

Genetic Diversity of Plasmodium vivax Field Isolates from the Thai–Myanmar Border during the Period of 2006–2016


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Sample Collection and Plasmodium Diagnosis
2.3. Genomic DNA Extraction
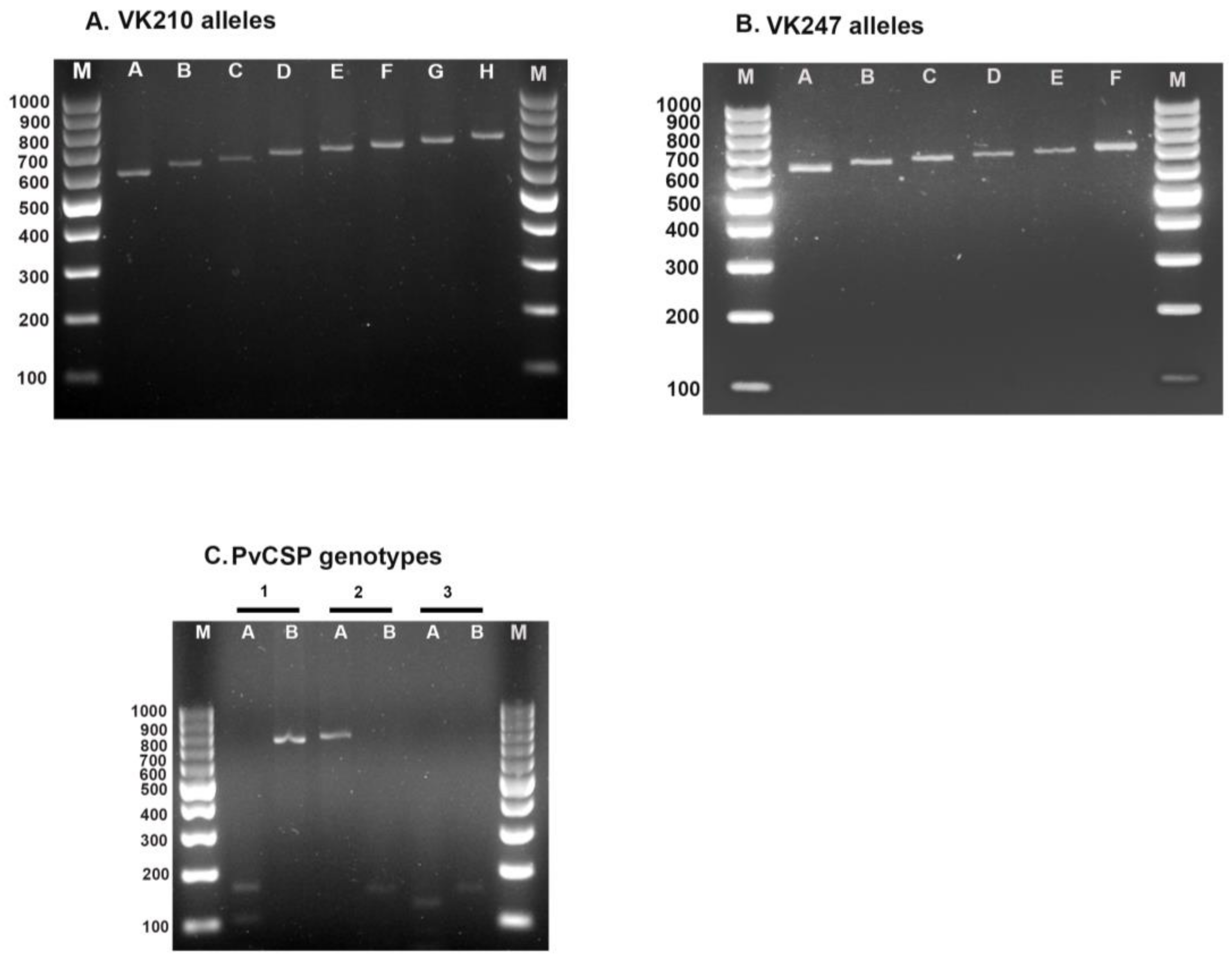
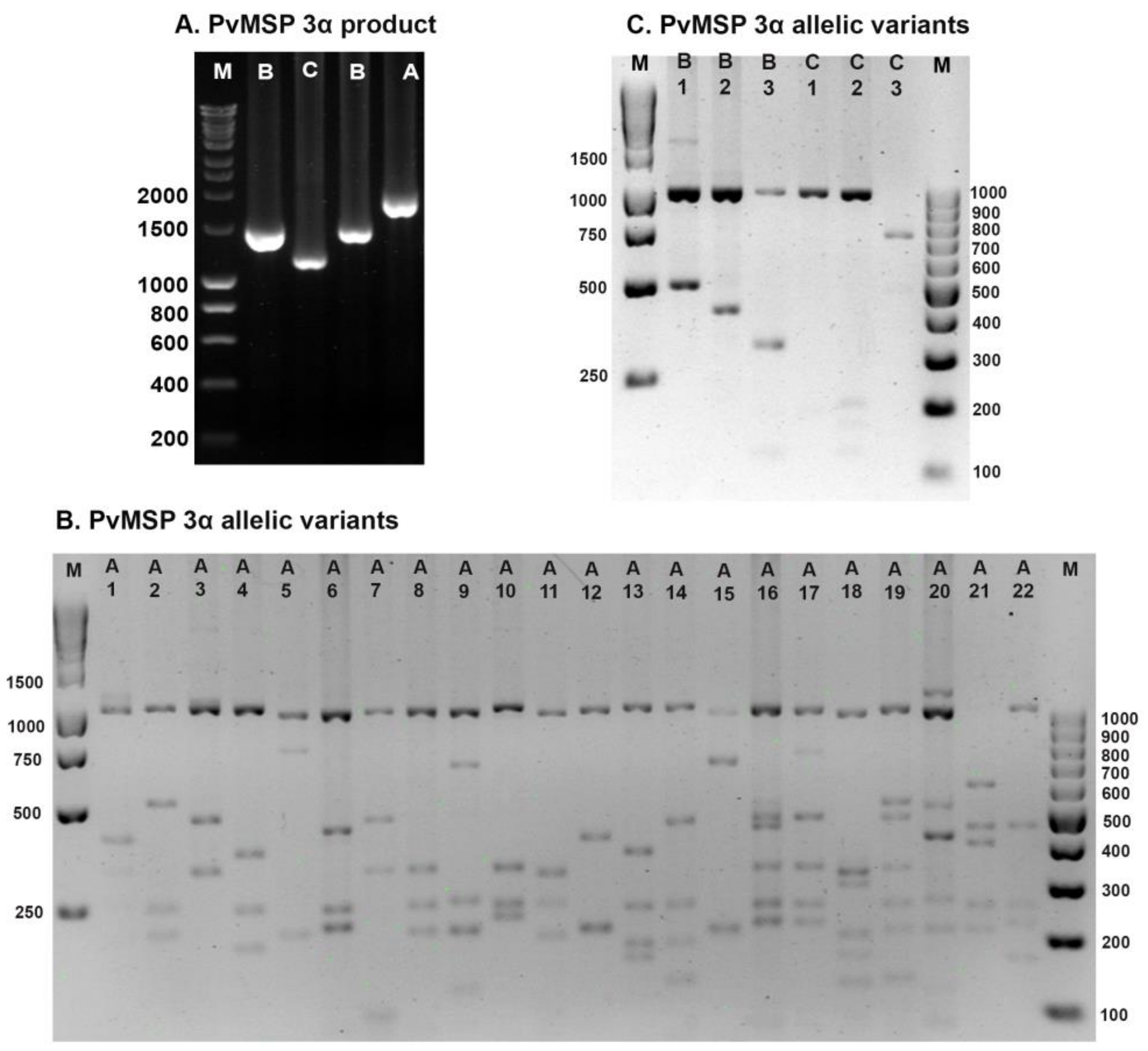
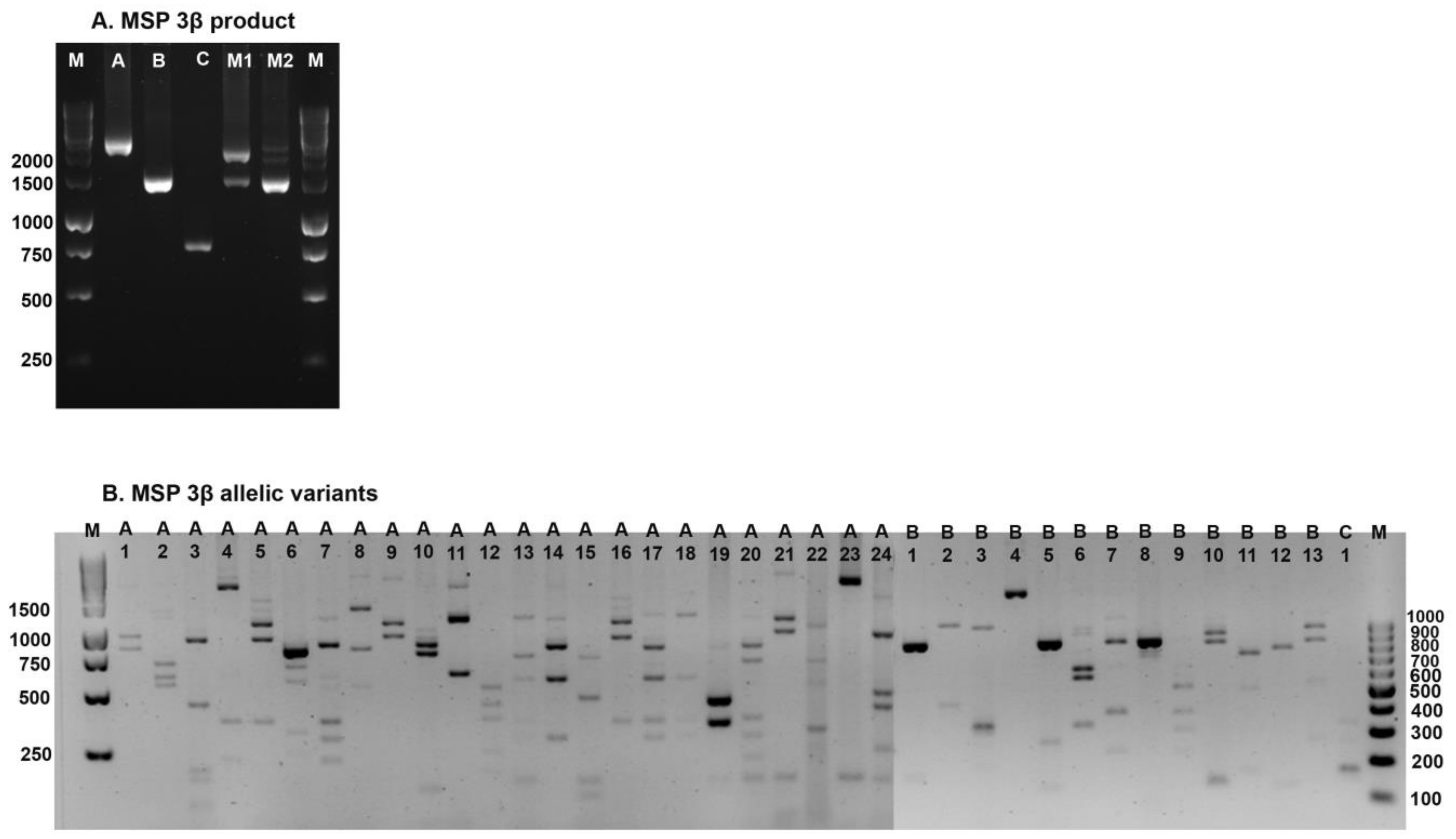
2.4. P. vivax Parasite Genotyping
2.5. Statistical Analysis
3. Results
3.1. Genetic Diversity of the PvCSP
3.2. Genetic Diversity of the PvMSP-3α
3.3. Genetic Diversity of the PvMSP-3β
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenwood, B.M.; Fidock, D.A.; Kyle, D.E.; Kappe, S.H.; Alonso, P.L.; Collins, F.H.; Duffy, P.E. Malaria: Progress, perils, and prospects for eradication. J. Clin. Investig. 2008, 118, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howes, R.E.; Battle, K.E.; Mendis, K.N.; Smith, D.L.; Cibulskis, R.E.; Baird, J.K.; Hay, S.I. Global Epidemiology of Plasmodium vivax. Am. J. Trop. Med. Hyg. 2016, 95, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Suphakhonchuwong, N.; Chaijaroenkul, W.; Rungsihirunrat, K.; Na-Bangchang, K.; Kuesap, J. Evaluation of Plasmodium vivax isolates in Thailand using polymorphic markers Plasmodium merozoite Surface Protein (PvMSP) 1 and PvMSP3. Parasitol. Res. 2018, 117, 3965–3978. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.F.; Khattak, A.A.; Zeeshan, N.; Zahid, H.; Awan, U.A.; Yaqoob, A.; Ashraf, N.M.; Gul, S.; Alam, S.; Ahmed, W. Surveillance of molecular markers of antimalarial drug resistance in Plasmodium falciparum and Plasmodium vivax in Federally Administered Tribal Area (FATA), Pakistan. Rev. Inst. Med. Trop. Sao Paulo. 2021, 63. [Google Scholar] [CrossRef]
- Crompton, P.D.; Pierce, S.K.; Miller, L.H. Advances and challenges in malaria vaccine development. J. Clin. Investig. 2010, 120, 4168–4178. [Google Scholar] [CrossRef]
- Barry, A.E.; Arnott, A. Strategies for designing and monitoring malaria vaccines targeting diverse antigens. Front. Immunol. 2014, 5, 359. [Google Scholar] [CrossRef] [Green Version]
- Thakur, A.; Alam, M.T.; Sharma, Y.D. Genetic diversity in the C-terminal 42 kDa region of merozoite surface protein-1 of Plasmodium vivax (PvMSP-142) among Indian isolates. Acta Trop. 2008, 108, 58–63. [Google Scholar] [CrossRef]
- Mzilahowa, T.; McCall, P.J.; Hastings, I.M. “Sexual” population structure and genetics of the malaria agent P. falciparum. PLoS ONE 2007, 2, e613. [Google Scholar] [CrossRef] [Green Version]
- Rungsihirunrat, K.; Chaijaroenkul, W.; Siripoon, N.; Seugorn, A.; Na-Bangchang, K. Genotyping of polymorphic marker (MSP3α and MSP3β) genes of Plasmodium vivax field isolates from malaria endemic of Thailand. Trop. Med. Int. Health 2011, 16, 794–801. [Google Scholar] [CrossRef]
- Bahk, Y.Y.; Kim, J.; Ahn, S.K.; Na, B.K.; Chai, J.Y.; Kim, T.S. Genetic Diversity of Plasmodium vivax Causing Epidemic Malaria in the Republic of Korea. Korean J. Parasitol. 2018, 56, 545–552. [Google Scholar] [CrossRef]
- Zakeri, S.; Barjesteh, H.; Djadid, N.D. Merozoite surface protein-3α is a reliable marker for population genetic analysis of Plasmodium vivax. Malar. J. 2006, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Huang, S.; Su, X.Z.; Guo, H.; Xu, Y.; Xu, F.; Hu, X.; Yang, Y.; Wang, S.; Lu, F. Genetic diversity of Plasmodium vivax population in Anhui province of China. Malar. J. 2014, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- López, C.; Yepes-Pérez, Y.; Hincapié-Escobar, N.; Díaz-Arévalo, D.; Patarroyo, M.A. What is known about the immune response induced by Plasmodium vivax malaria vaccine candidates? Front. Immunol. 2017, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Qari, S.H.; Shi, V.-P.; Povoa, M.M.; Alpers, M.P.; Deloron, P.; Murphy, G.S.; Harjosuwarno, S.; Lal, A.A. Global occurrence of Plasmodium vivax-like human malaria parasite. J. Infect. Dis. 1993, 168, 1485–1489. [Google Scholar] [CrossRef]
- Kaur, H.; Sehgal, R.; Kumar, A.; Sehgal, A.; Bharti, P.K.; Bansal, D.; Mohapatra, P.K.; Mahanta, J.; Sultan, A.A. Exploration of genetic diversity of Plasmodium vivax circumsporozoite protein (Pvcsp) and Plasmodium vivax sexual stage antigen (Pvs25) among North Indian isolates. Malar. J. 2019, 18, 1–8. [Google Scholar] [CrossRef]
- Jiang, J.; Barnwell, J.W.; Meyer, E.V.; Galinski, M.R. Plasmodium vivax merozoite surface protein-3 (PvMSP3): Expression of an 11 member multigene family in blood-stage parasites. PLoS ONE 2013, 8, e63888. [Google Scholar] [CrossRef] [Green Version]
- Kuesap, J.; Rungsihirunrat, K.; Chaijaroenkul, W.; Mungthin, M. Genetic diversity of Plasmodium vivax merozoite surface protein-3 alpha and beta from diverse geographic areas of Thailand. Jpn. J. Infect. Dis. 2022, 75, 241–248. [Google Scholar] [CrossRef]
- Yang, Z.; Miao, J.; Huang, Y.; Li, X.; Putaporntip, C.; Jongwutiwes, S.; Gao, Q.; Udomsangpetch, R.; Sattabongkot, J.; Cui, L. Genetic structures of geographically distinct Plasmodium vivax populations assessed by PCR/RFLP analysis of the merozoite surface protein 3β gene. Acta Trop. 2006, 100, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Bruce, M.C.; Galinski, M.R.; Barnwell, J.W.; Snounou, G.; Day, K.P. Polymorphism at the merozoite surface protein-3alpha locus of Plasmodium vivax: Global and local diversity. Am. J. Trop. Med. Hyg. 1999, 61, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Imwong, M.; Pukrittayakamee, S.; Grüner, A.C.; Rénia, L.; Letourneur, F.; Looareesuwan, S.; White, N.J.; Snounou, G. Practical PCR genotyping protocols for Plasmodium vivax using Pvcs and Pvmsp1. Malar. J. 2005, 4, 1–13. [Google Scholar] [CrossRef]
- Maneerattanasak, S.; Gosi, P.; Krudsood, S.; Tongshoob, J.; Lanteri, C.A.; Snounou, G.; Khusmith, S. Genetic diversity among Plasmodium vivax isolates along the Thai–Myanmar border of Thailand. Malar. J. 2016, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Ceron, L.; Rodriguez, M.H.; Nettel, J.C.; Villarreal, C.; Kain, K.C.; Hernandez, J.E. Differential susceptibilities of Anopheles albimanus and Anopheles pseudopunctipennis to infections with coindigenous Plasmodium vivax variants VK210 and VK247 in southern Mexico. Infect. Immun. 1999, 67, 410–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosaisavee, V.; Hastings, I.; Craig, A.; Lek-Uthai, U. The genetic polymorphism of Plasmodium vivax genes in endemic regions of Thailand. Asian Pac. J. Trop. Med. 2011, 4, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Ghanchi, N.K.; Thaver, A.M.; Jafri, S.; Beg, M.A. Genetic diversity of Plasmodium vivax clinical isolates from southern Pakistan using pvcsp and pvmsp1 genetic markers. Malar. J. 2013, 12, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, J.A.; Validum, L.; Cummings, R.; Palmer, C.J. Genetic diversity of Plasmodium vivax pvcsp and pvmsp1 in Guyana, South America. Am. J. Trop. Med. Hyg. 2006, 75, 830–835. [Google Scholar] [CrossRef]
- González-Cerón, L.; Martinez-Barnetche, J.; Montero-Solís, C.; Santillán, F.; Soto, A.M.; Rodríguez, M.H.; Espinosa, B.J.; Chávez, O.A. Molecular epidemiology of Plasmodium vivax in Latin America: Polymorphism and evolutionary relationships of the circumsporozoite gene. Malar. J. 2013, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Chenet, S.M.; Tapia, L.L.; Escalante, A.A.; Durand, S.; Lucas, C.; Bacon, D.J. Genetic diversity and population structure of genes encoding vaccine candidate antigens of Plasmodium vivax. Malar. J. 2012, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.L.; Póvoa, M.M. Distribution of Plasmodium vivax variants (VK210, VK247 and P. vivax-like) in three endemic areas of the Amazon region of Brazil and their correlation with chloroquine treatment. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 377–381. [Google Scholar] [CrossRef]
- Da Silva, A.N.; Santos, C.C.; Lacerda, R.N.; Machado, R.L.; Póvoa, M.M. Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247. Mem. Inst. Oswaldo Cruz. 2006, 101, 547–550. [Google Scholar] [CrossRef]
- Kim, J.-R.; Imwong, M.; Nandy, A.; Chotivanich, K.; Nontprasert, A.; Tonomsing, N.; Maji, A.; Addy, M.; Day, N.P.; White, N.J. Genetic diversity of Plasmodium vivax in Kolkata, India. Malar. J. 2006, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Mascorro, C.N.; Fan, Q.; Rzomp, K.A.; Khuntirat, B.; Zhou, G.; Chen, H.; Yan, G.; Sattabongkot, J. Genetic diversity and multiple infections of Plasmodium vivax malaria in Western Thailand. Am. J. Trop. Med. Hyg. 2003, 68, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bai, Y.; Wu, Y.; Zeng, W.; Xiang, Z.; Zhao, H.; Zhao, W.; Chen, X.; Duan, M.; Wang, X. PvMSP-3α and PvMSP-3β genotyping reveals higher genetic diversity in Plasmodium vivax parasites from migrant workers than residents at the China-Myanmar border. Infect. Genet. Evol. 2022, 106, 105387. [Google Scholar] [CrossRef]
- Kaul, A.; Bali, P.; Anwar, S.; Sharma, A.K.; Gupta, B.K.; Singh, O.P.; Adak, T.; Sohail, M. Genetic diversity and allelic variation in MSP3α gene of paired clinical Plasmodium vivax isolates from Delhi, India. J. Infect. Public Health. 2019, 12, 576–584. [Google Scholar] [CrossRef]
- Khan, S.N.; Khan, A.; Khan, S.; Ayaz, S.; Attaullah, S.; Khan, J.; Khan, M.A.; Ali, I.; Shah, A.H. PCR/RFLP-based analysis of genetically distinct Plasmodium vivax population of Pvmsp-3α and Pvmsp-3β genes in Pakistan. Malar. J. 2014, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-C.; Wang, G.-Z.; Meng, F.; Zeng, W.; He, C.-h.; Hu, X.-M.; Wang, S.-Q. Genetic diversity of Plasmodium vivax population before elimination of malaria in Hainan Province, China. Malar. J. 2015, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Joshi, H.; Singh, V.; Anvikar, A.; Valecha, N. Plasmodium vivax msp-3α polymorphisms: Analysis in the Indian subcontinent. Malar. J. 2016, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.-U.; Lee, H.-W.; Kim, J.-Y.; Na, B.-K.; Cho, S.-H.; Lin, K.; Sohn, W.-M.; Kim, T.-S. High frequency of genetic diversity of Plasmodium vivax field isolates in Myanmar. Acta Trop. 2009, 109, 30–36. [Google Scholar] [CrossRef]
- Zakeri, S.; Raeisi, A.; Afsharpad, M.; Kakar, Q.; Ghasemi, F.; Atta, H.; Zamani, G.; Memon, M.S.; Salehi, M.; Djadid, N.D. Molecular characterization of Plasmodium vivax clinical isolates in Pakistan and Iran using pvmsp-1, pvmsp-3α and pvcsp genes as molecular markers. Parasitol. Int. 2010, 59, 15–21. [Google Scholar] [CrossRef]
- Thanapongpichat, S.; Khammanee, T.; Sawangjaroen, N.; Buncherd, H.; Tun, A.W. Genetic diversity of Plasmodium vivax in clinical isolates from Southern Thailand using PvMSP1, PvMSP3 (PvMSP3α, PvMSP3β) genes and eight microsatellite markers. Korean J. Parasitol. 2019, 57, 469. [Google Scholar] [CrossRef] [Green Version]
- Kibria, M.G.; Elahi, R.; Mohon, A.N.; Khan, W.A.; Haque, R.; Alam, M.S. Genetic diversity of Plasmodium vivax in clinical isolates from Bangladesh. Malar. J. 2015, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| First Period 1 | Second Period | |||
|---|---|---|---|---|
| Sai Yok | Mae Sot | Total | Mae Sot | |
| N % | N % | N % | N % | |
| VK210 | 160 (92.5) | 136 (95.8) | 296 (94.0) | 89 (87.3) |
| VK247 | 11 (6.4) | 6 (4.2) | 17 (5.4) | 13 (12.7) |
| Mixed | 2 (1.2) | 0 (0.0) | 2 (0.6) | 0 (0.0) |
| Total | 173 (100.0) | 142 (100.0) | 315 (100.0) | 102 (100.0) |
| First Period 1 | Second Period | ||||||
|---|---|---|---|---|---|---|---|
| Sai Yok | Mae Sot 3 | Mae Sot | |||||
| N | % | N | % | N | % | ||
| Type A (77.3 %) | 136 | 78.6 | 78 | 75 | Type A (78.1%) | 75 | 78.1 |
| A1 | 8 | 4.6 | 3 | 2.9 | A3 | 4 | 4.2 |
| A2 | 1 | 0.6 | 0 | 0.0 | A4 | 1 | 1.0 |
| A3 | 15 | 8.7 | 4 | 3.8 | A5 | 3 | 3.1 |
| A4 | 5 | 2.9 | 2 | 1.9 | A7 | 1 | 1.0 |
| A5 | 8 | 4.6 | 8 | 7.7 | A8 | 12 | 12.5 |
| A6 | 5 | 2.9 | 0 | 0.0 | A10 | 2 | 2.1 |
| A7 | 1 | 0.6 | 1 | 1.0 | A11 | 31 | 32.3 |
| A8 | 31 | 17.9 | 11 | 10.6 | A12 | 5 | 5.2 |
| A9 | 7 | 4.0 | 8 | 7.7 | A13 | 12 | 12.5 |
| A10 | 2 | 1.2 | 0 | 0.0 | A18 | 1 | 1.0 |
| A11 | 1 | 0.6 | 11 | 10.6 | A19 | 1 | 1.0 |
| A12 | 11 | 6.4 | 9 | 8.7 | A22 | 2 | 2.1 |
| A13 | 4 | 2.3 | 2 | 1.9 | Type B (21.9%) | 21 | 21.9 |
| A14 | 2 | 1.2 | 2 | 1.9 | B2 | 7 | 7.3 |
| A15 | 0 | 0.0 | 3 | 2.9 | B3 | 14 | 14.6 |
| A16 | 1 | 0.6 | 3 | 2.9 | Total | 96 | 100.0 |
| A17 * | 2 | 1.2 | 2 | 1.9 | Pattern (14, 14.6%) | 14 | |
| A18 | 27 | 15.6 | 7 | 6.7 | |||
| A19 * | 4 | 2.3 | 1 | 1.0 | |||
| A20 | 1 | 0.6 | 0 | 0.0 | |||
| A21 | 0 | 0.0 | 1 | 1.0 | |||
| Type B (7.9 %) | 13 | 7.6 | 9 | 8.7 | |||
| B1 | 6 | 3.5 | 5 | 4.8 | |||
| B2 | 6 | 3.5 | 2 | 1.9 | |||
| B3 | 1 | 0.6 | 2 | 1.9 | |||
| Type C (13%) | 19 | 11.0 | 17 | 16.3 | |||
| C1 | 17 | 9.8 | 15 | 14.4 | |||
| C2 | 2 | 1.2 | 1 | 1.0 | |||
| C3 | 0 | 0.0 | 1 | 1.0 | |||
| M 2 (1.8%) | 5 | 2.9 | 0.0 | 0.0 | |||
| Total | 173 | 100.0 | 104 | 100.0 | |||
| Pattern (28, 10.1 %) | 25 | 23 | |||||
| First Period | Second Period | ||||||
|---|---|---|---|---|---|---|---|
| Sai Yok | Mae Sot | Mae Sot | |||||
| N | % | N | % | N | % | ||
| Type A (63.4%) | 102 | 63.0 | 55 | 66.3 | Type A (51.2%) | 41 | 51.2 |
| A1 | 32 | 19.4 | 16 | 19.3 | A1 | 14 | 17.5 |
| A2 | 2 | 1.2 | 0 | 0.0 | A3 | 2 | 2.5 |
| A3 | 0 | 0.0 | 4 | 4.8 | A4 | 1 | 1.3 |
| A4 | 3 | 1.8 | 1 | 1.2 | A7 * | 8 | 10.0 |
| A5 * | 3 | 1.8 | 0 | 0.0 | A10 | 2 | 2.5 |
| A6 * | 0 | 0.0 | 1 | 1.2 | A11 | 2 | 2.5 |
| A7 * | 11 | 6.7 | 7 | 8.4 | A12 | 1 | 1.3 |
| A8 * | 5 | 3 | 0 | 0.0 | A14 | 10 | 12.5 |
| A9 | 0 | 0.0 | 1 | 1.2 | A23 | 1 | 1.3 |
| A10 | 1 | 0.6 | 2 | 2.4 | Type B (47.5%) | 38 | 47.5 |
| A11 | 23 | 13.9 | 9 | 10.8 | B1 | 2 | 2.5 |
| A13 * | 2 | 1.2 | 0 | 0.0 | B2 | 1 | 1.3 |
| A14 | 1 | 0.6 | 2 | 2.4 | B3 | 2 | 2.5 |
| A15 | 2 | 1.2 | 1 | 1.2 | B4 | 22 | 27.5 |
| A16 | 4 | 2.4 | 7 | 8.4 | B5 | 2 | 2.5 |
| A17 | 3 | 1.8 | 1 | 1.2 | B6 | 1 | 1.3 |
| A18 * | 2 | 1.2 | 1 | 1.2 | B7 * | 2 | 2.5 |
| A19 | 1 | 0.6 | 0 | 0.0 | B8 | 1 | 1.3 |
| A20 * | 1 | 0.6 | 0 | 0.0 | B9 | 2 | 2.5 |
| A21 * | 3 | 1.8 | 0 | 0.0 | B12 | 3 | 3.8 |
| A22 * | 3 | 1.8 | 0 | 0.0 | Type M (1.3%) | 1 | 1.3 |
| A24 | 0 | 0.0 | 2 | 2.4 | Total | 80 | 100.0 |
| Type B (30.2%) | 52 | 30.3 | 23 | 27.7 | Pattern (20, 25% | 20 | |
| B1 | 2 | 1.2 | 3 | 3.6 | |||
| B2 | 2 | 1.2 | 0 | 0.0 | |||
| B3 | 4 | 2.4 | 1 | 1.2 | |||
| B4 | 27 | 16.4 | 14 | 16.9 | |||
| B5 | 3 | 1.8 | 0 | 0.0 | |||
| B6 * | 1 | 0.6 | 0 | 0.0 | |||
| B7 * | 2 | 1.2 | 0 | 0.0 | |||
| B8 | 1 | 0.6 | 0 | 0.0 | |||
| B9 | 5 | 3.0 | 2 | 2.4 | |||
| B10 * | 0 | 0.0 | 2 | 2.4 | |||
| B11 | 0 | 0.0 | 1 | 1.2 | |||
| B12 | 4 | 2.4 | 0 | 0.0 | |||
| B13 * | 1 | 0.6 | 0 | 0.0 | |||
| Type C (0.4 %) | 0 | 0.0 | 0 | 1.2 | |||
| C1 | 0 | 0.0 | 1 | 1.2 | |||
| Type M (6 %) | 11 | 6.7 | 4 | 4.8 | |||
| Total | 165 | 100.0 | 83 | 100.0 | |||
| Pattern (36, 14.5%) | 29 | 22 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalei, A.A.; Chaijaroenkul, W.; Na-Bangchang, K. Genetic Diversity of Plasmodium vivax Field Isolates from the Thai–Myanmar Border during the Period of 2006–2016. Trop. Med. Infect. Dis. 2023, 8, 210. https://doi.org/10.3390/tropicalmed8040210
Jalei AA, Chaijaroenkul W, Na-Bangchang K. Genetic Diversity of Plasmodium vivax Field Isolates from the Thai–Myanmar Border during the Period of 2006–2016. Tropical Medicine and Infectious Disease. 2023; 8(4):210. https://doi.org/10.3390/tropicalmed8040210
Chicago/Turabian StyleJalei, Abdifatah Abdullahi, Wanna Chaijaroenkul, and Kesara Na-Bangchang. 2023. "Genetic Diversity of Plasmodium vivax Field Isolates from the Thai–Myanmar Border during the Period of 2006–2016" Tropical Medicine and Infectious Disease 8, no. 4: 210. https://doi.org/10.3390/tropicalmed8040210