
Mapping and Validation of Peptides Differentially Recognized by Antibodies from the Serum of Yellow Fever Virus-Infected or 17DD-Vaccinated Patients

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
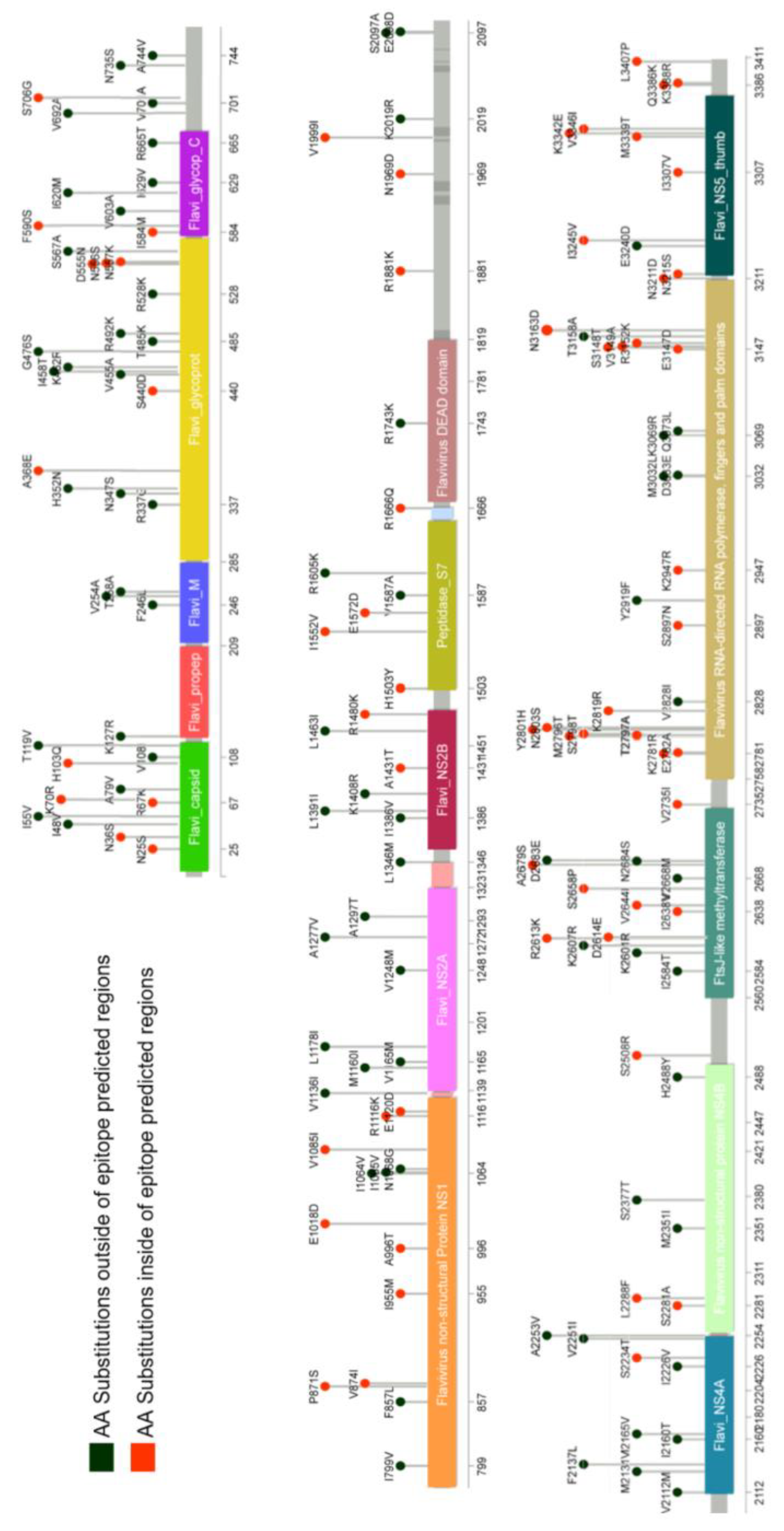
2.2. Design of Peptides
2.3. Microarray Procedures
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Statistical Analysis
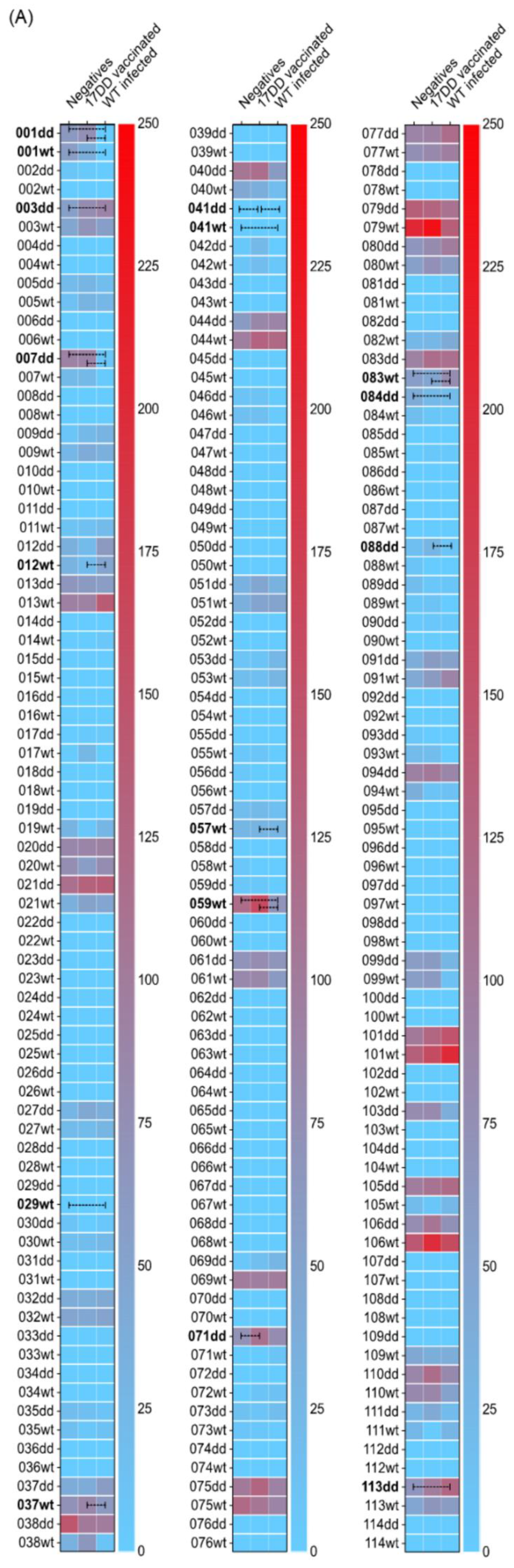
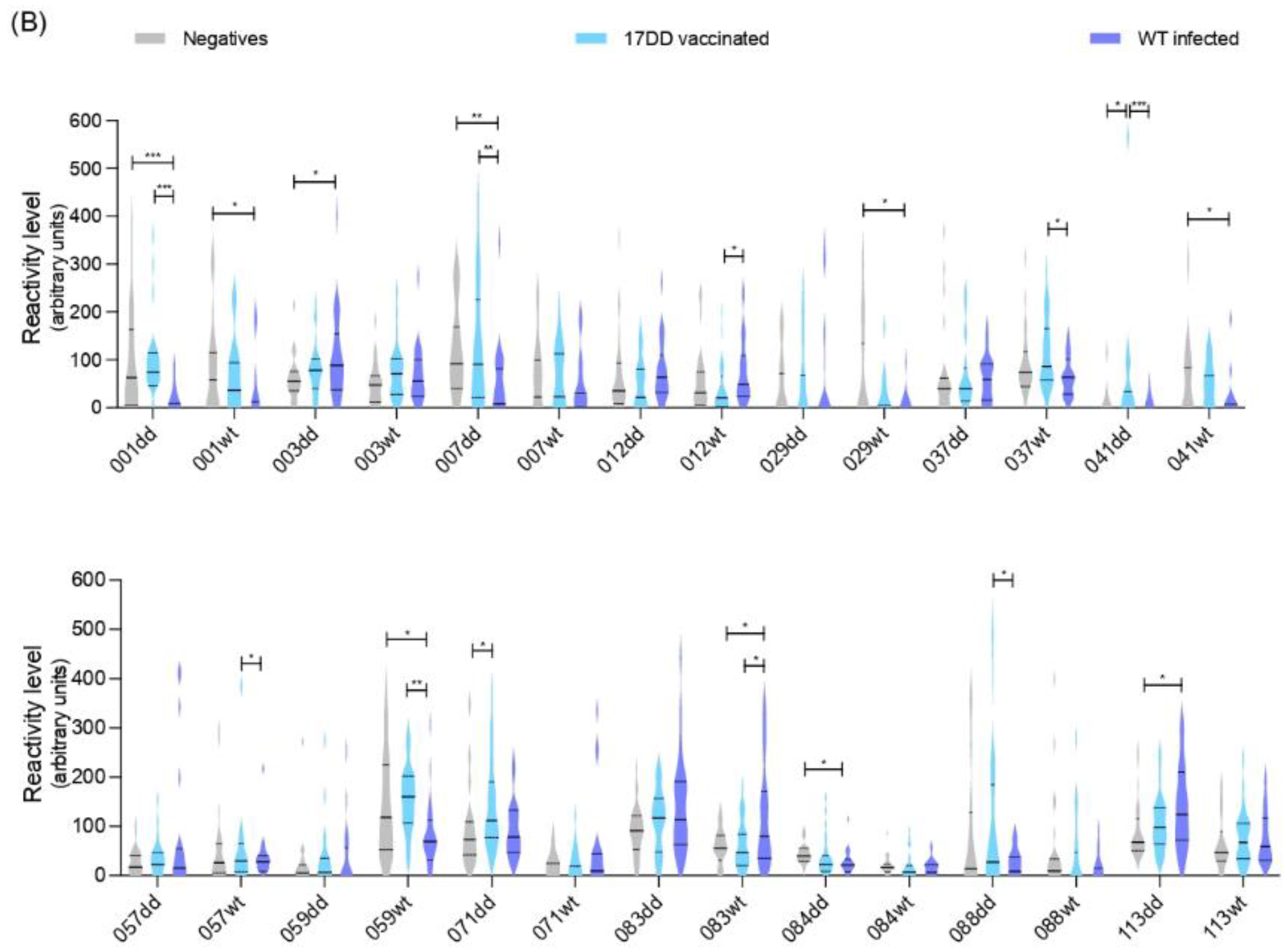
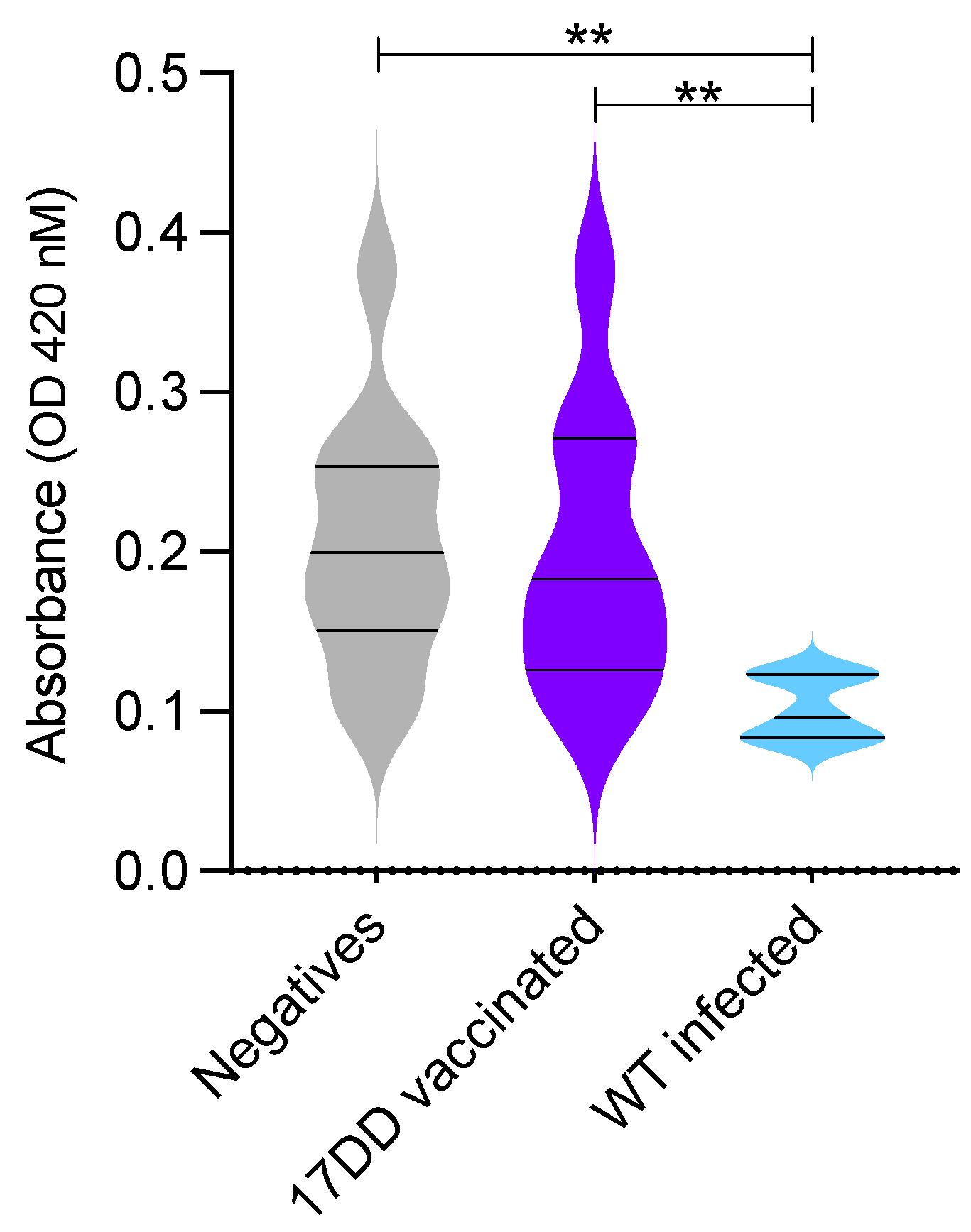
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindenbach, B.D.; Murray, C.L.; Thiel, H.J.; Rice, C.M. Flaviviridae. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 747–794. [Google Scholar]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide Sequence of Yellow Fever Virus: Implications for Flavivirus Gene Expression and Evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus Infection—A Review of Immunopathogenesis, Immunological Response, and Immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, R.; Shen, H.; Wang, M.; Yin, Z.; Cheng, A. Structures and Functions of the Envelope Glycoprotein in Flavivirus Infections. Viruses 2017, 9, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, F.X.; Stiasny, K.; Püschner-Auer, G.; Holzmann, H.; Allison, S.L.; Mandl, C.W.; Kunz, C. Structural Changes and Functional Control of the Tick-Borne Encephalitis Virus Glycoprotein E by the Heterodimeric Association with Protein PrM. Virology 1994, 198, 109–117. [Google Scholar] [CrossRef]
- Ma, L.; Jones, C.T.; Groesch, T.D.; Kuhn, R.J.; Post, C.B. Solution Structure of Dengue Virus Capsid Protein Reveals Another Fold. Proc. Natl. Acad. Sci. USA 2004, 101, 3414–3419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, C.L.; Jones, C.T.; Rice, C.M. Architects of Assembly: Roles of Flaviviridae Non-Structural Proteins in Virion Morphogenesis. Nat. Rev. Microbiol. 2008, 6, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Barrows, N.J.; Campos, R.K.; Liao, K.-C.; Prasanth, K.R.; Soto-Acosta, R.; Yeh, S.-C.; Schott-Lerner, G.; Pompon, J.; Sessions, O.M.; Bradrick, S.S.; et al. Biochemistry and Molecular Biology of Flaviviruses. Chem. Rev. 2018, 118, 4448–4482. [Google Scholar] [CrossRef]
- Akey, D.L.; Brown, W.C.; Dutta, S.; Konwerski, J.; Jose, J.; Jurkiw, T.J.; DelProposto, J.; Ogata, C.M.; Skiniotis, G.; Kuhn, R.J.; et al. Flavivirus NS1 Structures Reveal Surfaces for Associations with Membranes and the Immune System. Science 2014, 343, 881–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avirutnan, P.; Hauhart, R.E.; Somnuke, P.; Blom, A.M.; Diamond, M.S.; Atkinson, J.P. Binding of Flavivirus Nonstructural Protein NS1 to C4b Binding Protein Modulates Complement Activation. J. Immunol. 2011, 187, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.A.; Barrett, A.D.T. The Present and Future of Yellow Fever Vaccines. Pharmaceuticals 2021, 14, 891. [Google Scholar] [CrossRef] [PubMed]
- Theiler, M.; Smith, H.H. The Effect of Prolonged Cultivation in Vitro upon the Pathogenicity of Yellow Fever Virus. J. Exp. Med. 1937, 65, 767–786. [Google Scholar] [CrossRef]
- Gubler, D.J. Human Arbovirus Infections Worlwide. Ann. N. Y. Acad. Sci. 2001, 951, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Sall, A.; Faye, O.; Diallo, M.; Firth, C.; Kitchen, A.; Holmes, E.C. Yellow Fever Virus Exhibits Slower Evolutionary Dynamics than Dengue Virus. J. Virol. 2010, 84, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, P.O.; Stoffella-Dutra, A.G.; Costa, G.B.; Oliveira, J.S.; Amaral, C.D.; Santos, J.D.; Rocha, K.L.S.; Júnior, J.P.A.; Nogueira, M.L.; Borges, M.A.; et al. Re-Emergence of Yellow Fever in Brazil during 2016–2019: Challenges, Lessons Learned, and Perspectives. Viruses 2020, 12, 1233. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D.T. Yellow Fever Vaccines. Biologicals 1997, 25, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Ministério da Saúde; Secretaria de Vigilância em Saúde; Coordenação-Geral de Desenvolvimento da Epidemiologia em Serviços (Eds.). Febre Amarela. In Guia de Vigilância em Saúde, 3rd ed.; Ministério da Saúde: Brasília, Brazil, 2019; pp. 362–387. [Google Scholar]
- Ministério da Saúde. Manual de Vigilância Epidemiológica Da Febre Amarela; Ministério da Saúde, Ed.; Ministério da Saúde: Brasília, Brazil, 1999; pp. 11–16. [Google Scholar]
- Franco, O. História Da Febre-Amarela No Brasil; Ministério da Saúde: Rio de Janeiro, Brazil, 1969. [Google Scholar]
- da Costa Vasconcelos, P.F. Febre Amarela. Rev. Soc. Bras. Med. Trop. 2003, 36, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Delatorre, E.; Abreu, F.V.S.; Ribeiro, I.P.; Gómez, M.M.; Santos, A.A.C.; Ferreira-de-Brito, A.; Neves, M.S.A.S.; Bonelly, I.; Miranda, R.M.; Furtado, N.D.; et al. Distinct YFV Lineages Co-Circulated in the Central-Western and Southeastern Brazilian Regions from 2015 to 2018. Front. Microbiol. 2019, 10, 1079. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Martínez, Y.; Patiño-Barbosa, A.M.; Rodriguez-Morales, A.J. Yellow Fever in the Americas: The Growing Concern about New Epidemics. F1000Research 2017, 6, 398. [Google Scholar] [CrossRef]
- Paules, C.I.; Fauci, A.S. Yellow Fever—Once Again on the Radar Screen in the Americas. N. Engl. J. Med. 2017, 376, 1397–1399. [Google Scholar] [CrossRef]
- Rezende, I.M.; Sacchetto, L.; Mello, É.M.; Alves, P.A.; Iani, F.C.M.; Adelino, T.É.R.; Duarte, M.M.; Cury, A.L.F.; Bernardes, A.F.L.; Santos, T.A.; et al. Persistence of Yellow Fever Virus Outside the Amazon Basin, Causing Epidemics in Southeast Brazil, from 2016 to 2018. PLoS Negl. Trop. Dis. 2018, 12, e0006538. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, P.F.C. Yellow Fever in Brazil: Thoughts and Hypotheses on the Emergence in Previously Free Areas. Rev. Saude Publica 2010, 44, 1144–1149. [Google Scholar] [CrossRef] [Green Version]
- Bifani, A.M.; Ong, E.Z.; Alwis, R. Vaccination and Therapeutics: Responding to the Changing Epidemiology of Yellow Fever. Curr. Treat. Options Infect. Dis. 2020, 12, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, M.C.; Gómez, M.M.; Santos, A.A.; Abreu, F.V.S.; Ferreira-de-Brito, A.; Miranda, R.M.; Castro, M.G.; Oliveira, R.L. Genome Analysis of Yellow Fever Virus of the Ongoing Outbreak in Brazil Reveals Polymorphisms. Mem. Inst. Oswaldo Cruz 2017, 112, 447–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, M.M.; Abreu, F.V.S.; Santos, A.A.C.; Mello, I.S.; Santos, M.P.; Ribeiro, I.P.; Brito, A.F.; Miranda, R.M.; Castro, M.G.; Ribeiro, M.S.; et al. Genomic and Structural Features of the Yellow Fever Virus from the 2016–2017 Brazilian Outbreak. J. Gen. Virol. 2018, 99, 536–548. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Boronat, M.; Diez-Rivero, C.M.; Reinherz, E.L.; Reche, P.A. PVS: A web server for protein sequence variability analysis tuned to facilitate conserved epitope discovery. Nucleic Acids Res. 2008, 36, W35–W41. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Caciula, A.; Price, A.; Thakkar, R.; Ng, J.; Chauhan, L.V.; Jain, K.; Che, X.; Espinosa, D.A.; Montoya Cruz, M.; et al. Diagnosis of Zika Virus Infection by Peptide Array and Enzyme-Linked Immunosorbent Assay. mBio 2018, 9, e00095-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombarolli, S.G.; Batista, I.C.A.; Tavares, N.C.; Oliveira, E.S.; Nascimento, C.S.; Felgner, P.L.; Assis, R.R.; Silva, C.E.C. Peptide Microarrays for Flavivirus Diagnosis. In Peptide Microarrays, 3rd ed.; Cretich, M., Ed.; Springer Nature: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving Sequence-Based B-Cell Epitope Prediction Using Conformational Epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [Green Version]
- Rosser, J.I.; Nielsen-Saines, K.; Saad, E.; Fuller, T. Reemergence of Yellow Fever Virus in Southeastern Brazil, 2017–2018: What Sparked the Spread? PLoS Negl. Trop. Dis. 2022, 16, e0010133. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Torres, M.A.N.; Paula, M.B.; Fernandes, A.; Marassá, A.M.; Consales, C.A.; Fonseca, D.F. Ecologia de Haemagogus e Sabethes (Diptera: Culicidae) Em Áreas Epizoóticas Do Vírus Da Febre Amarela, Rio Grande Do Sul, Brasil. Epidemiol. Serviços Saúde 2010, 19, 101–113. [Google Scholar] [CrossRef]
- Muller, D.A.; Young, P.R. The Flavivirus NS1 Protein: Molecular and Structural Biology, Immunology, Role in Pathogenesis and Application as a Diagnostic Biomarker. Antiviral Res. 2013, 98, 192–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assis, R.R.; Jain, A.; Nakajima, R.; Jasinskas, A.; Felgner, J.; Obiero, J.M.; Norris, P.J.; Stone, M.; Simmons, G.; Bagri, A.; et al. Analysis of SARS-CoV-2 Antibodies in COVID-19 Convalescent Blood Using a Coronavirus Antigen Microarray. Nat. Commun. 2021, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Vieira, D.F.; Couto-Lima, D.; Jácome, F.C.; Caldas, G.C.; Barth, O.M. Dengue, Yellow Fever, Zika and Chikungunya Epidemic Arboviruses in Brazil: Ultrastructural Aspects. Mem. Inst. Oswaldo Cruz 2020, 115, e200278. [Google Scholar] [CrossRef] [PubMed]
- Estofolete, C.F.; Terzian, A.C.B.; Colombo, T.E.; de Freitas Guimarães, G.; Ferraz, H.C.; da Silva, R.A.; Greque, G.V.; Nogueira, M.L. Co-Infection between Zika and Different Dengue Serotypes during DENV Outbreak in Brazil. J. Infect. Public Health 2019, 12, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Souza, N.C.S.; Félix, A.C.; Paula, A.V.; Levi, J.E.; Pannuti, C.S.; Romano, C.M. Evaluation of Serological Cross-Reactivity between Yellow Fever and Other Flaviviruses. Int. J. Infect. Dis. 2019, 81, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, J.J.; Monath, T.P.; Cropp, C.B.; Brandriss, M.W.; Prange, W.C. Yellow Fever Monoclonal Antibodies: Type-Specific and Cross-Reactive Determinants Identified by Immunofluorescence. Am. J. Trop. Med. Hyg. 1984, 33, 695–698. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AVQ67911.1 | AVQ67912.1 | AVQ67913.1 | AVQ67914.1 | AVQ67915.1 | AVQ67916.1 | AVQ94372.1 | ES-505 | AVQ94369.1 | AVQ94370.1 | AVQ94371.1 | |
| AVQ67911.1 | 1 | ||||||||||
| AVQ67912.1 | 0.9997 | 1 | |||||||||
| AVQ67913.1 | 0.9997 | 1 | 1 | ||||||||
| AVQ67914.1 | 0.9997 | 1 | 1 | 1 | |||||||
| AVQ67915.1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | ||||||
| AVQ67916.1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | 1 | |||||
| AVQ94372.1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | 1 | 1 | ||||
| ES-505 | 0.9997 | 1 | 1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | |||
| AVQ94369.1 | 0.9997 | 1 | 1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | 1 | ||
| AVQ94370.1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | 1 | 1 | 0.99 | 0.9997 | 1 | |
| AVQ94371.1 | 0.9997 | 1 | 1 | 1 | 0.9997 | 0.9997 | 0.9997 | 1 | 1 | 0.9997 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, E.S.; Tavares, N.C.; Colombarolli, S.G.; Batista, I.C.A.; Nascimento, C.S.; Felgner, P.L.; de Assis, R.R.; Calzavara-Silva, C.E. Mapping and Validation of Peptides Differentially Recognized by Antibodies from the Serum of Yellow Fever Virus-Infected or 17DD-Vaccinated Patients. Viruses 2022, 14, 1645. https://doi.org/10.3390/v14081645
Oliveira ES, Tavares NC, Colombarolli SG, Batista ICA, Nascimento CS, Felgner PL, de Assis RR, Calzavara-Silva CE. Mapping and Validation of Peptides Differentially Recognized by Antibodies from the Serum of Yellow Fever Virus-Infected or 17DD-Vaccinated Patients. Viruses. 2022; 14(8):1645. https://doi.org/10.3390/v14081645
Chicago/Turabian StyleOliveira, Eneida Santos, Naiara Clemente Tavares, Stella Garcia Colombarolli, Izabella Cristina Andrade Batista, Camila Sales Nascimento, Philip Louis Felgner, Rafael Ramiro de Assis, and Carlos Eduardo Calzavara-Silva. 2022. "Mapping and Validation of Peptides Differentially Recognized by Antibodies from the Serum of Yellow Fever Virus-Infected or 17DD-Vaccinated Patients" Viruses 14, no. 8: 1645. https://doi.org/10.3390/v14081645