
Extracellular Vesicles and Their Impact on the Biology of Protozoan Parasites


1
Division of Infectious Diseases, School of Medicine, Stanford University, Stanford, CA 94305, USA
2
Department of Microbiology and Immunology, School of Medicine, Stanford University, Stanford, CA 94305, USA
*
Author to whom correspondence should be addressed.
Trop. Med. Infect. Dis. 2023, 8(9), 448; https://doi.org/10.3390/tropicalmed8090448
Submission received: 7 August 2023
/
Revised: 11 September 2023
/
Accepted: 11 September 2023
/
Published: 15 September 2023
(This article belongs to the Special Issue Emerging Insights in Pathogenesis of Infectious Protozoa and Algae)
{kind=link}
Abstract
:Extracellular vesicles (EVs) are lipid-membrane-bound structures produced naturally by all cells and have a variety of functions. EVs act as vehicles for transporting important molecular signals from one cell to another. Several parasites have been shown to secrete EVs, and their biological functions have been extensively studied. EVs have been shown to facilitate communication with the host cells (such as modulation of the host’s immune system or promoting attachment and invasion into the host cells) or for communication between parasitic cells (e.g., transferring drug-resistance genes or factors modulating stage conversion). It is clear that EVs play an important role in host–parasite interactions. In this review, we summarized the latest research on the EVs secreted by protozoan parasites and their role in host–parasite and parasite–parasite communications.
1. Introduction
Protozoan pathogens are responsible for many diseases in both humans and animals, accounting for a significant economic burden globally. These unicellular parasites surmount several challenges in establishing a successful infection. In their effort to survive in the host environment, one approach that the parasites utilize is the secretion of extracellular vesicles (EVs) to communicate with each other or to interact with distal host cells. The transport of biologically active molecules through EVs can modulate the behavior of the recipient cells, which can be beneficial for the parasite in adhering to and infecting the host [1]. However, at other times, the host’s immune cells internalize the parasitic EVs, and this primes the immune cells against the infection [2].
EVs are lipid-bilayer-bound vesicles that contain a selectively packaged cargo of proteins, DNA, RNA, and lipids [3,4]. They are heterogenous and vary in size, morphology, and composition depending on the type of cell and the environment. To maintain conformity, the International Society for Extracellular Vesicles (ISEV) defines EV as a collective term for “particles naturally released from the cell that is delimited by a lipid bilayer and cannot replicate” [5,6,7].
Particle tracking analysis has shown heterogeneity in the size of the vesicles released by cells. EVs have been classified according to their biogenesis and size. Exosomes are typically 50–150 nm in size, originate from the endosomal membrane, and are released into the extracellular environment when multivesicular bodies fuse with the plasma membrane [4]. Microvesicles are bigger than exosomes, with diameters ranging from 200 nm to 1 µm; they are released by budding from the plasma membrane [8]. Protozoan parasites have been reported to shed both exosome-like vesicles and larger microvesicle-like vesicles. For example, Trichomonas vaginalis has been reported to release vesicles with a size ranging from 40 nm to 1000 nm [9].
However, as well as the variance in their size, there is also heterogeneity in the EVs’ cargo. As the contents of EVs have been characterized in depth with newer lipidomic, proteomic, and RNA sequencing techniques, it has become clear that EVs’ cargoes differ depending on the parental cell type and the culture conditions [10]. Protozoan parasites have also been reported to control the cargo being packaged in their EVs to modulate the specific signal sent to the recipient cells.
Additionally, intracellular stages of parasites, such as Leishmania or Plasmodium, have the ability to modulate the cargo of the EVs secreted by the infected host cells to promote proliferation and pathogenesis. Interestingly, viruses have been found inside the EVs secreted from some cells [11]. This has been seen in parasites as well. Atayde et al. discovered that Leishmania EVs are used as a viral envelope by the Leishmania RNA Virus 1, facilitating transmission. Similarly, Rada et al. reported that Trichomonas virus particles are released from T. vaginalis cells within EVs [12,13].
The selective packaging of effector molecules into lipid-membrane-bound vesicles, which protect the cargo from external insults, makes EVs the ideal vehicle for intercellular communication [14]. Recent research has identified small RNA such as tRNA fragments and tRNA halves, which are enriched in EVs and could have a potential role in regulating gene expression in the target cells [15,16].
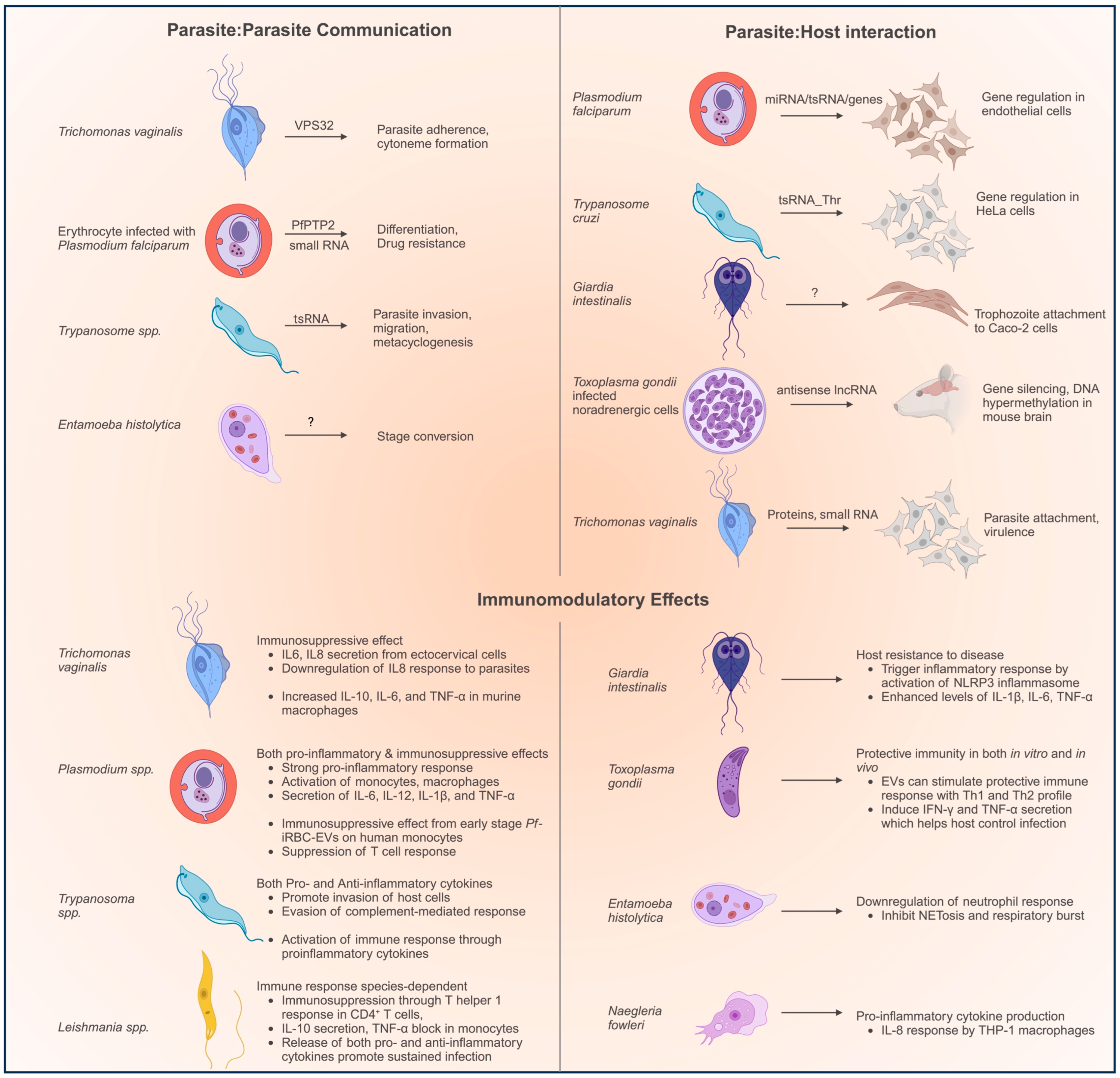
Parasites use EVs to transmit biologically active molecules to enable (a) parasite–parasite communication to help coordinate a concerted action, (b) parasite–host communication to facilitate adherence and virulence, and (c) parasite–host communication targeting the immune cells to suppress the immune response (Figure 1). At the same time, the host’s immune cells can intercept pathogenic EVs by recognizing pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs). PRRs include the family of Toll-like receptors (TLR) and nucleotide-binding oligomerization domain (NOD)-like receptors [17].
Thus, EVs play a central role in the interaction between the parasites and the immune system, which results in a spectrum of outcomes that encompass both pro- and anti-inflammatory effects. This complex interplay ultimately shapes the overall impact on the host’s health and the outcome of the parasitic infection.
In the following sections, we discuss the features of the EVs secreted by different protozoan parasites in terms of their morphology, protein/RNA cargo, and their main biological functions. We summarize the functions of EVs in Figure 1.
2. Extracellular Vesicles Secreted by Protozoan Parasites
2.1. Trichomonas vaginalis
Trichomonas vaginalis is responsible for the most common nonviral sexually transmitted infection worldwide, known as trichomoniasis. This disease leads to complications such as vaginitis, cervicitis, urethritis, and pelvic inflammatory disease, and increases the risk of transmitting HIV. Trichomoniasis has also been associated with the higher incidence and severity of cervical and prostate cancers [18,19,20].
T. vaginalis trophozoites colonize the human urogenital tract, where they adhere extracellularly to the epithelial cells. The EVs secreted by T. vaginalis are some of the best characterized parasitic EVs. Twu et al. first reported the secretion of exosome-like vesicles (Tv-ELVs) from the adherent B7RC2 strain of T. vaginalis [21]. Electron microscopy revealed that Tv-ELVs had a cup-shaped morphology with a size of around 50–150 nm. A different study reported that larger vesicles were also secreted by these parasites. Vesicles with diameters ranging from 100 to 1000 nm were termed “microvesicle-like structures” (MVs) and those larger than 1 µm were termed “large vesicles” (LVs). Electron microscopy showed that the MVs were being shed from the flagellar membrane of the parasite. Both MVs and LVs were found to be secreted in response to stimulation by the host cells and were selectively enriched for individual characterization of their proteomes.
The Tv-ELVs’ proteome was characterized, and around 73% of the Tv-ELVs’ proteins were found to be orthologs of mammalian exosome proteins [21]. A member of the ESCRT III complex, VPS32, was shown to be important for biogenesis and the sorting of the protein cargo into the EVs (the protocol used did not separate the vesicles on the basis of their size and therefore, the generic term EV was used). Overexpression of VPS32 in the T. vaginalis trophozoites increased the secretion of EVs and improved the binding and uptake by the host cells [21].
Characterization of the RNA cargo showed small RNAs ranging in size from between 25 and 200 nt in the Tv-ELVs [21]. A later study showed that small RNAs made up ~55% of the total RNA, and tRNA fragments constituted the majority of these small RNA. Interestingly, the most abundant tRNA fragments were 5′ tRNA halves. T. vaginalis does not have miRNA genes, and tRNA halves could be important regulators of gene expression in these parasites [22]. Their presence in the Tv-ELVs suggests a possible role in parasite–parasite and parasite–host communications through gene regulation. However, the enrichment of tRNA halves due to nutrient deprivation such as serum starvation during the preparation of EVs cannot be ruled out. Further work is required to elucidate the role of the tRNA fragments packaged in the Tv-ELVs.
Trichomonas parasites can harbor an double-stranded RNA virus endosymbiont, Trichomonasvirus, TVV. Govender et al. showed that TVV-positive parasites secreted EVs that carried a modified protein cargo that helped the parasite subvert the host’s immunity. [23]. However, recently, Rada et al. reported that the viral particles from TVV loaded inside T. vaginalis EVs resulted in a markedly higher proinflammatory immune response [13]. Future work should elucidate the role of the viral particles in promoting inflammation-related pathogenesis during trichomoniasis.
Furthermore, the selective packaging of certain proteins involved in pathogenesis, such as some proteases, as well as surface proteins involved in parasite attachment was also observed. Rai et al. showed that 4-α-glucanotransferase (Tv4AGT) on the surface of Tv-ELVs binds to the heparan sulfate (HS) present on the host cell’s surface proteoglycans. This interaction was shown to be critical for the internalization of EVs through a lipid raft-mediated endocytosis process [24].
After adhering, Tv-ELVs fuse with the host cells to deliver their cargo into the cells. Once internalized by the recipient cells, these vesicles can modulate their behavior in various ways. Tv-ELVs can modulate both parasite–parasite and parasite–host cell adherence. Data have shown that pathogenic factors specific to parasitic strains were specifically packed into Tv-ELVs, and thus highly adherent strains of T. vaginalis secreted EVs that helped increase the attachment of a less adherent parasite strain to the host cells [21].
Internalization of the Tv-ELVs by ectocervical cells elicited the secretion of IL-6 and, to a lesser extent, IL-8 from the ectocervical cells. Preincubation of the cells with Tv-ELVs prior to the addition of T. vaginalis parasites led to a significant inhibition of the secretion of IL-8 but not IL-6 by the cells, indicating that Tv-ELVs specifically modulate the production of IL-8 by the ectocervical cells [21].
An immunomodulatory role of Tv-ELVs was found in an in vivo murine model of trichomoniasis [25]. T. vaginalis ELVs dampen the inflammatory response both in vitro and in vivo. Anti-inflammatory IL-10 was dramatically increased, whereas pro-inflammatory TNF-α and IL-6 were moderately induced in RAW264.7 macrophages stimulated with Tv-ELVs. Mice pretreated with the vesicles had an elevated production of IL-10 but reduced levels of IL-6, IL-13, and IL-17 [1]. The reduction in the inflammatory response by the Tv-ELVs likely favors the persistence of T. vaginalis parasites.
Thus, the EVs secreted by T. vaginalis parasites can promote the pathogenesis of the disease by modulating the host immune response mediated by the secretion of cytokines and by facilitating the parasites’ adherence and persistence in the host.
2.2. Entamoeba histolytica
Entamoeba histolytica is an anaerobic parasite that infects the colon. It has a biphasic lifecycle consisting of the dormant cyst stage, which survives in the environment, and a motile trophozoite stage, which causes invasive disease [26]. While most infections are localized to the colon (colitis), extra-intestinal amoebiasis can sometimes occur through invasion of the colonic mucosa by trophozoites, which can spread to the liver and cause amebic liver abscesses. Entamoeba is one of the primary causes of parasitic mortality in humans and represents a significant health concern in the developing world.
E. histolytica EVs consist of the characteristic cup-shaped vesicles of around 125 nm in diameter [27]. A second study reported more variation in the size of the amoebic EVs: the amoebic EVs had an average size of 167 nm, but EVs (microvesicles) of up to 600 nm were also observed [28]. Both studies used the same EV isolation protocol (a commercial PEG-based kit) and the variation in size could be due to different culture conditions or variance in the amoeba cultures maintained in the different labs.
Characterization of the EVs’ proteomes showed similar results in both studies, with selective enrichment of proteins associated with vesicle formation, cytoskeletal proteins, and common EV marker proteins such as elongation factor 1-alpha, heat shock protein 70, and ADP-ribosylation factor [27,28]. However, tetraspanins, which are common EV marker proteins, were not detected in the E. histolytica EVs’ proteome in either of these reports. The E. histolytica genome has 17 tetraspanins, some of which are highly expressed in the parasites, but none of them was found on the EVs, suggesting a distinct mechanism of biogenesis of these EVs that is not dependent on tetraspanins.
RNAseq of the EV RNA showed that a 27 nt antisense RNA population was enriched in the EVs [27]. E. histolytica parasites contain a 27 nt antisense small RNA (AS sRNA) population that can regulate the expression levels of specific target genes to very low levels [29,30]. Interestingly, the EVs were also found to contain some proteins that have been identified as members of the RNA-induced silencing complex (RISC) in Entamoeba, such as Argonaute 2-2 and a Tudor-domain-containing protein. This could suggest that E. histolytica EVs might serve to transport these antisense small RNA effector molecules to other cells.
Recently, the presence of tRNA halves in the E. histolytica EVs was also reported. tRNA halves were accumulated in E. histolytica parasites during stress [16]. This accumulation of tRNA halves was seen to be concomitant with a decrease in protein translation during stress. The packaging of the tRNA halves in the EVs suggests a role for the tRNA halves in modulating gene expression in distal cells. However, more work is required to confirm this hypothesis. As mentioned before, the packaging of tRNA halves in EVs from unstressed cells needs to be confirmed. The usual EV isolation protocols require serum starvation, which would lead to the accumulation of tRNA halves in the parasites and might cause their subsequent packaging in the EVs.
Amoebic EVs were seen to be instrumental in inter-parasite communication. EVs isolated from encysting parasites promoted encystation in other parasites, whereas EVs from metabolically active trophozoites impeded encystation (these experiments were conducted on Entamoeba invadens as an encystation model) [27,31]. Further investigations to identify the cargo content of EVs from encysting cells could help to elucidate the regulatory factor(s) and their roles in parasite–parasite communication.
Díaz-Godínez et al. characterized the immunomodulatory role of the EVs of E. histolytica on neutrophils [28]. Amoebic EVs were found to be incorporated into neutrophils and transfer their cargo into the neutrophils following membrane fusion. This resulted in a drastic reduction in the oxidative burst and NETosis from the neutrophils stimulated by PMA, ionophore A23187, or the amoeba itself. Although the amoebic EVs contained ROS and were able to transfer them to the neutrophils, they still had a suppressive effect on the NETosis induced by other stimuli (chemical or biological). The mechanism of the inhibition of NETosis by amoebic EVs are still not understood and further work is required.
Thus, the data suggest that Entamoeba EVs are capable of impacting the parasite’s biology (conversion to cysts) and modulating the host (suppressing the immune response and promoting infection). However, further work needs to be carried out to elucidate the role of these EVs in these biological processes, and the specific biomolecules they carry.
2.3. Giardia
The protozoa Giardia duodenalis is the causative agent for the zoonotic enteric infection called giardiasis. The parasite is transmitted by the fecal–oral route through ingestion of the cysts from contaminated water, leading to colonization of the intestine by the trophozoites. The trophozoites attach tightly to the outside of the epithelial cells of the small intestine though their adhesive disks [32]. Giardia has undergone reductive evolution and lacks an endosomal/lysosomal system, a Golgi complex, peroxisomes, and mitochondria [33]. Instead, a network of peripheral vesicles (PVs) performs several roles, such as those of multivesicular bodies, including the release of extracellular vesicles.
Evans-Osses et al. reported that EVs were secreted by Giardia in response to different environmental conditions such as pH levels and the concentration of calcium [34]. The vesicles induced in the presence of calcium, a known inducer of MVs, were defined as microvesicles (MVs), since their size was around 200 nm. Consistent with what was seen in other systems, the origin of the Giardia MVs was found to be the plasma membrane and required a lipid raft for synthesis [35]. In contrast, Zhao et al. reported that G. duodenalis trophozoites secrete cup-shaped vesicles with a membrane bilayer and a diameter of around 100–200 nm and called them EVs [36]. A different group reported two distinct types of EVs secreted by Giardia: small vesicles (SVs) that were smaller than 100 nm, and large vesicles (LVs) that were 100–400 nm in size [37]. The SVs were exosome-like particles, and their biogenesis and protein content were dependent on lipid rafts. Inhibitors of lipid rafts affected the morphology and composition of SVs (and, to a smaller extent, LVs) and disrupted the attachment of the trophozoites to the host cells [37].
Analysis of the MVs’ cargo showed the presence of proteins seen in MVs from other systems [34]. Proteins involved in pathogenesis such as VSPs and giardins were found in the MVs, along with a high number of uncharacterized proteins that were found exclusively in the MVs of the cyst stage. The MVs’ cargo also contained small RNA, though this was not sequenced. The MVs were reported to aid in the trophozoites’ attachment to Caco-2 cells, although MVs alone did not affect cell viability (though Giardia can induce apoptosis of the host cell) [34].
The MVs have a moderate immunomodulatory effect on their internalization by human immature dendritic cells (iDCs). Subsequent studies showed a stronger response of the host cells’ innate immune system after exposure to Giardia EVs. Endocytosis of G. duodenalis EVs (GEVs) by murine peritoneal macrophages induced inflammatory cytokines such as IL-6, IL-10, IL-12, IL-17, IL-18, IFN-γ, TNF-α, IL-1β, Cxcl2, and Ccl20, which may contribute to the host’s resistance to the development of the disease. Several Toll-like receptors (TLRs) and Nod-like receptors (NLRs) were seen to be upregulated, and TLR2 and NLRP3 inflammasome signaling pathways were activated after treatment with GEVs. Pretreatment of murine macrophages with the GEVs enhanced the inflammatory response to G. intestinalis trophozoites [2].
A more recent study characterized the RNA content of the vesicles secreted by Giardia duodenalis [38]. The vesicles were found to be similar in size, shape, and protein and lipid composition to those described for exosomes from other eukaryotic cells and were therefore called “exosomal-like vesicles (ElVs)”. RNAseq of the ElVs secreted from different assemblages of Giardia showed distinct small RNA populations, including ribosomal small RNAs (rsRNAs), messenger small RNAs (msRNAs), and transfer small RNAs (tsRNAs). No miRNA could be identified in the ElVs. This was expected, since Giardia lacks the usual miRNA machinery, such as DROSHA and XPO5 [38].
The presence of tRNA fragments in EVs from Giardia adds to the growing body of evidence that tRNA fragments are ubiquitously packaged in EVs and suggests a possible role in the modulation of gene expression in the target cells. However, further work would be required to confirm the biological role of tRNA fragments in these EVs.
Thus, the EVs secreted by Giardia can promote the pathogenesis of the disease by aiding the trophozoites’ attachment and internalization. However, exposure of the immune cells to the EVs elicited a pro-inflammatory cytokine response that contributed to resistance against the parasites.
2.4. Leishmania
Leishmania spp. is the causative agent for a variety of diseases called Leishmaniasis, which is spread by the bite of infected phlebotomine sandflies. Leishmaniasis vary from cutaneous infections to the potentially lethal visceral infections that have a high mortality rate and economic burden globally [39]. Leishmania parasites exist as two structural variants in two distinct hosts, depending on the stage of their lifecycle: the Leishmania amastigote is the intracellular stage that lives inside macrophages in mammals, while the promastigote is the motile extracellular form in the digestive tract of the vector, sandflies.
Several reports have characterized the EVs secreted by different Leishmania parasites. Both exosome-like [40,41] and microvesicle-like vesicles were observed [42,43]. The EV proteome was analyzed and found to contain immunogenic proteins such as active proteinases, and virulence factors such as HSP100, LPG2, and glycoprotein 63 kDa (gp63) [40,44,45,46,47]. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was found to be highly enriched in EVs of L. major and contributed to the inhibition of TNF-α [43]. The protein kinase casein kinase 1.2 (CK1.2) was released by the promastigotes’ exosomes in several Leishmania species and directly phosphorylated host’s extracellular substrates [48,49,50].
The RNA content of Leishmania EVs showed selectively and specifically enriched small RNAs derived exclusively from noncoding RNAs such as rRNAs and tRNAs. Interestingly, tRNA fragments, predominantly tRNA halves (tsRNAs) were found to be amongst the highly enriched RNA populations. The majority of tsRNAs in the EVs of both L. donovani and L. braziliensis were 5′ tRNA halves. Northern blot experiments demonstrated that the tRNA-Asp and tRNA-Leu were abundant in the EVs, though no detectable amounts were observed in the parental parasite’s RNA [51]. Interestingly, EVs from another parasite, E. histolytica, were also reported to contain tRNA-Asp [16]. A recent study has reported that Leishmania-derived small RNAs, including tsRNA, can be found in the blood of mice infected with L. donovani and L. amazonensis [52]. The enrichment and dissemination of small RNA through EVs in the host merits the investigation of the regulation of the host’s genes by these parasites.
EVs play an important role in the Leishmania infection cycle in both the promastigote and amastigote stages [48,53,54]. Promastigotes can secrete EVs into the lumen of the sandfly midgut, from where they are egested along with the parasites when the insect bites the mammalian host. Co-inoculation of the EVs with the parasites has been shown to facilitate infection [55]. Moreover, macrophages infected with amastigotes secrete EVs that can modulate innate and adaptive immune responses in the mammalian host [41,54].
Martin Olivier’s group published seminal work on the secretion of EVs from Leishmania promastigotes in the sandfly midgut, elucidating their role as vehicles for biologically active molecules. Midguts from sandflies infected with L. infantum and L. major were analyzed by TEM, and vesicles were imaged on the promastigotes’ membrane surfaces [55]. EVs were egested along with the parasites when feeding on blood and were found to exacerbate infection through the induction of inflammatory cytokines such as IL-17a and IL-10. The EV proteome was found to be highly conserved among strains, except for a few specific proteins that were enriched differently in the parental parasites in response to drugs. The EVs contained several virulence factors, transcription factors, and proteins encoded by drug resistance genes [56]. The EVs secreted by drug resistant Leishmania parasites were highly enriched in genomic regions containing drug resistance genes, and these EVs were capable of transmitting the drug resistance genes to distal parasites and promoted their survival [57]. In another interesting discovery, they showed that Leishmania RNA virus 1 (LRV1) can hijack Leishmania’s exosome as a vehicle to reach the extracellular environment. Leishmania EVs enabled viral transmission and infectivity by acting as an envelope for LRV1, which protected the virus from RNAses such as RNAseIII [12].
The effect of Leishmania EVs on the host’s immune response has been studied in detail and was found to vary with species [25]. L. donovani EVs were reported to have an immunosuppressive effect, inducing a T helper 1 response in CD4+ T cells, promoting the production of IL-10, and inhibiting the production of TNF-α in monocytes [49,58]. EVs from L. amazonensis promoted the progression of the disease by increasing the expression of IL-6 and IL-10 by naïve macrophages [54]. Recent work has shown that the EVs secreted by Leishmania-infected macrophages (LiEVs) contain parasite-derived molecules and can carry these to the liver in murine models of infection [59,60]. Although there was some initial evidence that LiEVs are associated with the M2 polarization of macrophages, further work is required to verify that the EVs are responsible for the alternatively activation of M2 polarization.
Weber et al. analyzed the EVs isolated from axenic cultures of the promastigotes of the cutaneous parasites L. shawi and L. guyanensis [41]. Zeta potential measurements indicated that the EVs were likely to flocculate and integrate with the cellular membranes in a nondisruptive way. Incorporation of the EVs into macrophages facilitated the recognition of parasitic antigens by surface and intracellular pattern recognition receptors (PRR) such TLR4, NOD1, and TLR9. A mix of both pro- and anti-inflammatory cytokines was observed, suggesting that the EVs can promote a balanced and sustained low level of infection.
Thus, Leishmania EVs promote the parasites’ survival and infectivity during Leishmaniasis. However, the impact of these EVs on disease outcomes varies among Leishmania species, and both immunosuppressive and immunostimulatory effects have been observed.
2.5. Plasmodium
The apicomplexan protozoan Plasmodium spp. is the causative agent of malaria, one of the most important infectious diseases in the world. Infection with P. falciparum is potentially life-threatening, while P. vivax is commonly responsible for less severe forms of malaria. The parasite’s sporozoites are transmitted to humans by the bite of an infected female Anopheles mosquito when it feeds on blood. The sporozoites migrate to the liver, infect the hepatocytes, and multiply to release merozoites. During the blood stage, merozoites replicate inside the erythrocytes. Mature gametocytes are released into the bloodstream and can be taken up by the mosquito. Sexual development of the parasites occurs in the mosquito midgut, resulting in ookinetes which penetrate the epithelium and form oocysts. These replicate to form the sporozoites that migrate to the salivary glands to complete the lifecycle [61].
During infection, EVs can be released into the body fluids directly by the extracellular sporozoites, and by the erythrocytes and hepatocytes infected by the parasite. At the same time, parasitic infection can modulate the secretion of EVs from activated effector cells [62]. However, research on the EVs related to malaria has predominantly focused on the intraerythrocytic stages of the parasite. High levels of plasma EVs have been reported in patients and are known to contribute to malaria-associated clinical symptoms, such as cerebral malaria [63].
EVs from Plasmodium-infected RBCs (iRBCs) have been very well characterized and were found to contain a variety of molecules mediating pathogenesis and intercellular communication between the host cells, and between the host and the parasite [64]. Interestingly, Pf-iRBC-EVs from RBCs infected with different stages of malaria parasites had distinct protein expression profiles, which indicates the tight control of the EV cargo by the intracellular parasites [65]. Different proteomic analyses have confirmed that virulence-associated proteins such as PfPTP2 and MSP-1 are enriched in these EVs, and they were shown to have a role in EV-mediated communication [66,67,68]. In a recent report, Abou Karam et al. identified two EV subpopulations derived from Pf-iRBC with distinct size ranges of 30–70 nm and 70–300 nm. The proteomes of these two populations were distinct, which indicated that these EVs could induce different phenotypic effects in separate recipient cells [69].
The functional role of the nucleic acid content of Pf-iRBC EVs has also been characterized well. It was reported that the EVs mediate cell–cell communication between the infected erythrocytes by delivering genes, and this was found to promote their differentiation to gametocytes [67]. The DNA content of the EVs also has a role in altering the host’s gene expression: Pf-iRBC EVs carry P. falciparum gDNA, which, when internalized by monocytes, is recognized by the host’s PRRs, which triggers a signaling cascade, resulting in modulation of the induction of the host’s genes [70,71].
Pf-iRBC EVs can also carry small RNA molecules to distal cells. In an elegant study, Mantel et al. showed that the EVs contain functional miRNA derived from the host’s RBCs. The miRNA–Argonaute 2 complexes in the EVs could modulate the expression of the target gene and the properties of the barrier in target endothelial cells [72]. Small regulatory RNA populations from these EVs consisted of mRNAs coding for exported proteins and proteins involved in drug resistance, as well as noncoding RNAs, such as rRNAs, small nuclear (snRNAs), and tRNAs. tRNA-derived fragments from both the host and the parasite were identified in the EVs, with tRNA halves being the predominant species [73,74].
The role of Pf-iRBC EVs in helping the parasite evade the host’s immune mechanism by regulating the cytokine levels from various immune cells has been reviewed extensively before [62,64,75]. The pro-inflammatory response (characterized by the activation of monocytes and macrophages, and the secretion of pro-inflammatory cytokines such as IL-6, IL-12, and IL-1β) has been most commonly reported. Recently, these EVs have also been reported to prime naïve RBCs to enable invasion through a 20S proteasome secretion mechanism [76]. Pf-iRBC EVs have also been shown to suppress the immune response under certain situations, indicating the complex and dynamic role of these EVs [77].
Pf-iRBC EVs have also been proposed to have a role in quorum sensing (QS) by the parasites by regulating their growth density. Plasmodium EVs could induce apoptosis by transporting PfLDH to the recipient cells if the parasite population was high [78].
Several studies have been carried out to investigate Plasmodium EVs, and there is mounting evidence for their role in driving the progression of the disease. The functions of different biomolecules transported inside the EVs have been elucidated, and they were found to contribute to a complex interplay between the parasite and the host’s immune system.
2.6. Toxoplasma
The obligate intracellular parasite Toxoplasma gondii infects nucleated cells in humans and animals, leading to toxoplasmosis, an important zoonotic disease which causes a significant economic burden worldwide.
EVs have been reported from extracellular tachyzoites of T. gondii. Both exosome-like and microvesicle-like EVs were identified. A fraction of these vesicles was morphologically similar to mammalian exosomes, with a size of around 138–170 nm. These EVs were found to contain miRNA and were found to activate murine macrophages to induce TNFα, iNOS, and IL-10 [79,80]. In a different study, EVs from tachyzoites were found to induce several changes in the host’s cells that helped enhance the survival, transmission, and progression of T. gondii parasites [81]. Interestingly, immunization with EVs from T. gondii tachyzoites could confer protection in murine infections, activating cellular and humoral responses, and could elicit a combined Th1 and Th2 profile. The immunization reduced parasitemia and increased the survival index [82].
EVs from cells infected with T. gondii have also been well characterized [83]. The parasite can package its proteins into the host’s EVs, which helps protect the parasite from the host’s immune system. Proteomic analysis of the EVs from human foreskin fibroblasts (HFF) infected with T. gondii identified classical EV marker proteins such as extracellular matrix glycoproteins, calcium-binding proteins, filament-associated proteins, members of the annexin family, HSP 70, and CD63 [84]. RNA characterization of these EVs showed the enrichment of mRNA and miRNA. Four specific mRNA species with neurologic activity, namely Rab-13, eukaryotic translation elongation factor 1 alpha 1, thymosin beta 4, and an LLP homolog, were the most predominant [85].
T. gondii EVs were seen to impart protective immunity in both in vitro and in vivo studies. EVs from infected dendritic cells of the host had antigen and adjuvant properties, and could confer protection against T. gondii in mouse models [86,87]. Moreover, immunization of BALB/c mice with T. gondii exosomes improved the survival time, with both humoral and cellular responses [88].
Recently, Tedord et al. described a novel type of the regulation of neurotransmission mediated by EVs from T. gondii that may contribute to maintaining chronic infection. T. gondii EVs from infected noradrenergic cells were shown to contain a natural mammalian antisense lncRNA that induced gene silencing and DNA hypermethylation in vitro and in the brain [89].
2.7. Trypanosoma cruzi
The obligate intracellular parasite Trypanosoma cruzi is the causative agent for Chagas disease (CD) or American trypanosomiasis.
EVs from T. cruzi epimastigotes were first described by da Silveria et al. in 1979 [90]. Later, these EVs were characterized and found to consist of two populations with distinct sizes. Proteomic analysis showed proteins involved in metabolism, host–parasite interactions, signaling, nucleic acid binding, parasite survival, and virulence factors such as trans-sialidases, mucin, mucin-associated surface protein, cruzipain, and phosphatases [91,92,93]. Distinct proteome profiles were observed in EVs from different strains of the parasites and corresponded to different infection profiles in these strains [94].
Characterization of the RNA content of the EVs showed the enrichment of tRNA halves as well as the argonaute TcPIWI-tryp protein, suggesting an endocytic/exocytic pathway in T. cruzi [95,96]. The EVs deliver tRNA halves to the recipient parasites as well as the mammalian host’s cells. The EVs secreted by epimastigotes were seen to strongly promote metacyclogenesis (transformation of the epimastigote to the trypomastigote form) in axenic cultures [97]. Garcia et al. also found that T. cruzi EVs elicit changes in the gene expression of recipient HeLa cells through a microarray analysis. Genes related to the host’s cell cytoskeleton, extracellular matrix, and immune responses pathways were found to be differentially expressed upon EV treatment. Interestingly, the parasitic tRNA halves packaged in the EVs were identified as effector molecules regulating gene expression in the HeLa cells. Overexpression of tstRNA_Thr in HeLa cells resulted in upregulation of the target CXCL2 gene [96]. Recently, Cornet-Gomez et al. carried out a transcriptomic analysis of Vero cells stimulated by EVs derived from the trypomastigote stage of T. cruzi and found the overexpression of genes related to ubiquitin-related processes and downregulation of Rho-GTPase. The authors also reported the anti-apoptotic role of these EVs [98].
EVs released by several T. cruzi strains were shown to trigger an proinflammatory response mediated by TNF-α, IL-12, IL-6, and NO which enhance the invasion of the host cells via Toll-like receptor 2 [99], although the secretion of anti-inflammatory IL-10 from T and B cells has also been reported [25].
2.8. Trypanosoma brucei
Trypanosoma brucei is the causative agent of human African trypanosomiasis (HAT), also known as sleeping sickness.
T. brucei EVs were first described by Szempruch et al. The EVs were found to be 70–80 nm in size and were formed by budding of the flagellar membrane. Proteomic analysis of the EVs identified several flagellar proteins that contribute to virulence. Trypanosome EVs were found to fuse with mammalian erythrocytes, resulting in rapid erythrocyte clearance and anemia [100].
Eliaz et al. showed that T. brucei EVs had a role in parasite–parasite communication and could affect a change in the migration of the recipient parasites. EVs from stressed trypanosomes were incorporated in the recipient parasites and affected their social motility by causing repulsion. Thus, the parasites were proposed to signal to other parasites about immediate stressful environments [101].
2.9. Naegleria
Naegleria fowleri, known popularly as the “brain-eating amoeba”, is a thermophilic free-living amoeba that is the causative agent for the fulminant disease known as primary amoebic meningoencephalitis (PAM). N. fowleri does not require a host in nature or a vector for transmission; however, it can thrive in the environment, particularly in warm rivers and lakes [102].
The EVs secreted by N. fowleri have a mean size of around 206 nm. Preliminary proteomic analysis revealed a predominance of serine proteases in the EVs. [103]. A different study characterized N. fowleri-derived microparticles (Nf-MP) and N. fowleri-derived exosomes (Nf-Exo), although the sizes of these two were not too different, with an average diameter of 150 nm and 140 nm, respectively [104]. The EVs elicited an immune response that was distinct from that known to be induced by the N. fowleri trophozoites. Macrophages and microglia cells play an instrumental role in the immune response to N. fowleri, though the infection leads to necrosis and apoptosis in these cells [105]. However, no apoptosis was observed in THP-1 monocytes and macrophages exposed to the Nf EVs.
Stimulation with Nf EVs was seen to activate Thp-1 macrophages and induce the production of IL-8. This was proposed to trigger the migration of neutrophils, leading to their accumulation in the brain and their involvement in the pathogenesis of PAM [104].
Further work is required to elucidate the role of Naegleria EVs in the progression and pathogenesis of the disease.
Extracellular vesicles play several important biological functions during the pathogenesis of protozoan diseases. With their cargo of biologically active molecules protected by a lipid bilayer, EVs are the ideal vehicle to communicate with distal cells. Various studies have identified different effector molecules that mediate the signals, including proteins, nucleic acids, and even reactive oxygen species (ROS). Proteins, such as the GP63 protease found in Leishmania EVs and the PfPTP2 packaged in Plasmodium EVs, play a pivotal role in transmitting signals to the recipient cells [45,47,67]. Small RNA species, including miRNA, antisense RNA, and tRNA halves (tsRNAs), can regulate gene expression in the target cells, as well as initiate the host’s immune response. EVs have also been shown to communicate with the target cells through their DNA cargo. Plasmodium EVs can carry drug resistance genes to the recipient parasites and can also regulate gene expression in the host’s cells [73]. Through these manipulations, EVs can also facilitate adherence to the host and promote the progression of the disease.
It is not surprising, therefore, that EVs are emerging as important mediators of the immune response for various parasites. EVs carry immunogenic molecules that can be identified by pattern recognition receptors (PRRs) and activate the host’s innate immune responses. In contrast, EVs can also carry molecules that have an immunosuppressive effect (e.g., the agonists of PRRs) to have an overall immune-protective effect. Moreover, the specific packaging of these molecules allows for a finely nuanced modulation of the host’s immune response, leading to the secretion of anti-inflammatory or pro-inflammatory cytokines by the same parasite, depending on its situation.
On the basis of these properties, EVs’ functions can be broadly classified into three main categories.
- Parasite–parasite communication. EVs can coordinate the interactions between parasitic cells. E. histolytica EVs from encysting parasites were shown to carry encystation signals to trophozoites and promote stage conversion [27]. Similarly, T. vaginalis parasites can transfer adherence-promoting factors to other parasites through EVs [21]. RBCs infected with P. falciparum secrete EVs that can carry signals for the differentiation of gametocytes or drug resistance to other parasites [67]. T. brucei parasites can transmit factors that promote invasion and migration [101].
- Parasite–host interactions: EVs can also benefit parasites by regulating the host’s cells to promote infection. P. falciparum-infected RBCs secrete EVs which can carry small RNA such as miRNA and tsRNA to the endothelial cells [72]. Similarly, tsRNA in EVs from T. cruzi parasites have been shown to regulate mRNA levels in HeLa cells [96]. T. gondii EVs carry antisense RNA to downregulate gene expression in murine brain models [89]. Giardia EVs promote attachment to the host’s cells [34].
- The role of EVs in immunomodulation has been extensively studied, and both pro-inflammatory and anti-inflammatory functions have been observed. For example, T. vaginalis EVs stimulate the host’s macrophages to secrete pro-inflammatory cytokines such as IL-6 and IL-8, which help promote the infection [21]. P. falciparum-infected RBCs secreted EVs with a complex role in promoting the pathogenesis of the disease by eliciting the production of cytokines and immunosuppression in various immune cells of the host [62,64,75]. Trypanosoma and Leishmania EVs also facilitate the progression of the disease by immunosuppression [53,58,99]. T. cruzi EVs have been observed to stimulate the secretion of both pro- and anti-inflammatory cytokines, and consequently play various regulatory roles in the host’s cells [106]. However, sometimes, EVs can also stimulate the host’s immune response and promote clearance of the disease by the host. Giardia EVs stimulate the secretion of IL-1β, IL-6, and TNF-α, which enhance protection against the disease [2]. EVs from N. fowleri stimulate the production of IL-8 from the macrophages that would affect the pathogenesis of the disease [104].
3. Conclusions
The data demonstrate that EVs play a central role in the interplay between parasites and the host cells. Parasites control and modulate the cargo of not only the EVs secreted directly by them into the extracellular environment, but also of the EVs secreted by the host cells they infect. This ability to manipulate the content of host-derived EVs grants a distinct advantage to the parasites concealed within the host cells, enabling them to wield their influence over distant target cells.
Recent research has identified newer effector molecules in the cargo. Functional small RNA species such as tsRNA and miRNA, along with associated proteins such as argonautes, could help regulate the gene expression profiles in distant cells.
EVs initiate a spectrum of immune responses from the host and are capable of inducing both pro-inflammatory and anti-inflammatory effects. Several studies have provided evidence that the cargo inside EVs is dynamically regulated by the parasites in adapting to their immediate environment and conditions. Therefore, the same parasitic EVs can elicit different immune responses at different stages of the lifecycle, indicating the complex and nuanced modulation of the host’s immune response.
Understanding the precise role of EVs in the physiological and natural contexts demands more sophisticated approaches. Despite these challenges, the study of EVs in parasite–host interactions holds great promise in advancing our understanding of infectious diseases and potential therapeutic targets.
Author Contributions
M.S., D.L.-A., D.C. and U.S. all contributed to the review. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olmos-Ortiz, L.M.; Barajas-Mendiola, M.A.; Barrios-Rodiles, M.; Castellano, L.E.; Arias-Negrete, S.; Avila, E.E.; Cuellar-Mata, P. Trichomonas vaginalis exosome-like vesicles modify the cytokine profile and reduce inflammation in parasite-infected mice. Parasite Immunol. 2017, 39, e12426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cao, L.; Wang, X.; Li, J.; Dong, J.; Zhang, N.; Li, X.; Li, S.; Sun, M.; Zhang, X.; et al. Giardia duodenalis extracellular vesicles regulate the proinflammatory immune response in mouse macrophages in vitro via the MAPK, AKT and NF-kappaB pathways. Parasites Vectors 2021, 14, 358. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Soekmadji, C.; Hill, A.F.; Wauben, M.H.; Buzas, E.I.; Di Vizio, D.; Falcon-Perez, J.M.; Gardiner, C.; Hochberg, F.; Kurochkin, I.V.; et al. Updating the MISEV minimal requirements for extracellular vesicle studies: Building bridges to reproducibility. J. Extracell. Vesicles 2017, 6, 1396823. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Goberdhan, D.C.; O’Driscoll, L.; Thery, C.; Welsh, J.A.; Blenkiron, C.; Buzas, E.I.; Di Vizio, D.; Erdbrugger, U.; Falcon-Perez, J.M.; et al. Updating MISEV: Evolving the minimal requirements for studies of extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12182. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Nievas, Y.R.; Coceres, V.M.; Midlej, V.; de Souza, W.; Benchimol, M.; Pereira-Neves, A.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J.; de Miguel, N. Membrane-shed vesicles from the parasite Trichomonas vaginalis: Characterization and their association with cell interaction. Cell Mol. Life Sci. 2018, 75, 2211–2226. [Google Scholar] [CrossRef]
- Dixson, A.C.; Dawson, T.R.; Di Vizio, D.; Weaver, A.M. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat. Rev. Mol. Cell Biol. 2023, 24, 454–476. [Google Scholar] [CrossRef]
- Kerviel, A.; Zhang, M.; Altan-Bonnet, N. A New Infectious Unit: Extracellular Vesicles Carrying Virus Populations. Annu. Rev. Cell Dev. Biol. 2021, 37, 171–197. [Google Scholar] [CrossRef]
- Atayde, V.D.; da Silva Lira Filho, A.; Chaparro, V.; Zimmermann, A.; Martel, C.; Jaramillo, M.; Olivier, M. Exploitation of the Leishmania exosomal pathway by Leishmania RNA virus 1. Nat. Microbiol. 2019, 4, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Hrdy, I.; Zdrha, A.; Narayanasamy, R.K.; Smutna, T.; Horackova, J.; Harant, K.; Benes, V.; Ong, S.C.; Tsai, C.Y.; et al. Double-Stranded RNA Viruses Are Released From Trichomonas vaginalis Inside Small Extracellular Vesicles and Modulate the Exosomal Cargo. Front. Microbiol. 2022, 13, 893692. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Zhang, H.; Ehrenkaufer, G.; Singh, U. Stress Response in Entamoeba histolytica Is Associated with Robust Processing of tRNA to tRNA Halves. mBio 2023, 14, e0345022. [Google Scholar] [CrossRef]
- Buzas, E.I. The roles of extracellular vesicles in the immune system. Nat. Rev. Immunol. 2023, 23, 236–250. [Google Scholar] [CrossRef]
- Sutcliffe, S.; Alderete, J.F.; Till, C.; Goodman, P.J.; Hsing, A.W.; Zenilman, J.M.; De Marzo, A.M.; Platz, E.A. Trichomonosis and subsequent risk of prostate cancer in the Prostate Cancer Prevention Trial. Int. J. Cancer 2009, 124, 2082–2087. [Google Scholar] [CrossRef]
- Tsang, S.H.; Peisch, S.F.; Rowan, B.; Markt, S.C.; Gonzalez-Feliciano, A.G.; Sutcliffe, S.; Platz, E.A.; Mucci, L.A.; Ebot, E.M. Association between Trichomonas vaginalis and prostate cancer mortality. Int. J. Cancer 2019, 144, 2377–2380. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, W.; Wang, H.; Wang, Y.; Li, J.; Wu, X. Trichomonas vaginalis infection-associated risk of cervical cancer: A meta-analysis. Eur. J. Obs. Gynecol. Reprod. Biol. 2018, 228, 166–173. [Google Scholar] [CrossRef]
- Twu, O.; de Miguel, N.; Lustig, G.; Stevens, G.C.; Vashisht, A.A.; Wohlschlegel, J.A.; Johnson, P.J. Trichomonas vaginalis exosomes deliver cargo to host cells and mediate hostratioparasite interactions. PLoS Pathog. 2013, 9, e1003482. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.S.; Zhou, H.C.; Wei, C.Y.; Wang, Z.H.; Hao, X.; Zhang, L.H.; Li, J.Z.; Wang, Z.L.; Wang, H. Global survey of miRNAs and tRNA-derived small RNAs from the human parasitic protist Trichomonas vaginalis. Parasit. Vectors 2021, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Govender, Y.; Chan, T.; Yamamoto, H.S.; Budnik, B.; Fichorova, R.N. The Role of Small Extracellular Vesicles in Viral-Protozoan Symbiosis: Lessons From Trichomonasvirus in an Isogenic Host Parasite Model. Front. Cell. Infect. Microbiol. 2020, 10, 591172. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Johnson, P.J. Trichomonas vaginalis extracellular vesicles are internalized by host cells using proteoglycans and caveolin-dependent endocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 21354–21360. [Google Scholar] [CrossRef]
- Nogueira, P.M.; de Menezes-Neto, A.; Borges, V.M.; Descoteaux, A.; Torrecilhas, A.C.; Xander, P.; Revach, O.Y.; Regev-Rudzki, N.; Soares, R.P. Immunomodulatory Properties of Leishmania Extracellular Vesicles During Host-Parasite Interaction: Differential Activation of TLRs and NF-kappaB Translocation by Dermotropic and Viscerotropic Species. Front. Cell. Infect. Microbiol. 2020, 10, 380. [Google Scholar] [CrossRef]
- Stanley, S.L. Pathophysiology of amoebiasis. Trends Parasitol. 2001, 17, 280–285. [Google Scholar] [CrossRef]
- Sharma, M.; Morgado, P.; Zhang, H.; Ehrenkaufer, G.; Manna, D.; Singh, U. Characterization of Extracellular Vesicles from Entamoeba histolytica Identifies Roles in Intercellular Communication That Regulates Parasite Growth and Development. Infect. Immun. 2020, 88, 10.1128. [Google Scholar] [CrossRef]
- Diaz-Godinez, C.; Rios-Valencia, D.G.; Garcia-Aguirre, S.; Martinez-Calvillo, S.; Carrero, J.C. Immunomodulatory effect of extracellular vesicles from Entamoeba histolytica trophozoites: Regulation of NETs and respiratory burst during confrontation with human neutrophils. Front. Cell. Infect. Microbiol. 2022, 12, 1018314. [Google Scholar] [CrossRef]
- Zhang, H.; Ehrenkaufer, G.M.; Pompey, J.M.; Hackney, J.A.; Singh, U. Small RNAs with 5’-polyphosphate termini associate with a Piwi-related protein and regulate gene expression in the single-celled eukaryote Entamoeba histolytica. PLoS Pathog. 2008, 4, e1000219. [Google Scholar] [CrossRef]
- Zhang, H.; Ehrenkaufer, G.M.; Hall, N.; Singh, U. Small RNA pyrosequencing in the protozoan parasite Entamoeba histolytica reveals strain-specific small RNAs that target virulence genes. BMC Genom. 2013, 14, 53. [Google Scholar] [CrossRef]
- Vazquezdelara-Cisneros, L.G.; Arroyo-Begovich, A. Induction of encystation of Entamoeba invadens by removal of glucose from the culture medium. J. Parasitol. 1984, 70, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef]
- Xu, F.; Jimenez-Gonzalez, A.; Einarsson, E.; Astvaldsson, A.; Peirasmaki, D.; Eckmann, L.; Andersson, J.O.; Svard, S.G.; Jerlstrom-Hultqvist, J. The compact genome of Giardia muris reveals important steps in the evolution of intestinal protozoan parasites. Microb. Genom. 2020, 6, e000402. [Google Scholar] [CrossRef] [PubMed]
- Evans-Osses, I.; Mojoli, A.; Monguio-Tortajada, M.; Marcilla, A.; Aran, V.; Amorim, M.; Inal, J.; Borras, F.E.; Ramirez, M.I. Microvesicles released from Giardia intestinalis disturb host-pathogen response in vitro. Eur. J. Cell Biol. 2017, 96, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Pollet, H.; Conrard, L.; Cloos, A.S.; Tyteca, D. Plasma Membrane Lipid Domains as Platforms for Vesicle Biogenesis and Shedding? Biomolecules 2018, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cao, L.; Wang, X.; Dong, J.; Zhang, N.; Li, X.; Li, J.; Zhang, X.; Gong, P. Extracellular vesicles secreted by Giardia duodenalis regulate host cell innate immunity via TLR2 and NLRP3 inflammasome signaling pathways. PLoS Negl. Trop. Dis. 2021, 15, e0009304. [Google Scholar] [CrossRef]
- Grajeda, B.I.; De Chatterjee, A.; Villalobos, C.M.; Pence, B.C.; Ellis, C.C.; Enriquez, V.; Roy, S.; Roychowdhury, S.; Neumann, A.K.; Almeida, I.C.; et al. Giardial lipid rafts share virulence factors with secreted vesicles and participate in parasitic infection in mice. Front. Cell. Infect. Microbiol. 2022, 12, 974200. [Google Scholar] [CrossRef]
- Natali, L.; Luna Pizarro, G.; Moyano, S.; de la Cruz-Thea, B.; Musso, J.; Ropolo, A.S.; Eichner, N.; Meister, G.; Musri, M.M.; Feliziani, C.; et al. The Exosome-like Vesicles of Giardia Assemblages A, B, and E Are Involved in the Delivering of Distinct Small RNA from Parasite to Parasite. Int. J. Mol. Sci. 2023, 24, 9559. [Google Scholar] [CrossRef]
- Pace, D. Leishmaniasis. J. Infect. 2014, 69 (Suppl. S1), S10–S18. [Google Scholar] [CrossRef]
- da Silva Lira Filho, A.; Fajardo, E.F.; Chang, K.P.; Clement, P.; Olivier, M. Leishmania Exosomes/Extracellular Vesicles Containing GP63 Are Essential for Enhance Cutaneous Leishmaniasis Development Upon Co-Inoculation of Leishmania amazonensis and Its Exosomes. Front. Cell. Infect. Microbiol. 2021, 11, 709258. [Google Scholar] [CrossRef]
- Weber, J.I.; Rodrigues, A.V.; Valerio-Bolas, A.; Nunes, T.; Carvalheiro, M.; Antunes, W.; Alexandre-Pires, G.; da Fonseca, I.P.; Santos-Gomes, G. Insights on Host-Parasite Immunomodulation Mediated by Extracellular Vesicles of Cutaneous Leishmania shawi and Leishmania guyanensis. Cells 2023, 12, 1101. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.M.; Reiner, N.E. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Front. Cell. Infect. Microbiol. 2011, 1, 26. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Mukherjee, A.; Adak, S. Glyceraldehyde-3-phosphate dehydrogenase present in extracellular vesicles from Leishmania major suppresses host TNF-alpha expression. J. Biol. Chem. 2021, 297, 101198. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Kolli, B.K.; Yadava, N.; Lu, H.G.; Gilman-Sachs, A.; Peterson, D.A.; Chang, K.P. Episomal expression of specific sense and antisense mRNAs in Leishmania amazonensis: Modulation of gp63 level in promastigotes and their infection of macrophages in vitro. Infect. Immun. 2000, 68, 80–86. [Google Scholar] [CrossRef]
- Gomez, M.A.; Contreras, I.; Halle, M.; Tremblay, M.L.; McMaster, R.W.; Olivier, M. Leishmania GP63 alters host signaling through cleavage-activated protein tyrosine phosphatases. Sci. Signal 2009, 2, ra58. [Google Scholar] [CrossRef]
- Halle, M.; Gomez, M.A.; Stuible, M.; Shimizu, H.; McMaster, W.R.; Olivier, M.; Tremblay, M.L. The Leishmania surface protease GP63 cleaves multiple intracellular proteins and actively participates in p38 mitogen-activated protein kinase inactivation. J. Biol. Chem. 2009, 284, 6893–6908. [Google Scholar] [CrossRef]
- Isnard, A.; Shio, M.T.; Olivier, M. Impact of Leishmania metalloprotease GP63 on macrophage signaling. Front. Cell. Infect. Microbiol. 2012, 2, 72. [Google Scholar] [CrossRef]
- Silverman, J.M.; Chan, S.K.; Robinson, D.P.; Dwyer, D.M.; Nandan, D.; Foster, L.J.; Reiner, N.E. Proteomic analysis of the secretome of Leishmania donovani. Genome Biol. 2008, 9, R35. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania exosomes modulate innate and adaptive immune responses through effects on monocytes and dendritic cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef]
- Smirlis, D.; Dingli, F.; Sabatet, V.; Roth, A.; Knippschild, U.; Loew, D.; Spath, G.F.; Rachidi, N. Identification of the Host Substratome of Leishmania-Secreted Casein Kinase 1 Using a SILAC-Based Quantitative Mass Spectrometry Assay. Front. Cell. Dev. Biol. 2021, 9, 800098. [Google Scholar] [CrossRef]
- Lambertz, U.; Oviedo Ovando, M.E.; Vasconcelos, E.J.; Unrau, P.J.; Myler, P.J.; Reiner, N.E. Small RNAs derived from tRNAs and rRNAs are highly enriched in exosomes from both old and new world Leishmania providing evidence for conserved exosomal RNA Packaging. BMC Genom. 2015, 16, 151. [Google Scholar] [CrossRef]
- Kusakisako, K.; Nakao, R.; Katakura, K. Detection of parasite-derived tRNA and rRNA fragments in the peripheral blood of mice experimentally infected with Leishmania donovani and Leishmania amazonensis using next-generation sequencing analysis. Parasitol. Int. 2023, 93, 102716. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; de’Oliveira, C.C.; Shirvani, O.; Fang, Y.; Wang, C.; Foster, L.J.; Reiner, N.E. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci. 2010, 123, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.M.C.; Dupin, T.V.; Toledo, M.D.S.; Reis, N.; Ribeiro, K.; Cronemberger-Andrade, A.; Rugani, J.N.; De Lorenzo, B.H.P.; Novaes, E.B.R.R.; Soares, R.P.; et al. Extracellular Vesicles Released by Leishmania (Leishmania) amazonensis Promote Disease Progression and Induce the Production of Different Cytokines in Macrophages and B-1 Cells. Front. Microbiol. 2018, 9, 3056. [Google Scholar] [CrossRef] [PubMed]
- Atayde, V.D.; Aslan, H.; Townsend, S.; Hassani, K.; Kamhawi, S.; Olivier, M. Exosome Secretion by the Parasitic Protozoan Leishmania within the Sand Fly Midgut. Cell Rep. 2015, 13, 957–967. [Google Scholar] [CrossRef]
- Douanne, N.; Dong, G.; Douanne, M.; Olivier, M.; Fernandez-Prada, C. Unravelling the proteomic signature of extracellular vesicles released by drug-resistant Leishmania infantum parasites. PLoS Negl. Trop. Dis. 2020, 14, e0008439. [Google Scholar] [CrossRef]
- Douanne, N.; Dong, G.; Amin, A.; Bernardo, L.; Blanchette, M.; Langlais, D.; Olivier, M.; Fernandez-Prada, C. Leishmania parasites exchange drug-resistance genes through extracellular vesicles. Cell Rep. 2022, 40, 111121. [Google Scholar] [CrossRef]
- Castelli, G.; Bruno, F.; Saieva, L.; Alessandro, R.; Galluzzi, L.; Diotallevi, A.; Vitale, F. Exosome secretion by Leishmania infantum modulate the chemotactic behavior and cytokinic expression creating an environment permissive for early infection. Exp. Parasitol. 2019, 198, 39–45. [Google Scholar] [CrossRef]
- Gioseffi, A.; Hamerly, T.; Van, K.; Zhang, N.; Dinglasan, R.R.; Yates, P.A.; Kima, P.E. Leishmania-infected macrophages release extracellular vesicles that can promote lesion development. Life Sci. Alliance 2020, 3, e202000742. [Google Scholar] [CrossRef]
- Emerson, L.E.; Gioseffi, A.; Barker, H.; Sheppe, A.; Morrill, J.K.; Edelmann, M.J.; Kima, P.E. Leishmania infection-derived extracellular vesicles drive transcription of genes involved in M2 polarization. Front. Cell. Infect. Microbiol. 2022, 12, 934611. [Google Scholar] [CrossRef]
- Aly, A.S.; Vaughan, A.M.; Kappe, S.H. Malaria parasite development in the mosquito and infection of the mammalian host. Annu. Rev. Microbiol. 2009, 63, 195–221. [Google Scholar] [CrossRef] [PubMed]
- Opadokun, T.; Rohrbach, P. Extracellular vesicles in malaria: An agglomeration of two decades of research. Malar. J. 2021, 20, 442. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, N.G.; Cheng, L.; Eriksson, E.M. The role of extracellular vesicles in malaria biology and pathogenesis. Malar. J. 2017, 16, 245. [Google Scholar] [CrossRef] [PubMed]
- Babatunde, K.A.; Yesodha Subramanian, B.; Ahouidi, A.D.; Martinez Murillo, P.; Walch, M.; Mantel, P.Y. Role of Extracellular Vesicles in Cellular Cross Talk in Malaria. Front. Immunol. 2020, 11, 22. [Google Scholar] [CrossRef]
- Opadokun, T.; Agyapong, J.; Rohrbach, P. Protein Profiling of Malaria-Derived Extracellular Vesicles Reveals Distinct Subtypes. Membranes 2022, 12, 397. [Google Scholar] [CrossRef]
- Vimonpatranon, S.; Roytrakul, S.; Phaonakrop, N.; Lekmanee, K.; Atipimonpat, A.; Srimark, N.; Sukapirom, K.; Chotivanich, K.; Khowawisetsut, L.; Pattanapanyasat, K. Extracellular Vesicles Derived from Early and Late Stage Plasmodium falciparum-Infected Red Blood Cells Contain Invasion-Associated Proteins. J. Clin. Med. 2022, 11, 4250. [Google Scholar] [CrossRef]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef]
- Abdi, A.; Yu, L.; Goulding, D.; Rono, M.K.; Bejon, P.; Choudhary, J.; Rayner, J. Proteomic analysis of extracellular vesicles from a Plasmodium falciparum Kenyan clinical isolate defines a core parasite secretome. Wellcome Open Res. 2017, 2, 50. [Google Scholar] [CrossRef]
- Abou Karam, P.; Rosenhek-Goldian, I.; Ziv, T.; Ben Ami Pilo, H.; Azuri, I.; Rivkin, A.; Kiper, E.; Rotkopf, R.; Cohen, S.R.; Torrecilhas, A.C.; et al. Malaria parasites release vesicle subpopulations with signatures of different destinations. EMBO Rep. 2022, 23, e54755. [Google Scholar] [CrossRef]
- Sisquella, X.; Ofir-Birin, Y.; Pimentel, M.A.; Cheng, L.; Abou Karam, P.; Sampaio, N.G.; Penington, J.S.; Connolly, D.; Giladi, T.; Scicluna, B.J.; et al. Malaria parasite DNA-harbouring vesicles activate cytosolic immune sensors. Nat. Commun. 2017, 8, 1985. [Google Scholar] [CrossRef]
- Ofir-Birin, Y.; Ben Ami Pilo, H.; Cruz Camacho, A.; Rudik, A.; Rivkin, A.; Revach, O.Y.; Nir, N.; Block Tamin, T.; Abou Karam, P.; Kiper, E.; et al. Malaria parasites both repress host CXCL10 and use it as a cue for growth acceleration. Nat. Commun. 2021, 12, 4851. [Google Scholar] [CrossRef] [PubMed]
- Mantel, P.Y.; Hjelmqvist, D.; Walch, M.; Kharoubi-Hess, S.; Nilsson, S.; Ravel, D.; Ribeiro, M.; Gruring, C.; Ma, S.; Padmanabhan, P.; et al. Infected erythrocyte-derived extracellular vesicles alter vascular function via regulatory Ago2-miRNA complexes in malaria. Nat. Commun. 2016, 7, 12727. [Google Scholar] [CrossRef] [PubMed]
- Babatunde, K.A.; Mbagwu, S.; Hernandez-Castaneda, M.A.; Adapa, S.R.; Walch, M.; Filgueira, L.; Falquet, L.; Jiang, R.H.Y.; Ghiran, I.; Mantel, P.Y. Malaria infected red blood cells release small regulatory RNAs through extracellular vesicles. Sci. Rep. 2018, 8, 884. [Google Scholar] [CrossRef]
- Wang, Z.; Wei, C.; Hao, X.; Deng, W.; Zhang, L.; Wang, Z.; Wang, H. Genome-wide identification and characterization of transfer RNA-derived small RNAs in Plasmodium falciparum. Parasit Vectors 2019, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Ofir-Birin, Y.; Regev-Rudzki, N. Extracellular vesicles in parasite survival. Science 2019, 363, 817–818. [Google Scholar] [CrossRef]
- Dekel, E.; Yaffe, D.; Rosenhek-Goldian, I.; Ben-Nissan, G.; Ofir-Birin, Y.; Morandi, M.I.; Ziv, T.; Sisquella, X.; Pimentel, M.A.; Nebl, T.; et al. 20S proteasomes secreted by the malaria parasite promote its growth. Nat. Commun. 2021, 12, 1172. [Google Scholar] [CrossRef]
- Sampaio, N.G.; Emery, S.J.; Garnham, A.L.; Tan, Q.Y.; Sisquella, X.; Pimentel, M.A.; Jex, A.R.; Regev-Rudzki, N.; Schofield, L.; Eriksson, E.M. Extracellular vesicles from early stage Plasmodium falciparum-infected red blood cells contain PfEMP1 and induce transcriptional changes in human monocytes. Cell Microbiol. 2018, 20, e12822. [Google Scholar] [CrossRef]
- Correa, R.; Coronado, L.; Caballero, Z.; Faral-Tello, P.; Robello, C.; Spadafora, C. Extracellular vesicles carrying lactate dehydrogenase induce suicide in increased population density of Plasmodium falciparum in vitro. Sci. Rep. 2019, 9, 5042. [Google Scholar] [CrossRef]
- Silva, V.O.; Maia, M.M.; Torrecilhas, A.C.; Taniwaki, N.N.; Namiyama, G.M.; Oliveira, K.C.; Ribeiro, K.S.; Toledo, M.D.S.; Xander, P.; Pereira-Chioccola, V.L. Extracellular vesicles isolated from Toxoplasma gondii induce host immune response. Parasite Immunol. 2018, 40, e12571. [Google Scholar] [CrossRef]
- Quiarim, T.M.; Maia, M.M.; da Cruz, A.B.; Taniwaki, N.N.; Namiyama, G.M.; Pereira-Chioccola, V.L. Characterization of extracellular vesicles isolated from types I, II and III strains of Toxoplasma gondii. Acta Trop. 2021, 219, 105915. [Google Scholar] [CrossRef]
- Retana Moreira, L.; Rodriguez Serrano, F.; Osuna, A. Extracellular vesicles of Trypanosoma cruzi tissue-culture cell-derived trypomastigotes: Induction of physiological changes in non-parasitized culture cells. PLoS Negl. Trop. Dis. 2019, 13, e0007163. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.M.; da Cruz, A.B.; Taniwaki, N.N.; Namiyama, G.M.; Gava, R.; Gomes, A.H.S.; Kanamura, C.T.; Barbo, M.L.P.; Pereira-Chioccola, V.L. Immunization with extracellular vesicles excreted by Toxoplasma gondii confers protection in murine infection, activating cellular and humoral responses. Int. J. Parasitol. 2021, 51, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Chavez, F.; Murrieta-Coxca, J.M.; Caballero-Ortega, H.; Morales-Prieto, D.M.; Markert, U.R. Host-pathogen interactions mediated by extracellular vesicles in Toxoplasma gondii infection during pregnancy. J. Reprod. Immunol. 2023, 158, 103957. [Google Scholar] [CrossRef] [PubMed]
- Wowk, P.F.; Zardo, M.L.; Miot, H.T.; Goldenberg, S.; Carvalho, P.C.; Morking, P.A. Proteomic profiling of extracellular vesicles secreted from Toxoplasma gondii. Proteomics 2017, 17, 201600477. [Google Scholar] [CrossRef]
- Pope, S.M.; Lasser, C. Toxoplasma gondii infection of fibroblasts causes the production of exosome-like vesicles containing a unique array of mRNA and miRNA transcripts compared to serum starvation. J. Extracell. Vesicles 2013, 2, 22484. [Google Scholar] [CrossRef]
- Aline, F.; Bout, D.; Amigorena, S.; Roingeard, P.; Dimier-Poisson, I. Toxoplasma gondii antigen-pulsed-dendritic cell-derived exosomes induce a protective immune response against T. gondii infection. Infect. Immun. 2004, 72, 4127–4137. [Google Scholar] [CrossRef]
- Beauvillain, C.; Ruiz, S.; Guiton, R.; Bout, D.; Dimier-Poisson, I. A vaccine based on exosomes secreted by a dendritic cell line confers protection against T. gondii infection in syngeneic and allogeneic mice. Microbes Infect. 2007, 9, 1614–1622. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Xiu, F.; Wang, J.; Cong, H.; He, S.; Shi, Y.; Wang, X.; Li, X.; Zhou, H. Characterization of exosomes derived from Toxoplasma gondii and their functions in modulating immune responses. Int. J. Nanomed. 2018, 13, 467–477. [Google Scholar] [CrossRef]
- Tedford, E.; Badya, N.B.; Laing, C.; Asaoka, N.; Kaneko, S.; Filippi, B.M.; McConkey, G.A. Infection-induced extracellular vesicles evoke neuronal transcriptional and epigenetic changes. Sci. Rep. 2023, 13, 6913. [Google Scholar] [CrossRef]
- Da Silveira, J.F.; Abrahamsohn, P.A.; Colli, W. Plasma membrane vesicles isolated from epimastigote forms of Trypanosoma cruzi. Biochim. Biophys. Acta 1979, 550, 222–232. [Google Scholar] [CrossRef]
- Bayer-Santos, E.; Aguilar-Bonavides, C.; Rodrigues, S.P.; Cordero, E.M.; Marques, A.F.; Varela-Ramirez, A.; Choi, H.; Yoshida, N.; da Silveira, J.F.; Almeida, I.C. Proteomic analysis of Trypanosoma cruzi secretome: Characterization of two populations of extracellular vesicles and soluble proteins. J. Proteome. Res. 2013, 12, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, R.M.; Ricart, C.A.; Machado, M.O.; Bastos, I.M.; de Santana, J.M.; de Sousa, M.V.; Roepstorff, P.; Charneau, S. Insight into the Exoproteome of the Tissue-Derived Trypomastigote form of Trypanosoma cruzi. Front. Chem. 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Lopez, N.L.; Ndao, M.; Camargo, F.V.; Nara, T.; Annoura, T.; Hardie, D.B.; Borchers, C.H.; Jardim, A. Characterization and Diagnostic Application of Trypanosoma cruzi Trypomastigote Excreted-Secreted Antigens Shed in Extracellular Vesicles Released from Infected Mammalian Cells. J. Clin. Microbiol. 2017, 55, 744–758. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, K.S.; Vasconcellos, C.I.; Soares, R.P.; Mendes, M.T.; Ellis, C.C.; Aguilera-Flores, M.; de Almeida, I.C.; Schenkman, S.; Iwai, L.K.; Torrecilhas, A.C. Proteomic analysis reveals different composition of extracellular vesicles released by two Trypanosoma cruzi strains associated with their distinct interaction with host cells. J. Extracell. Vesicles 2018, 7, 1463779. [Google Scholar] [CrossRef]
- Bayer-Santos, E.; Lima, F.M.; Ruiz, J.C.; Almeida, I.C.; da Silveira, J.F. Characterization of the small RNA content of Trypanosoma cruzi extracellular vesicles. Mol. Biochem. Parasitol. 2014, 193, 71–74. [Google Scholar] [CrossRef]
- Garcia-Silva, M.R.; Cabrera-Cabrera, F.; das Neves, R.F.; Souto-Padron, T.; de Souza, W.; Cayota, A. Gene expression changes induced by Trypanosoma cruzi shed microvesicles in mammalian host cells: Relevance of tRNA-derived halves. Biomed. Res. Int. 2014, 2014, 305239. [Google Scholar] [CrossRef]
- Garcia-Silva, M.R.; das Neves, R.F.; Cabrera-Cabrera, F.; Sanguinetti, J.; Medeiros, L.C.; Robello, C.; Naya, H.; Fernandez-Calero, T.; Souto-Padron, T.; de Souza, W.; et al. Extracellular vesicles shed by Trypanosoma cruzi are linked to small RNA pathways, life cycle regulation, and susceptibility to infection of mammalian cells. Parasitol. Res. 2014, 113, 285–304. [Google Scholar] [CrossRef]
- Cornet-Gomez, A.; Retana Moreira, L.; Kronenberger, T.; Osuna, A. Extracellular vesicles of trypomastigotes of Trypanosoma cruzi induce changes in ubiquitin-related processes, cell-signaling pathways and apoptosis. Sci. Rep. 2023, 13, 7618. [Google Scholar] [CrossRef]
- Cronemberger-Andrade, A.; Xander, P.; Soares, R.P.; Pessoa, N.L.; Campos, M.A.; Ellis, C.C.; Grajeda, B.; Ofir-Birin, Y.; Almeida, I.C.; Regev-Rudzki, N.; et al. Trypanosoma cruzi-Infected Human Macrophages Shed Proinflammatory Extracellular Vesicles That Enhance Host-Cell Invasion via Toll-Like Receptor 2. Front. Cell. Infect. Microbiol. 2020, 10, 99. [Google Scholar] [CrossRef]
- Szempruch, A.J.; Sykes, S.E.; Kieft, R.; Dennison, L.; Becker, A.C.; Gartrell, A.; Martin, W.J.; Nakayasu, E.S.; Almeida, I.C.; Hajduk, S.L.; et al. Extracellular Vesicles from Trypanosoma brucei Mediate Virulence Factor Transfer and Cause Host Anemia. Cell 2016, 164, 246–257. [Google Scholar] [CrossRef]
- Eliaz, D.; Kannan, S.; Shaked, H.; Arvatz, G.; Tkacz, I.D.; Binder, L.; Waldman Ben-Asher, H.; Okalang, U.; Chikne, V.; Cohen-Chalamish, S.; et al. Exosome secretion affects social motility in Trypanosoma brucei. PLoS Pathog. 2017, 13, e1006245. [Google Scholar] [CrossRef] [PubMed]
- Ekici, A.; Alkan, S.; Aydemir, S.; Gurbuz, E.; Unlu, A.H. Trends in Naegleria fowleri global research: A bibliometric analysis study. Acta Trop. 2022, 234, 106603. [Google Scholar] [CrossRef] [PubMed]
- Retana Moreira, L.; Steller Espinoza, M.F.; Chacon Camacho, N.; Cornet-Gomez, A.; Saenz-Arce, G.; Osuna, A.; Lomonte, B.; Abrahams Sandi, E. Characterization of Extracellular Vesicles Secreted by a Clinical Isolate of Naegleria fowleri and Identification of Immunogenic Components within Their Protein Cargo. Biology 2022, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Lertjuthaporn, S.; Somkird, J.; Lekmanee, K.; Atipimonpat, A.; Sukapirom, K.; Sawasdipokin, H.; Tiewcharoen, S.; Pattanapanyasat, K.; Khowawisetsut, L. Extracellular Vesicles from Naegleria fowleri Induce IL-8 Response in THP-1 Macrophage. Pathogens 2022, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.H.; Jeong, S.R.; Kim, J.H.; Song, K.J.; Kim, K.; Park, S.; Sohn, S.; Shin, H.J. Cytopathic changes and pro-inflammatory cytokines induced by Naegleria fowleri trophozoites in rat microglial cells and protective effects of an anti-Nfa1 antibody. Parasite Immunol. 2005, 27, 453–459. [Google Scholar] [CrossRef]
- Cortes-Serra, N.; Gualdron-Lopez, M.; Pinazo, M.J.; Torrecilhas, A.C.; Fernandez-Becerra, C. Extracellular Vesicles in Trypanosoma cruzi Infection: Immunomodulatory Effects and Future Perspectives as Potential Control Tools against Chagas Disease. J. Immunol. Res. 2022, 2022, 5230603. [Google Scholar] [CrossRef]
Figure 1.
Examples of the biological functions mediated by EVs.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharma, M.; Lozano-Amado, D.; Chowdhury, D.; Singh, U. Extracellular Vesicles and Their Impact on the Biology of Protozoan Parasites. Trop. Med. Infect. Dis. 2023, 8, 448. https://doi.org/10.3390/tropicalmed8090448
AMA Style
Sharma M, Lozano-Amado D, Chowdhury D, Singh U. Extracellular Vesicles and Their Impact on the Biology of Protozoan Parasites. Tropical Medicine and Infectious Disease. 2023; 8(9):448. https://doi.org/10.3390/tropicalmed8090448
Chicago/Turabian StyleSharma, Manu, Daniela Lozano-Amado, Debabrata Chowdhury, and Upinder Singh. 2023. "Extracellular Vesicles and Their Impact on the Biology of Protozoan Parasites" Tropical Medicine and Infectious Disease 8, no. 9: 448. https://doi.org/10.3390/tropicalmed8090448