
First Isolation and Identification of Homologous Recombination Events of Porcine Adenovirus from Wild Boar
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation from Fecal Samples of Wild Boar
2.2. Construction of DNA and cDNA Libraries and Deep Sequencing
2.3. Genome Analysis
2.4. Identification of PAdVs from Fecal Samples of Wild Boar and Pigs Using PCR
3. Results
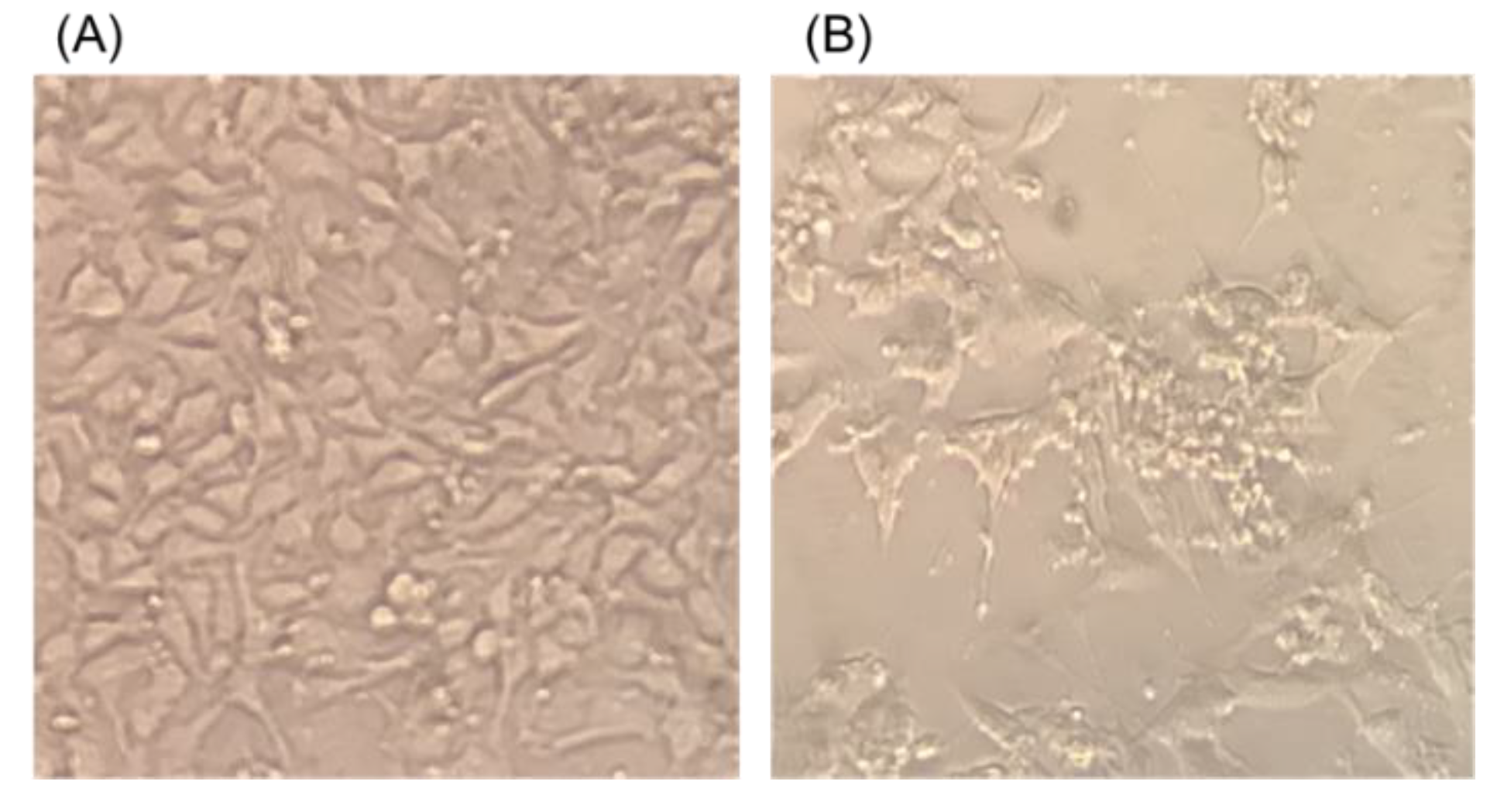
3.1. Isolation and Identification of Porcine Adenovirus
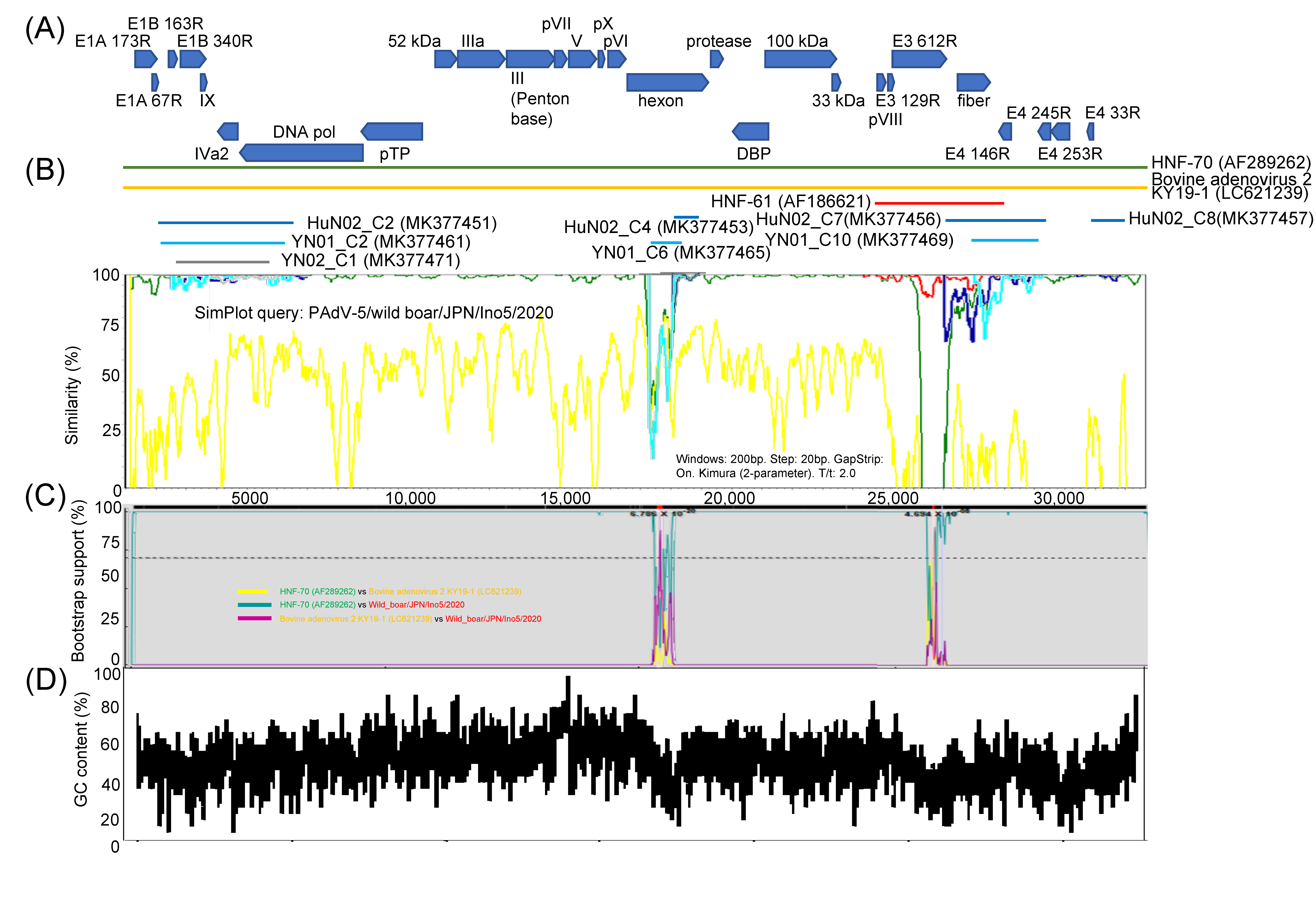
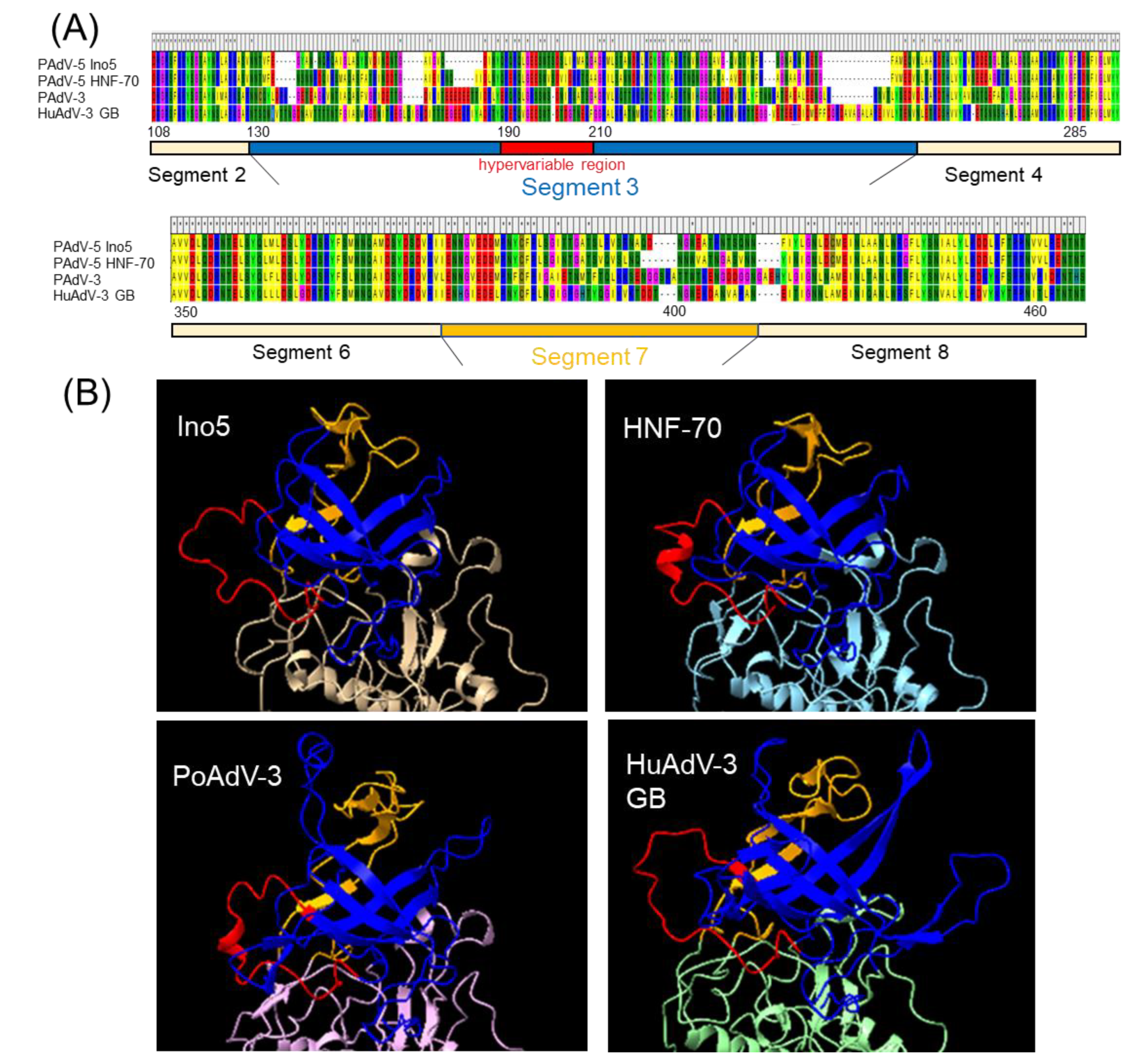
3.2. Genome Analysis and Structural Modeling of the Hexon and Fiber Proteins
3.3. Prevalence of PAdV in Wild Boars and Pigs, and Phylogenetic Analyses of the Hexon and Fiber Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. Ictv Report Consortium. ICDTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.M.; Johnson, M.A. Porcine adenovirus as a delivery system for swine vaccines and immunotherapeutics. Vet. J. 2005, 169, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Haig, D.A.; Clarke, M.C.; Pereira, M.S. Isolation of an adenovirus from a pig. J. Comp. Path. 1964, 74, 81–84. [Google Scholar] [CrossRef]
- Clarke, M.C.; Sharpe, H.B.; Derbyshire, J.B. Some characteristics of three porcine adenoviruses. Arch. Gesamte. Virusforsch. 1967, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kasza, L. Isolation of an adenovirus from the brain of a pig. Am. J. Vet. Res. 1966, 27, 751–758. [Google Scholar] [PubMed]
- Hirahara, T.; Yasuhara, H.; Matsui, O.; Yamanaka, M.; Tanaka, M.; Fukuyama, S.; Izumida, A.; Yoshiki, K.; Kodama, K.; Nakai, M.; et al. Isolation of porcine adenovirus from the respiratory tract of pigs in Japan. Nihon Juigaku Zasshi 1990, 52, 407–409. [Google Scholar] [CrossRef]
- Kadoi, K.; Inoue, Y.; Ikeda, T.; Kamata, H.; Yukawa, M.; Iwabuchi, M.; Inaba, Y. Isolation of porcine adenovirus as a candidate of 5th serotype. J. Basic. Microbiol. 1995, 35, 195–204. [Google Scholar] [CrossRef]
- Jerman, U.D.; Kolenc, M.; Steyer, A.; Veranič, P.; Prijatelj, M.P.; Kreft, M.E. A novel strain of porcine adenovirus detected in urinary bladder urothelial cell culture. Viruses 2014, 23, 2505–2518. [Google Scholar] [CrossRef] [Green Version]
- Bigoraj, E.; Paszkiewicz, W.; Rzeżutka, A. Porcine blood and liver as sporadic sources of Hepatitis E virus (HEV) in the production chain of offal-derived foodstuffs in Poland. Food Environ. Virol. 2021, 13, 347–356. [Google Scholar] [CrossRef]
- Elazhary, M.A.; Dea, S.; Mittal, K.R.; Higgins, R. Prevalence of antibodies to swine influenza virus, porcine adenovirus type 4 and haemophilus pleuropneumoniae in Quebec pig farms with respiratory problems. Can. Vet. J. 1985, 26, 190–192. [Google Scholar]
- Hirahara, T.; Yasuhara, H.; Matsui, O.; Fukuyama, S.; Yamanaka, M.; Izumida, A.; Yoshiki, K.; Kodama, K.; Nakai, M.; Sasaki, N. Serological survey on porcine adenovirus infection of pigs in Japan. J. Jpn. Vet. Med. Assoc. 1990, 43, 779–783. (In Japanese) [Google Scholar] [CrossRef]
- Kumthip, K.; Khamrin, P.; Kongkaew, A.; Vachirachewin, R.; Malasao, R.; Ushijima, H.; Maneekarn, N. Molecular epidemiology and characterization of porcine adenoviruses in pigs with diarrhea in Thailand. Infect. Genet. Evol. 2019, 67, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Nagy, M.; Nagy, É.; Tuboly, T. The complete nucleotide sequence of porcine adenovirus serotype 5. J. Gen. Virol. 2001, 82, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Aoki, H.; Sakoda, Y.; Kozasa, T.; Tominaga-Teshima, K.; Mine, J.; Abe, Y.; Tamura, T.; Kobayashi, T.; Nishine, K.; et al. Molecular, biological, and antigenic characterization of a Border disease virus isolated from a pig during classical swine fever surveillance in Japan. J. Vet. Diagn. Investig. 2014, 26, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Komaniwa, H.; Fukusho, A.; Shimizu, Y. Micro method for performing titration and neutralization test of hog cholera virus using established porcine kidney cell strain. Natl. Inst. Anim. Health. Q 1981, 21, 153–158. [Google Scholar]
- Suzuki, T.; Murakami, S.; Takahashi, O.; Kodera, A.; Masuda, T.; Itoh, S.; Miyazaki, A.; Ohashi, S.; Tsutsui, T. Molecular characterization of pig epidemic diarrhoea viruses isolated in Japan from 2013 to 2014. Infect. Genet. Evol. 2015, 36, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic. Acids. Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus. Evol. 2015, 26, vev003. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Wellehan, J.F.; Johnson, A.J.; Harrach, B.; Benkö, M.; Pessier, A.P.; Johnson, C.M.; Garner, M.M.; Childress, A.; Jacobson, E.R. Detection and analysis of six lizard adenoviruses by consensus primer PCR provides further evidence of a reptilian origin for the atadenoviruses. J. Virol. 2004, 78, 13366–13369. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Z.; Shang, L.; Gao, H.; Du, X.; Li, F.; Gao, Y.; Qi, G.; Guo, W.; Qu, Z.; et al. Immunological Study of Reconstructed Common Ancestral Sequence of Adenovirus Hexon Protein. Front. Microbiol. 2021, 27, 717047. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Mortlock, M.; Weyer, J.; Bezuidt, O.; Seamark, E.C.J.; Kearney, T.; Gleasner, C.; Erkkila, T.H.; Cui, H.; Markotter, W. A metagenomic viral discovery approach identifies potential zoonotic and novel mammalian viruses in Neoromicia bats within South Africa. PloS. One. 2018, 26, e0194527. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 3, 178. [Google Scholar] [CrossRef]
- Nietfeld, J.C.; Leslie-Steen, P. Interstitial nephritis in pigs with adenovirus infection. J. Vet. Diagn. Invest. 1993, 5, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Lee, J.S.; Lee, J.Y.; Singh, G.; Dyer, D.W.; Seto, D.; Chodosh, J.; Rajaiya, J. Adenoviromics: Mining the Human Adenovirus Species D Genome. Front. Microbiol. 2018, 11, 2178. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Padilla, J.; Papp, T.; Kaján, G.L.; Benkő, M.; Havenga, M.; Lemckert, A.; Harrach, B.; Baker, A.H. Development of novel adenoviral vectors to overcome challenges observed With HAdV-5-based constructs. Mol. Ther. 2016, 24, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, F.; Tachibana, M.; Mizuguchi, H. Adenovirus vector-based vaccine for infectious diseases. Drug Metab. Pharmacokinet. 2022, 42, 100432. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nishikawaji, Y.; Kawakami, H.; Kosai, K.I. Adenovirus biology, recombinant adenovirus, and adenovirus usage in gene therapy. Viruses 2021, 14, 2502. [Google Scholar] [CrossRef]
- Walsh, M.P.; Chintakuntlawar, A.; Robinson, C.M.; Madisch, I.; Harrach, B.; Hudson, N.R.; Schnurr, D.; Heim, A.; Chodosh, J.; Seto, D.; et al. Evidence of molecular evolution driven by recombination events influencing tropism in a novel human adenovirus that causes epidemic keratoconjunctivitis. PLoS ONE 2009, 4, e5635. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, A.; Hage, E.; Ganzenmueller, T.; Böttcher, S.; Hofmann, J.; Hamprecht, K.; Obermeier, P.; Rath, B.; Hausmann, F.; Dobner, T.; et al. Molecular Evolution of Human Adenovirus (HAdV) Species C. Sci. Rep. 2018, 31, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Qian, Y.; Jia, L.; Dong, H.; Deng, L.; Huang, H.; Zhao, L.; Zhu, R. Genetic diversity and molecular evolution of human adenovirus serotype 41 strains circulating in Beijing, China, during 2010-2019. Infect. Genet. Evol. 2021, 95, 105056. [Google Scholar] [CrossRef]
- Horwitz, M.S. Function of adenovirus E3 proteins and their interactions with immunoregulatory cell proteins. J. Gene. Med. 2004, 6 (Suppl. 1), S172–S183. [Google Scholar] [CrossRef]
- Tuboly, T.; Nagy, E. Sequence analysis and deletion of porcine adenovirus serotype 5 E3 region. Virus Res. 2000, 68, 109–117. [Google Scholar] [CrossRef]
- Sohaimi, N.M.; Hair-Bejo, M. A recent perspective on fiber and hexon genes proteins analyses of fowl adenovirus toward virus infectivity-A review. Open Vet. J. 2021, 11, 569–580. [Google Scholar] [CrossRef]
- Lukashev, A.N.; Ivanova, O.E.; Eremeeva, T.P.; Iggo, R.D. Evidence of frequent recombination among human adenoviruses. J. Gen. Virol. 2008, 89, 380–388. [Google Scholar] [CrossRef]
- Homonnay, Z.; Jakab, S.; Bali, K.; Kaszab, E.; Mató, T.; Kiss, I.; Palya, V.; Bányai, K. Genome sequencing of a novel variant of fowl adenovirus B reveals mosaicism in the pattern of homologous recombination events. Arch. Virol. 2021, 166, 1477–1480. [Google Scholar] [CrossRef]
- Singh, G.; Zhou, X.; Lee, J.Y.; Yousuf, M.A.; Ramke, M.; Ismail, A.M.; Lee, J.S.; Robinson, C.M.; Seto, D.; Dyer, D.W.; et al. Recombination of the epsilon determinant and corneal tropism: Human adenovirus species D types 15, 29, 56, and 69. Virology 2015, 485, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Xu, B.; Li, C.; Bao, Y.; An, S.; Zhou, Y.; Chen, A.; Deng, L.; Ning, L.; Zhu, Y.; et al. Molecular Characteristics of Human Adenovirus Type 3 Circulating in Parts of China During 2014-2018. Front. Microbiol. 2021, 29, 688661. [Google Scholar] [CrossRef] [PubMed]
- Gruss, A.; Moretto, V.; Ehrlich, S.D.; Duwat, P.; Dabert, P. GC-rich DNA sequences block homologous recombination in vitro. J. Biol. Chem. 1991, 15, 6667–66669. [Google Scholar] [CrossRef]
- Nagy, P.D.; Bujarski, J.J. Engineering of homologous recombination hotspots with AU-rich sequences in brome mosaic virus. J. Virol. 1997, 71, 3799–3810. [Google Scholar] [CrossRef] [Green Version]
- Adám, E.; Nász, I.; Hudecz, F.; Lengyel, A.; Mezö, G.; Dobay, O. Characterization of intertype specific epitopes on adenovirus hexons. Arch. Virol. 1998, 143, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Maluquer de Motes, C.; Clemente-Casares, P.; Hundesa, A.; Martín, M.; Girones, R. Detection of bovine and porcine adenoviruses for tracing the source of fecal contamination. Appl. Environ. Microbiol. 2004, 70, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Hundesa, A.; Maluquer de Motes, C.; Bofill-Mas, S.; Albinana-Gimenez, N.; Girones, R. Identification of human and animal adenoviruses and polyomaviruses for determination of sources of fecal contamination in the environment. Appl. Environ. Microbiol. 2006, 72, 7886–7893. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Hewitt, J.; Greening, G.E. Viral multiplex quantitative PCR assays for tracking sources of fecal contamination. Appl. Environ. Microbiol. 2010, 76, 1388–1394. [Google Scholar] [CrossRef] [Green Version]
- Nagy, M.; Nagy, E.; Tuboly, T. Sequence analysis of porcine adenovirus serotype 5 fibre gene: Evidence for recombination. Virus Genes 2002, 24, 181–185. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oba, M.; Borjigin, S.; Kikuchi, F.; Oi, T.; Takemae, H.; Ishida, H.; Murakami, H.; Aihara, N.; Shiga, T.; Kamiie, J.; et al. First Isolation and Identification of Homologous Recombination Events of Porcine Adenovirus from Wild Boar. Viruses 2022, 14, 2400. https://doi.org/10.3390/v14112400
Oba M, Borjigin S, Kikuchi F, Oi T, Takemae H, Ishida H, Murakami H, Aihara N, Shiga T, Kamiie J, et al. First Isolation and Identification of Homologous Recombination Events of Porcine Adenovirus from Wild Boar. Viruses. 2022; 14(11):2400. https://doi.org/10.3390/v14112400
Chicago/Turabian StyleOba, Mami, Sumiya Borjigin, Fuka Kikuchi, Toru Oi, Hitoshi Takemae, Hiroho Ishida, Hironobu Murakami, Naoyuki Aihara, Takanori Shiga, Junichi Kamiie, and et al. 2022. "First Isolation and Identification of Homologous Recombination Events of Porcine Adenovirus from Wild Boar" Viruses 14, no. 11: 2400. https://doi.org/10.3390/v14112400