Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies
2.3. gB1s Production and Neutralization
2.4. Experimental Design
2.5. Quantitative RT-PCR
2.6. Histology—Light Microscopy
2.7. Immunofluorescence
2.8. Laser Confocal Immunofluorescence
3. Results
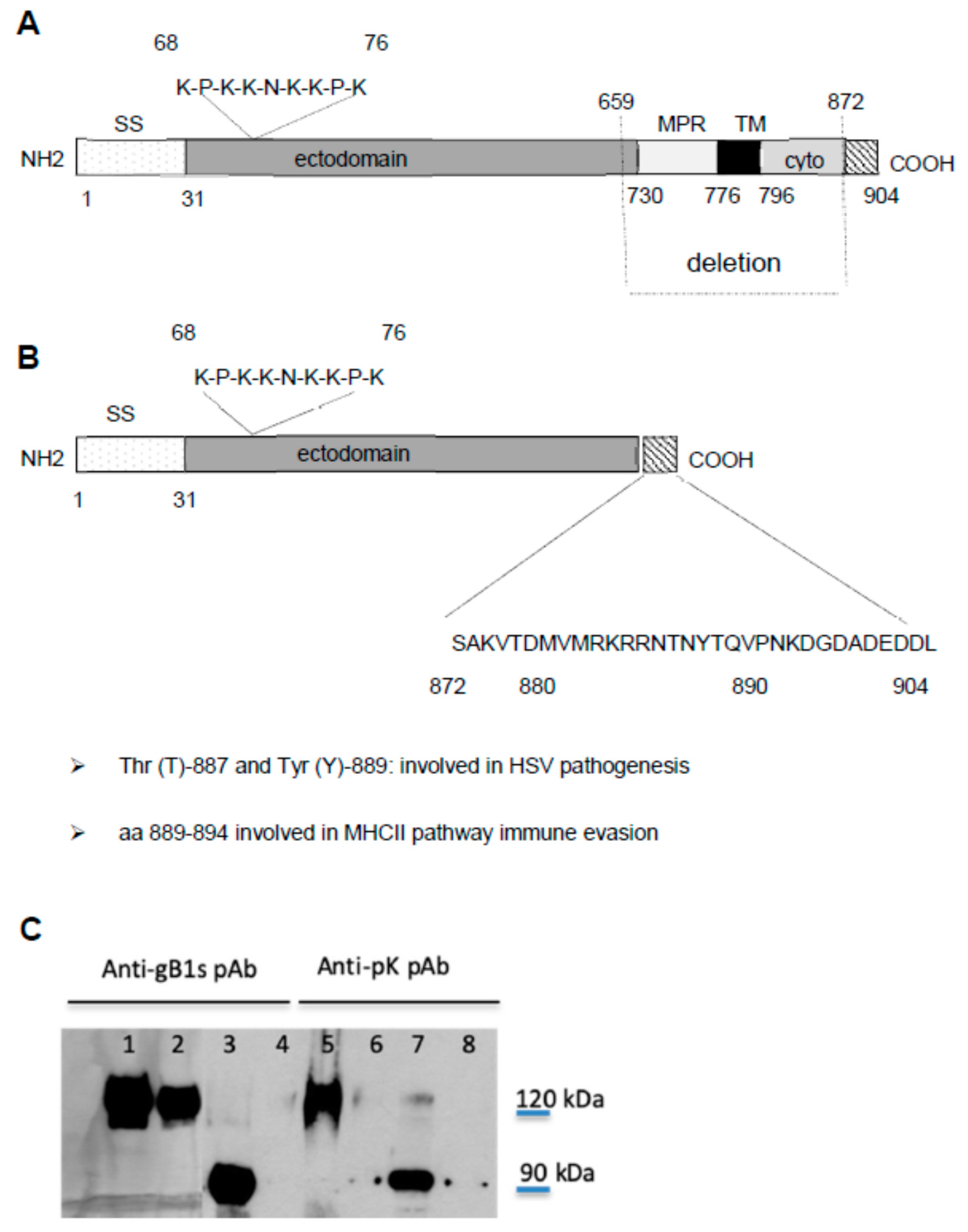
3.1. Characterization of Polyclonal Antibodies against gB1s and Anti gB1-pK Sequences
3.2. Biological Activity of gB1s in an ex-vivo Rabbit Corneal Model
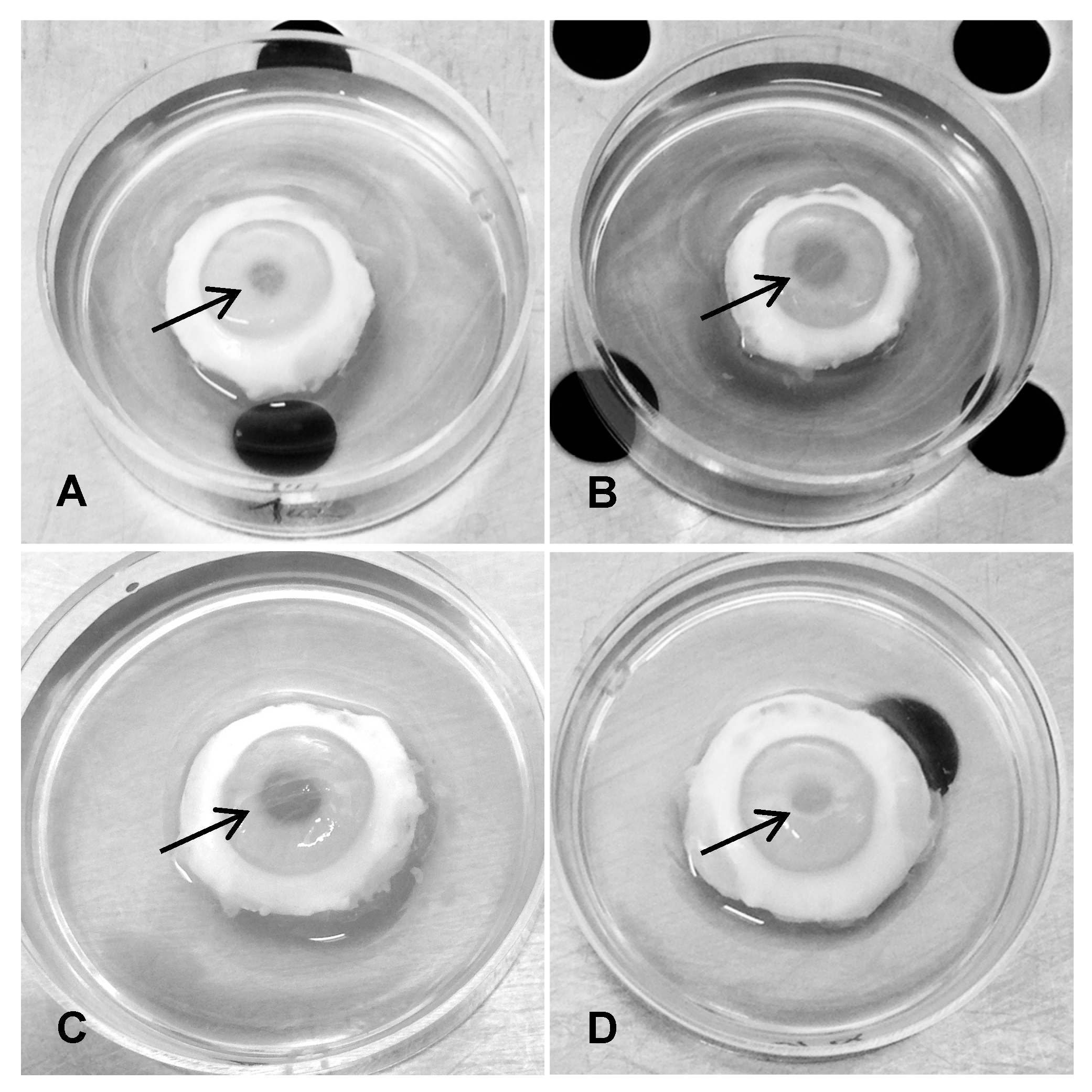
3.2.1. Mydriasis in gB1s Treated Corneas
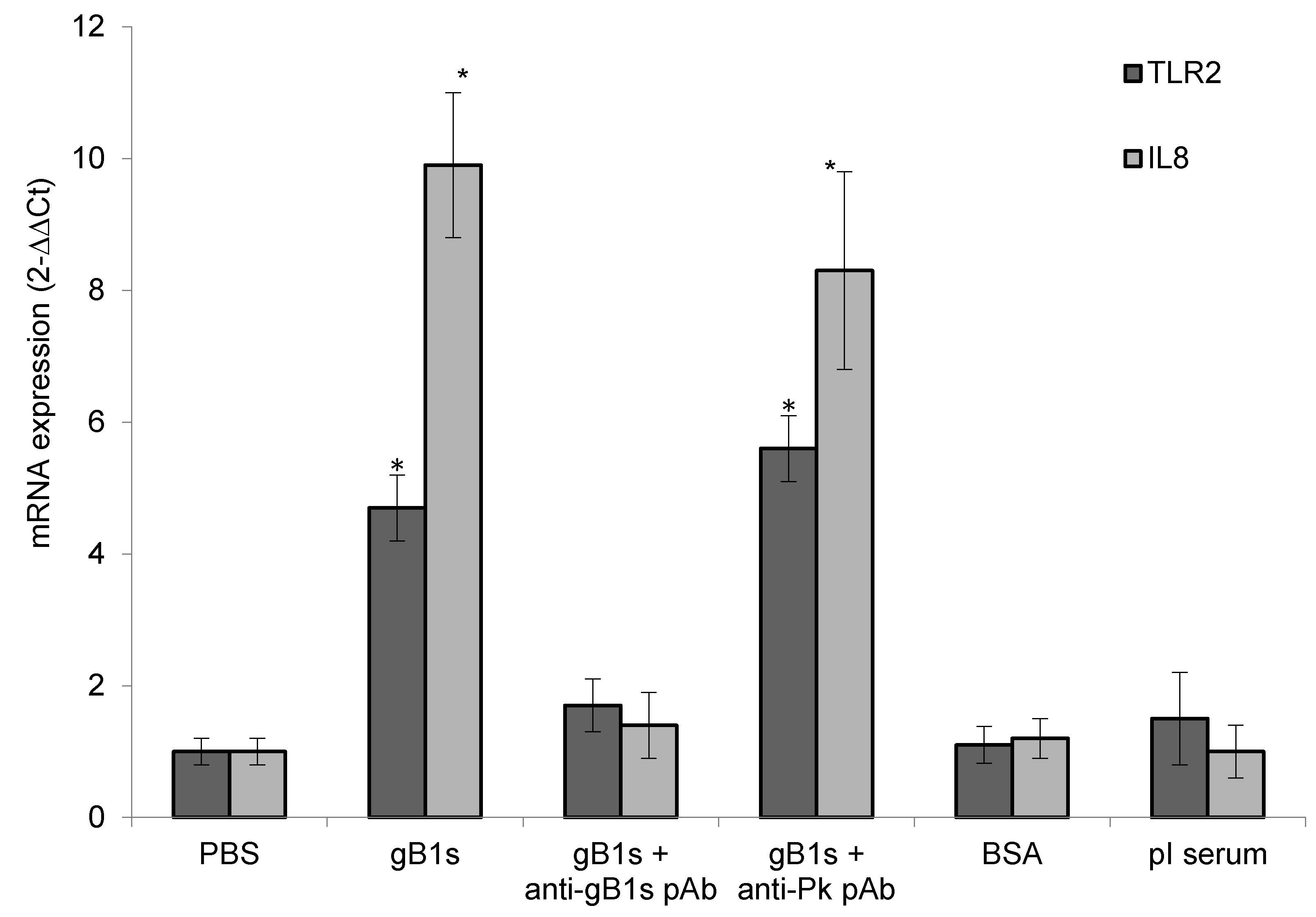
3.2.2. TLR2 and IL8 mRNA Expression in Superficial Layers of Corneal Epithelial Cells Exposed to gB1s
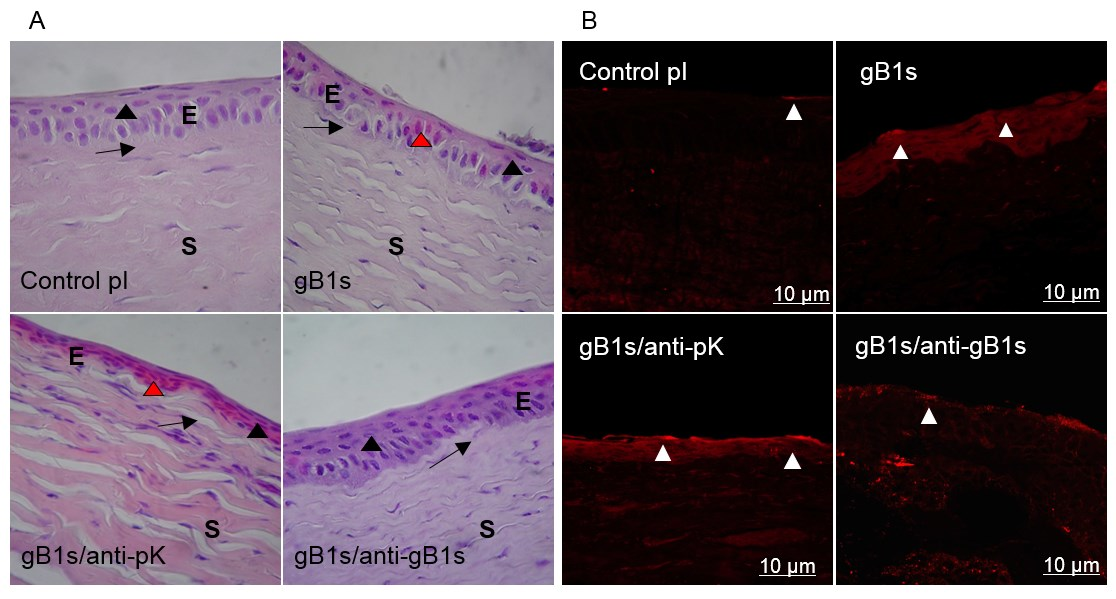
3.2.3. Morphological Changes in Sections of Rabbit Corneal Epithelium and Expression of TLR2 on Cell Surface
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farooq, A.V.; Shukla, D. Herpes simplex epithelial and stromal keratitis: An epidemiologic update. Surv. Ophthalmol. 2012, 57, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global and Regional Estimates of Prevalent and Incident Herpes Simplex Virus Type 1 Infections in 2012. PLoS ONE 2015, 10, e0140765. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.M.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Tsatsos, M.; MacGregor, C.; Athanasiadis, I.; Moschos, M.M.; Hossain, P.; Anderson, D. Herpes simplex virus keratitis: An update of the pathogenesis and current treatment with oral and topical antiviral agents-response. Clin. Exp. Ophthalmol. 2017, 45, 317. [Google Scholar] [CrossRef] [PubMed]
- Hlinomazova, Z.; Loukotova, V.; Horackova, M.; Sery, O. The treatment of HSV1 ocular infections using quantitative real-time PCR results. Acta Ophthalmol. 2012, 90, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, C.S.; Streeten, B.W.; Dawson, C.R. Histopathology of herpes simplex virus keratouveitis. Curr. Eye Res. 1987, 6, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Micera, A.; Sacchetti, M.; Mantelli, F.; Bonini, S. Toll-like receptors in ocular surface diseases: Overview and new findings. Clin. Sci. 2011, 120, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Mai, K.; Chui, J.J.; Di Girolamo, N.; McCluskey, P.J.; Wakefield, D. Role of toll-like receptors in human iris pigment epithelial cells and their response to pathogen-associated molecular patterns. J. Inflamm. 2014, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, B. Recognition of herpes simplex viruses: Toll-like receptors and beyond. J. Mol. Biol. 2014, 426, 1133–1147. [Google Scholar] [CrossRef]
- Sarangi, P.P.; Kim, B.; Kurt-Jones, E.; Rouse, B.T. Innate recognition network driving herpes simplex virus-induced corneal immunopathology: Role of the toll pathway in early inflammatory events in stromal keratitis. J. Virol. 2007, 81, 11128–11138. [Google Scholar] [CrossRef]
- Agelidis, A.M.; Shukla, D. Cell entry mechanisms of HSV: What we have learned in recent years. Future Virol. 2015, 10, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Campadelli-Fiume, G.; Menotti, L.; Avitabile, E.; Gianni, T. Viral and cellular contributions to herpes simplex virus entry into the cell. Curr. Opin. Virol. 2012, 2, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Cheshenko, N.; Trepanier, J.B.; Stefanidou, M.; Buckley, N.; Gonzalez, P.; Jacobs, W.; Herold, B.C. HSV activates Akt to trigger calcium release and promote viral entry: Novel candidate target for treatment and suppression. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 2584–2599. [Google Scholar] [CrossRef] [PubMed]
- Koujah, L.; Suryawanshi, R.K.; Shukla, D. Pathological processes activated by herpes simplex virus-1 (HSV-1) infection in the cornea. Cell Mol. Life Sci. 2019, 76, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Oh, M.J.; Kovacs, M.; Shukla, S.Y.; Valyi-Nagy, T.; Shukla, D. Role for nectin-1 in herpes simplex virus 1 entry and spread in human retinal pigment epithelial cells. Febs J. 2008, 275, 5272–5285. [Google Scholar] [CrossRef] [Green Version]
- Gianni, T.; Massaro, R.; Campadelli-Fiume, G. Dissociation of HSV gL from gH by alphavbeta6- or alphavbeta8-integrin promotes gH activation and virus entry. Proc. Natl. Acad. Sci. USA 2015, 112, E3901–E3910. [Google Scholar] [CrossRef] [PubMed]
- Manservigi, R.; Spear, P.G.; Buchan, A. Cell fusion induced by herpes simplex virus is promoted and suppressed by different viral glycoproteins. Proc. Natl. Acad. Sci. USA 1977, 74, 3913–3917. [Google Scholar] [CrossRef]
- Compton, T.; Kurt-Jones, E.A.; Boehme, K.W.; Belko, J.; Latz, E.; Golenbock, D.T.; Finberg, R.W. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2. J. Virol. 2003, 77, 4588–4596. [Google Scholar] [CrossRef]
- Boehme, K.W.; Guerrero, M.; Compton, T. Human cytomegalovirus envelope glycoproteins B and H are necessary for TLR2 activation in permissive cells. J. Immunol. 2006, 177, 7094–7102. [Google Scholar] [CrossRef]
- Cai, M.; Li, M.; Wang, K.; Wang, S.; Lu, Q.; Yan, J.; Mossman, K.L.; Lin, R.; Zheng, C. The herpes simplex virus 1-encoded envelope glycoprotein B activates NF-kappaB through the Toll-like receptor 2 and MyD88/TRAF6-dependent signaling pathway. PLoS ONE 2013, 8, e54586. [Google Scholar]
- Leoni, V.; Gianni, T.; Salvioli, S.; Campadelli-Fiume, G. Herpes simplex virus glycoproteins gH/gL and gB bind Toll-like receptor 2, and soluble gH/gL is sufficient to activate NF-kappaB. J. Virol. 2012, 86, 6555–6562. [Google Scholar] [CrossRef] [PubMed]
- Manservigi, R.; Grossi, M.P.; Gualandri, R.; Balboni, P.G.; Marchini, A.; Rotola, A.; Rimessi, P.; Di Luca, D.; Cassai, E.; Barbanti-Brodano, G. Protection from herpes simplex virus type 1 lethal and latent infections by secreted recombinant glycoprotein B constitutively expressed in human cells with a BK virus episomal vector. J. Virol. 1990, 64, 431–436. [Google Scholar] [PubMed]
- Marino, A.; Pergolizzi, S.; Lauriano, E.R.; Santoro, G.; Spataro, F.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G. TLR2 activation in corneal stromal cells by Staphylococcus aureus-induced keratitis. Apmis Acta Pathol. Microbiol. Immunol. Scand. 2015, 123, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.A.; Mis, A.A.; Oh, F.S.; Deschenes, J.G. Persistent pupillary dilation in herpes simplex uveitis. Can. J. Ophthalmol. 2009, 44, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Lin, P. Infectious Uveitis. Curr. Ophthalmol. Rep. 2015, 3, 170–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-la-Torre, A.; Valdes-Camacho, J.; Foster, C.S. Bilateral Herpes Simplex Uveitis: Review of the Literature and Own Reports. Ocul. Immunol. Inflamm. 2017, 25, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Laquerre, S.; Argnani, R.; Anderson, D.B.; Zucchini, S.; Manservigi, R.; Glorioso, J.C. Heparan sulfate proteoglycan binding by herpes simplex virus type 1 glycoproteins B and C, which differ in their contributions to virus attachment, penetration, and cell-to-cell spread. J. Virol. 1998, 72, 6119–6130. [Google Scholar]
- Marconi, P.; Manservigi, R. Herpes simplex virus growth, preparation, and assay. Methods Mol. Biol. 2014, 1144, 19–29. [Google Scholar]
- Revello, M.G.; Gualandri, R.; Manservigi, R.; Gerna, G. Development and Evaluation of an Elisa Using Secreted Recombinant Glycoprotein-B for Determination of IgG Antibody to Herpes-Simplex Virus. J. Virol. Methods 1991, 34, 57–70. [Google Scholar] [CrossRef]
- Cortesi, R.; Argnani, R.; Esposito, E.; Dalpiaz, A.; Scatturin, A.; Bortolotti, F.; Lufino, M.; Guerrini, R.; Cavicchioni, G.; Incorvaia, C.; et al. Cationic liposomes as potential carriers for ocular administration of peptides with anti-herpetic activity. Int. J. Pharm. 2006, 317, 90–100. [Google Scholar] [CrossRef]
- Blanco, A.R.; Nostro, A.; D’Angelo, V.; D’Arrigo, M.; Mazzone, M.G.; Marino, A. Efficacy of a Fixed Combination of Tetracycline, Chloramphenicol, and Colistimethate Sodium for Treatment of Candida albicans Keratitis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4292–4298. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Santoro, G.; Spataro, F.; Lauriano, E.R.; Pergolizzi, S.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G.; Dugo, G. Resveratrol role in Staphylococcus aureus-induced corneal inflammation. Pathog. Dis. 2013, 68, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Pergolizzi, S.; Capillo, G.; Kuciel, M.; Alesci, A.; Faggio, C. Immunohistochemical characterization of Toll-like receptor 2 in gut epithelial cells and macrophages of goldfish Carassius auratus fed with a high-cholesterol diet. Fish Shellfish Immunol. 2016, 59, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Silvestri, G.; Kuciel, M.; Zuwala, K.; Zaccone, D.; Palombieri, D.; Alesci, A.; Pergolizzi, S. Immunohistochemical localization of Toll-like receptor 2 in skin Langerhans’ cells of striped dolphin (Stenella coeruleoalba). Tissue Cell 2014, 46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Revello, M.G.; Percivalle, E.; Zannino, M.; Rossi, V.; Gerna, G. Development and Evaluation of a Capture Elisa for Igm Antibody to the Human Cytomegalovirus Major DNA-Binding Protein. J. Virol. Methods 1991, 35, 315–329. [Google Scholar] [CrossRef]
- Caselli, E.; Balboni, P.G.; Incorvaia, C.; Argnani, R.; Parmeggiani, F.; Cassai, E.; Manservigi, R. Local and systemic inoculation of DNA or protein gB1s-based vaccines induce a protective immunity against rabbit ocular HSV-1 infection. Vaccine 2000, 19, 1225–1231. [Google Scholar] [CrossRef]
- Manservigi, R.; Boero, A.; Argnani, R.; Caselli, E.; Zucchini, S.; Miriagou, V.; Mavromara, P.; Cilli, M.; Grossi, M.P.; Balboni, P.G.; et al. Immunotherapeutic activity of a recombinant combined gB-gD-gE vaccine against recurrent HSV-2 infections in a guinea pig model. Vaccine 2005, 23, 865–872. [Google Scholar] [CrossRef]
- Cortesi, R.; Ravani, L.; Rinaldi, F.; Marconi, P.; Drechsler, M.; Manservigi, M.; Argnani, R.; Menegatti, E.; Esposito, E.; Manservigi, R. Intranasal immunization in mice with non-ionic surfactants vesicles containing HSV immunogens: A preliminary study as possible vaccine against genital herpes. Int. J. Pharm. 2013, 440, 229–237. [Google Scholar] [CrossRef]
- Alekseev, O.; Tran, A.H.; Azizkhan-Clifford, J. Ex vivo organotypic corneal model of acute epithelial herpes simplex virus type I infection. J. Vis. Exp. Jove 2012, 69, e3631. [Google Scholar] [CrossRef]
- Cowdry, E.V.; Nicholson, F.M. Inclusion Bodies in Experimental Herpetic Infection of Rabbits. J. Exp. Med. 1923, 38, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V.; Saghizadeh, M. Progress in corneal wound healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Arii, J.; Minowa, A.; Kakimoto, A.; Koyanagi, N.; Kato, A.; Kawaguchi, Y. Role of the herpes simplex virus 1 Us3 kinase phosphorylation site and endocytosis motifs in the intracellular transport and neurovirulence of envelope glycoprotein B. J. Virol. 2011, 85, 5003–5015. [Google Scholar] [CrossRef] [PubMed]
- Niazy, N.; Temme, S.; Bocuk, D.; Giesen, C.; Konig, A.; Temme, N.; Ziegfeld, A.; Gregers, T.F.; Bakke, O.; Lang, T.; et al. Misdirection of endosomal trafficking mediated by herpes simplex virus-encoded glycoprotein B. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, H.; Ghazanfari, T.; Yaraee, R.; Faghihzadeh, S.; Hassan, Z.M. Roles of IL-8 in ocular inflammations: A review. Ocul. Immunol. Inflamm. 2011, 19, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, S.; Huang, Y.Y.; Fong, Y.; Wong, R.J. Calcium depletion enhances nectin-1 expression and herpes oncolytic therapy of squamous cell carcinoma. Cancer Gene Ther. 2007, 14, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Roos, J.C.; Haridas, A.S. Prolonged mydriasis after inadvertent topical administration of the calcium channel antagonist amlodipine: Implications for glaucoma drug development. Cutan. Ocul. Toxicol. 2015, 34, 84–87. [Google Scholar] [CrossRef]
- Nazli, A.; Kafka, J.K.; Ferreira, V.H.; Anipindi, V.; Mueller, K.; Osborne, B.J.; Dizzell, S.; Chauvin, S.; Mian, M.F.; Ouellet, M.; et al. HIV-1 gp120 induces TLR2- and TLR4-mediated innate immune activation in human female genital epithelium. J. Immunol. 2013, 191, 4246–4258. [Google Scholar] [CrossRef]
- Krawczyk, A.; Dirks, M.; Kasper, M.; Buch, A.; Dittmer, U.; Giebel, B.; Wildschutz, L.; Busch, M.; Goergens, A.; Schneweis, K.E.; et al. Prevention of herpes simplex virus induced stromal keratitis by a glycoprotein B-specific monoclonal antibody. PLoS ONE 2015, 10, e0116800. [Google Scholar] [CrossRef]
- Khan, A.A.; Srivastava, R.; Spencer, D.; Garg, S.; Fremgen, D.; Vahed, H.; Lopes, P.P.; Pham, T.T.; Hewett, C.; Kuang, J.; et al. Phenotypic and functional characterization of herpes simplex virus glycoprotein B epitope-specific effector and memory CD8+ T cells from symptomatic and asymptomatic individuals with ocular herpes. J. Virol. 2015, 89, 3776–3792. [Google Scholar] [CrossRef]
- Henrick, B.M.; Yao, X.D.; Taha, A.Y.; German, J.B.; Rosenthal, K.L. Insights into Soluble Toll-Like Receptor 2 as a Downregulator of Virally Induced Inflammation. Front. Immunol. 2016, 7, 291. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, A.; Pergolizzi, S.; Cimino, F.; Lauriano, E.R.; Speciale, A.; D’Angelo, V.; Sicurella, M.; Argnani, R.; Manservigi, R.; Marconi, P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model. Viruses 2019, 11, 819. https://doi.org/10.3390/v11090819
Marino A, Pergolizzi S, Cimino F, Lauriano ER, Speciale A, D’Angelo V, Sicurella M, Argnani R, Manservigi R, Marconi P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model. Viruses. 2019; 11(9):819. https://doi.org/10.3390/v11090819
Chicago/Turabian StyleMarino, Andreana, Simona Pergolizzi, Francesco Cimino, Eugenia Rita Lauriano, Antonio Speciale, Valeria D’Angelo, Mariaconcetta Sicurella, Rafaela Argnani, Roberto Manservigi, and Peggy Marconi. 2019. "Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model" Viruses 11, no. 9: 819. https://doi.org/10.3390/v11090819