
Impact of Predator Exclusion and Habitat on Seroprevalence of New World Orthohantavirus Harbored by Two Sympatric Rodents within the Interior Atlantic Forest

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Grid Enclosure and Predator Removal
2.3. Rodent Collection
2.4. Hantavirus Antibody Screening by Immunofluorescence Assays (IFA)
2.5. Statistical Analyses
2.6. Ethics Statements
3. Results
3.1. Rodent Sampling
3.2. Population Size, Abundance, and Experimental Variables
3.3. Effects of Grid Enclosure, Session, and Forest Degradation Level on Rodent Species Diversity
3.4. Hantavirus Seroprevalence
3.5. Effects of Grid Enclosure, Session, and Forest Degradation Level on Rodent Seroprevalence
3.6. Effects of Species Diversity on Rodent Seroprevalence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current Classification and Future Perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plyusnin, A.; Morzunov, S.P. Virus evolution and genetic diversity of hantaviruses and their rodent hosts. In Hantaviruses; Schmaljohn, C., Nichol, S.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 256, pp. 47–75. [Google Scholar]
- Plyusnin, A.; Sironen, T. Evolution of hantaviruses: Co-speciation with reservoir hosts for more than 100MYR. Virus Res. 2014, 187, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-P.; Lin, X.-D.; Wang, W.; Tian, J.-H.; Cong, M.-L.; Zhang, H.-L.; Wang, M.-R.; Zhou, R.-H.; Wang, J.-B.; Li, M.-H.; et al. Phylogeny and Origins of Hantaviruses Harbored by Bats, Insectivores, and Rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.L.; Friedman, R. Evolutionary Diversification of Protein-Coding Genes of Hantaviruses. Mol. Biol. Evol. 2000, 17, 1558–1568. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.S.; Khabbaz, R.F.; Armstrong, L.R.; Holman, R.C.; Bauer, S.P.; Graber, J.; Strine, T.; Miller, G.; Reef, S.; Tappero, J.; et al. Hantavirus Pulmonary Syndrome: The First 100 US Cases. J. Infect. Dis. 1996, 173, 1297–1303. [Google Scholar] [CrossRef]
- Bi, Z.; Formenty, P.B.H.; Roth, C.E. Hantavirus Infection: A review and global update. J. Infect. Dev. Ctries. 2008, 2. [Google Scholar] [CrossRef]
- Toro, J.; Vega, J.D.; Khan, A.S.; Mills, J.N.; Padula, P.; Terry, W.; Yadón, Z.; Valderrama, R.; Ellis, B.A.; Pavletic, C.; et al. An Outbreak of Hantavirus Pulmonary Syndrome, Chile, 1997. Emerg. Infect. Dis. 1998, 4, 687–694. [Google Scholar] [CrossRef]
- López, N.; Padula, P.; Rossi, C.; Lázaro, M.E.; Franze-Fernández, M.T. Genetic Identification of a New Hantavirus Causing Severe Pulmonary Syndrome in Argentina. Virology 1996, 220, 223–226. [Google Scholar] [CrossRef]
- Padula, P.J.; Edelstein, A.; Miguel, S.D.L.; López, N.M.; Rossi, C.M.; Rabinovich, R.D. Hantavirus Pulmonary Syndrome Outbreak in Argentina: Molecular Evidence for Person-to-Person Transmission of Andes Virus. Virology 1998, 241, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Ferrés, M.; Vial, P.; Marco, C.; Yañez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.J.; Mertz, G.J.; et al. Prospective Evaluation of Household Contacts of Persons with Hantavirus Cardiopulmonary Syndrome in Chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, V.P.; Bellomo, C.; San Juan, J.; Pinna, D.; Forlenza, R.; Elder, M.; Padula, P.J. Person-to-Person Transmission of Andes Virus. Emerg. Infect. Dis. 2005, 11, 1848–1853. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M.; Moreli, M.L.; De Sousa, R.L.M.; Borges, A.A.; De Figueiredo, G.G.; Machado, A.M.; Bisordi, I.; Nagasse-Sugahara, T.K.; Suzuki, A.; Pereira, L.E.; et al. Hantavirus Pulmonary Syndrome, Central Plateau, Southeastern, and Southern Brazil. Emerg. Infect. Dis. 2009, 15, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Bisordi, I.; Levis, S.; Garcia, J.; Pereira, L.E.; Souza, R.P.; Sugahara, T.K.N.; Pini, N.; Enria, D.; Souza, L.T.M. Identifying Rodent Hantavirus Reservoirs, Brazil. Emerg. Infect. Dis. 2004, 10, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.; Guterres, A.; Fernandes, J.; D’Andrea, P.; Bonvicino, C.; De Lemos, E. Hantavirus Reservoirs: Current Status with an Emphasis on Data from Brazil. Viruses 2014, 6, 1929–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guterres, A.; De Oliveira, R.C.; Fernandes, J.; Maia, R.M.; Teixeira, B.R.; Oliveira, F.C.G.; Bonvicino, C.R.; D’Andrea, P.S.; Schrago, C.G.; De Lemos, E.R.S. Co-circulation of Araraquara and Juquitiba Hantavirus in Brazilian Cerrado. Microb. Ecol. 2018, 75, 783–789. [Google Scholar] [CrossRef]
- Vasconcelos, M.I.; Lima, V.P.; Iversson, L.B.; Rosa, M.D.; da Rosa, A.P.; da Rosa, E.S.; Pereira, L.E.; Nassar, E.; Katz, G.; Matida, L.H.; et al. Hantavirus pulmonary syndrome in the rural area of Juquitiba, São Paulo metropolitan area, Brazil. Rev. Do Inst. Med. Trop. São Paulo 1997, 39, 237–238. [Google Scholar] [CrossRef] [Green Version]
- Padula, P.; Martinez, V.P.; Bellomo, C.; Maidana, S.; San Juan, J.; Tagliaferri, P.; Bargardi, S.; Vazquez, C.; Colucci, N.; Estévez, J.; et al. Pathogenic Hantaviruses, Northeastern Argentina and Eastern Paraguay. Emerg. Infect. Dis. 2007, 13, 1211–1214. [Google Scholar] [CrossRef]
- Johnson, A.M.; Bowen, M.D.; Ksiazek, T.G.; Williams, R.J.; Bryan, R.T.; Mills, J.N.; Peters, C.J.; Nichol, S.T. Laguna Negra Virus Associated with HPS in Western Paraguay and Bolivia. Virology 1997, 238, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Levis, S.; Morzunov, S.P.; Rowe, J.E.; Enria, D.; Pini, N.; Calderon, G.; Sabattini, M.; Jeor, S.S.C. Genetic Diversity and Epidemiology of Hantaviruses in Argentina. J. Infect. Dis. 1998, 177, 529–538. [Google Scholar] [CrossRef]
- Chu, Y.K.; Milligan, B.; Goodin, D.G.; Owen, R.D.; Jonsson, C.B. Phlyogenetic and geographical relationships of hantavirus strains in eastern and western Paraguay. Am. J. Trop. Med. Hyg. 2006, 75, 1127–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, P.C.; Ittig, R.E.G.; Fraire, H.J.R.; Levis, S.; Gardenal, C.N. Molecular identification and phylogenetic relationships among the species of the genus Oligoryzomys (Rodentia, Cricetidae) present in Argentina, putative reservoirs of hantaviruses. Zool. Scr. 2007, 36, 231–239. [Google Scholar] [CrossRef]
- Chu, Y.-K.; Jonsson, C.B.; Gonzalez, L.M.; Owen, R.D. The complex ecology of hantaviruses in Paraguay. Am. J. Trop. Med. Hyg. 2003, 69, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellomo, C.M.; Alonso, D.; Ricardo, T.; Coelho, R.; Kehl, S.; Periolo, N.; Casas, N.; Bergero, L.C.; Previtali, M.A.; Martinez, V.P. Emerging Hantaviruses in Central Argentina: First case of Hantavirus Pulmonary Syndrome caused by Alto Paraguay Virus and a novel orthohantavirus in Scapteromys aquaticus rodent. BioRxiv 2021. [Google Scholar] [CrossRef]
- Chu, Y.-K.; Goodin, D.; Owen, R.D.; Koch, D.; Jonsson, C.B. Sympatry of 2 Hantavirus Strains, Paraguay, 2003–2007. Emerg. Infect. Dis. 2009, 15, 1977–1980. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-K.; Owen, R.D.; Jonsson, C.B. Phylogenetic exploration of hantaviruses in paraguay reveals reassortment and host switching in South America. Virol. J. 2011, 8, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastwood, G.; Camp, J.V.; Chu, Y.K.; Sawyer, A.M.; Owen, R.D.; Cao, X.; Taylor, M.K.; Valdivieso-Torres, L.; Sage, R.D.; Yu, A.; et al. Habitat, species richness and hantaviruses of sigmodontine rodents within the Interior Atlantic Forest, Paraguay. PLoS ONE 2018, 13, e0201307. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.F.; Hjelle, B.; Jonsson, C.B.; Torrez-Martinez, N.; Galligan, D.; Esteban, E.; Callahan, J.; Basombrio, M.A.; Peralta-Ramos, M.; Bharadwaj, M.; et al. High prevalence of hantavirus infection in Indian communities of the Paraguayan and Argentinean Gran Chaco. Am. J. Trop. Med. Hyg. 1998, 59, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, J.F.; Galligan, D.; Esteban, E.; Rey, V.; Murua, A.; Gutierrez, S.; Gonzalez, L.; Thakuri, M.; Feldman, L.; Poiesz, B.; et al. Hantavirus infection in people inhabiting a highly endemic region of the Gran Chaco territory, Paraguay: Association with Trypanosoma cruzi infection, epidemiological features and haematological characteristics. Ann. Trop. Med. Parasitol. 2003, 97, 269–280. [Google Scholar] [CrossRef]
- Yates, T.L.; Mills, J.N.; Parmenter, C.A.; Ksiazek, T.G.; Parmenter, R.R.; Vande Castle, J.R.; Calisher, C.H.; Nichol, S.T.; Abbott, K.D.; Young, J.C.; et al. The Ecology and Evolutionary History of an Emergent Disease: Hantavirus Pulmonary Syndrome. BioScience 2002, 52, 989. [Google Scholar] [CrossRef] [Green Version]
- Prist, P.R.; D’Andrea, P.S.; Metzger, J.P. Landscape, Climate and Hantavirus Cardiopulmonary Syndrome Outbreaks. EcoHealth 2017, 14, 614–629. [Google Scholar] [CrossRef]
- Clement, J.; Vercauteren, J.; Verstraeten, W.W.; Ducoffre, G.; Barrios, J.M.; Vandamme, A.-M.; Maes, P.; Van Ranst, M. Relating increasing hantavirus incidences to the changing climate: The mast connection. Int. J. Health Geogr. 2009, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Stenseth, N.C. The ecological dynamics of hantavirus diseases: From environmental variability to disease prevention largely based on data from China. PLoS Negl. Trop. Dis. 2019, 13, e0006901. [Google Scholar] [CrossRef] [Green Version]
- Douglass, R.J.; Semmens, W.J.; Matlock-Cooley, S.J.; Kuenzi, A.J. Deer Mouse Movements in Peridomestic and Sylvan Settings in Relation to Sin Nombre Virus Antibody Prevalence. J. Wildl. Dis. 2006, 42, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Suzán, G.; Marcé, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armién, B.; Pascale, J.M.; Yates, T.L. Experimental Evidence for Reduced Rodent Diversity Causing Increased Hantavirus Prevalence. PLoS ONE 2009, 4, e5461. [Google Scholar] [CrossRef] [PubMed]
- Orrock, J.L.; Allan, B.F.; Drost, C.A. Biogeographic and Ecological Regulation of Disease: Prevalence of Sin Nombre Virus in Island Mice Is Related to Island Area, Precipitation, and Predator Richness. Am. Nat. 2011, 177, 691–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niklasson, B.; Hornfeldt, B.; Lundkvist, A.; Leduc, J.; Bjorsten, S. Temporal Dynamics of Puumala Virus Antibody Prevalence in Voles and of Nephropathia Epidemica Incidence in Humans. Am. J. Trop. Med. Hyg. 1995, 53, 134–140. [Google Scholar] [CrossRef] [PubMed]
- French, R.K.; Holmes, E.C. An Ecosystems Perspective on Virus Evolution and Emergence. Cell Press Trends Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodin, D.G.; Koch, D.E.; Owen, R.D.; Chu, Y.-K.; Hutchinson, J.M.S.; Jonsson, C.B. Land cover associated with hantavirus presence in Paraguay. Glob. Ecol. Biogeogr. 2006, 15, 519–527. [Google Scholar] [CrossRef]
- Goodin, D.G.; Paige, R.; Owen, R.D.; Ghimire, K.; Koch, D.E.; Chu, Y.-K.; Jonsson, C.B. Microhabitat characteristics of Akodon montensis, a reservoir for hantavirus, and hantaviral seroprevalence in an Atlantic forest site in eastern Paraguay. J. Vector Ecol. 2009, 34, 104–113. [Google Scholar] [CrossRef]
- Suzán, G.; Marcé, E.; Giermakowski, J.T.; Armién, B.; Pascale, J.; Mills, J.; Ceballos, G.; Gómez, A.; Aguirre, A.A.; Salazar-Bravo, J.; et al. The Effect of Habitat Fragmentation and Species Diversity Loss on Hantavirus Prevalence in Panama. Ann. N. Y. Acad. Sci. 2008, 1149, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzán, G.; Armién, A.; Mills, J.N.; Marcé, E.; Ceballos, G.; Ávila, M.; Salazar-Bravo, J.; Ruedas, L.; Armién, B.; Yates, T.L. Epidemiological considerations of rodent community composition in fragmented landscapes in Panama. J. Mammal. 2008, 89, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Suzán, G.; Marcé, E.; Yates, T.L.; Giermakowski, J.T.; Armién, B.; Suzán-Azpiri, H. Modeling hantavirus reservoir species dominance in high seroprevalence areas on the Azuero Peninsula of Panama. Am. J. Trop. Med. Hyg. 2006, 74, 1103–1110. [Google Scholar] [CrossRef]
- Ruedas, L.A.; Salazar–Bravo, J.; Tinnin, D.S.; Armién, B.; Cáceres, C.; García, A.; Díaz, M.A.; Gracia, F.; Suzán, G.; Peters, C.J.; et al. Community ecology of small mammal populations in Panamá following an outbreak of Hantavirus pulmonary syndrome. J. Vector Ecol. 2003, 29, 177–191. [Google Scholar]
- Owen, R.D.; Goodin, D.G.; Koch, D.E.; Chu, Y.-K.; Jonsson, C.B. Spatiotemporal variation in Akodon montensis (Cricetidae: Sigmodontinae) and hantaviral seroprevalence in a subtropical forest ecosystem. J. Mammal. 2010, 91, 467–481. [Google Scholar] [CrossRef] [Green Version]
- Camp, J.V.; Spruill-Harrell, B.; Owen, R.D.; Solà-Riera, C.; Williams, E.P.; Eastwood, G.; Sawyer, A.M.; Jonsson, C.B. Mixed Effects of Habitat Degradation and Resources on Hantaviruses in Sympatric Wild Rodent Reservoirs within a Neotropical Forest. Viruses 2021, 13, 85. [Google Scholar] [CrossRef]
- Muylaert, R.L.; Bovendorp, R.S.; Sabino-Santos, G.; Prist, P.R.; Melo, G.L.; Priante, C.D.F.; Wilkinson, D.A.; Ribeiro, M.C.; Hayman, D.T.S. Hantavirus host assemblages and human disease in the Atlantic Forest. PLoS Negl. Trop. Dis. 2019, 13, e0007655. [Google Scholar] [CrossRef] [Green Version]
- Pardini, R. Effects of forest fragmentation on small mammals in an Atlantic Forest landscape. Biodivers. Conserv. 2004, 13, 2567–2586. [Google Scholar] [CrossRef]
- Katz, G.; Williams, R.J.; Burt, M.S.; de Souza, L.T.; Pereira, L.E.; Mills, J.N.; Suzuki, A.; Ferreira, I.B.; Souza, R.P.; Alves, V.A.; et al. Hantavirus Pulmonary Syndrome in the State of São Paulo, Brazil, 1993–1998. Vector Borne Zoonotic Dis. 2001, 1, 181–190. [Google Scholar] [CrossRef]
- Rubio, A.V.; Ávila-Flores, R.; Suzán, G. Responses of Small Mammals to Habitat Fragmentation: Epidemiological Considerations for Rodent-Borne Hantaviruses in the Americas. EcoHealth 2014, 11, 526–533. [Google Scholar] [CrossRef]
- De Castro, E.C. The Atlantic Forest of South America: Biodiversity Status, Threats and Outlook; Leal, C.G., de Gusmão Câmara, I., Eds.; Island Press: Washington, DC, USA, 2003; Volume 1, pp. 1–488. [Google Scholar]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Kim, S.; Altstatt, A.; Townshend, J.R.G.; Davis, P.; Song, K.; Tucker, C.J.; Rodas, O.; Yanosky, A.; Clay, R.; et al. Rapid loss of Paraguay’s Atlantic forest and the status of protected areas—A Landsat assessment. Remote Sens. Environ. 2007, 106, 460–466. [Google Scholar] [CrossRef]
- Huang, C.; Kim, S.; Song, K.; Townshend, J.R.G.; Davis, P.; Altstatt, A.; Rodas, O.; Yanosky, A.; Clay, R.; Tucker, C.J.; et al. Assessment of Paraguay’s forest cover change using Landsat observations. Glob. Planet. Chang. 2009, 67, 1–12. [Google Scholar] [CrossRef]
- Owen, R.D.; Camp, J.V.; Sage, R.; Rodríguez, L.; Martínez Bruyn, V.J.; McAllister, R.C.; Jonsson, C.B. Sympatry and habitat associations of sigmodontine rodents in a neotropical forest-savanna interface. Mammalia 2019, 84, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, H.P.; Sundell, J.; Ecke, F.; Halle, S.; Haapakoski, M.; Henttonen, H.; Huitu, O.; Jacob, J.; Johnsen, K.; Koskela, E.; et al. Population cycles and outbreaks of small rodents: Ten essential questions we still need to solve. Oecologia 2021, 195, 601–622. [Google Scholar] [CrossRef]
- Duffy, J.E. Biodiversity and ecosystem function: The consumer connection. Oikos 2002, 99, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Ordiz, A.; Aronsson, M.; Persson, J.; Støen, O.-G.; Swenson, J.E.; Kindberg, J. Effects of Human Disturbance on Terrestrial Apex Predators. Diversity 2021, 13, 68. [Google Scholar] [CrossRef]
- Hayward, M.W.; Kerley, G.I.H. Fencing for conservation: Restriction of evolutionary potential or a riposte to threatening processes? Biol. Conserv. 2009, 142, 1–13. [Google Scholar] [CrossRef]
- Zuercher, G.L.; Owen, R.D.; Torres, J.; Gipson, P.S. Mechanisms of coexistence in a diverse Neotropical mammalian carnivore community. J. Mammal. in press.
- Zuercher, G.L.; Gipson, P.S.; Stewart, G.C. Identification of Carnivore Feces by Local Peoples and Molecular Analyses. Wildl. Soc. Bullentin 2003, 31, 961–970. [Google Scholar]
- Santori, R.T.; Astua De Moraes, D.; Cerqueira, R. Diet composition of Metachirus nudicaudatus and Didelphis aurita (Marsupialia, Didelphoidea) in southeastern Brazil. Mammalia 1995, 60, 511–516. [Google Scholar] [CrossRef]
- Cacciali, P.; Bauer, F.; Martínez, N. Herpetofauna de la Reserva Natural del Bosque Mbaracayú, Paraguay. Kempffiana 2015, 11, 29–47. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Quintana, J.; Morse, S. Social interactions and resource ownership in two private protected areas of Paraguay. J. Environ. Manag. 2005, 77, 64–78. [Google Scholar] [CrossRef]
- D’Elia, G.; Pardinas, U.F.J. Subfamily Sigmodontinae Wagner, 1843. In Mammals of South America; The University of Chicago Press: Chicago, IL, USA, 2015; Volume 2, pp. 63–687. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 July 2021).
- Huggins, R.M. On the statistical analysis of capture experiments. Biometrika 1989, 76, 133–140. [Google Scholar] [CrossRef]
- Otis, D.; Burnham, K.; White, G.; Anderson, D. Statistical Inference from Capture Data on Closed Animal Populations; Wildlife Society: Washington, DC, USA, 1978; Volume 62. [Google Scholar]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, S120–S139. [Google Scholar] [CrossRef]
- Gorbunova, V.; Bozzella, M.J.; Seluanov, A. Rodents for comparative aging studies: From mice to beavers. AGE 2008, 30, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 July 2021).
- Lande, R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 1996, 76, 5–13. [Google Scholar] [CrossRef]
- Heinze, G.; Ploner, M.; Jiricka, L. logistf: Firth’s Bias-Reduced Logistic Regression, R Package Version 1.24. 2020. Available online: https://CRAN.R-project.org/package=logistf (accessed on 16 September 2021).
- King, G.; Zeng, L. Logistic Regression in Rare Events Data. Political Anal. 2001, 9, 137–163. [Google Scholar] [CrossRef] [Green Version]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N. Biodiversity loss and emerging infectious disease: An example from the rodent-borne hemorrhagic fevers. Biodiversity 2006, 7, 9–17. [Google Scholar] [CrossRef]
- Williams, E.P.; Spruill-Harrell, B.M.; Taylor, M.K.; Lee, J.; Nywening, A.V.; Yang, Z.; Nichols, J.H.; Camp, J.V.; Owen, R.D.; Jonsson, C.B. Common Themes in Zoonotic Spillover and Disease Emergence: Lessons Learned from Bat- and Rodent-Borne RNA Viruses. Viruses 2021, 13, 1509. [Google Scholar] [CrossRef]
- Mark, E.J.; Woolhouse, K.A.; Brierley, L. RNA Viruses: A Case Study of the Biology of Emerging Infectious Diseases; American Society of Microbiology: Washington, DC, USA, 2014; pp. 83–97. [Google Scholar]
- Woolhouse, M.E.J.; Brierley, L. Epidemiological characteristics of human-infective RNA viruses. Sci. Data 2018, 5, 180017. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of acute illness--southwestern United States, 1993. MMWR. Morb. Mortal. Wkly. Rep. 1993, 42, 421–424. [Google Scholar]
- Johnson, A.M.; de Souza, L.T.M.; Ferreira, I.B.; Pereira, L.E.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J.; Nichol, S.T. Genetic investigation of novel hantaviruses causing fatal HPS in Brazil. J. Med. Virol. 1999, 59, 527–535. [Google Scholar] [CrossRef]
- Mills, J.N.; Ellis, B.A.; McKee, K.T., Jr.; Calderon, G.E.; Maiztegui, J.I.; Nelson, G.O.; Ksiazek, T.G.; Peters, C.J.; Childs, J.E. A Longitudinal Study of Junin Virus Activity in the Rodent Reservoir of Agrentine Hemorrhagic Fever. Am. J. Trop. Med. Hyg. 1992, 47, 749–763. [Google Scholar] [CrossRef]
- Olayemi, A.; Cadar, D.; Magassouba, N.F.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New Hosts of The Lassa Virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef] [Green Version]
- Dan-Nwafor, C.C.; Ipadeola, O.; Smout, E.; Ilori, E.; Adeyemo, A.; Umeokonkwo, C.; Nwidi, D.; Nwachukwu, W.; Ukponu, W.; Omabe, E.; et al. A cluster of nosocomial Lassa fever cases in a tertiary health facility in Nigeria: Description and lessons learned, 2018. Int. J. Infect. Dis. 2019, 83, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Escalera-Antezana, J.P.; Rodriguez-Villena, O.J.; Arancibia-Alba, A.W.; Alvarado-Arnez, L.E.; Bonilla-Aldana, D.K.; Rodríguez-Morales, A.J. Clinical features of fatal cases of Chapare virus hemorrhagic fever originating from rural La Paz, Bolivia, 2019: A cluster analysis. Travel Med. Infect. Dis. 2020, 36, 101589. [Google Scholar] [CrossRef] [PubMed]
- Mull, N.; Jackson, R.; Sironen, T.; Forbes, K.M. Ecology of Neglected Rodent-Borne American Orthohantaviruses. Pathogens 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Bernshtein, A.D.; Apekina, N.S.; Mikhailova, T.V.; Myasnikov, Y.A.; Khlyap, L.A.; Korotkov, Y.S.; Gavrilovskaya, I.N. Dynamics of Puumala hantavirus infection in naturally infected bank voles (Clethrinomys glareolus). Arch. Virol. 1999, 144, 2415–2428. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Sweeney, W.; Mills, J.N.; Beaty, B.J. Natural History of Sin Nombre Virus in Western Colorado. Emerg. Infect. Dis. 1999, 5, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Douglass, R.J.; Semmens, W.J.; Mills, J.N.; Zanto, S.N.; Bond, C.W.; Wilson, T.; Van Horn, R.C. Longitudinal studies of Sin Nombre virus in deer mouse-dominated ecosystems of Montana. Am. J. Trop. Med. Hyg. 2001, 65, 33–41. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, N.E.; Chu, Y.K.; Owen, R.D.; Abuzeineh, A.; De la Sancha, N.; Dick, C.W.; Holsomback, T.; Nisbett, R.A.; Jonsson, C. A longitudinal study of Bayou virus, hosts, and habitat. Am. J. Trop. Med. Hyg. 2005, 73, 1043–1049. [Google Scholar] [CrossRef]
- Evander, M.; Ahlm, C. Milder winters in northern Scandinavia may contribute to larger outbreaks of haemorrhagic fever virus. Glob. Health Action 2009, 2, 2020. [Google Scholar] [CrossRef]
- Glass, G.E.; Yates, T.L.; Fine, J.B.; Shields, T.M.; Kendall, J.B.; Hope, A.G.; Parmenter, C.A.; Peters, C.J.; Ksiazek, T.G.; Li, C.S.; et al. Satellite imagery characterizes local animal reservoir populations of Sin Nombre virus in the southwestern United States. Proc. Natl. Acad. Sci. USA 2002, 99, 16817–16822. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.N.; Ksiazek, T.G.; Peters, C.J.; Childs, J.E. Long-Term Studies of Hantavirus Reservoir Populations in the Southwestern United States: A Synthesis. Emerg. Infect. Dis. 1999, 5, 135–142. [Google Scholar] [CrossRef]
- Mills, J.N.; Ksiazek, T.G.; Ellis, B.A.; Rollin, P.E.; Nichol, S.T.; Yates, T.L.; Gannon, W.L.; Levy, C.E.; Engelthaler, D.M.; Davis, T.; et al. Patterns of Association with Host and Habitat: Antibody Reactive with Sin Nombre Virus in Small Mammals in the Major Biotic Communities of the Southwestern United States. Am. J. Trop. Med. Hyg. 1997, 56, 273–284. [Google Scholar] [CrossRef]
- Gubler, D.J.; Reiter, P.; Ebi, K.L.; Yap, W.; Nasci, R.; Patz, J.A. Climate variability and change in the United States: Potential impacts on vector- and rodent-borne diseases. Environ. Health Perspect. 2001, 109, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.J.S.; McCormack, R.K.; Jonsson, C.B. Mathematical Models for Hantavirus Infection in Rodents. Bull. Math. Biol. 2006, 68, 511–524. [Google Scholar] [CrossRef]
- Allen, L.J.S.; Wesley, C.L.; Owen, R.D.; Goodin, D.G.; Koch, D.; Jonsson, C.B.; Chu, Y.-K.; Shawn Hutchinson, J.M.; Paige, R.L. A habitat-based model for the spread of hantavirus between reservoir and spillover species. J. Theor. Biol. 2009, 260, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Wesley, C.L.; Allen, L.J.S.; Jonsson, C.B.; Chu, Y.-K.; Owen, R.D. A discrete-time rodent-hantavirus model structured by infection and developmental stages. Adv. Stud. Pure Math. 2009, 2009, 387–398. [Google Scholar] [CrossRef]
- Gray, A.; Greenhalgh, D.; Hu, L.; Mao, X.; Pan, J. A Stochastic Differential Equation SIS Epidemic Model. J. Appl. Math. 2011, 71, 876–902. [Google Scholar] [CrossRef] [Green Version]
- McCormack, R.; Allen, L.J.S. Stochastic SIS and SIR Multihost Epidemic Models. In Proceedings of the Conference on Differential and Difference Equations and Applications, Rajecké Teplice, Slovakia, 26–30 June 2006; Hindawi Publishing Corporation: New York, NY, USA, 2006. [Google Scholar]
- McCormack, R.K.; Allen, L.J.S. Multi-patch deterministic and stochastic models for wildlife diseases. J. Biol. Dyn. 2007, 1, 63–85. [Google Scholar] [CrossRef]
- Allen, E.J.; Allen, L.J.S.; Arciniega, A.; Greenwood, P.E. Construction of Equivalent Stochastic Differential Equation Models. Stoch. Anal. Appl. 2008, 26, 274–297. [Google Scholar] [CrossRef]
- Wesley, C.L.; Allen, L.J.S. The basic reproduction number in epidemic models with periodic demographics. J. Biol. Dyn. 2009, 3, 116–129. [Google Scholar] [CrossRef]
- Banerjee, C.; Allen, L.J.S.; Salazar-Bravo, J. Models for an arenavirus infection in a rodent population: Consequences of horizontal, vertical and sexual transmission. Math. Biosci. Eng. 2008, 5, 617–645. [Google Scholar] [CrossRef]
- Wesley, C.L.; Allen, L.J.S.; Langlais, M. Models for the spread and persistence of hantavirus infection in rodents with direct and indirect transmission. Math. Biosci. Eng. 2010, 7, 195–211. [Google Scholar] [CrossRef]
- Holsomback, T.S.; McIntyre, N.E.; Nisbett, R.A.; Strauss, R.E.; Chu, Y.-K.; Abuzeineh, A.A.; De La Sancha, N.; Dick, C.W.; Jonsson, C.B.; Morris, B.E.L. Bayou virus detected in non-oryzomyine rodent hosts: An assessment of habitat composition, reservoir community structure, and marsh rice rat social dynamics. J. Vector Ecol. 2009, 34, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Andreo, V.; Provensal, C.; Levis, S.; Pini, N.; Enría, D.; Polop, J. Summer—autumn distribution and abundance of the hantavirus host, Oligoryzomys longicaudatus, in northwestern Chubut, Argentina. J. Mammal. 2012, 93, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Palma, R.E.; Polop, J.J.; Owen, R.D.; Mills, J.N. Ecology of rodent-associated hantaviruses in the Southern Cone of South America: Argentina, Chile, Paraguay, and Uruguay. J. Wildl. Dis. 2012, 48, 267–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto Cáceres, M.B.; Owen, R.D. Relación de los pequeños mamíferos terrestres (Rodentia y Didelphimorphia) con la estructura de la vegetación en el Bosque Atlántico Interior—Un análisis multivariado. Therya 2019, 10, 359–369. [Google Scholar] [CrossRef]
- Owen, R.; Camp, J.V.; Jonsson, C.B. Sigmodontine community and species responses to El Niño and precipitation in different levels of forest degradation. Therya 2019, 10, 255–265. [Google Scholar] [CrossRef]
- Owen, R.D.; SÁnchez, H.; RodrÍguez, L.; Jonsson, C.B. Composition and characteristics of a diverse didelphid community (Mammalia: Didelphimorphia) in sub-tropical South America. Occas. Pap. Tex. Tech. Univ. Mus. 2018, 2018, 358. [Google Scholar]
- Haapakoski, M.; Sundell, J.; Ylönen, H. Predation risk and food: Opposite effects on overwintering survival and onset of breeding in a boreal rodent. J. Anim. Ecol. 2012, 81, 1183–1192. [Google Scholar] [CrossRef]
- Orrock, J.L. Rodent foraging is affected by indirect, but not by direct, cues of predation risk. Behav. Ecol. 2004, 15, 433–437. [Google Scholar] [CrossRef] [Green Version]
- Fanson, B.G. Effect of Direct and Indirect Cues of Predation Risk on the Foraging Behavior of the White-footed Mouse (Peromyscus leucopus). Northeast. Nat. 2010, 17, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Borowski, Z.; Owadowska, E. Field vole (Microtus agrestis) seasonal spacing behavior: The effect of predation risk by mustelids. Naturwissenschaften 2010, 97, 487–493. [Google Scholar] [CrossRef]
- Dupuch, A.; Morris, D.W.; Ale, S.B.; Wilson, D.J.; Moore, D.E. Landscapes of fear or competition? Predation did not alter habitat choice by Arctic rodents. Oecologia 2014, 174, 403–412. [Google Scholar] [CrossRef]
- Korpimäki, E.; Norrdahl, K.; Huitu, O.; Klemola, T. Predator–induced synchrony in population oscillations of coexisting small mammal species. Proc. R. Soc. B Biol. Sci. 2005, 272, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, J.L.; Pearson, D.E.; Fletcher, R.J. Counterintuitive effects of large-scale predator removal on a midlatitude rodent community. Ecology 2010, 91, 3719–3728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selonen, V.; Remm, J.; Hanski, I.K.; Henttonen, H.; Huitu, O.; Jokinen, M.; Korpimäki, E.; Mäkelä, A.; Sulkava, R.; Wistbacka, R. Population fluctuations and spatial synchrony in an arboreal rodent. Oecologia 2019, 191, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, M.; Stenseth, N.C.; Jaksic, F.M. Population dynamics of a South American rodent: Seasonal structure interacting with climate, density dependence and predator effects. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 2579–2586. [Google Scholar] [CrossRef] [Green Version]
- Stryjek, R.; Mioduszewska, B.; Spaltabaka-Gędek, E.; Juszczak, G.R. Wild Norway Rats Do Not Avoid Predator Scents When Collecting Food in a Familiar Habitat: A Field Study. Sci. Rep. 2018, 8, 9475. [Google Scholar] [CrossRef] [PubMed]
- Cozzoli, F.; Gjoni, V.; Basset, A. Size dependency of patch departure behavior: Evidence from granivorous rodents. Ecology 2019, 100, e02800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos, V.O.; Contreras, L.C.; Meserve, P.L.; Gutiérrez, J.R.; Jaksic, F.M. Effects of predation risk on space use by small mammals: A field experiment with a Neotropical rodent. Oikos 1995, 74, 259–264. [Google Scholar] [CrossRef]
- Püttker, T.; Pardini, R.; Meyer-Lucht, Y.; Sommer, S. Responses of five small mammal species to micro-scale variations in vegetation structure in secondary Atlantic Forest remnants, Brazil. BMC Ecol. 2008, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Lima, D.O.D.; Azambuja, B.O.; Camilotti, V.L.; Cáceres, N.C. Small mammal community structure and microhabitat use in the austral boundary of the Atlantic Forest, Brazil. Zoologia 2010, 27, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Dalmagro, A.D.; Vieira, E.M. Patterns of habitat utilization of small rodents in an area of Araucaria forest in Southern Brazil. Austral. Ecol. 2005, 30, 353–362. [Google Scholar] [CrossRef]
- Abreu, M.S.; Oliveira, L.R. Patterns of arboreal and terrestrial space use by non-volant small mammals in an Araucaria forest of southern Brazil. An. Acad. Bras. Ciências 2014, 86, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.F.; Marks, C.F.; Peres, B.; Melo, G.L.; Cáceres, N.C. Movement and use of environmental structures, climbing supports and shelters by Akodon montensis (Sigmodontinae, Rodentia) in the Atlantic Forest of southern Brazil. Mammalia 2020, 84, 107–113. [Google Scholar] [CrossRef]
- Puettker, T.; Meyer-Lucht, Y.; Sommer, S. Fragmentation effects on population density of three rodent species in secondary Atlantic Rainforest, Brazil. Stud. Neotrop. Fauna Environ. 2008, 43, 11–18. [Google Scholar] [CrossRef]
- Umetsu, F.; Pardini, R. Small mammals in a mosaic of forest remnants and anthropogenic habitats—evaluating matrix quality in an Atlantic forest landscape. Landsc. Ecol. 2007, 22, 517–530. [Google Scholar] [CrossRef]
- Gheler-Costa, C.; Vettorazzi, C.A.; Pardini, R.; Verdade, L.M. The distribution and abundance of small mammals in agroecosystems of southeastern Brazil. Mammalia 2012, 76, 185–191. [Google Scholar] [CrossRef]
- Prist, P.R.; Prado, A.; Tambosi, L.R.; Umetsu, F.; de Arruda Bueno, A.; Pardini, R.; Metzger, J.P. Moving to healthier landscapes: Forest restoration decreases the abundance of Hantavirus reservoir rodents in tropical forests. Sci. Total Environ. 2021, 752, 141967. [Google Scholar] [CrossRef]
- Pardini, R.; de Souza, S.M.; Braga-Neto, R.; Metzger, J.P. The role of forest structure, fragment size and corridors in maintaining small mammal abundance and diversity in an Atlantic forest landscape. Biol. Conserv. 2005, 124, 253–266. [Google Scholar] [CrossRef]
- Milholland, M.T.; Castro-Arellano, I.; Arellano, E.; Nava-García, E.; Rangel-Altamirano, G.; Gonzalez-Cozatl, F.X.; Suzán, G.; Schountz, T.; González-Padrón, S.; Vigueras, A.; et al. Species Identity Supersedes the Dilution Effect Concerning Hantavirus Prevalence at Sites across Texas and México. ILAR J. 2017, 58, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.C.D.; Guterres, A.; Schrago, C.G.; Fernandes, J.; Teixeira, B.R.; Zeccer, S.; Bonvicino, C.R.; D’Andrea, P.S.; Lemos, E.R.S.D. Detection of the first incidence of Akodon paranaensis naturally infected with the Jabora virus strain (Hantavirus) in Brazil. Memórias Do Inst. Oswaldo Cruz 2012, 107, 424–428. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, B.R.; Oliveira, R.C.; Guterres, A.; Bonvicino, C.R.; Fernandes, J.; Duarte Dos Santos, C.N.; Lemos, E.R.S.; D’Andrea, P.S.; Loureiro, N.; Mattos, L.H.B.V.; et al. Population Ecology of Hantavirus Rodent Hosts in Southern Brazil. Am. J. Trop. Med. Hyg. 2014, 91, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raboni, S.M.; De Araujo, M.R.; Delfraro, A.; Lemos, E.R.S.; Carstensen, S.; De Borba, L.; D’Andrea, P.S.; Rubio, G.; Teixeira, B.R.; Maron, A.; et al. Hantavirus Infection Prevalence in Wild Rodents and Human Anti-Hantavirus Serological Profiles from Different Geographic Areas of South Brazil. Am. J. Trop. Med. Hyg. 2012, 87, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moll, R.J.; Eaton, J.T.; Cepek, J.D.; Lorch, P.D.; Dennis, P.M.; Robison, T.; Tsao, J.; Montgomery, R.A. Dynamic rodent behavioral response to predation risk: Implications for disease ecology. Oecologia 2020, 192, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Holt, R.D. Are predators good for your health? Evaluating evidence for top-down regulation of zoonotic disease reservoirs. Front. Ecol. Environ. 2004, 2, 13–20. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
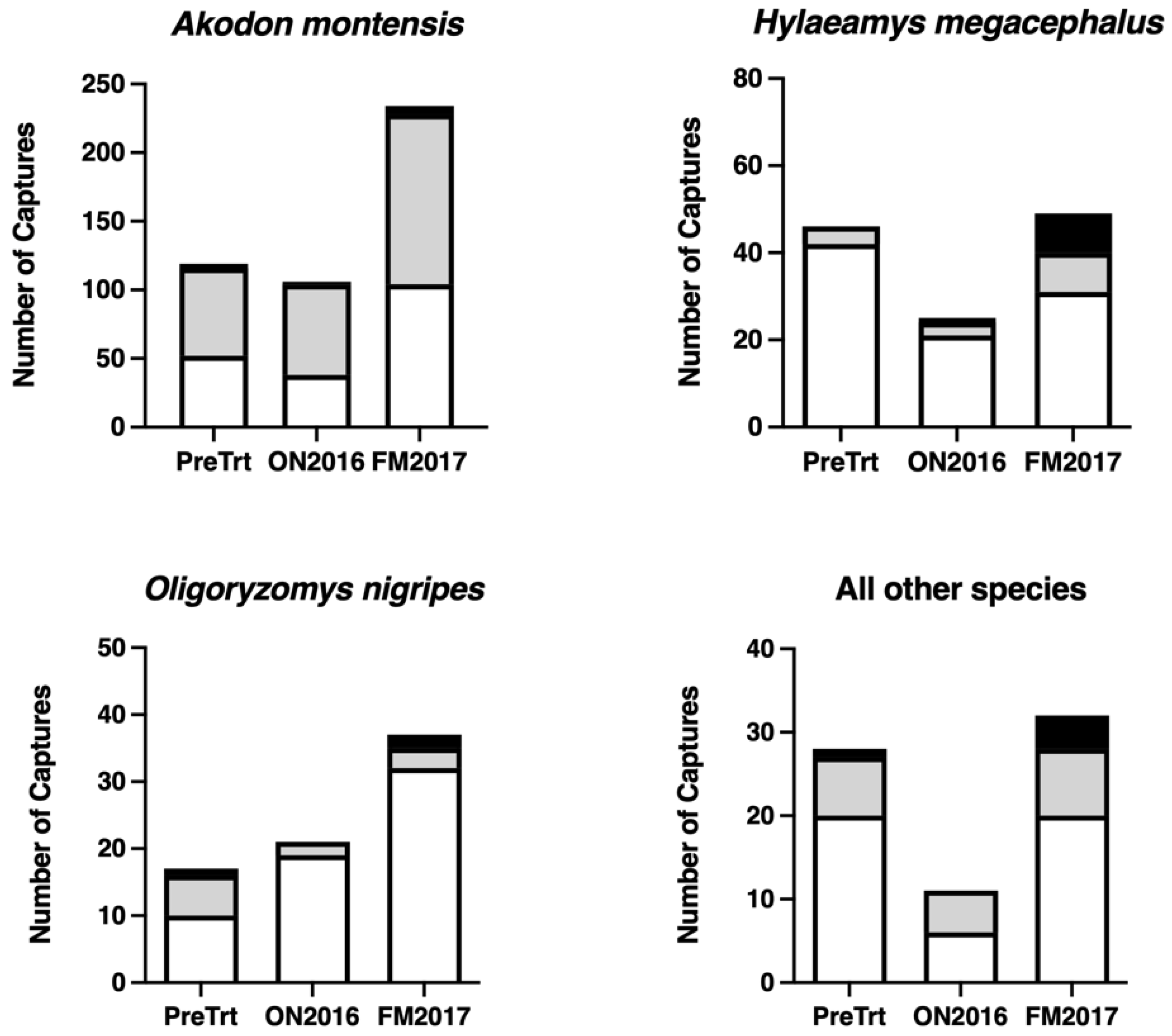{kind=link}
| PreTrt | ON2016 | FM2017 | Total | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grid | B | A | G | H | D | C | B | A | G | H | D | C | B | A | G | H | D | C | |
| Enclosure | − | − | − | − | − | − | − | − | − | + | + | + | − | − | − | + | + | + | |
| Predator Removal | − | − | − | − | − | − | − | − | − | + | + | + | − | − | − | + | + | + | |
| Degradation Level | Least | Moderate | Most | Least | Moderate | Most | Least | Moderate | Most | Least | Moderate | Most | Least | Moderate | Most | Least | Moderate | Most | |
| Species | |||||||||||||||||||
| Akodon montensis | 27 | 28 | 16 | 21 | 7 | 20 | 13 | 18 | 12 | 18 | 15 | 30 | 42 | 61 | 18 | 48 | 10 | 55 | 459 |
| Calomys callosus | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 3 |
| Euryoryzomys russatus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Hylaeamys megacephalus | 4 | 12 | 7 | 4 | 6 | 13 | 1 | 7 | 3 | 5 | 5 | 4 | 7 | 15 | 2 | 7 | 8 | 10 | 120 |
| Juliomys pictipes | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 2 |
| Nectomys squamipes | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| Oecomys mamorae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 |
| Oligoryzomys flavescens | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 2 | 9 |
| Oligoryzomys mattogrossae | 4 | 2 | 3 | 0 | 1 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 7 | 4 | 3 | 0 | 3 | 37 |
| Oligoryzomys nigripes | 3 | 6 | 3 | 0 | 0 | 5 | 2 | 1 | 5 | 1 | 5 | 7 | 4 | 14 | 3 | 0 | 3 | 13 | 75 |
| Scapteromys aquaticus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Sooretamys angouya | 1 | 0 | 1 | 0 | 0 | 1 | 2 | 0 | 3 | 0 | 1 | 2 | 0 | 1 | 1 | 0 | 0 | 1 | 14 |
| Total | 40 | 50 | 32 | 25 | 14 | 49 | 18 | 27 | 23 | 25 | 26 | 44 | 55 | 101 | 30 | 60 | 22 | 84 | 725 |
| Species Richness | 6 | 5 | 6 | 2 | 3 | 6 | 4 | 4 | 4 | 4 | 4 | 5 | 5 | 6 | 7 | 4 | 4 | 6 | |
| Effect a | Level | Estimate (β) | SE | p |
|---|---|---|---|---|
| Treatment | Yes | −0.043 | 0.040 | 0.3950 |
| Session | ON2016 | −0.040 | 0.044 | 0.3884 |
| FM2017 | −0.045 | 0.044 | 0.3343 | |
| Degradation Level | Moderately | 0.178 | 0.049 | 0.0689 |
| Most | 0.212 | 0.049 | 0.0504 |
| Akodon montensis | Oligoryzomys nigripes | Total | |||||
|---|---|---|---|---|---|---|---|
| Variable | Level | Pos/Tested | % Pos | Pos/Tested | % Pos | Pos/Tested | % Pos |
| Session a | PreTrt | 2/100 | 2.0% | 2/13 | 15.4% | 4/167 | 2.4% |
| ON2016 | 5/74 | 6.8% | 3/20 | 15.0% | 8/124 | 6.5% | |
| FM2017 | 7/228 | 3.1% | 5/33 | 15.2% | 12/341 | 3.5% | |
| Degradation Level | Least | 1/146 | 0.7% | 1/8 | 12.5% | 2/190 | 1.1% |
| Moderately | 4/126 | 3.2% | 3/27 | 11.1% | 7/216 | 3.2% | |
| Most | 9/130 | 6.9% | 6/31 | 19.4% | 15/226 | 6.6% | |
| Total | 14/402 | 3.5% | 10/66 | 15.2% | 24/632 | 3.8% | |
| Species | Predictor | Level | No. Neg (%) | No. Pos (%) | Odds Ratio a | 95% CI | p |
|---|---|---|---|---|---|---|---|
| A. montensis | Age | Adult | 175 (94%) | 11 (6%) | 3.847 | 1.251–15.297 | 0.0176 |
| Juvenile | 8 (100%) | 0 | 3.454 | 0.025–40.153 | 0.4877 | ||
| Subadult | 205 (99%) | 3 (1%) | |||||
| Sex | Male | 205 (94%) | 12 (6%) | 4.465 | 1.314–23.063 | 0.0144 | |
| Female | 183 (99%) | 2 (1%) | |||||
| Weight b | 388 (6~63) | 14 (14~59) | 1.135 | 1.083–1.196 | <0.0001 | ||
| Reprod. Condition | Active | 68 (92%) | 6 (8%) | 3.578 | 1.195–10.251 | 0.0240 | |
| Inactive | 320 (98%) | 8 (2%) | |||||
| Tail Scar | Yes | 135 (92%) | 11 (8%) | 6.123 | 1.988–24.367 | 0.0012 | |
| No | 252 (99%) | 3 (1%) | |||||
| O. nigripes | Age | Adult | 46 (82%) | 10 (18%) | 4.290 | 0.473–569.134 | 0.2358 |
| Juvenile | 1 (100%) | 0 | 6.333 | 0.029–1443.403 | 0.4150 | ||
| Subadult | 9 (100%) | 0 | |||||
| Sex | Male | 33 (77%) | 10 (23%) | 14.731 | 1.742–1928.789 | 0.0083 | |
| Female | 23 (100%) | 0 | |||||
| Weight b | 56 (6~30) | 10 (19~31) | 1.264 | 1.052–1.577 | 0.0109 | ||
| Reprod. Condition | Active | 28 (87.5%) | 4 (12.5%) | 0.692 | 0.175–2.540 | 0.5796 | |
| Inactive | 28 (82%) | 6 (18%) | |||||
| Tail Scar | Yes | 1 (50%) | 1 (50%) | 5.842 | 0.440–78.007 | 0.1620 | |
| No | 55 (86%) | 9 (14%) |
| Species | Predictor a | Level | Odds Ratio a | 95% CI | p |
|---|---|---|---|---|---|
| A. montensis | Treatment | Yes | 0.679 | 0.198–1.981 | 0.4901 |
| Session | ON2016 | 3.118 | 0.728–17.749 | 0.1270 | |
| FM2017 | 1.334 | 0.349–7.239 | 0.6907 | ||
| Degradation Level | Moderately | 3.563 | 0.649–35.88 | 0.1488 | |
| Most | 7.584 | 1.713–71.41 | 0.0054 | ||
| O. nigripes | Treatment | Yes | 0.692 | 0.175–2.540 | 0.5796 |
| Session | ON2016 | 0.920 | 0.153–6.285 | 0.9272 | |
| FM2017 | 0.888 | 0.182–5.514 | 0.8877 | ||
| Degradation Level | Moderately | 0.714 | 0.097–8.211 | 0.7552 | |
| Most | 1.275 | 0.213–13.624 | 0.8046 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spruill-Harrell, B.; Pérez-Umphrey, A.; Valdivieso-Torres, L.; Cao, X.; Owen, R.D.; Jonsson, C.B. Impact of Predator Exclusion and Habitat on Seroprevalence of New World Orthohantavirus Harbored by Two Sympatric Rodents within the Interior Atlantic Forest. Viruses 2021, 13, 1963. https://doi.org/10.3390/v13101963
Spruill-Harrell B, Pérez-Umphrey A, Valdivieso-Torres L, Cao X, Owen RD, Jonsson CB. Impact of Predator Exclusion and Habitat on Seroprevalence of New World Orthohantavirus Harbored by Two Sympatric Rodents within the Interior Atlantic Forest. Viruses. 2021; 13(10):1963. https://doi.org/10.3390/v13101963
Chicago/Turabian StyleSpruill-Harrell, Briana, Anna Pérez-Umphrey, Leonardo Valdivieso-Torres, Xueyuan Cao, Robert D. Owen, and Colleen B. Jonsson. 2021. "Impact of Predator Exclusion and Habitat on Seroprevalence of New World Orthohantavirus Harbored by Two Sympatric Rodents within the Interior Atlantic Forest" Viruses 13, no. 10: 1963. https://doi.org/10.3390/v13101963