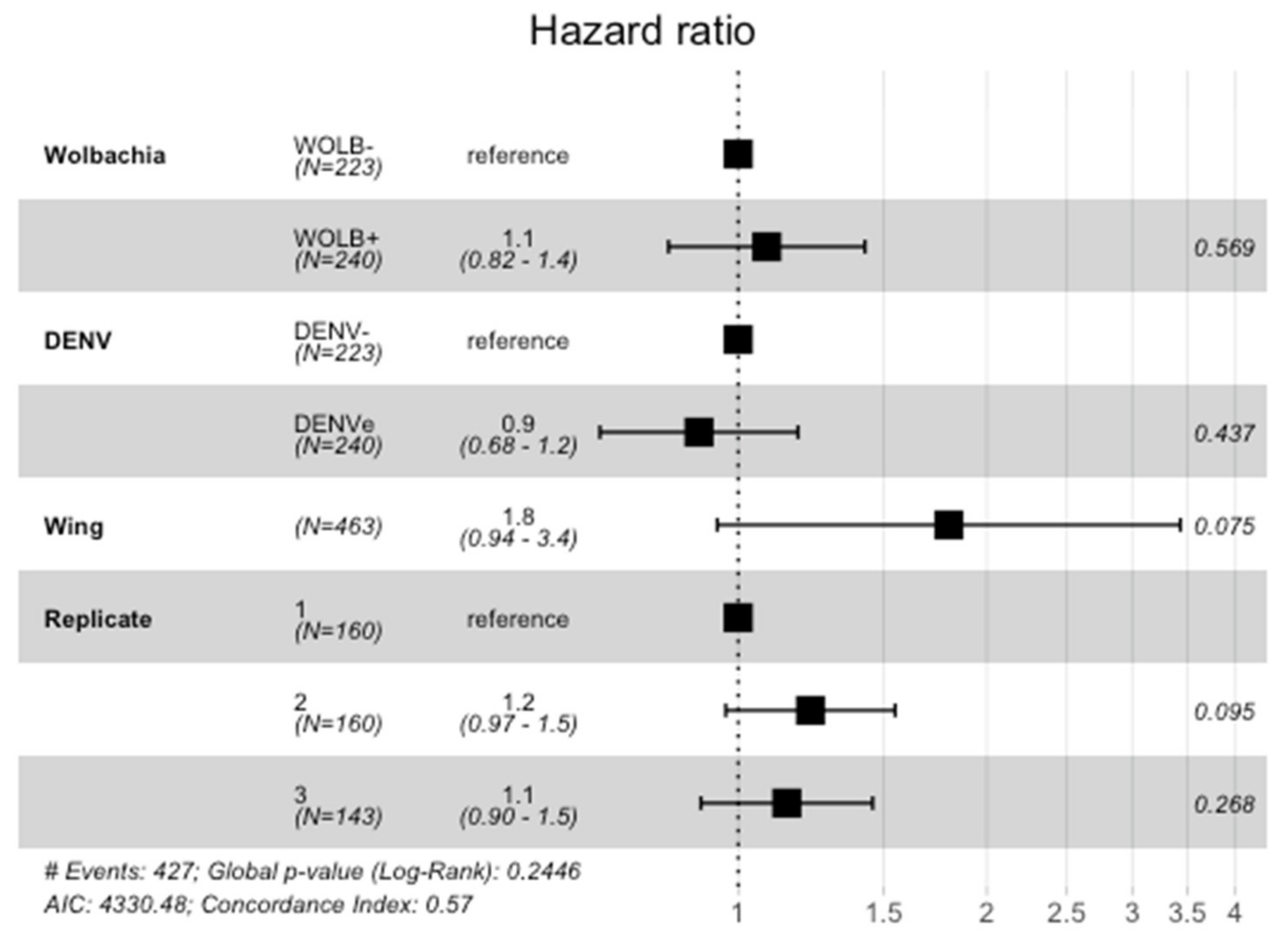
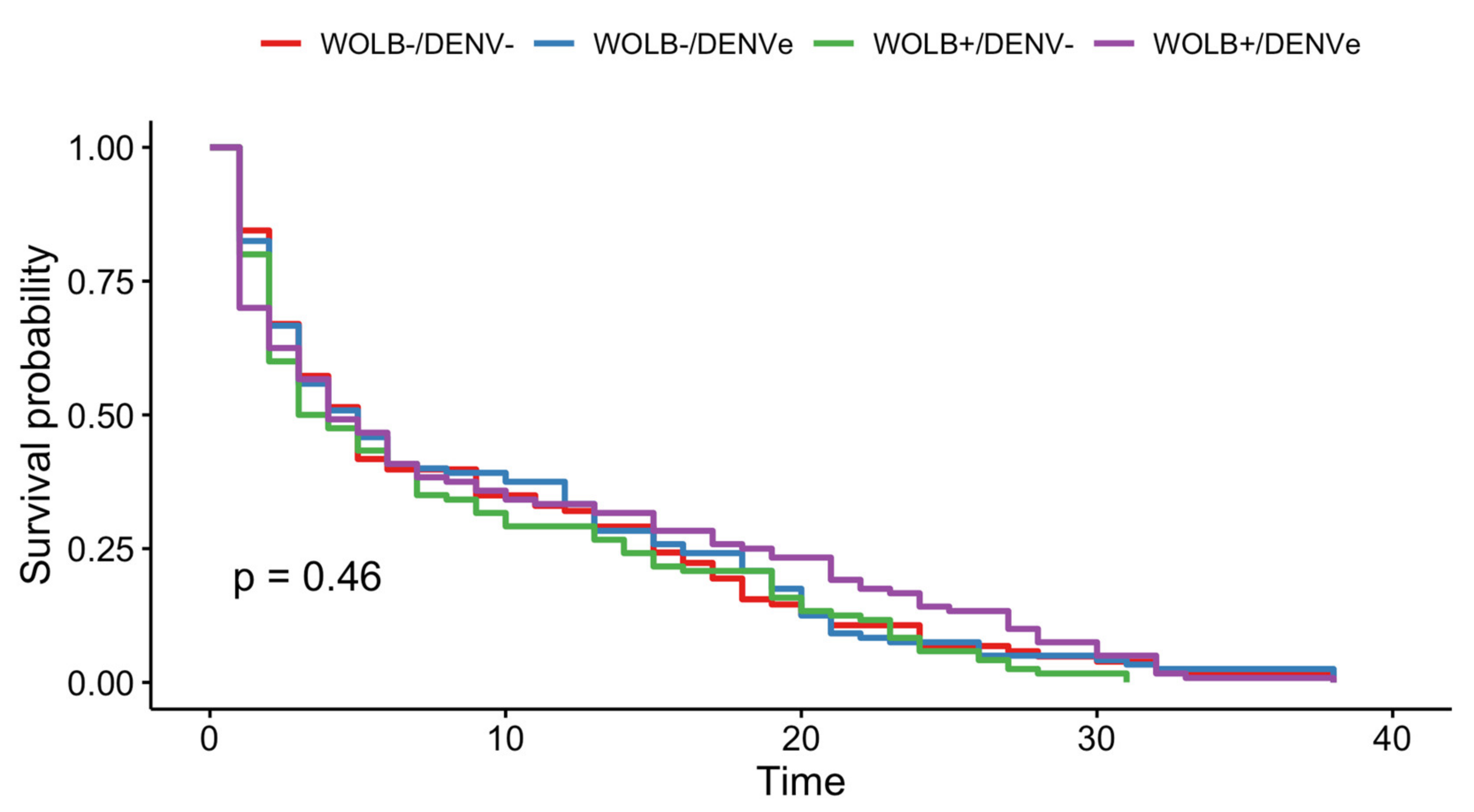
3.3. Wolbachia and DENV-1 Had No Effect on Mosquito Survival
3.4. Oviposition Success Decreased over Time
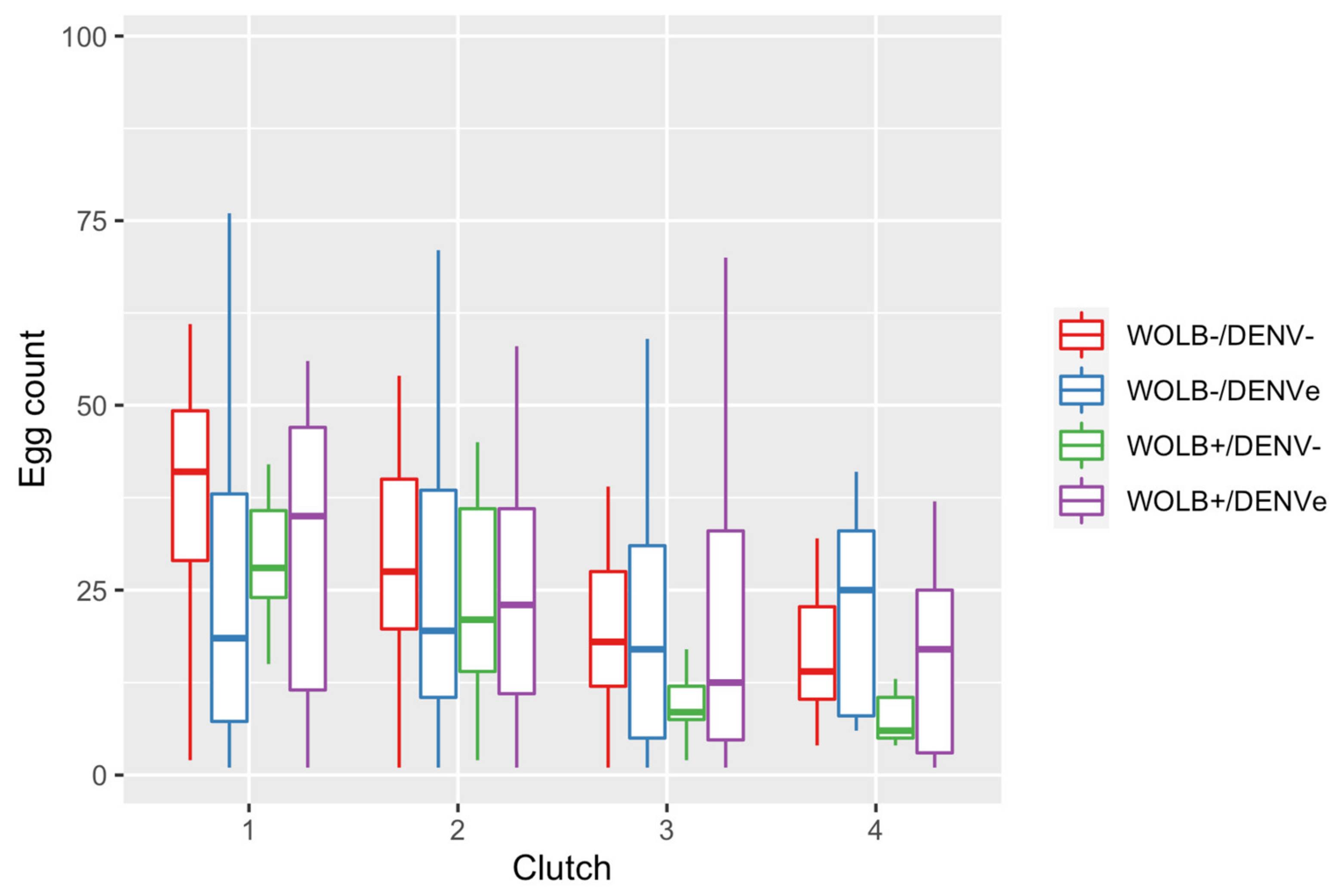
3.5. Fecundity Decreased with Ageing
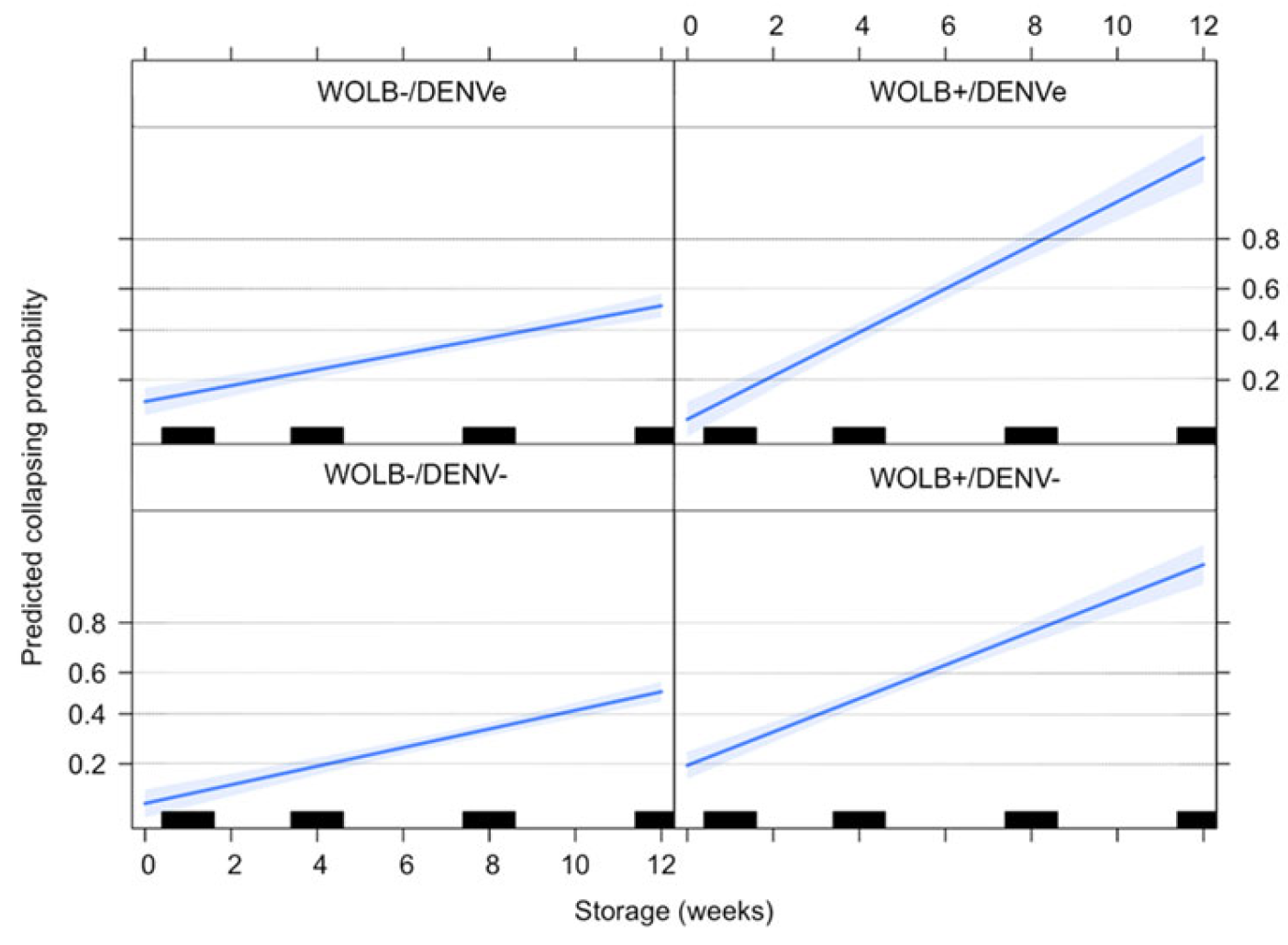
3.6. Quiescent Egg Viability Depends on Wolbachia Infection but Not on DENV Exposure
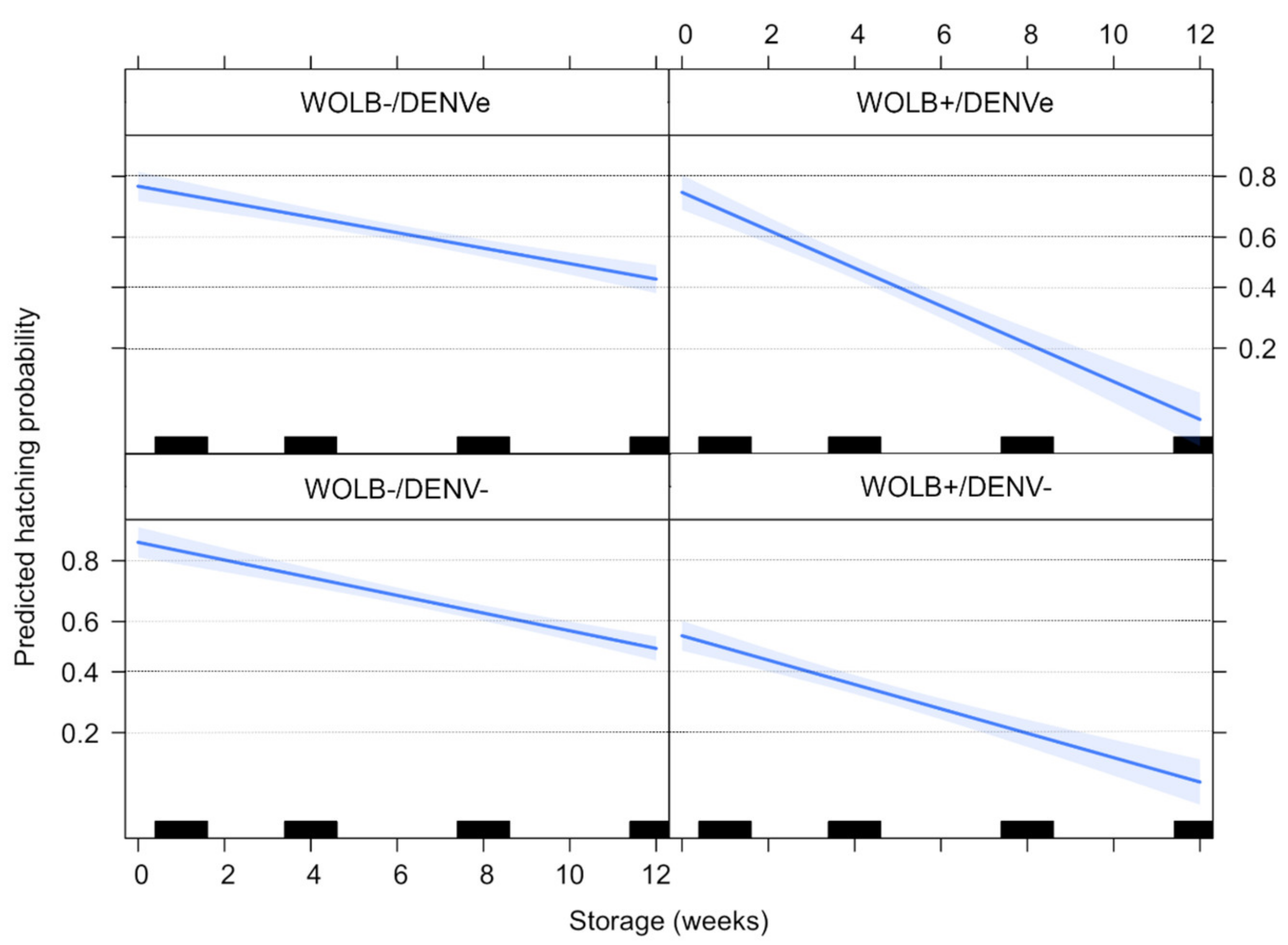
3.7. Quiescent Eggs Show a Decrease in Fertility, Especially on Wolbachia-Infected Groups
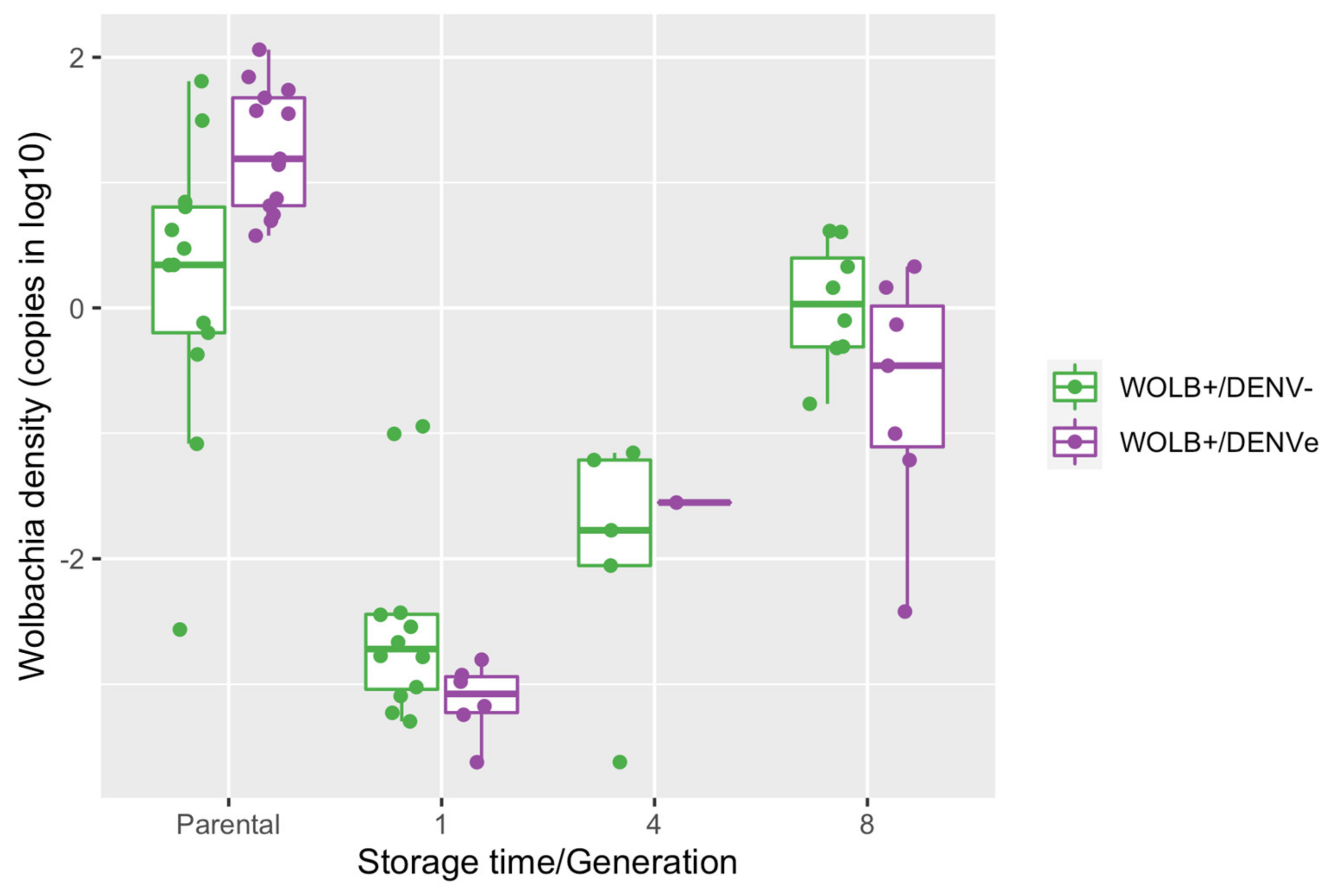
3.8. Wolbachia Density in Adult Females Reared from Quiescent Eggs Increased with Egg Storage Time
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How Aedes aegypti Mosquitoes Enabled the First Pandemic Arbovirus. Annu. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Reisen, W.K. Present and Future Arboviral Threats. Antiviral Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J. Dengue and Dengue Hemorrhagic Fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffner, F.; Mathis, A. Dengue and Dengue Vectors in the WHO European Region: Past, Present, and Scenarios for the Future. Lancet Infect. Dis. 2014, 14, 1271–1280. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The Global Distribution of the Arbovirus Vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Marques, W.A.; Peres, R.C.; Cunha, S.P.; Lourenço-de-Oliveira, R. Variation in Aedes aegypti (Diptera: Culicidae) Container Productivity in a Slum and a Suburban District of Rio de Janeiro during Dry and Wet Seasons. Mem. Inst. Oswaldo Cruz 2007, 102, 489–496. [Google Scholar] [CrossRef] [Green Version]
- David, M.R.; Dantas, E.S.; Maciel-de-Freitas, R.; Codeço, C.T.; Prast, A.E.; Lourenço-de-Oliveira, R. Influence of Larval Habitat Environmental Characteristics on Culicidae Immature Abundance and Body Size of Adult Aedes aegypti. Front. Ecol. Evol. 2021, 9, 626757. [Google Scholar] [CrossRef]
- Tun-Lin, W.; Lenhart, A.; Nam, V.S.; Rebollar-Téllez, E.; Morrison, A.C.; Barbazan, P.; Cote, M.; Midega, J.; Sanchez, F.; Manrique-Saide, P.; et al. Reducing Costs and Operational Constraints of Dengue Vector Control by Targeting Productive Breeding Places: A Multi-Country Non-Inferiority Cluster Randomized Trial. Trop. Med. Int. Health 2009, 14, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- Maciel-de-Freitas, R.; Peres, R.C.; Souza-Santos, R.; Lourenço-de-Oliveira, R. Occurrence, Productivity and Spatial Distribution of Key-Premises in Two Dengue-Endemic Areas of Rio de Janeiro and Their Role in Adult Aedes aegypti Spatial Infestation Pattern. Trop. Med. Int. Health 2008, 13, 1488–1494. [Google Scholar] [CrossRef]
- Ferdousi, F.; Yoshimatsu, S.; Ma, E.; Sohel, N.; Wagatsuma, Y. Identification of Essential Containers for Aedes Larval Breeding to Control Dengue in Dhaka, Bangladesh. Trop. Med. Health 2015, 43, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Maciel-de-Freitas, R.; Avendanho, F.C.; Santos, R.; Sylvestre, G.; Araújo, S.C.; Lima, J.B.P.; Martins, A.J.; Coelho, G.E.; Valle, D. Undesirable Consequences of Insecticide Resistance Following Aedes aegypti Control Activities Due to a Dengue Outbreak. PLoS ONE 2014, 9, e92424. [Google Scholar] [CrossRef]
- Barrera, R.; Amador, M.; Clark, G.G. Ecological Factors Influencing Aedes aegypti (Diptera: Culicidae) Productivity in Artificial Containers in Salinas, Puerto Rico. J. Med. Entomol. 2006, 43, 484–492. [Google Scholar] [CrossRef] [Green Version]
- David, M.R.; Garcia, G.A.; Valle, D.; Maciel-de-Freitas, R. Insecticide Resistance and Fitness: The Case of Four Aedes aegypti Populations from Different Brazilian Regions. Biomed Res. Int. 2018, 2018, 6257860. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.d.A.; David, M.R.; de Martins, A.J.; Maciel-de-Freitas, R.; Linss, J.G.B.; Araújo, S.C.; Lima, J.B.P.; Valle, D. The Impact of Insecticide Applications on the Dynamics of Resistance: The Case of Four Aedes aegypti Populations from Different Brazilian Regions. PLoS Negl. Trop. Dis. 2018, 12, e0006227. [Google Scholar] [CrossRef] [Green Version]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.-P.; et al. Contemporary Status of Insecticide Resistance in the Major Aedes Vectors of Arboviruses Infecting Humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef]
- Lazaro, A.; Han, W.W.; Manrique-Saide, P.; George, L.; Velayudhan, R.; Toledo, J.; Runge Ranzinger, S.; Horstick, O. Community Effectiveness of Copepods for Dengue Vector Control: Systematic Review. Trop. Med. Int. Health 2015, 20, 685–706. [Google Scholar] [CrossRef]
- Morales-Pérez, A.; Nava-Aguilera, E.; Legorreta-Soberanis, J.; Cortés-Guzmán, A.J.; Balanzar-Martínez, A.; Harris, E.; Coloma, J.; Alvarado-Castro, V.M.; Bonilla-Leon, M.V.; Morales-Nava, L.; et al. “Where We Put Little Fish in the Water There Are No Mosquitoes:” A Cross-Sectional Study on Biological Control of the Aedes aegypti Vector in 90 Coastal-Region Communities of Guerrero, Mexico. BMC Public Health 2017, 17, 433. [Google Scholar] [CrossRef] [Green Version]
- Baldacchino, F.; Caputo, B.; Chandre, F.; Drago, A.; della Torre, A.; Montarsi, F.; Rizzoli, A. Control Methods against Invasive Aedes Mosquitoes in Europe: A Review. Pest Manag. Sci. 2015, 71, 1471–1485. [Google Scholar] [CrossRef] [Green Version]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-NG, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C.; et al. Alternative Strategies for Mosquito-Borne Arbovirus Control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar] [CrossRef] [Green Version]
- Beier, J.C.; Keating, J.; Githure, J.I.; Macdonald, M.B.; Impoinvil, D.E.; Novak, R.J. Integrated Vector Management for Malaria Control. Malar. J. 2008, 7, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leandro, A.d.S.; Lopes, R.D.; Martins, C.A.; Rivas, A.V.; da Silva, I.; Galvão, S.R.; Maciel-de-Freitas, R. The Adoption of the One Health Approach to Improve Surveillance of Venomous Animal Injury, Vector-Borne and Zoonotic Diseases in Foz Do Iguaçu, Brazil. PLoS Negl. Trop. Dis. 2021, 15, e0009109. [Google Scholar] [CrossRef] [PubMed]
- Flores, H.A.; O’Neill, S.L. Controlling Vector-Borne Diseases by Releasing Modified Mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- McGraw, E.A.; O’Neill, S.L. Beyond Insecticides: New Thinking on an Ancient Problem. Nat. Rev. Microbiol. 2013, 11, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How Many Species Are Infected with Wolbachia?—A Statistical Analysis of Current Data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Aliota, M.T.; Walker, E.C.; Uribe Yepes, A.; Dario Velez, I.; Christensen, B.M.; Osorio, J.E. The WMel Strain of Wolbachia Reduces Transmission of Chikungunya Virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2016, 10, e0004677. [Google Scholar] [CrossRef] [Green Version]
- Dutra, H.L.C.; Rocha, M.N.; Dias, F.B.S.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia Blocks Currently Circulating Zika Virus Isolates in Brazilian Aedes aegypti Mosquitoes. Cell Host Microbe 2016, 19, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The WMel Wolbachia Strain Blocks Dengue and Invades Caged Aedes aegypti Populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Garcia, G.d.A.; Sylvestre, G.; Aguiar, R.; da Costa, G.B.; Martins, A.J.; Lima, J.B.P.; Petersen, M.T.; Lourenço-de-Oliveira, R.; Shadbolt, M.F.; Rašić, G.; et al. Matching the Genetics of Released and Local Aedes aegypti Populations Is Critical to Assure Wolbachia Invasion. PLoS Negl. Trop. Dis. 2019, 13, e0007023. [Google Scholar] [CrossRef]
- Lau, M.-J.; Ross, P.A.; Hoffmann, A.A. Infertility and Fecundity Loss of Wolbachia-Infected Aedes aegypti Hatched from Quiescent Eggs Is Expected to Alter Invasion Dynamics. PLoS Negl. Trop. Dis. 2021, 15, e0009179. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Koella, J.C.; Lourenço-de-Oliveira, R. Lower Survival Rate, Longevity and Fecundity of Aedes aegypti (Diptera: Culicidae) Females Orally Challenged with Dengue Virus Serotype 2. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 452–458. [Google Scholar] [CrossRef]
- Da Silveira, I.D.; Petersen, M.T.; Sylvestre, G.; Garcia, G.A.; David, M.R.; Pavan, M.G.; Maciel-de-Freitas, R. Zika Virus Infection Produces a Reduction on Aedes aegypti Lifespan but No Effects on Mosquito Fecundity and Oviposition Success. Front. Microbiol. 2018, 9, 3011. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.T.; Silveira, I.D.d.; Tátila-Ferreira, A.; David, M.R.; Chouin-Carneiro, T.; Van den Wouwer, L.; Maes, L.; Maciel-de-Freitas, R. The Impact of the Age of First Blood Meal and Zika Virus Infection on Aedes aegypti Egg Production and Longevity. PLoS ONE 2018, 13, e0200766. [Google Scholar] [CrossRef]
- Maciel-de-Freitas, R.; Sylvestre, G.; Gandini, M.; Koella, J.C. The Influence of Dengue Virus Serotype-2 Infection on Aedes aegypti (Diptera: Culicidae) Motivation and Avidity to Blood Feed. PLoS ONE 2013, 8, e65252. [Google Scholar] [CrossRef]
- Sylvestre, G.; Gandini, M.; Maciel-de-Freitas, R. Age-Dependent Effects of Oral Infection with Dengue Virus on Aedes aegypti (Diptera: Culicidae) Feeding Behavior, Survival, Oviposition Success and Fecundity. PLoS ONE 2013, 8, e59933. [Google Scholar] [CrossRef]
- De Jesus, C.P.; Dias, F.B.S.; Villela, D.M.A.; Maciel-de-Freitas, R. Ovitraps Provide a Reliable Estimate of Wolbachia Frequency during WMelBr Strain Deployment in a Geographically Isolated Aedes aegypti Population. Insects 2020, 11, 92. [Google Scholar] [CrossRef] [Green Version]
- Souto-Maior, C.; Sylvestre, G.; Braga Stehling Dias, F.; Gomes, M.G.M.; Maciel-de-Freitas, R. Model-Based Inference from Multiple Dose, Time Course Data Reveals Wolbachia Effects on Infection Profiles of Type 1 Dengue Virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006339. [Google Scholar] [CrossRef]
- Caragata, E.; Dutra, H.; Moreira, L. Inhibition of Zika Virus by Wolbachia in Aedes aegypti. Microb. Cell 2016, 3, 293–295. [Google Scholar] [CrossRef]
- Harbach, R.E.; Knight, K.L. Taxonomist’s Glossary of Mosquito Anatomy; Plexus Publications Co.: Marlton, NJ, USA, 1980; pp. 1–54. ISBN 978-0937548004. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2020. Available online: https://www.R-project.org/ (accessed on 11 March 2023).
- Fox, J. Effect Displays in R for Generalised Linear Models. J. Stat. Softw. 2003, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ryan, P.A.; Turley, A.P.; Wilson, G.; Hurst, T.P.; Retzki, K.; Brown-kenyon, J.; Hodgson, L.; Kenny, N.; Cook, H.; Montgomery, B.L.; et al. Establishment of w Mel Wolbachia in Aedes aegypti Mosquitoes and Reduction of Local Dengue Transmission in Cairns and Surrounding Locations in Northern Queensland, Australia. Gates Open Res. 2019, 3, 1547. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Nguyen, H.L.; Nguyen, T.Y.; Vu, S.N.; Tran, N.D.; Le, T.N.; Vien, Q.M.; Bui, T.C.; Le, H.T.; Kutcher, S.; et al. Field Evaluation of the Establishment Potential of Wmelpop Wolbachia in Australia and Vietnam for Dengue Control. Parasit. Vectors 2015, 8, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, G.A.; Hoffmann, A.A.; Maciel-de-Freitas, R.; Villela, D.A.M. Aedes aegypti Insecticide Resistance Underlies the Success (and Failure) of Wolbachia Population Replacement. Sci. Rep. 2020, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Javed, N.; Bhatti, A.; Paradkar, P.N. Advances in Understanding Vector Behavioural Traits after Infection. Pathogens 2021, 10, 1376. [Google Scholar] [CrossRef]
- Garcia, G.A.; Lord, A.R.; Santos, L.M.B.; Kariyawasam, T.N.; David, M.R.; Couto-Lima, D.; Tátila-Ferreira, A.; Pavan, M.G.; Sikulu-Lord, M.T.; Maciel-de-Freitas, R. Rapid and Non-Invasive Detection of Aedes aegypti Co-Infected with Zika and Dengue Viruses Using Near Infrared Spectroscopy. Viruses 2022, 15, 11. [Google Scholar] [CrossRef]
- Lu, P.; Sun, Q.; Fu, P.; Li, K.; Liang, X.; Xi, Z. Wolbachia Inhibits Binding of Dengue and Zika Viruses to Mosquito Cells. Front. Microbiol. 2020, 11, 1750. [Google Scholar] [CrossRef]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia Induces Density-Dependent Inhibition to Dengue Virus in Mosquito Cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.I.L.; Suzuki, Y.; Carvajal, T.; Muñoz, M.N.M.; Watanabe, K. Intracellular Interactions Between Arboviruses and Wolbachia in Aedes aegypti. Front. Cell. Infect. Microbiol. 2021, 11, 690087. [Google Scholar] [CrossRef]
- Novelo, M.; Audsley, M.D.; McGraw, E.A. The Effects of DENV Serotype Competition and Co-Infection on Viral Kinetics in Wolbachia-Infected and Uninfected Aedes aegypti Mosquitoes. Parasit. Vectors 2021, 14, 314. [Google Scholar] [CrossRef]
- Ribeiro dos Santos, G.; Durovni, B.; Saraceni, V.; Souza Riback, T.I.; Pinto, S.B.; Anders, K.L.; Moreira, L.A.; Salje, H. Estimating the Effect of the WMel Release Programme on the Incidence of Dengue and Chikungunya in Rio de Janeiro, Brazil: A Spatiotemporal Modelling Study. Lancet Infect. Dis. 2022, 22, 1587–1595. [Google Scholar] [CrossRef]
- Gesto, J.S.M.; Pinto, S.B.; Dias, F.B.S.; Peixoto, J.; Costa, G.; Kutcher, S.; Montgomery, J.; Green, B.R.; Anders, K.L.; Ryan, P.A.; et al. Large-Scale Deployment and Establishment of Wolbachia into the Aedes aegypti Population in Rio de Janeiro, Brazil. Front. Microbiol. 2021, 12, 711107. [Google Scholar] [CrossRef]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic Specificity and Potential for Local Adaptation between Dengue Viruses and Mosquito Vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Farnesi, L.C.; Belinato, T.A.; Gesto, J.S.M.; Martins, A.J.; Bruno, R.V.; Moreira, L.A. Embryonic Development and Egg Viability of WMel-Infected Aedes aegypti. Parasit. Vectors 2019, 12, 211. [Google Scholar] [CrossRef]
- Caragata, E.P.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Competition for Amino Acids Between Wolbachia and the Mosquito Host, Aedes aegypti. Microb. Ecol. 2014, 67, 205–218. [Google Scholar] [CrossRef]
- Farnesi, L.C.; Menna-Barreto, R.F.S.; Martins, A.J.; Valle, D.; Rezende, G.L. Physical Features and Chitin Content of Eggs from the Mosquito Vectors Aedes aegypti, Anopheles Aquasalis and Culex Quinquefasciatus: Connection with Distinct Levels of Resistance to Desiccation. J. Insect Physiol. 2015, 83, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Embryonic Development of Aedes aegypti (Diptera: Culicidae): Influence of Different Constant Temperatures. Mem. Inst. Oswaldo Cruz 2009, 104, 124–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allman, M.J.; Lin, Y.-H.; Joubert, D.A.; Addley-Cook, J.; Mejía-Torres, M.C.; Simmons, C.P.; Flores, H.A.; Fraser, J.E. Enhancing the Scalability of Wolbachia-Based Vector-Borne Disease Management: Time and Temperature Limits for Storage and Transport of Wolbachia-Infected Aedes aegypti Eggs for Field Releases. Parasit. Vectors 2023, 16, 108. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | d.f. | χ2 | p Value |
|---|---|---|---|
| Wolbachia | 1 | 1.691 | 0.1933 |
| DENV exposure | 1 | 0.566 | 0.4515 |
| Clutch | 3 | 14.262 | 0.0026 * |
| Wing | 1 | 0.317 | 0.5732 |
| Wolbachia vs. DENV exposure | 1 | 0.038 | 0.8436 |
| Wolbachia vs. Clutch | 3 | 15.337 | 0.0016 * |
| DENV exposure vs. Clutch | 3 | 2.430 | 0.4880 |
| Source | Numerator df | Denominator df | F | p-Value |
|---|---|---|---|---|
| Clutch vs. Wolbachia | 1 | 66 | 0.6369 | 0.4277 |
| Clutch vs. DENV | 1 | 66 | 0.7470 | 0.3906 |
| Clutch vs. Wing | 1 | 66 | 0.4748 | 0.4932 |
| Clutch vs. Wolbachia vs. DENV | 1 | 66 | 1.6259 | 0.2067 |
| Variable | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|
| Intercept | −2.17 | 0.1400 | −15.48 | <0.001 * |
| Wolbachia (WOLB+) | 0.7464 | 0.1958 | 3.811 | <0.001 * |
| DENV (DENVe) | 0.3533 | 0.1957 | 1.805 | 0.0711 |
| Storage | 0.1829 | 0.0165 | 11.03 | <0.001 * |
| Wolbachia vs. DENV | −1.0970 | 0.2964 | −3.701 | <0.001 * |
| Wolbachia vs. Storage | 0.1459 | 0.0294 | 4.953 | <0.001 * |
| DENV vs. Storage | −0.0260 | 0.0242 | −1.075 | 0.2825 |
| Wolbachia vs. DENV vs. Storage | 0.1251 | 0.0460 | 2.719 | 0.0060 * |
| Variable | Estimate | Std. Error | z Value | p-Value |
|---|---|---|---|---|
| Intercept | 1.68 | 0.12 | 13.52 | <0.001 |
| Wolbachia (WOLB+) | −1.51 | 0.17 | −8.60 | <0.001 * |
| DENV (DENVe) | −0.46 | 0.17 | −2.64 | 0.008 * |
| Storage time | −0.14 | 0.01 | −9.28 | <0.001 * |
| Wolbachia vs. DENV | 1.41 | 0.26 | 5.49 | <0.001 * |
| Wolbachia vs. Storage time | −0.05 | 0.027 | −2.00 | 0.0452 * |
| DENV vs. Storage time | 0.02 | 0.02 | 0.80 | 0.4261 |
| Wolbachia vs. DENV vs. Storage | −0.13 | 0.04 | −3.08 | 0.002 * |
| Variable | Estimate | Std. Error | t Value | p-Value |
|---|---|---|---|---|
| Intercept | −2.98 | 0.24 | −12.44 | <0.001 * |
| DENV (DENVe) | −0.44 | 0.42 | −1.06 | 0.30 |
| Storage time | 0.36 | 0.05 | 7.42 | <0.001 * |
| DENV * Storage time | −0.01 | 0.08 | −0.19 | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petersen, M.T.; Couto-Lima, D.; Garcia, G.A.; Pavan, M.G.; David, M.R.; Maciel-de-Freitas, R. Dengue Exposure and Wolbachia wMel Strain Affects the Fertility of Quiescent Eggs of Aedes aegypti. Viruses 2023, 15, 952. https://doi.org/10.3390/v15040952
Petersen MT, Couto-Lima D, Garcia GA, Pavan MG, David MR, Maciel-de-Freitas R. Dengue Exposure and Wolbachia wMel Strain Affects the Fertility of Quiescent Eggs of Aedes aegypti. Viruses. 2023; 15(4):952. https://doi.org/10.3390/v15040952
Chicago/Turabian StylePetersen, Martha Thieme, Dinair Couto-Lima, Gabriela Azambuja Garcia, Márcio Galvão Pavan, Mariana Rocha David, and Rafael Maciel-de-Freitas. 2023. "Dengue Exposure and Wolbachia wMel Strain Affects the Fertility of Quiescent Eggs of Aedes aegypti" Viruses 15, no. 4: 952. https://doi.org/10.3390/v15040952