
Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection and Pathological Examination
2.3. Cell Lines, Virus and Antibodies
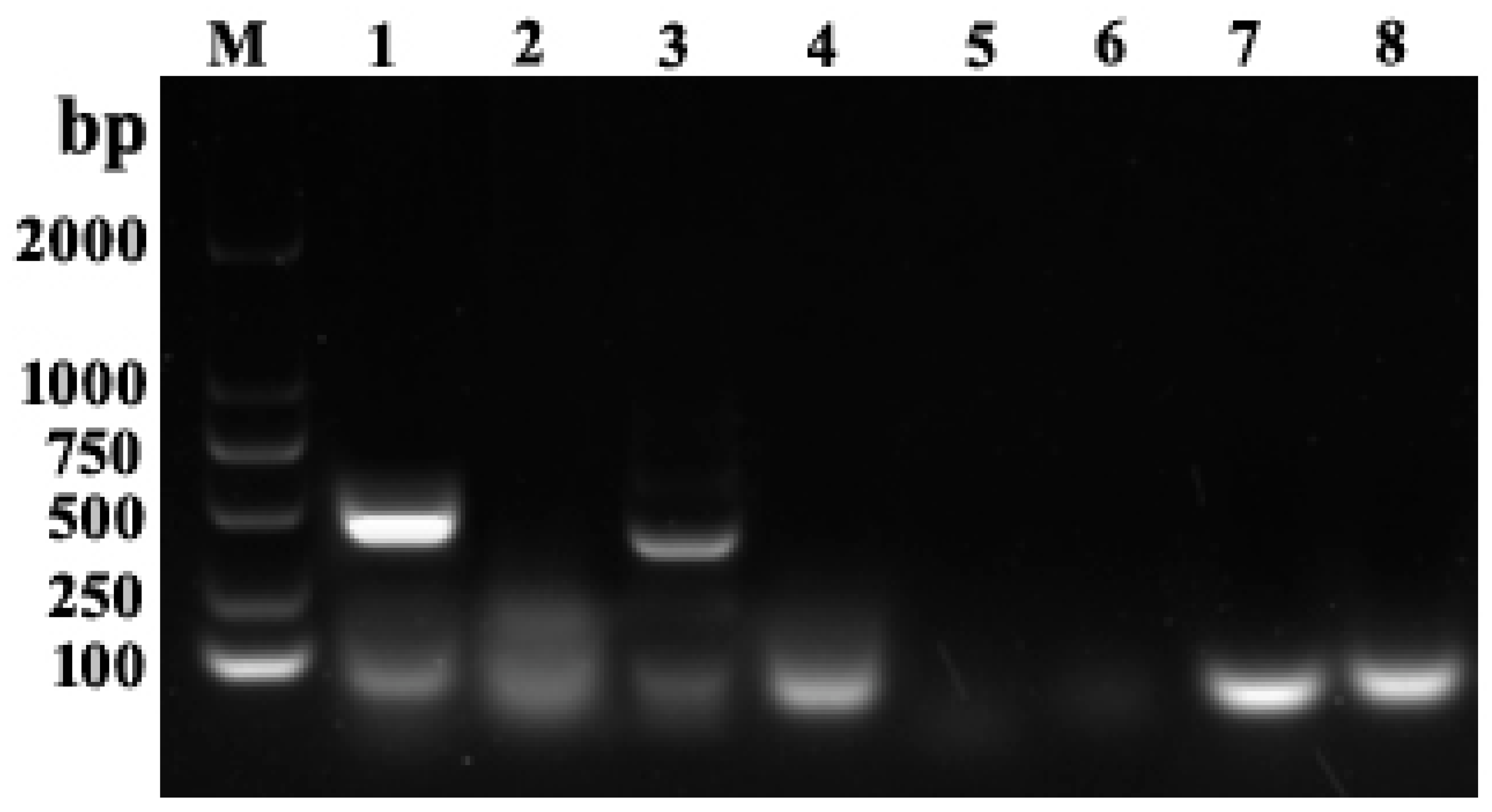
2.4. Primers and Probes
2.5. Virus Detection
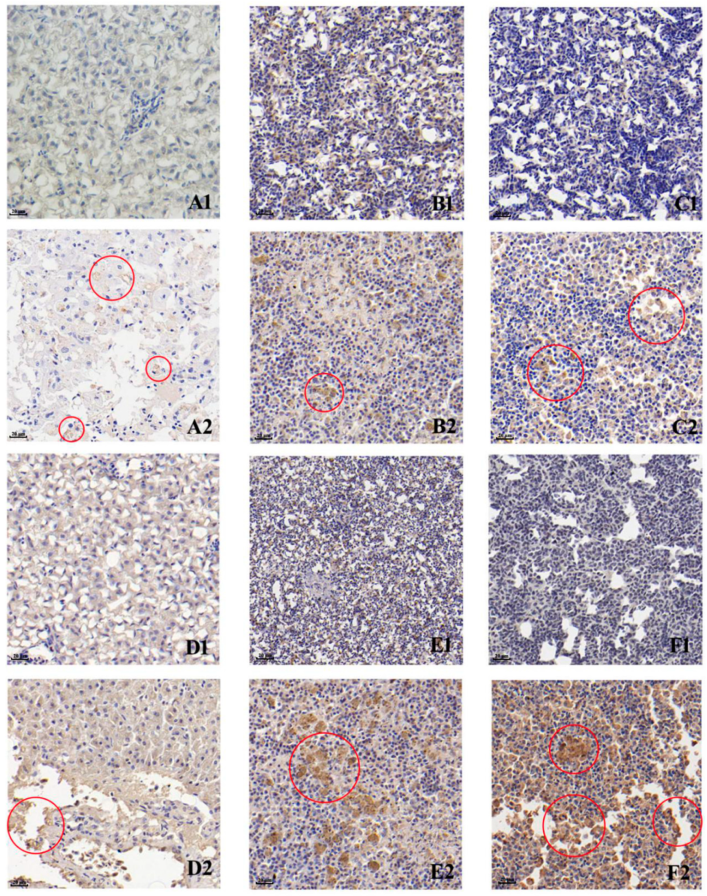
2.6. Immunohistochemistry
2.7. Virus Isolation
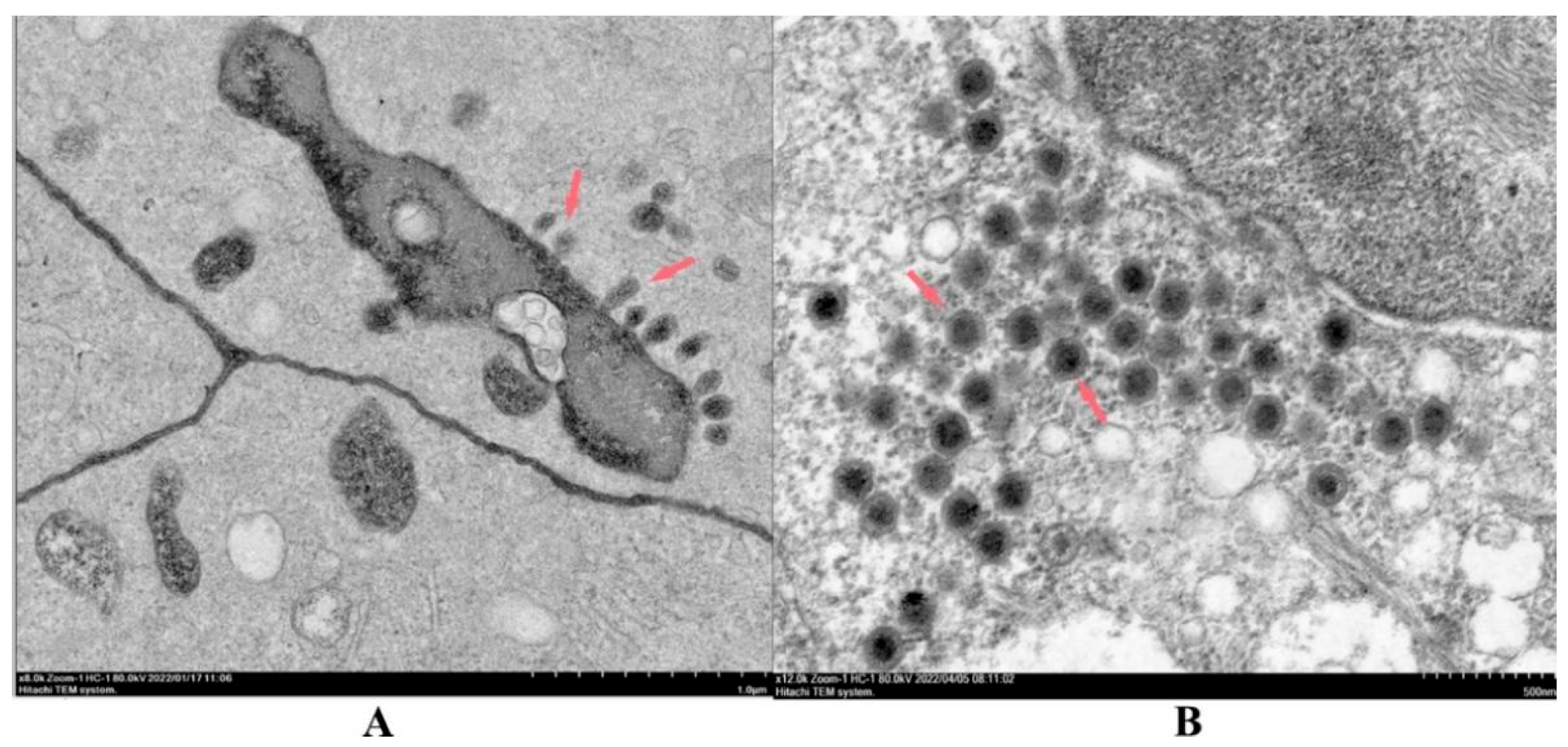
2.8. Morphology of Virus Particles
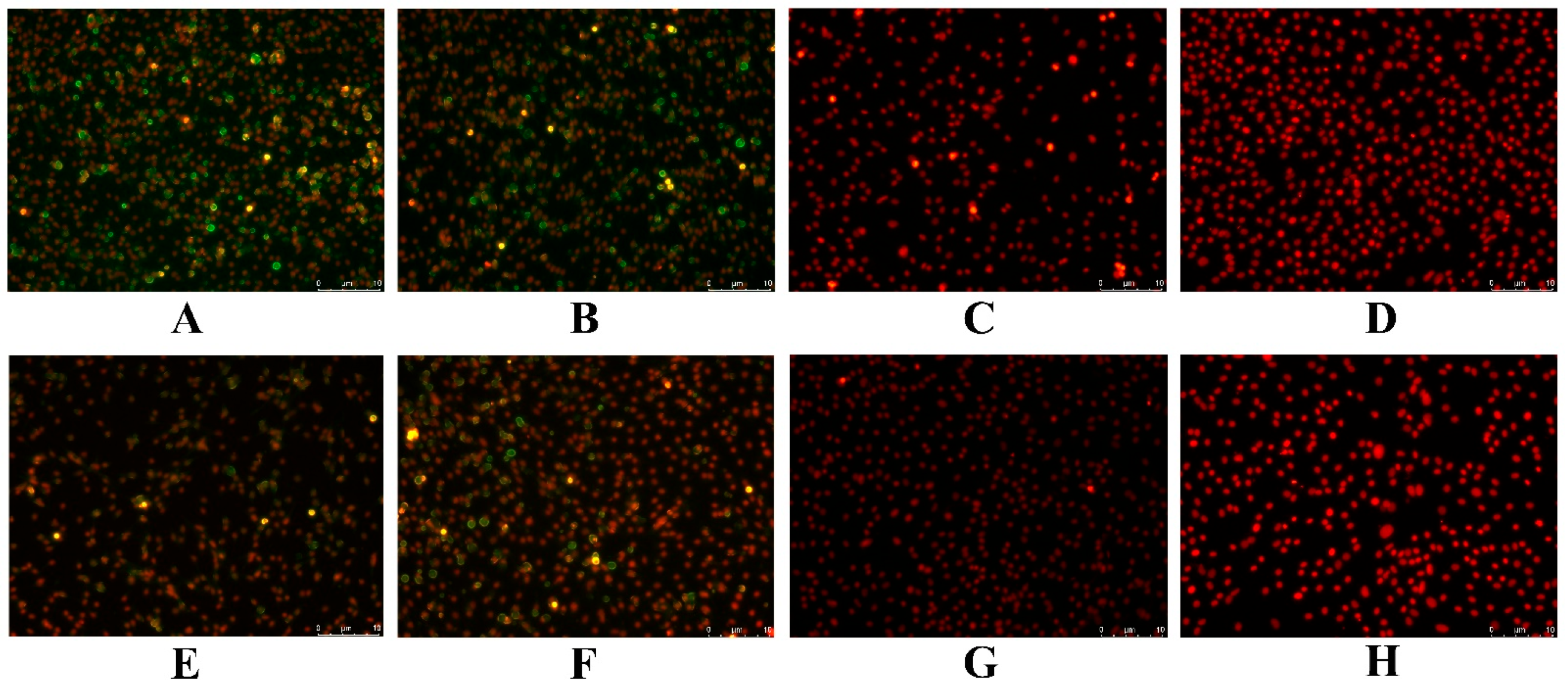
2.9. Antigen Identification
2.10. Virus Plaque Assay
2.11. Sequence and Evolutionary Analysis
2.12. Regression Infection Test
2.13. Statistical Analyses
3. Results
3.1. Clinical Characteristics
3.2. Bacterial Isolation and Virus Detection
3.3. Immunohistochemistry (IHC)
3.4. Virus Isolation
3.5. Morphology of Virus Particles
3.6. Antigen Identification
3.7. Virus Plaque Assays
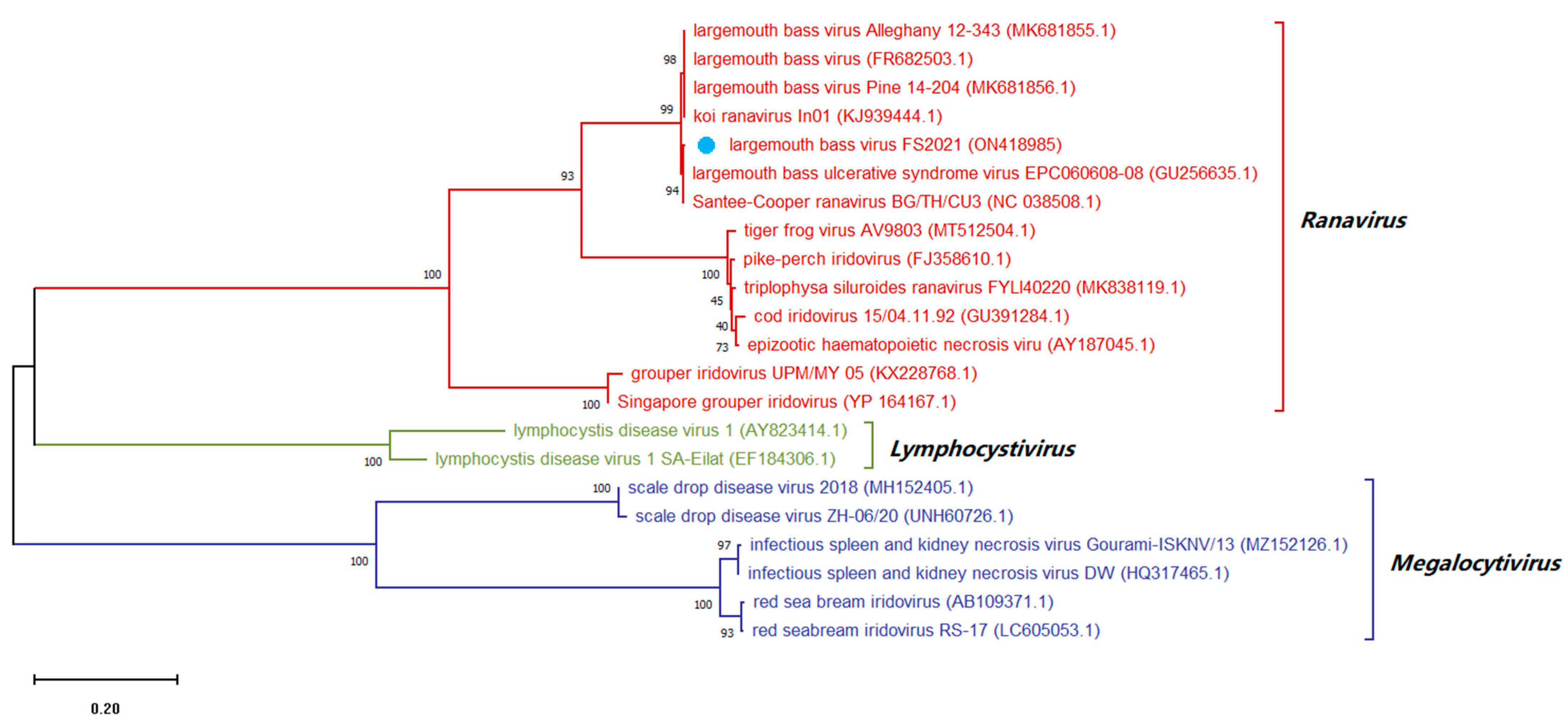
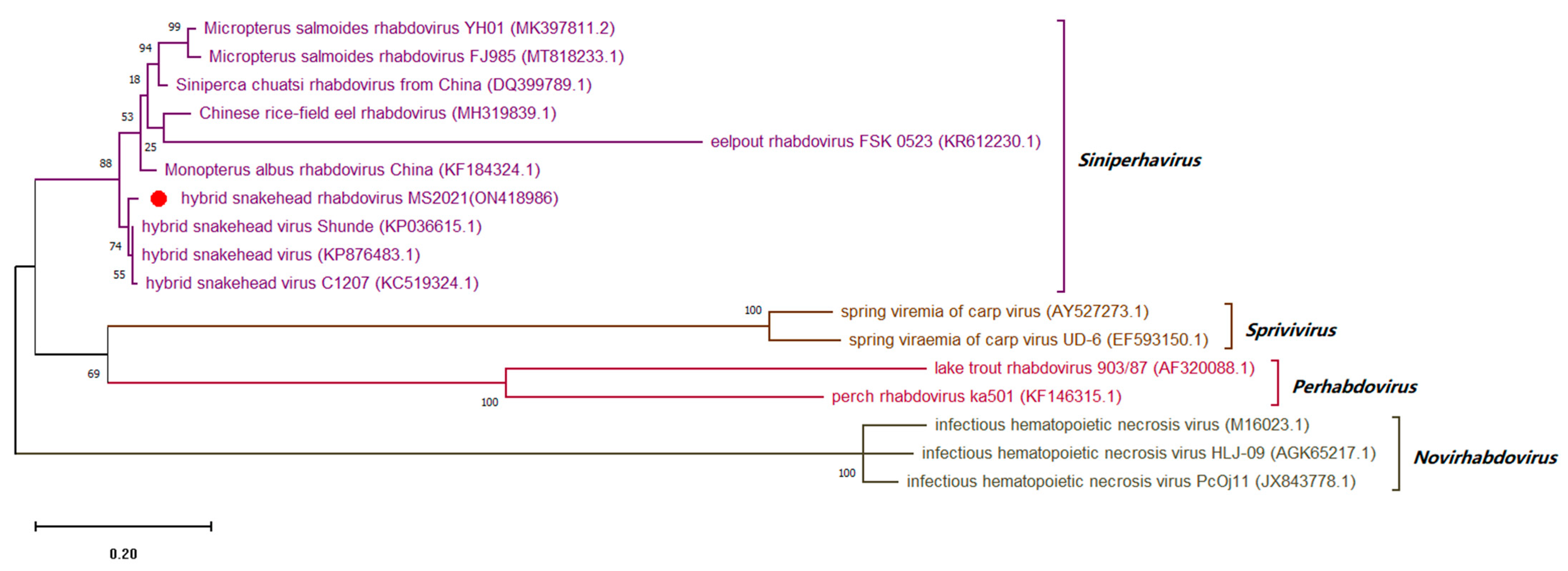
3.8. Sequence Analysis of Conserved Regions of Viral Genome
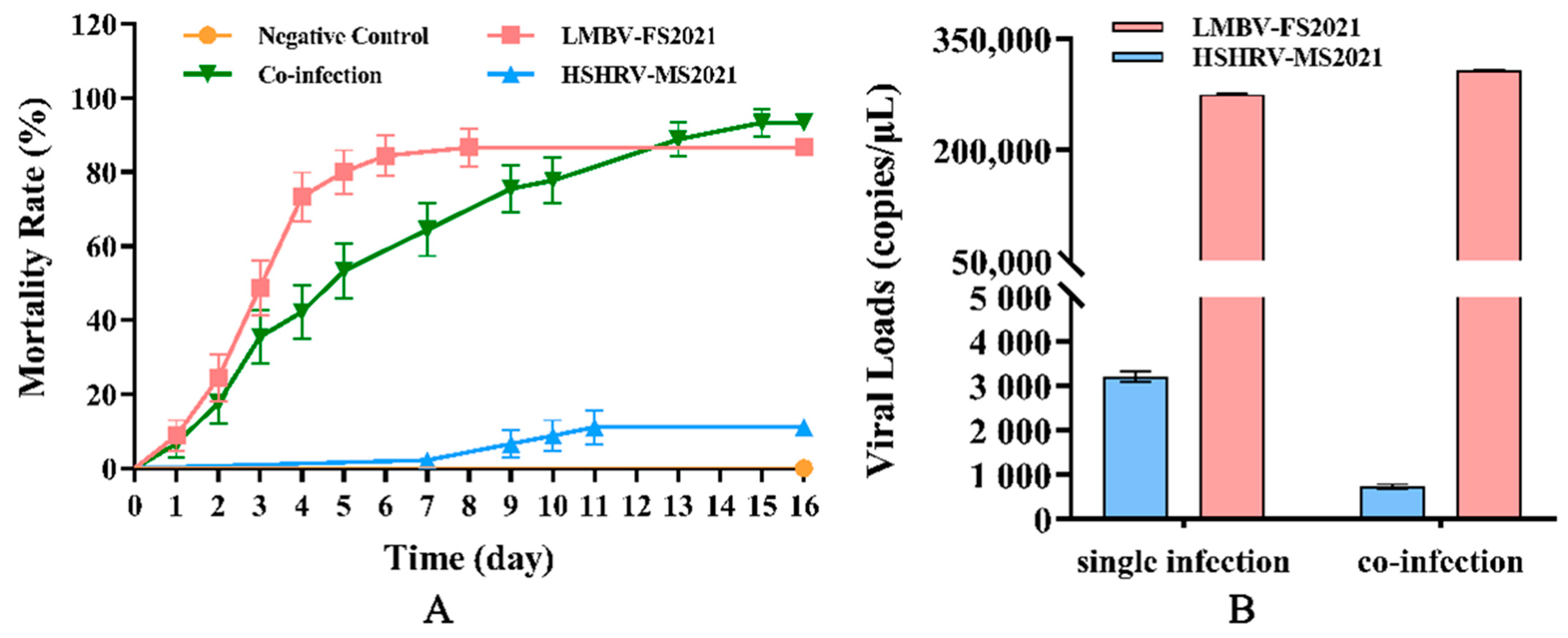
3.9. Regression Infection Experiments
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, E.B.; Chen, G. Micropterus salmoides rhabdovirus (MSRV) infection induced apoptosis and activated interferon signaling pathway in largemouth bass skin cells. Fish Shellfish Immunol. 2018, 76, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yao, H.; Li, Y.H.; Xu, Y.J.; Ma, X.F.; Wang, H.P. Global diversity and genetic landscape of natural populations and hatchery stocks of largemouth bass micropterus salmoides across American and Asian regions. Sci. Rep. 2019, 9, 16697. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhong, L.; Kang, Q.; Liu, S.; Feng, Y.; Geng, Y.; Chen, D.; Ou, Y.; Yang, S.; Yin, L.; et al. A High Starch Diet Alters the Composition of the Intestinal Microbiota of Largemouth Bass Micropterus salmoides, Which May Be Associated With the Development of Enteritis. Front. Microbiol. 2021, 12, 696588. [Google Scholar] [CrossRef] [PubMed]
- China Fisheries Statistical Yearbook Committee. China Fishery Statistical Yearbook; Agriculture Press: Beijing, China, 2021; Available online: https://www.yearbookchina.com/navibooklist-n3022013079-1.html (accessed on 30 May 2022).
- Feng, Z.; Chu, X.; Han, M.; Yu, C.; Jiang, Y.; Wang, H.; Lu, L.; Xu, D. Rapid visual detection of Micropterus salmoides rhabdovirus using recombinase polymerase amplification combined with lateral flow dipsticks. J. Fish Dis. 2022, 45, 461–469. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, H.; Zhang, L.; Li, Q.; Huang, Z.; Zhao, Z.; Ke, H.; Xiao, Y.; Su, X.; Liu, Q.; et al. MiRNA-seq analysis of spleen and head kidney tissue from aquacultured largemouth bass (Micropterus salmoides) in response to Aeromonas hydrophila infection. Funct. Integr. Genom. 2021, 21, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Li, M.; Liu, M.; Huang, S.; Wang, G.; Wang, T.; Li, P. Selection and Characterization of ssDNA Aptamers Targeting Largemouth Bass Virus Infected Cells With Antiviral Activities. Front. Microbiol. 2021, 12, 785318. [Google Scholar] [CrossRef]
- Jia, Y.J.; Guo, Z.R.; Ma, R.; Qiu, D.K.; Wang, G.X.; Zhu, B. Protective immunity of largemouth bass immunized with immersed DNA vaccine against largemouth bass ulcerative syndrome virus. Fish Shellfish Immunol. 2020, 107, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Luo, M.; Zheng, G.; Liang, H.; Liu, L.; Lin, Q.; Niu, Y.; Luo, X.; Li, N. Determination and Characterization of a Novel Birnavirus Associated with Massive Mortality in Largemouth Bass. Microbiol. Spectr. 2022, 10, e0171621. [Google Scholar] [CrossRef]
- Ma, D.; Deng, G.; Bai, J.; Li, S.; Yu, L.; Quan, Y.; Yang, X.; Jiang, X.; Zhu, Z.; Ye, X. A strain of Siniperca chuatsi rhabdovirus causes high mortality among cultured Largemouth Bass in South China. J. Aquat. Anim. Health 2013, 25, 197–204. [Google Scholar] [CrossRef]
- Sibley, S.D.; Finley, M.A.; Baker, B.B.; Puzach, C.; Armién, A.G.; Giehtbrock, D.; Goldberg, T.L. Novel reovirus associated with epidemic mortality in wild largemouth bass (Micropterus salmoides). J. Gen. Virol. 2016, 97, 2482–2487. [Google Scholar] [CrossRef]
- Bovo, G.; Gustinelli, A.; Quaglio, F.; Gobbo, F.; Panzarin, V.; Fusaro, A.; Mutinelli, F.; Caffara, M.; Fioravanti, M.L. Viral encephalopathy and retinopathy outbreak in freshwater fish farmed in Italy. Dis. Aquat. Org. 2011, 96, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Godahewa, G.I.; Lee, S.; Kim, J.; Perera, N.C.N.; Kim, M.J.; Kwon, M.G.; Jee, B.Y.; Hwang, S.D.; Lee, J. Analysis of complete genome and pathogenicity studies of the spring viremia of carp virus isolated from common carp (Cyprinus carpio carpio) and largemouth bass (Micropterus salmoides): An indication of SVC disease threat in Korea. Virus Res. 2018, 255, 105–116. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Z.; Xiang, Y.; Jia, P.; Liu, W.; Yi, M.; Jia, K. Isolation and identification of a viral haemorrhagic septicaemia virus (VHSV) isolate from wild largemouth bass Micropterus salmoides in China. J. Fish Dis. 2019, 42, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Zhao, R.; Geng, Y.; Wang, K.; Yang, P.O.; Chen, D.; Huang, X.; Zuo, Z.; He, C.; Chen, Z.; et al. Nocardia seriolae: A serious threat to the largemouth bass Micropterus salmoides industry in Southwest China. Dis. Aquat. Org. 2020, 142, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Camus, A.; Griffin, M.; Armwood, A.; Soto, E. A Spontaneous Outbreak of Systemic Edwardsiella piscicida Infection in Largemouth Bass Micropterus salmoides (Lacépède, 1802) in California, USA. J. Fish Dis. 2019, 42, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Bebak, J.; Matthews, M.; Shoemaker, C. Survival of vaccinated, feed-trained largemouth bass fry (Micropterus salmoides floridanus) during natural exposure to Flavobacterium columnare. Vaccine 2009, 27, 4297–4301. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Lu, G.; Du, J.; Wang, J.; Hu, Y.; Su, Y.; Li, A. First report of trypanosomiasis in farmed largemouth bass (Micropterus salmoides) from China: Pathological evaluation and taxonomic status. Parasitol. Res. 2019, 118, 1731–1739. [Google Scholar] [CrossRef] [PubMed]
- Pomposini, A.; Blubaugh, J.; Boyce, R.C.; Gauthier, D.T. Leech (Myzobdella lugubris) infestations in largemouth bass (Micropterus salmoides) in Back Bay, Virginia, USA. J. Fish Dis. 2019, 42, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Grizzle, J.M.; Altinok, I.; Fraser, W.A.; Francis-Floyd, R. First isolation of largemouth bass virus. Dis. Aquat. Org. 2002, 50, 233–235. [Google Scholar] [CrossRef]
- Dong, C.; Wang, Z.; Weng, S.; He, J. Occurrence of a lethal ranavirus in hybrid mandarin (Siniperca scherzeri × Siniperca chuatsi) in Guangdong, South China. Vet. Microbiol. 2017, 203, 28–33. [Google Scholar] [CrossRef]
- Boonthai, T.; Loch, T.P.; Yamashita, C.J.; Smith, G.D.; Winters, A.D.; Kiupel, M.; Brenden, T.O.; Faisal, M. Laboratory investigation into the role of largemouth bass virus (Ranavirus, Iridoviridae) in smallmouth bass mortality events in Pennsylvania rivers. BMC Vet. Res. 2018, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, A.M.; Stone, D.M.; Way, K.; Torhy, C.; Chilmonczyk, S.; Benmansour, A.; de Kinkelin, P. Emerging vesiculo-type virus infections of freshwater fishes in Europe. Dis. Aquat. Org. 2003, 57, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, H.L.; Blazer, V.S.; Smith, G.D.; Lookenbill, M.; Alvarez, D.A.; Smalling, K.L. Risk Factors Associated with Mortality of Age-0 Smallmouth Bass in the Susquehanna River Basin, Pennsylvania. J. Aquat. Anim. Health 2018, 30, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Diamanka, A.; Loch, T.P.; LaFrentz, B.R.; Winters, A.D.; García, J.C.; Toguebaye, B.S. Isolation and characterization of Flavobacterium columnare strains infecting fishes inhabiting the Laurentian Great Lakes basin. J. Fish Dis. 2017, 40, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Dong, H.; Chen, X.; Bergmann, S.M.; Yang, Y.; Wei, X.; Tong, G.; Li, H.; Yu, H.; Chen, Y. Establishment and characterization of a permanent heart cell line from largemouth bass Micropterus salmoides and its application to fish virology and immunology. Aquaculture 2022, 547, 737427. [Google Scholar] [CrossRef]
- Ramírez-Paredes, J.G.; Paley, R.K.; Hunt, W.; Feist, S.W.; Stone, D.M.; Field, T.R.; Haydon, D.J.; Ziddah, P.A.; Nkansa, M.; Guilder, J.; et al. First detection of infectious spleen and kidney necrosis virus (ISKNV) associated with massive mortalities in farmed tilapia in Africa. Transbound. Emerg. Dis. 2021, 68, 1550–1563. [Google Scholar] [CrossRef]
- Yinnan, M.; Kebing, L.; Xinhua, C.; Jingqun, A. Diagnosis of nervous necrosis virus in orange-spotted grouper, Epinephelus coioides, by a rapid and convenient RT-PCR method. Acta Oceanol. Sin. (China) 2013, 32, 88–92. [Google Scholar]
- Guo, Y.; Wang, Y.; Fan, Z.; Zhao, X.; Bergmann, S.M.; Dong, H.; Jin, Y.; Sun, D.; Mai, Q.; Liu, W.; et al. Establishment and evaluation of qPCR and real-time recombinase-aided amplification assays for detection of largemouth bass ranavirus. J. Fish Dis. 2022, 45, 1033–1043. [Google Scholar] [CrossRef]
- Liang, H.R.; Cai, X.Z.; Fan, Z.Y.; Lin, Q.; Fu, X.Z.; Liu, L.H.; Huang, Z.B.; Niu, Y.J.; Lin, L.; Li, N.Q. Establishment and application of a Taq Man real-time PCR assay for the detection of Siniperca chuatsi rhabdovirus. Chin. J. Prev. Vet. Med. (China) 2019, 41, 929–934. [Google Scholar]
- Yavuz, O.; Erer, H. Immunohistochemical and immunocytochemical findings associated with Marek’s disease virus in naturally infected laying hens. Biotech. Histochem. Off. Publ. Biol. Stain. Comm. 2017, 92, 498–505. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J. Immunohistochemical Staining of Influenza Virus in Tissues. Methods Mol. Biol. (Clifton N. J.) 2020, 2123, 29–36. [Google Scholar]
- Tamošiūnaitė, A.; Hoffmann, D.; Franke, A.; Schluckebier, J.; Tauscher, K.; Tischer, B.K.; Beer, M.; Klopfleisch, R.; Osterrieder, N. Histopathological and Immunohistochemical Studies of Cowpox Virus Replication in a Three-Dimensional Skin Model. J. Comp. Pathol. 2016, 155, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Lin, Q.; Liang, H.; Liu, L.; Huang, Z.; Li, N.; Su, J. The biological features and genetic diversity of novel fish rhabdovirus isolates in China. Arch. Virol. 2017, 162, 2829–2834. [Google Scholar] [CrossRef] [PubMed]
- Sunarto, A.; McColl, K.A.; Crane, M.S.; Sumiati, T.; Hyatt, A.D.; Barnes, A.C.; Walker, P.J. Isolation and characterization of koi herpesvirus (KHV) from Indonesia: Identification of a new genetic lineage. J. Fish Dis. 2011, 34, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, H.; Zeng, W.; Wang, L.; Liu, C.; Wu, J.; Wang, Y.; Li, Y.; Bergmann, S.M. Development of indirect immunofluorescence assay for TCID(50) measurement of grass carp reovirus genotype II without cytopathic effect onto cells. Microb. Pathog. 2018, 114, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.; Subramaniam, K.; Waltzek, T.B.; Whittington, R.; Williams, T.; et al. ICTV Virus Taxonomy Profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef]
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef]
- Walker, P.J.; Bigarré, L.; Kurath, G.; Dacheux, L.; Pallandre, L. Revised Taxonomy of Rhabdoviruses Infecting Fish and Marine Mammals. Animals 2022, 12, 1363. [Google Scholar] [CrossRef]
- Lyu, S.J.; Yuan, X.M.; Zhang, H.Q.; Shi, W.D.; Hang, X.Y.; Liu, L.; Wu, Y.L. Isolation and characterization of a novel strain (YH01) of Micropterus salmoides rhabdovirus and expression of its glycoprotein by the baculovirus expression system. J. Zhejiang Univ. (China) 2019, 20, 728–739. [Google Scholar] [CrossRef]
- Zilberg, D.; Grizzle, J.M.; Plumb, J.A. Preliminary description of lesions in juvenile largemouth bass injected with largemouth bass virus. Dis. Aquat. Org. 2000, 39, 143–146. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, Q.; Wang, Y.; Liu, C.; Liang, H.; Fang, X.; Wu, S. Genomic characterization and taxonomic position of a rhabdovirus from a hybrid snakehead. Arch. Virol. 2014, 159, 2469–2473. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Fu, X.; Li, N.; Wan, Q.; Chen, W.; Huang, Y.; Huang, Z.; Li, J.; Zhao, L.; Lin, L. Co-infections of infectious spleen and kidney necrosis virus and Siniperca chuatsi rhabdovirus in Chinese perch (Siniperca chuatsi). Microb. Pathog. 2017, 111, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.; Castric, J.; Lamour, F.; Cabon, J.; Quentel, C. Study of the viral interference between infectious pancreatic necrosis virus (IPNV) and infectious haematopoietic necrosis virus (IHNV) in rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2008, 24, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, K.; Praveenraj, J.; Kiruba-Sankar, R.; Devi, V.; Biswas, U.; Kumar, T.S.; Sudhagar, A.; El-Matbouli, M.; Kumar, G. Co-Infection of Infectious Hypodermal and Hematopoietic Necrosis Virus (IHHNV) and White Spot Syndrome Virus (WSSV) in the Wild Crustaceans of Andaman and Nicobar Archipelago, India. Viruses 2021, 13, 1378. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Product Size (bp) | References/Purpose |
|---|---|---|---|
| LMBV-DT-F | GCTGGCGGCCAACCAGTTTAAC | 430 | This study Virus detection |
| LMBV-DT-R | GGCCACGATTGGCTTGACTTCT | ||
| MSRV-DT-F | GGGCTGGATGATAGACGATTG | 349 | This study Virus detection |
| MSRV-DT-R | TGGCGGAGGTGCTTGATATGG | ||
| ISKNV-DT-F | GGTTCATCGACATCTCCGCG | 431 | [27] Virus detection |
| ISKNV-DT-R | AGGTCGCTGCGCATGCCAATC | ||
| NNV-P0 | CGAGTCAACACGGGTGAAGACAG | 326 | [28] Virus detection |
| NNV-P1 | ACCGCTCCCATCATGACACAA | ||
| NNV-P2 | AACAGGCAGCAGAATTTGACG | ||
| MCP-F | ACCAACATTTCTATCGCTTAT | 1456 | This study Sequencing |
| MCP-R | TGCGATATGGAAACGTAGTAA | ||
| G1-F | ATTAATCAATGGTGTTGGTGG | 946 | This study Sequencing |
| G1-R | CCACCAACACCATTGATTAAT | ||
| G2-F | CATATCCGAATTGCGAAGAGC | 637 | This study Sequencing |
| G2-R | TTCTTGAGAATAATCCATGAT | ||
| LMBV-qPCR-F | GGCCACCACCTCTACTCTTAC | 120 | [29] Virus loads determination |
| LMBV-qPCR-R | GGCAGACAGAGACACGTTGA | ||
| LMBV-probe | FAM-CTTCAGGGTCTACCAATTTCGGTC-TAMRA | ||
| MSRV-qPCR-F | GACATGTTCTTCTACAGATTCAAC | 140 | [30] Virus loads determination |
| MSRV-qPCR-R | CAATCCAGCACTCCACTG | ||
| MSRV-probe | FAM-AGGTTCAAAGACTGTGCAGCTCTGT-TAMRA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Bergmann, S.M.; Mai, Q.; Yang, Y.; Liu, W.; Sun, D.; Chen, Y.; Yu, Y.; Liu, Y.; Cai, W.; et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses 2022, 14, 1643. https://doi.org/10.3390/v14081643
Jin Y, Bergmann SM, Mai Q, Yang Y, Liu W, Sun D, Chen Y, Yu Y, Liu Y, Cai W, et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses. 2022; 14(8):1643. https://doi.org/10.3390/v14081643
Chicago/Turabian StyleJin, Yuqi, Sven M. Bergmann, Qianyi Mai, Ying Yang, Weiqiang Liu, Dongli Sun, Yanfeng Chen, Yingying Yu, Yuhong Liu, Wenlong Cai, and et al. 2022. "Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides)" Viruses 14, no. 8: 1643. https://doi.org/10.3390/v14081643