Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management


1
Institute of Crop Science and Resource Conservation (INRES), Plant Diseases and Plant Protection, University of Bonn, 53115 Bonn, Germany
2
Institute for Grapevine Breeding, Julius Kühn-Institut, 76833 Siebeldingen, Germany
3
Institute of Sugar Beet Research (IfZ), 37079 Goettingen, Germany
*
Author to whom correspondence should be addressed.
Toxins 2023, 15(3), 192; https://doi.org/10.3390/toxins15030192
Submission received: 23 January 2023
/
Revised: 28 February 2023
/
Accepted: 1 March 2023
/
Published: 3 March 2023
(This article belongs to the Special Issue Mycotoxin Spectrum in Food and Feed)
Abstract
:Fusarium head blight (FHB) is a major threat for wheat production worldwide. Most reviews focus on Fusarium graminearum as a main causal agent of FHB. However, different Fusarium species are involved in this disease complex. These species differ in their geographic adaptation and mycotoxin profile. The incidence of FHB epidemics is highly correlated with weather conditions, especially rainy days with warm temperatures at anthesis and an abundance of primary inoculum. Yield losses due to the disease can reach up to 80% of the crop. This review summarizes the Fusarium species involved in the FHB disease complex with the corresponding mycotoxin profiles, disease cycle, diagnostic methods, the history of FHB epidemics, and the management strategy of the disease. In addition, it discusses the role of remote sensing technology in the integrated management of the disease. This technology can accelerate the phenotyping process in the breeding programs aiming at FHB-resistant varieties. Moreover, it can support the decision-making strategies to apply fungicides via monitoring and early detection of the diseases under field conditions. It can also be used for selective harvest to avoid mycotoxin-contaminated plots in the field.
Keywords:
wheat scab; mycotoxins; yield losses; epidemics; monitoring; decision making; control measures; resistance; remote sensingKey Contribution: This review summarizes Fusarium species involved in Fusarium head blight on wheat with their mycotoxin profile; diagnostic and monitoring methods, and management strategies.
1. Introduction
Wheat is an essential food around the world, in addition to its use as feed and fiber for livestock and energy production. It is the first crop in the world with a harvested area of 219 million hectares/year and a production of 760 million tons/year (average of 10 years). During the last decade, wheat yield reached 3475 kg/ha with an average increase of 605 kg/ha over the previous decade [1]. The genus wheat (Triticum L. 1753) is a member of the grass family Poaceae with different cultivated species, mainly the tetraploid durum wheat Triticum durum 2n = 28 with the genome set AABB and the hexaploid wheat Triticum aestivum 2n = 42 with the genome set AABBDD [2]. Wheat yield is affected by biotic (pests and pathogens) and abiotic stresses as a result of environmental conditions, stress occurrence, and genetic prevalence. Among the biotic stresses, pathogens (i.e., fungi, viruses, and bacteria) may contribute to average global losses of 21.5% of wheat yield [3].
Fusarium head blight (FHB) is one of the most relevant fungal diseases of wheat associated with different fungal species from the genus Fusarium [4,5]. FHB causes significant losses in wheat yield because the affected grains are small, shrunken, of low mass and quality, and contaminated with mycotoxins, which are harmful to humans and in animal nutrition [6]. The main mycotoxins produced by Fusarium species are trichothecenes, zearalenones, fumonisins, and the emerging toxins, i.e., beauvericin, enniatins, fusaproliferin, and moniliformin [7]. FHB, also known as Fusarium ear blight or scab, was first described in England by Smith (1884) [8] as a new disease of wheat, barley, and ryegrass, attributing the infection to the causal agents Fusisporium culmorum, hordei and Lolii Wm.G. Sm. It was recorded as an important disease in the USA at the end of the 19th century. By the beginning of the 20th century, FHB was known in wheat production regions worldwide [5]. In the 1990s, McMullen et al. [9] characterized it as a re-emerging disease due to the frequent epidemics on wheat in the USA and Canada from 1991 to 1996. They assigned this to the fundamental changes in agricultural practices, mainly reduced tillage. FHB is also known as a disease complex since more than one species from the genus Fusarium are involved in this disease in addition to two species from the genus Microdochium [10]. The main difference between them is that Fusarium species produce a wide spectrum of mycotoxins while Microdochium species do not produce mycotoxins [10].
Because of the high toxicity of Fusarium mycotoxins and the effect of Fusarium head blight on wheat yield, it is important to integrate plant protection practices such as crop rotation, resistant varieties, and cultural practices up to the application of fungicide within the management strategy. To safeguard wheat yield and to produce products with high quality especially, the application of fungicides is necessary under certain environmental conditions. To increase knowledge about the relevant parameters of the epidemiology and to perform knowledge-based plant protection measures, it is important to investigate and develop new methods to predict and detect FHB epidemics early on cereals [11]. This is essential, for example, for making the decision to apply fungicides at a suitable time of infection.
Besides direct control, a highly effective strategy to control FHB is breeding varieties with appropriate resistance against this disease complex. Effective cultivar development needs interdisciplinary research, integrating plant breeding, phytopathology, and informatics. It also needs precise and innovative methods for identifying and characterizing disease symptoms at an early stage of pathogenicity [12]. Because conventional characterization of host plant genotypes is laborious, time-consuming and cost-intensive, this is a limiting factor in plant-breeding programs. Here, proximal sensing with optical sensors is a promising characterizing method. Various sensors are suitable for the detection, identification, and quantification of plant diseases, such as thermography, fluorescence, and spectral sensors [13]. Recently, hyperspectral imaging showed efficiency as a precise and non-destructive tool in characterizing the interaction of Fusarium spp. and wheat plants. It was efficiently used in quantifying wheat resistance to FHB [14], the assessment of Fusarium infection and mycotoxin contamination of wheat kernels and flour [15], and, combined with thermography and chlorophyll fluorescence, for the early detection and monitoring of FHB development on wheat [16].
Most available reviews focus on Fusarium graminearum as the main causal agent of FHB on wheat; therefore, this review summarizes the Fusarium species involved in the FHB disease complex with the corresponding mycotoxin profiles, disease cycle, and diagnostic methods. In addition, it displays the history of FHB epidemics, and the management strategy of the disease. Moreover, it discusses the role of remote sensing technology in diagnosing and phenotyping the disease symptoms on wheat spikes.
2. Fusarium Head Blight Epidemics on Wheat
The frequency of FHB epidemics has increased in the last decades due to the changes in agricultural practices, mainly the zero tillage regime in wheat fields, in addition to the increase in the area of wheat cultivation and the frequency of wheat in the crop rotation [17,18]. The incidence of FHB epidemics is highly correlated with weather conditions especially rainy days with warm temperatures at anthesis and an abundance of primary inoculum [17,18,19,20,21]. Several studies showed that FHB epidemics on wheat take place sporadically. In Europe, yield losses caused by FHB epidemics ranged between 40 and 50% in Romania and Hungary during the 1970s–1980s [5]. A recent study showed an increase in Fusarium mycotoxin contamination in European wheat during the years 2010–2019; however, there is a lack of data on FHB incidence in Europe [22]. Seven severe epidemics and 14 medium epidemics occurred during the second half of the last century in China. In 2012, more than 9.9 million hectares were affected in the main producing areas in China [23]. In Canada, FHB epidemics were recorded during the 1940s and 1980s [24]. In the last two decades, epidemics were reported in 11 out of 17 surveyed years with a disease severity of 1.5–57.8% [25]. In the 20th century, five severe epidemics were described in the USA from the 1910s to the 1930s. However, FHB re-emerged from 1991 to 1996 and caused a yield drop of up to 25% and economic losses of USD 1 billion [9]. It continued to occur frequently from 1997 to 2010 in several states in the USA, resulting in annual losses of up to 54.2% in 2003 in different states [6]. In Latin America, seventeen epidemics have been reported in Argentina from 1960 to 2012 with losses of up to 70% in some years [26]. Furthermore, a model-based assessment study analyzed the weather data from 1957 to 2006 in Brazil. It showed an increased FHB-risk index during the 1960s and higher frequency of high-risk years starting from 1990 [27]. Information about FHB epidemics in Australia are rare due to their sporadic nature. However, a severe epidemic was reported in 2010 with disease of 79% in some fields [20].
3. The Pathogen
3.1. Pathogen Taxonomy
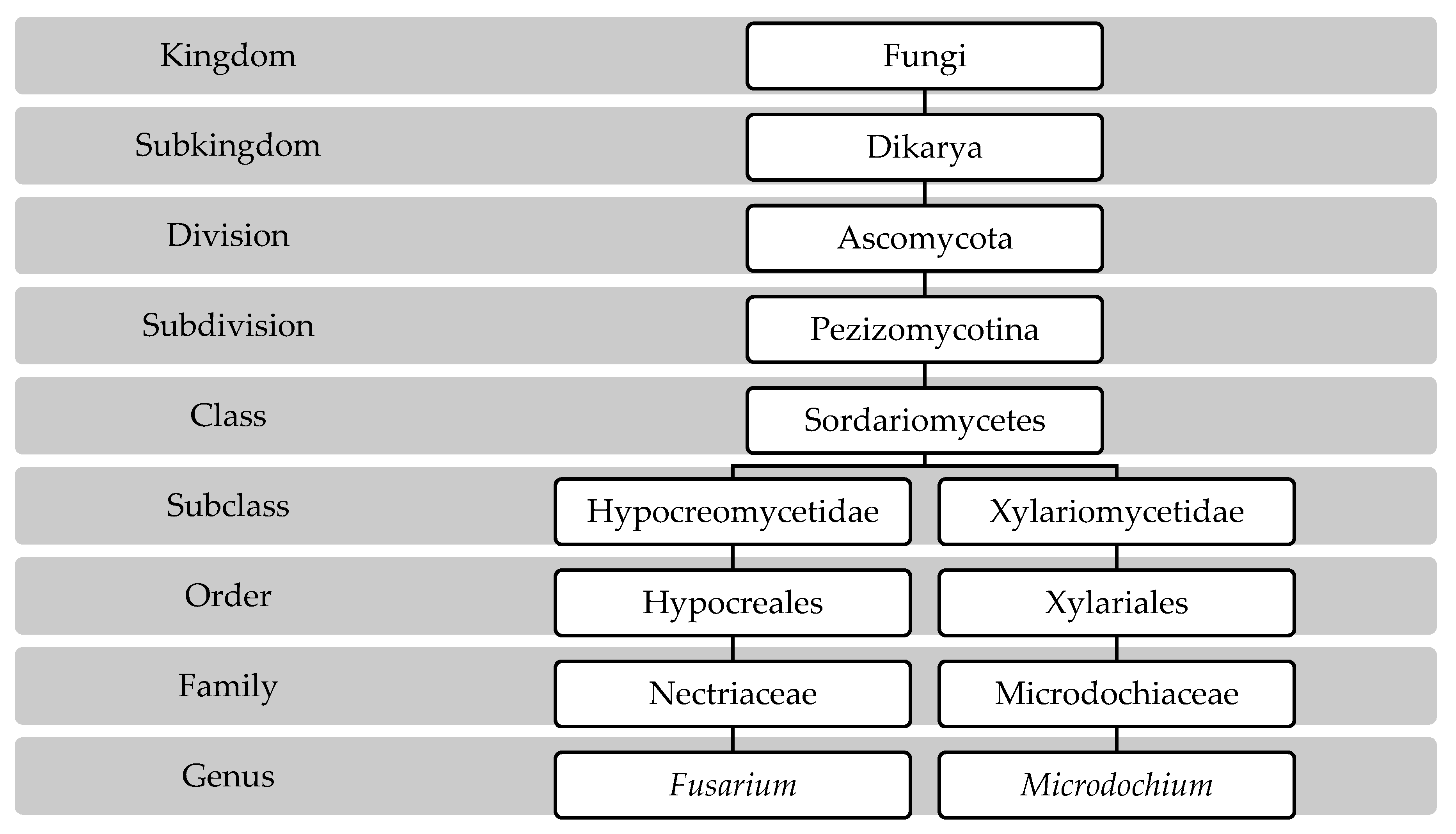
Since 2013, and after the changes in the International Code of Nomenclature for fungi, the name Fusarium was recognized as a unique name for all species including teleomorphs, which means that the name Gibberella and other names are not accepted anymore to indicate the sexual stage of these pathogens [28]. In addition, the subspecies of the genus Microdochium were elevated to species in 2005 and have become the accepted taxonomy. The following scheme shows the taxonomical position of the genus Fusarium and Microdochium according to the MycoBank database, 2022 [29] (Figure 1).
3.2. Fusarium Species Involved in FHB
Fusarium graminearum is the main pathogen of FHB worldwide [4,6,21,30]. However, different studies showed that other Fusarium species may contribute significantly to this disease in different areas of the world with different climate conditions. For example, F. graminearum, F. culmorum, F. avenaceum, F. poae, F. tricinctum, and M. majus were the dominant species in Europe [31,32,33]. In Canada, F. avenaceum, F. equiseti, F. graminearum, F. poae, and F. sporotrichioides were the most frequent species during the last two decades [25]. Alkadri et al. [34] recovered different Fusarium species from wheat in Syria. Table 1 shows the Fusarium species involved in FHB.
3.3. Life Cycle and Pathogenesis
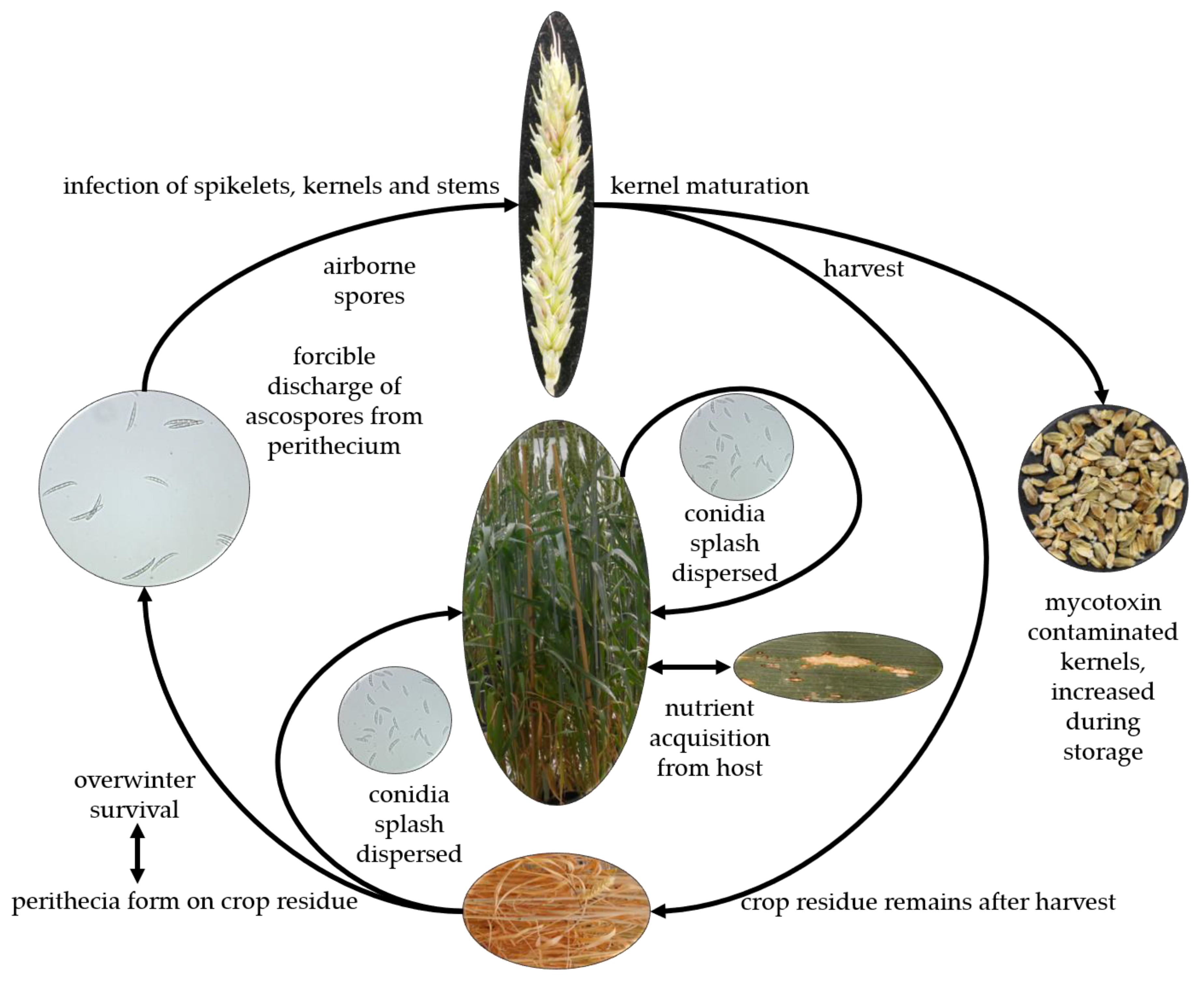
Fusarium head blight is a monocyclic disease (Figure 2). The pathogen survives in the debris of the previous crop as macroconidia or within sexual structures called perithecia as ascospores; or as asexual spores called macroconidia or microconidia for the species that have only anamorph stage. These spores are considered the primary inoculum of the disease. In addition, gramineous and non-gramineous weeds are not only a host range of Fusarium but they can also serve as an alternative host and source of inoculum. At wheat anthesis, which is the susceptible stage for infection, and in favorable weather conditions, the inoculum is blown by the wind or splashed by rain and lands on open spikelets. On the spikelet tissue, the spores germinate and produce germination tubes [6,21,35,36,37,38]. After germination, the fungal hyphae spread on the surface of the ovary, palea, and lemma and start to produce mycotoxins without penetrating the spikelet tissue. Thereafter, the pathogen penetrates the host tissue, starting a biotrophic infection with an intercellular growth in the spikelet and turns to the necrotrophic stage with inter- and intracellular growth laterally and vertically within the spike (Figure 2). During this stage of pathogenicity, mycotoxins accumulate in the spike tissue as well as in the kernels, reducing the crop yield and quality [39,40,41].
3.4. Symptoms
The first symptoms appear as water-soaked spots on the infected spikelets; these symptoms develop to necrosis and in an advanced stage, the infected spikelets become bleached (Figure 2). The bleaching spreads to cover the entire spike resulting in premature white wheat heads. Under warm and humid weather conditions, pinkish-red mycelium appears on the infected tissue. The kernels resulting from the infected spikes are known as tombstones because they are light in weight, shriveled, discolored with a pinkish or chalky appearance, and poor quality (Figure 2) [6,9,35,42,43]. However, some less virulent pathogens, such as F. poae, may cause infection and result in high levels of mycotoxin contamination in the infected kernels without detectable symptoms on the spikelets or the spike [44,45].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fusarium and Microdochium species involved in Fusarium head blight on wheat with their teleomorph and mycotoxin profile.
Table 1.
Fusarium and Microdochium species involved in Fusarium head blight on wheat with their teleomorph and mycotoxin profile.
| Pathogen | Teleomorph | Mycotoxin Profile | Reference | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trichothecenes | Zearalenone | Fusaric Acid | Fumonisins | Emerging Toxins | |||||||||||||||||
| Trichothecenes A | Trichothecenes B | Enniatins | Beauvericin | Fusaproliferin | Moniliformin | ||||||||||||||||
| T-2 toxin | HT-2 toxin | DAS | MAS | NEO | NX-2 | NX-3 | DON | 3-ADON | 15-ADON | NIV | 4-ANIV | ||||||||||
| F. acuminatum Ellis & Everh. | + | + | + | + | + | + | + | [46,47,48] | |||||||||||||
| F. avenaceum (Fr.) Sacc. | + | + | + | + | [46,47,49,50] | ||||||||||||||||
| F. crookwellense L.W. Burgess, P.E. Nelson & Toussoun synonym F. cerealis (Cooke) Sacc. | - | + | + | + | [47,50,51,52] | ||||||||||||||||
| F. culmorum (W.G. Sm.) Sacc. | - | + | + | + | + | + | + | [34,39,53,54,55,56] | |||||||||||||
| F. equiseti (Corda) Sacc. | + | + | + | + | + | + | + | + | [34,48,57] | ||||||||||||
| F. graminearum Schwabe | + | + | + | + | + | + | + | + | + | [34,46,53,56,58,59,60] | |||||||||||
| F. lateritium Nees | + | + | [61,62] | ||||||||||||||||||
| F. oxysporum Schltdl. | - | + | + | [7,63] | |||||||||||||||||
| F. poae (Peck) Wollenw. | - | + | + | + | + | + | + | + | + | [7,46,49,57,64] | |||||||||||
| F. proliferatum (Matsush.) Nirenberg | + | + | + | + | + | + | + | [49,65,66] | |||||||||||||
| F. sambucinum Fuckel | + | + | + | + | + | + | + | + | + | + | [49,61,67] | ||||||||||
| F. semitectum Berk. & Ravenel | - | + | + | [68,69] | |||||||||||||||||
| F. sporotrichioides Sherb. | - | + | + | + | + | + | + | + | + | + | [49,57,70] | ||||||||||
| F. subglutinans (Wollenw. & Reinking) P.E. Nelson, Toussoun & Marasas | + | + | + | + | + | [7,61,71] | |||||||||||||||
| F. tricinctum (Corda) Sacc. | + | + | + | [49] | |||||||||||||||||
| F. verticillioides (Sacc.) Nirenberg synonym F. moniliforme J. Sheld. | + | + | + | [7,72] | |||||||||||||||||
| Microdochium nivale (Fr.) Samuels & I.C. Hallett | + | [10] | |||||||||||||||||||
| Microdochium majus (Wollenw.) Glynn & S.G.Edwards | + | [10] | |||||||||||||||||||
Diacetoxyscirpenol (DAS); monoacetoxyscirpenol (MAS); neosolaniol (NEO); deoxynivalenol (DON); 3-acetyl-deoxynivalenol (3-ADON); 15-acetyl-deoxynivalenol (15-ADON); nivalenol (NIV); 4-acetyl-nivalenol (4-ANIV); zearalenone (ZEA); fusaric acid (FA); fumonisins (FUMs); enniatins (ENs); beauvericin (BEA); fusaproliferin (FP); moniliformin (MON).
4. Mycotoxins
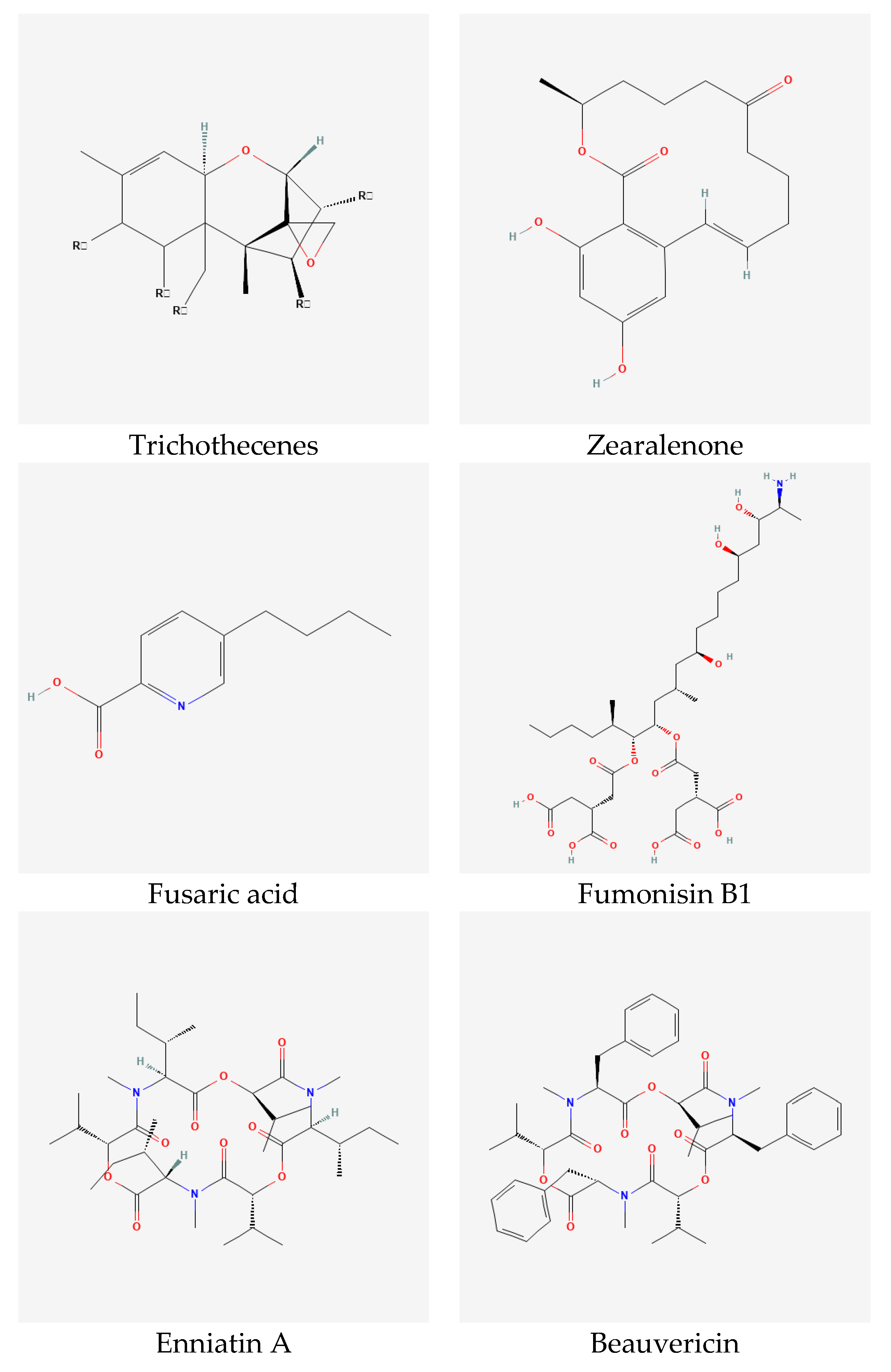
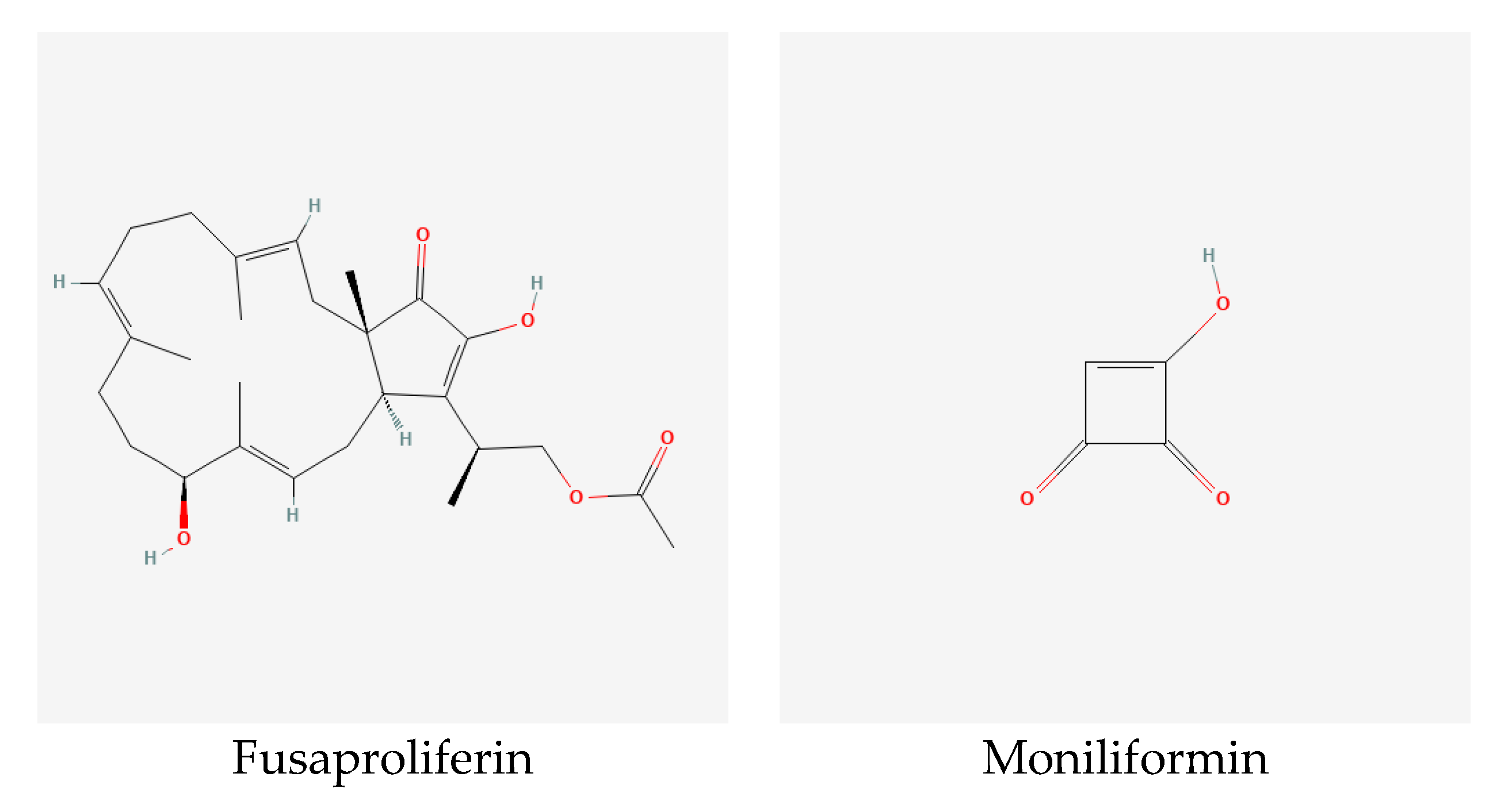
Mycotoxins are secondary metabolites produced by mold fungi such as Alternaria, Aspergillus, Claviceps, Fusarium, and Penicillium. Fusarium species involved in FHB produce a wide range of mycotoxins, mainly trichothecenes, zearalenone, fusaric acid, fumonisins, and emerging toxins, i.e., enniatins, beauvericin, moniliformin, and fusaproliferin (Figure 3). Due to the toxic effect of these metabolites on human and animal health, it is important to detect and quantify these toxins in food and feed. However, this process is challenging because it is expensive, time-consuming and laborious [15].
4.1. Trichothecenes
Trichothecenes are the most dominant group of Fusarium mycotoxins accompanying FHB infection on wheat worldwide [73]. This group is split, based on its chemical structure, into four subgroups A, B, C, and D [74]. However, trichothecenes produced by Fusarium spp. are A and B. The main difference between these two groups is the presence of ketone (=O) at C8 of trichothecenes backbone in trichothecenes B while it is absent in trichothecenes A [73]. In general, trichothecenes A are more toxic in Animalia compared with trichothecenes B; however, in Planta, trichothecenes B are more toxic [75]. Trichothecenes A include T-2 toxin, HT-2 toxin, diacetoxyscirpenol (DAS), monoacetoxyscirpenol (MAS), neosolaniol (NEO), NX-2 and NX-3. This group is mainly produced by F. acuminatum, F. equiseti, F. graminearum, F. poae, F. sambucinum, and F. sporotrichioides. Trichothecenes B include nivalenol (NIV), 4-acetyl-nivalenol (4-ANIV), deoxynivalenol (DON), 3-acetyl-deoxynivalenol (3-ADON) and 15-acetyl-deoxynivalenol (15-ADON). Fusarium species that produce trichothecenes B are F. acuminatum, F. crookwellense, F. culmorum, F. equiseti, F. graminearum, F. poae, F. sambucinum, F. semitectum, and F. sporotrichioides (Table 1). Trichothecenes B are known as a virulence factor of Fusarium spp. against wheat [76,77]. However, DON is more poisonous in Planta while NIV is more poisonous in Animalia [7]. The toxic effect of trichothecenes is the inhibition of protein synthesis in eukaryote by binding with 60S ribosomes [73,74,75].
4.2. Zearalenone
Zearalenone is also one of the dominant Fusarium mycotoxins on wheat worldwide; it is normally found in the same climate regions as trichothecenes [78,79,80,81]. Zearalenone derivatives, mainly, zearalanone, α- and β-zearalenol, and α- and β-zearalanol could be naturally produced by Fusarium spp. [7]. The main difference is the presence of ketone (=O) at C12 in zearalenone and zearalanone while it is hydroxyl (-OH) in α- and β- derivatives [82]. Zearalenone is of low acute toxicity either in Planta or in Animalia compared with trichothecenes [7,83]. Fusaria involved in zearalenone production are F. crookwellense, F. culmorum, F. equiseti, F. graminearum, F. semitectum, and F. sporotrichioides (Table 1). In Animalia, zearalenone has an estrogenic effect by binding to estrogen receptors which affect the sexual activities of animals [84].
4.3. Fusaric Acid
Fusaric acid is one of the first identified Fusarium mycotoxins; it is produced by a wide range of Fusarium species, and interestingly, by both trichothecene and fumonisin producers [63]. It is considered as a virulence factor for different Fusaria. Fusaric acid has virulent toxicity in Planta; however, its toxicity in Animalia is low to moderate. The toxic effects of fusaric acid include modifying the potential of the cell membrane and inhibiting ATP synthesis [85].
4.4. Fumonisins
Fumonisins were first identified in South Africa in 1988. They can be found mainly in maize products in regions with warm conditions. However, they can also be detected in other cereals especially wheat, barley and sorghum, and other plants such as soybean, asparagus, tea, and medicinal plants [86,87]. Fumonisins are polyketide hydrophilic mycotoxins and they contain a large number of derivatives. Therefore, they are classified in four main groups A, B, C, and P. Fumonisins B is the most widespread group and it contains FB1 which is of high concern regarding human and animal toxicity [7,87,88]. Exposure to fumonisins causes esophageal cancer and embryonal neural-tube defects in humans, leuko-encephalomalacia in equine and pulmonary edema in pigs [87,88,89]. A large number of Fusarium species are involved in fumonisin production; however, F. verticillioides and F. proliferatum are the main producers of these toxins [88].
4.5. Emerging Toxins
The emerging toxins of Fusarium are enniatins, beauvericin, fusaproliferin, and moniliformin [61]. The presence of emerging toxins is accompanied by traditional Fusarium toxins in cereals, particularly maize, wheat, barley, and oat, worldwide [62,67,71,90,91].
4.5.1. Enniatins and Beauvericin
Enniatins and beauvericin are cyclohexadepsipeptides with a lipophilic nature [61]. The main chemical derivatives of enniatins that can be detected in cereals are ENA, ENA1, ENB, and ENB1 [92]. Besides their antibacterial, antifungal, insecticidal activities [92], they showed cytotoxic effect for different cell cultures in vitro; however, toxicity in vivo is limited to poultry especially in the liver [67,93]. In addition, they exhibited cytotoxicity against cancer cell lines suggesting them as pharmacological candidates to fight cancer [94]. Enniatins and beauvericin are produced by a wide spectrum of Fusarium species [67]; however, on wheat, they were recorded for F. acuminatum, F. avenaceum, F. equiseti, F. lateritium, F. poae, F. proliferatum, F. sambucinum, F. sporotrichioides, F. subglutinans, and F. tricinctum (Table 1).
4.5.2. Fusaproliferin
Fusaproliferin is a bicyclic sesterterpene, which was later discovered in 1993 from F. proliferatum isolate. It can be produced simultaneously with a deacetylated form in a 3:1 ratio [61]. Fusaproliferin shows toxicity on insect and mammalian cells in addition to poultry embryos [7]. On wheat, fusaproliferin production was reported for F. proliferatum and F. subglutinans (Table 1).
4.5.3. Moniliformin
Moniliformin is a small molecule with high polarity; it can be found in nature as a sodium or potassium salt. Moniliformin was first identified as a mycotoxin of F. moniliforme that was renamed F. verticillioides [61]. The toxic effect of moniliformin is by disrupting thiamine enzymes that affect cellular energy supply. This leads to acute heart failure, pulmonary and immunity disruption in animals [67]. On wheat, moniliformin accompanied FHB infected with F. acuminatum, F. avenaceum, F. oxysporum, F. proliferatum, F. sambucinum, F. subglutinans, F. tricinctum, and F. verticillioides (Table 1).
4.6. Masked Mycotoxins
The term “masked mycotoxin” indicates a mycotoxin biologically modified by a conjugation reaction from the plants as a detoxification mechanism. This was suggested to differentiate them from other types of biological modification of mycotoxins, e.g., by animals, fungi, and microbiota of animals and humans. In addition, to discriminate them from chemically modified mycotoxins (i.e., thermally and non-thermally) and matrix-associated mycotoxins [95]. The main concern regarding masked mycotoxins is that they are not detectable by traditional analysis, and they are hydrolyzed through digestion into their parental mycotoxins or even more or less toxic compounds [95]. Different masked mycotoxins were reported in cereal grains. DON-3-glucoside (DON-3-G), NIV-3-glucoside (NIV-3-G), 4-ANIV-glucoside (4-ANIV-G), T-2-3-glucoside (T-2-3-G), HT-2-3-glucoside (HT-2-3-G), and ZEA-14-glucoside (ZEA-14-G) were reported to occur in wheat [96]. In their original situation, the toxicity of masked trichothecenes and zearalenone was shown to be significantly lower than their free parental toxins. They also showed stability in the upper gastro-intestinal tract. However, the main danger of these toxins is that they are hydrolyzed by the intestinal microbiota after ingestion [97,98]. Therefore, these masked mycotoxins must be legislated and controlled in the food and feed chain [53].
5. Fusarium Head Blight Diagnosis on Wheat
There are different methods to diagnose the fungal pathogens involved in FHB on wheat. The classical method is pathogen re-isolation on selective media and identifying the fungus based on the morphological characteristics of the spores or the colony. The immunological method uses specific antibodies against a specific protein or protein complex produced by the fungus. However, the most specific method is the molecular method using specific primers that target a specific region in the DNA of the fungus.
5.1. Selective Media
Fusarium species can be diagnosed based on the visual and microscopical characteristics of the colony and the spores after re-isolating the fungus on selective medium. Different media showed selectivity to Fusarium spp., e.g., Czapek Dox iprodione dichloran agar (CZID), dichloran-chloramphenicol peptone agar (DCPA), malachite green agar (MGA 2.5), modified Czapek Dox agar (MCz), Nash and Snyder medium (NS), and potato dextrose iprodione dichloran agar (PDID). However, MGA 2.5 was recommended as a selective medium for Fusarium re-isolation from naturally infected kernels [99]. Furthermore, differentiation between Fusarium species was possible based on their pigmentation on CZID [100]. Recently, different media containing the bacterial toxin “toxoflavin” produced by the Burkholderia glumae showed selectivity to Fusarium species [101]. However, this method is laborious and time-consuming and it needs experts in fungal taxonomy to diagnose the disease at the species scale.
5.2. Immunological Method
Enzyme-linked immunosorbent assay (ELISA) is used as a diagnostic method for Fusarium using poly- or monoclonal antibodies. These antibodies are obtained after immunization of animals or cell lines by exoantigens secreted by Fusarium [102,103]. However, the main drawback of this method is that it is genus specific [104].
5.3. Molecular Method
The polymerase chain reaction (PCR) was invented in 1984 and became widely used in plant pathogen detection and quantification with high sensitivity and specificity [105,106]. The PCR costs reduced with the introduction of the DNA polymerase (Taq) of the high-temperature tolerant bacteria Thermus aquaticus in 1988. This allowed automated thermal cycling and abolished the need for enzyme refreshment after each cycle [105]. The PCR allows the detection of plant diseases before the symptoms become visible. Moreover, it differentiates between fungal species scale even when they have morphological similarities. Different genomic regions are used to design species-specific primers, e.g., internal transcribed spacers (ITS), intergenic spacer (IGS) regions, and protein-coding genes [107]. Different primers were developed to detect Fusarium species involved in FHB (Table 2). However, primers targeting F. lateritium and F. semitectum showed cross-hybridization with other Fusarium species [108].
Table 2.
Forward and reverse primers sequences used to amplify specific fragments of fungal DNA of Fusarium species.
Table 2.
Forward and reverse primers sequences used to amplify specific fragments of fungal DNA of Fusarium species.
| Pathogen | Primer Name | Primer Sequence (5′-3′) | Amplified Fragment | Reference |
|---|---|---|---|---|
| F. acuminatum | FAC-F FAC-R | GGGATATCGGGCCTCA GGGATATCGGCAAGATCG | 602 bp | [109] |
| F. avenaceum | Fave574 fwd Fave627 rev | TATGTTGTCACTGTCTCACACCACC AGAGGGATGTTAGCATGATGAAG | EF1α gene | [110] |
| F. crookwellense synonym F. cerealis | CRO-A F CRO-A R | CTCAGTGTCCACCGCGTTGCGTAG CTCAGTGTCCCAATCAAATAGTCC | 842 bp | [111] |
| F. culmorum | OPT18 F OPT18 R | GATGCCAGACCAAGACGAAG GATGCCAGACGCACTAAGAT | 472 bp | [112] |
| F. equiseti | Feq-F Feq-R | GGCCTGCCGATGCGTC CGATACTGAAACCGACCTC | 990 bp | [113] |
| F. graminearum | Fg16N F Fg16N R | ACAGATGACAAGATTCAGGCACA TTCTTTGACATCTGTTCAACCCA | 280 bp | [114] |
| F. lateritium | ||||
| F. oxysporum | FOF1 FOR1 | ACATACCACTTGTTGCCTCG CGCCAATCAATTTGAGGAACG | 340 bp | [115] |
| F. poae | Fp82 F Fp82 R | CAAGCAAACAGGCTCTTCACC TGTTCCACCTCAGTGACAGGTT | 220 bp | [116] |
| F. proliferatum | Fp3-F Fp4-R | CGGCCACCAGAGGATGTG CAACACGAATCGCTTCCTGAC | 230 bp | [117] |
| F. sambucinum | FSF1 FSR1 | ACATACCTTTATGTTGCCTCG GGAGTGTCAGACGACAGCT | 315 bp | [115] |
| F. semitectum | ||||
| F. sporotrichioides | Fspor F1 Lanspo R1 | CGCACAACGCAAACTCATC TACAAGAAGACGTGGCGATAT | 332 bp | [118] |
| F. subglutinans | 61-2F 61-2R | GGCCACTCAAGAGGCGAAAG GTCAGACCAGAGCAATGGGC | 445 bp | [119] |
| F. tricinctum | Ftri573 fwd Ftri630 rev | TTGGTATGTTGTCACTGTCTCACACTAT TGACAGAGATGTTAGCATGATGCA | EF1α gene | [110] |
| F. verticillioides synonym F. moniliforme | VERT1 VERT2 | GTCAGAATCCATGCCAGAACG CACCCGCAGCAATCCATCAG | 800 bp | [120] |
| Microdochium nivale | Y13N F Y13N R | ACCAGCCGATTTGTGGTTATG GGTCACGAGGCAGAGTTCG | 300 bp | [121] |
| Microdochium majus | Y13M F Y13M R | CTTGAGGCGGAAGATCGC ATCCCTTTTCCGGGGTTG | 220 bp | [121] |
6. Integrated Management of Fusarium Head Blight
The effective management of FHB is challenging due to several factors. Firstly, maize intensification and reduced tillage increased the frequency of FHB epidemics during the last decades. This is because maize is the main host of Fusarium species, which serves as a source of the inoculum, and reduced tillage helps to keep this source available during wheat vegetation. In addition, wheat comes very often after maize in the crop rotation, which increases the disease incidence during the availability of the inoculum. Secondly, the visible FHB symptoms appear on wheat spikes at a later stage of pathogenicity, and during this stage, it is too late for fungicide application because the kernels have been contaminated with Fusarium mycotoxins. In addition, traditional disease control using fungicides involves different disadvantages mainly costs, bio- and eco-hazards, relatively short lifetime due to fungicide resistance, and low availability for smallholder farmers. Moreover, the environment and health protection measures lead to continuous regulatory changes regarding the availability and applicability of fungicides [122]. This shows the need for an integrated management strategy that incorporates cultural practices, resistant varieties, and bio- and chemical measures to control the disease.
6.1. Cultural Practices
Adopting moderately resistant varieties combined with variety rotation and using varieties with different maturities in addition to spreading anthesis times by disseminating planting dates showed efficiency in FHB control [123,124]. In addition, the reduction of inoculum pressure during wheat anthesis can play a significant role in disease management. This can be achieved by plowing the soil and burying the residues of the previous crop especially if this crop is one of the main hosts of Fusarium species, such as maize and barley. This practice prohibits perithecia formation and ascospore discharge during wheat spike development [21,125]. Another practice to reduce inoculum pressure is avoiding FHB cultural hosts, e.g., maize as a previous crop in wheat fields [126].
6.2. Host Plant Resistance to Fusarium Head Blight
Components of wheat resistance to FHB include passive resistance represented by morphological and phenological features and active resistance represented by physiological features [127]. Morphological and phenological features that are involved in passive resistance are plant height, wheat awns, narrow and short floral opening, and the time of retained anthers. Plant height: tallness helps wheat spikes to stand away from splashed rain droplets that carry the inoculum from the soil surface and crop residues. Wheat awns: awns trap the inoculum and increase natural infection while their absence reduces it [127]. A narrow and short floral opening reduces the floret’s exposure to the inoculum and increases resistance while retained anthers and pollen might trap the inoculum and catalyze spore germination and fungal penetration [128]. Active resistance can be classified into the following types: resistance to initial penetration or infection (Type I resistance), resistance to fungal spread within the spike from the infected spikelet (Type II resistance) [129], resistance to kernel infection and tolerance against FHB (Types III and IV, respectively) [130], and resistance to trichothecenes (Type V) [73].
Wheat resistance to FHB is a quantitative trait, which means that many genes with cumulative effects are involved in this trait. Environmental conditions have a significant effect on this trait resulting in various resistance levels in different environments [128]. Durum wheat is known to be highly susceptible to FHB due to the scarcity of resistance sources in the tetraploid gene pool [30]. To date, seven quantitative trait loci (QTLs) were officially given gene names, most of them from Chinese hexaploid wheat (Table 3) [131]. However, the direct introgression of these sources in breeding programs is still difficult due to undesirable agronomic traits [131]. Steiner et al. [128] suggested integrating genomic selection based on genome-wide prediction models with marker-assisted selection for QTL and classical phenotypic selection based on visible symptoms in breeding programs for FHB resistance.
Table 3.
QTLs involved in wheat resistance to Fusarium head blight.
| QTL | Location | Source | Resistance Type | Reference |
|---|---|---|---|---|
| Fhb1 | 3BS | Sumai 3 and Nyubai | Type II | [132] |
| Fhb2 | 6BS | Sumai 3 | Type II | [133] |
| Fhb3 | 7AS | Leymus racemosus | Type II | [134] |
| Fhb4 | 4BL | Wangshuibai | Type I | [135] |
| Fhb5 | 5AS | Wangshuibai and Sumai 3 | Type I | [136] |
| Fhb6 | 1AS | Elymus tsukushiensis | Type II | [137] |
| Fhb7 | 7D | Thinopyrum ponticum | Type II | [138] |
6.3. Biological Control
Biological control uses microorganisms antagonistic to Fusarium species or biological secondary metabolites to control FHB on wheat. These microorganisms can be applied to the residues of the previous crop to inhibit perithecia development or directly to wheat spikes. For example, bacteria from Bacillus spp., Lysobacter enzymogenes, Pseudomonas spp. and Streptomyces spp., and fungi from Aureobasidium pullulans, Clonostachys rosea, and Trichoderma spp. showed effectivity against Fusarium [139]. The fungus Clonostachys rosea was applied on wheat residues infected with different Fusarium species under field conditions. Fusarium growth measured as fungal DNA reduced between 68 and 98% after 90 days of treatment and was undetectable after 180 days [140]. Comby et al. [141] reported three new fungal species, namely Aureobasidium proteae, Phoma glomerate, and Sarocladium kiliense, with a high protection ratio between 75 and 100% on detached wheat spikelets. The basidiomycetous yeast Cryptococcus nodaensis OH 182.9 was isolated from wheat anthers [142]. This isolate presented reduced disease severity of between 45 and 60% under controlled and field conditions [143,144]. Zhang et al. [145] isolated 113 endophytes from roots, stems, leaves, and spikelets of wheat and tested their antagonistic effect against F. graminearum on detached wheat spikes. Six isolates were shown to inhibit F. graminearum growth while the strain XS-2 of Bacillus amyloliquefaciens reduced disease severity on detached wheat spikes significantly. In vitro, B. subtilis SG6 inhibited F. graminearum growth, sporulation, and DON concentration with ratios of 88, 96, and 100%, respectively, while in the field, the same strain significantly reduced disease incidence, FHB index, and kernel DON contamination when it was applied from anthesis until soft dough [146]. The usage of biochemical compounds proved to be effective in FHB control. Chitosan (the deacetylated derivative of chitin) inhibited the fungal growth and DON contamination in irradiated wheat kernels [147]. It also reduced disease severity and DON contamination by ≥74% under greenhouse and field conditions [148]. Drakopoulos et al. [149] tested botanical aqueous extracts of white mustard (Sinapis alba) and Chinese galls (Rhus chinensis) against F. graminearum in vitro. All these compounds fully inhibited mycelium growth, conidial, and ascospore germination. Moreover, they reduced perithecia formation and ascospore discharge up to 50 and 6%, respectively.
6.4. Chemical Control
Effective chemical control of FHB should be combined with other management practices [125,150]. The critical time for fungicide application is the susceptible stage, i.e., anthesis stage and 10 days after anthesis. However, a limited application period, anthesis heterogenicity, and weather conditions at this stage might be challenging for effective fungicide application; this may require multiple applications to achieve efficient disease control [139,151]. Demethylation inhibitor (DMI) fungicides, namely metconazole, prothioconazole, tebuconazole, prothioconazole + tebuconazole were shown to be more effective than propiconazole [6,152]. Another factor that affects fungicide efficiency is spike coverage during application, which is affected by the nozzle type, and spray angle. Lehoczki-Krsjaket al. [153] showed that two sideward-spraying (90 and 120° for forward and backward streams, respectively) increased fungicide content between 1.08 and 1.43 times in wheat spikes compared with vertical spraying. Moreover, increasing spike coverage from 19 to 37% reduced FHB incidence and DON content significantly for all tested fungicides [154].
6.5. Predicting and Detecting Disease Incidence
One of the prediction practices is risk assessment using disease prediction models based on weather conditions and the history of FHB epidemics in the growing region [6]. In addition, the effective monitoring of disease incidence in the field helps in early disease detection and supports the decision-making strategy to apply fungicides at a suitable time. Experts and prognosis models are based on information about the dominant Fusarium species, inoculum availability, resistance degree of the cultivated wheat variety, anthesis period, and the previous crops in the surrounding area. In addition, information about favorable weather conditions (e.g., rainfall, temperature, and relative humidity) for FHB incidence during wheat vegetation are required as input data for these models. These two practices could be effective tools to prevent quantity and quality losses in wheat yield caused by FHB. These models can be supported or improved by innovative digital technology and remote sensing data to realize knowledge-based plant protection management.
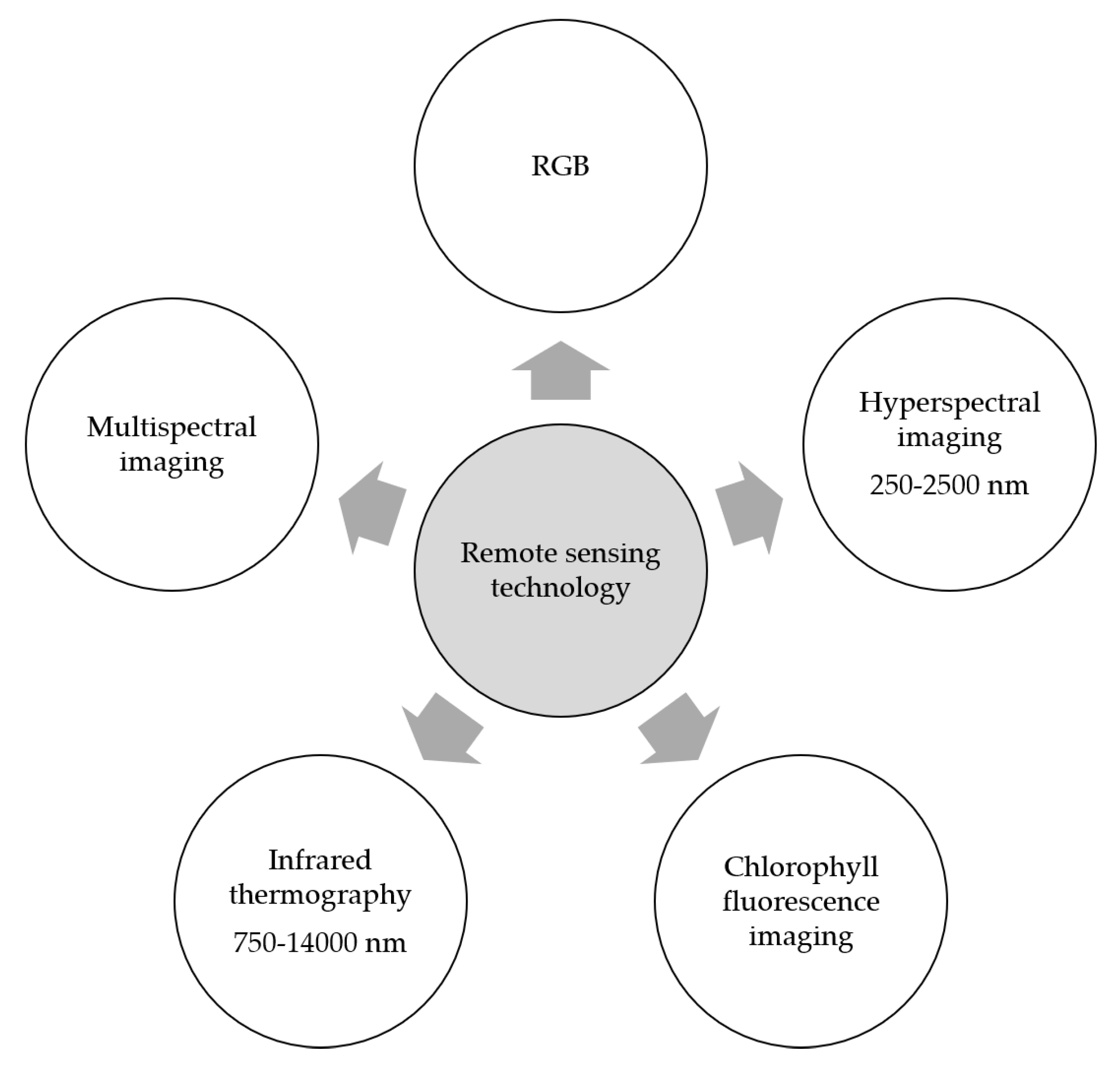
7. Remote Sensing for Monitoring and Phenotyping Fusarium Head Blight
Optical sensors are among the remote sensing technologies that have been widely investigated in monitoring plant diseases as well as in plant phenotyping (Figure 4). These sensors include RGB imaging (red, green, and blue bands), multi- and hyperspectral imaging in the visible–near infrared range and the shortwave infrared range, infrared thermography in the spectral range 7500–14,000 nm and chlorophyll fluorescence imaging [13].
7.1. Spectral Techniques
Spectral sensors acquire the spectral reflectance of the object. Based on the number of recorded wavebands, these sensors are classified as multispectral sensors and hyperspectral sensors. Multispectral sensors record the spectral reflectance of individual wavebands (e.g., RGB wavebands or specific wavebands in the NIR range) [13], while hyperspectral sensors record the spectral reflectance over a wide number of wavebands in the electromagnetic spectrum from 250 to 2500 nm. This information is correlated to the plant pigments, chemical compounds, and the water content of the plant [11].
The main flaw of using hyperspectral imaging sensors in plant-disease detection is data complexity. To reduce data complexity, spectral vegetation indices (SVIs) can be derived from the spectral data based on a ratio between individual wavebands. Each of these indices can be used as an indicator of a specific compound of the plant which might be affected during the pathogenicity (e.g., chlorophyll, water content, or tissue structure). This data can be utilized in plant-disease detection using supervised or unsupervised machine learning methods. Reducing data complexity reduces calculation time and improves the accuracy of the machine learning approach in plant-disease detection [155].
Bauriegel et al. [156] investigated the feasibility of HSI in the VIS and NIR ranges for the early detection of FHB using data from controlled and field conditions. They showed that the best time for disease detection is at the beginning of the medium milk stage GS 71–85 according to the Lancashire scale [157]. Principal component analysis (PCA) was used to disclose the relevant wavelength of healthy and diseased wheat tissue. Based on this approach, the healthy and diseased areas of wheat spikes were correctly classified with an accuracy of 100 and 94%, respectively. A spectral angle mapper (SAM) was also able to classify the diseased area with an accuracy of 87%. However, the main drawback of SAM is that it is time-consuming. Another study showed the superiority of HSI in FHB detection under controlled conditions compared with the field conditions at the growth stage GS 71–73 [158]. Alisaac et al. [14] used SVM to discriminate healthy and Fusarium-infected wheat spikes based on the mean spectral signature and the SVIs derived from the mean spectral signature of the spikes. They reached an accuracy of >93% in the period 8–17 days after inoculation. In addition, it was possible to rank wheat varieties automatically according to their resistance to FHB using a non-metric multidimensional scaling approach based on the SVIs of the spikes [14]. Moreover, HSI showed promising results as a fast, non-invasive, and non-destructive method for pre-screening Fusarium infection and mycotoxin contamination on the kernel and flour scale. This can accelerate the kernel sorting procedure by replacing the laborious and cost-effective chemical methods [15,159].
Under field conditions, Ma et al. [160] used the spectral reflectance in the VIS, NIR, and SWIR ranges to detect FHB. Six feature bands correlated to FHB were extracted by continuous wavelet analysis (CWA). Afterward, these feature bands were utilized to establish a discrimination model using Fisher linear discriminant analysis (FLDA). They revealed an accuracy of 89% using this model. Jin et al. [161] used HSI in the VIS and NIR ranges to detect FHB on wheat under field conditions. They reached an accuracy of 85% using the neural network approach (NN). Color imaging was also used as an input for the deep neural network (DNN) approach to detect FHB on spike scale in the field. The accuracy of the model reached 92% at the milk stage of wheat [162]. Zhang et al. [163] integrated spectral and image data to detect FHB using the same approach with an accuracy of R2 = 0.97.
Unmanned aerial vehicles (UAVs) are an effective and flexible tool for acquiring high resolution images of a large acreage in a short time and with low costs. Zhang et al. [164] used UAV hyperspectral images for a quantitative detection of FHB in the field. They classified the field infection into mild, moderate and severe infection by fusing the spectral and image features acquired by UAV. FHB-monitoring models with accuracies of 98% and R2 = 0.88 were also developed based on UAV hyperspectral images to detect FHB under field conditions [165,166].
7.2. Infrared Thermography
Infrared thermography determines the plant temperature which reflects the water status of the plant. Plant pathogens influence the water balance in the plant tissue and this effect can be indirectly measured and visualized as a false-color image by IRT [13]. Maximum temperature difference (MTD) and average temperature difference (∆T) are parameters derived from IRT and can be successfully used in plant-disease detection. MTD represents the differences between the maximum and the minimum temperature within the object, while ∆T represents the difference between the average temperature of the ambient air and the average temperature of the object [167]. These parameters were implemented successfully using the support vector machine (SVM) approach to detect FHB on the spikelet scale [16].
7.3. Chlorophyll Fluorescence Imaging
Chlorophyll fluorescence imaging assesses the status of photosystem II (PSII) of the plant [168]. The basic fluorescence (F0) is the minimum value of fluorescence for dark-adapted PSII after excitation with low-intensity light but not enough for electron transport through PSII. The maximum fluorescence (Fm) is the maximum value of fluorescence for dark-adapted PSII after excitation with a saturating pulse. The variable fluorescence (Fv) represents the difference between Fm and F0, while the ratio Fv/Fm represents the maximum quantum yield of PSII photochemistry with a constant value of ≈0.83 for healthy plants [169]. FHB infection causes a significant reduction in the photosynthetic activity of wheat spikes; this reduction can be detected by CFI [16,170,171].
8. Future Perspectives
The damage caused by FHB includes yield losses and contamination of wheat kernels with mycotoxins. Therefore, applying fungicides at an early stage of pathogenicity should be considered in the management strategy of the disease [21]. This, in turn, needs an accurate control of disease incidence under field conditions. In addition, the selective harvest of healthy spikes could be an option to avoid infected spikes and reduces mycotoxin contamination [171]. This highlights the need for effective tools for real-time detection and identification of plant diseases in the field.
Former studies successfully implemented HSI and CFI to detect and discriminate healthy and FHB-infected spikes [156,170]. Mahlein et al. [16] showed the applicability of optical sensors, i.e., IRT, CFI, and HSI to detect FHB on the spikelet scale as early as three days after inoculation using a machine learning approach. In these terms, optical sensors are promising tool for future applications to support the decision-making strategy with FHB incidence to apply fungicides against FHB at an early stage of pathogenicity.
Based on the SVIs of wheat spikes, it was also possible to discriminate between healthy and FHB-infected spikes as well as spikes infected with different Fusarium species [14]. Using SVIs in FHB detection reduces the required data and time for FHB detection. This shows the feasibility of using multispectral instead of hyperspectral sensors for future real-time detection and identification of FHB under field conditions. This, in turn, gives the possibility of local treatment of infected spots in the field and reduces the quantities of plant protection chemicals. In addition, selective harvest aided by spectral sensors helps to avoid the infected spikes during harvest and reduces mycotoxin contamination of the harvested wheat kernels.
High yielding varieties with sufficient resistance to FHB are required to reduce yield losses and the use of plant protection chemicals substantially. Besides yield and disease resistance, different plant traits (e.g., plant height, lodging) have to be assessed simultaneously. from this point of view, using optical sensors, especially multi- and hyperspectral imaging sensors in phenotyping, can provide an objective assessment for plant traits in addition to reducing time and labor in the field [11].
Alisaac et al. [14] designed an automated model to rank wheat varieties according to their resistance to FHB based on multiple hyperspectral assessments of disease development during the pathogenicity [14,16]. This model needs to be optimized for future application under field condition.
Another factor to be considered in the breeding programs is kernel phenotyping for infection and mycotoxin contamination. This needs the quantification of Fusarium DNA and mycotoxin content in wheat kernels. However, the quantification methods are destructive, time-consuming, laborious, and expensive [172]. HSI proved the feasibility of detecting wheat kernels and flour contaminated with multiple levels of DON and fungal DNA of different Fusarium species [15]. Applying this tool allows the screening of a large number of wheat entries within a short time according to their DON and Fusarium DNA contents. Moreover, this technology will reduce the costs, labor, and time required for wheat kernel phenotyping against FHB.
9. Conclusions
In summary, combining different control measures has been shown to be an effective tool in the integrated disease management of FHB. These control measures must be considered before and after planting wheat. Before planting, applying suitable cultural practices and planting resistant varieties play a significant role in reducing disease incidence and severity. However, predicting and monitoring the disease will help in the decision making to apply biological and chemical control products during the growing season. In addition, selective harvesting by avoiding mycotoxin-contaminated plots is a useful tool to reduce mycotoxin contamination in harvested kernels. From this point of view, optical sensors, mainly IRT and HSI, are promising tools to monitor FHB infection on wheat.
Author Contributions
Conceptualization, E.A. and A.-K.M.; writing—original draft preparation, E.A.; writing—review and editing, E.A. and A.-K.M.; visualization, E.A.; supervision, A.-K.M. All authors have read and agreed to the published version of the manuscript.
Funding
E.A. was funded by the German Federal Ministry of Education and Research (BMBF) within the funding program “Networks of excellence in agricultural and nutrition research—CROP.SENSe.net” (Funding Code: 0315529), Junior Research Group “Hyperspectral phenotyping of resistance reactions of barley” and the Catholic Academic Exchange Service (KAAD), Scholarship Program 2. A.-K.M. was partly funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under “Germany’s Excellence Strategy—EXC 2070–390732324”. The authors are thankful to Petr Karlovsky for helpful discussions and support.
Institutional Review Board Statement
The manuscript has been fully approved in the Publication Approval workflow at the Julius Kühn-Institute.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- FAO Food and Agriculture Organization of the United Nations FAOStat. Available online: https://www.fao.org/faostat/en/#data (accessed on 22 November 2022).
- CABI Centre for Agriculture and Bioscience International Crop Protection Compendium. Available online: https://www.cabi.org/publishing-products/crop-protection-compendium/ (accessed on 22 November 2022).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current Status of the Genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium Ear Blight (Scab) in Small Grain Cereals—A Review. Ptant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; de Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; van Sanford, D. A Unified Effort to Fight an Enemy of Wheat and Barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Smith, W. New Diseases of Wheat, Barley, and Rye-Grass. In Diseases of Field and Garden Crops; MacMillan and Co.: London, UK, 1884; pp. 208–213. [Google Scholar]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of Wheat and Barley: A Re-Emerging Disease of Devastating Impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Brennan, J.; Monaghan, S.; Moretti, A.; Mule, G.; et al. Relationship between the Fungal Complex Causing Fusarium Head Blight of Wheat and Environmental Conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral Sensors and Imaging Technologies in Phytopathology: State of the Art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.T.; Tester, M. Phenomics—Technologies to Relieve the Phenotyping Bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.K. Plant Disease Detection by Imaging Sensors—Parallels and Specific Demands for Precision Agriculture and Plant Phenotyping. Plant Dis. 2016, 100, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Alisaac, E.; Behmann, J.; Kuska, M.T.; Dehne, H.W.; Mahlein, A.K. Hyperspectral Quantification of Wheat Resistance to Fusarium Head Blight: Comparison of Two Fusarium Species. Eur. J. Plant Pathol. 2018, 152, 869–884. [Google Scholar] [CrossRef]
- Alisaac, E.; Behmann, J.; Rathgeb, A.; Karlovsky, P.; Dehne, H.W.; Mahlein, A.K. Assessment of Fusarium Infection and Mycotoxin Contamination of Wheat Kernels and Flour Using Hyperspectral Imaging. Toxins 2019, 11, 556. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Alisaac, E.; al Masri, A.; Behmann, J.; Dehne, H.W.; Oerke, E.C. Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale. Sensors 2019, 19, 2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; Culley, J.; Fraser, K.; Hubbard, S.; Meloche, F.; Ouellet, T.; Seaman, W.L.; Seifert, K.A.; Turkington, K.; Voldeng, H. Effect of Tillage Practice on Fusarium Head Blight of Wheat. Can. J. Plant Pathol. 1998, 20, 95–103. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The Effect of Previous Crop Residues and Tillage on Fusarium Head Blight of Wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.A.; de Wolf, E.D.; Paul, P.A.; Madden, L.v. Functional Data Analysis of Weather Variables Linked to Fusarium Head Blight Epidemics in the United States. Phytopathology 2019, 109, 96–110. [Google Scholar] [CrossRef] [Green Version]
- Obanor, F.; Neate, S.; Simpfendorfer, S.; Sabburg, R.; Wilson, P.; Chakraborty, S. Fusarium graminearum and Fusarium pseudograminearum Caused the 2010 Head Blight Epidemics in Australia. Plant Pathol. 2013, 62, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.M.; Palacios, S.A.; Yerkovich, N.; Palazzini, J.M.; Battilani, P.; Leslie, J.F.; Logrieco, A.F.; Chulze, S.N. Fusarium Head Blight and Mycotoxins in Wheat: Prevention and Control Strategies across the Food Chain. World Mycotoxin J. 2019, 12, 333–355. [Google Scholar] [CrossRef]
- Johns, L.E.; Bebber, D.P.; Gurr, S.J.; Brown, A. Emerging Health Threat and Cost of Fusarium Mycotoxins in European Wheat. Nat. Food 2022, 3, 1014–1019. [Google Scholar] [CrossRef]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding Wheat for Resistance to Fusarium Head Blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Aboukhaddour, R.; Fetch, T.; McCallum, B.D.; Harding, M.W.; Beres, B.L.; Graf, R.J. Wheat Diseases on the Prairies: A Canadian Story. Plant Pathol. 2020, 69, 418–432. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.; Seifert, K.; Guo, W.; Blackwell, B.A.; Harris, L.J.; Overy, D.P. Prevalence of Fusarium Species Causing Head Blight of Spring Wheat, Barley and Oat in Ontario during 2001–2017. Can. J. Plant Pathol. 2019, 41, 392–402. [Google Scholar] [CrossRef]
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and Deoxynivalenol during the 2012/13 Wheat Fusarium Head Blight Outbreak in Argentina. Cereal. Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef] [Green Version]
- del Ponte, E.M.; Fernandes, J.M.C.; Pavan, W.; Baethgen, W.E. A Model-Based Assessment of the Impacts of Climate Variability on Fusarium Head Blight Seasonal Risk in Southern Brazil. J. Phytopathol. 2009, 157, 675–681. [Google Scholar] [CrossRef]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One Fungus, One Name: Defining the Genus Fusarium in a Scientifically Robust Way That Preserves Longstanding Use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MycoBank An Initiative of the International Mycological Association (IMA) and the Westerdijk Fungal Biodiversity Institute MycoBank Database. Available online: https://www.mycobank.org/ (accessed on 22 November 2022).
- Haile, J.K.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Clarke, J.M.; Kutcher, H.R.; Steiner, B.; Buerstmayr, H.; Pozniak, C.J. Fusarium Head Blight in Durum Wheat: Recent Status, Breeding Directions, and Future Research Prospects. Phytopathology 2019, 109, 1664–1675. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.v. The Prevalence and Impact of Fusarium Head Blight Pathogens and Mycotoxins on Malting Barley Quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Oerke, E.C.; Meier, A.; Dehne, H.W.; Sulyok, M.; Krska, R.; Steiner, U. Spatial Variability of Fusarium Head Blight Pathogens and Associated Mycotoxins in Wheat Crops. Plant Pathol. 2010, 59, 671–682. [Google Scholar] [CrossRef]
- Spanic, V.; Lemmens, M.; Drezner, G. Morphological and Molecular Identification of Fusarium Species Associated with Head Blight on Wheat in East Croatia. Eur. J. Plant Pathol. 2010, 128, 511–516. [Google Scholar] [CrossRef]
- Alkadri, D.; Nipoti, P.; Döll, K.; Karlovsky, P.; Prodi, A.; Pisi, A. Study of Fungal Colonization of Wheat Kernels in Syria with a Focus on Fusarium Species. Int. J. Mol. Sci. 2013, 14, 5938–5951. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. For Blighted Waves of Grain: Fusarium graminearum in the Postgenomics Era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Mourelos, C.A.; Malbrán, I.; Balatti, P.A.; Ghiringhelli, P.D.; Lori, G.A. Gramineous and Non-Gramineous Weed Species as Alternative Hosts of Fusarium graminearum, Causal Agent of Fusarium Head Blight of Wheat, in Argentina. Crop Prot. 2014, 65, 100–104. [Google Scholar] [CrossRef]
- Suproniene, S.; Kadziene, G.; Irzykowski, W.; Sneideris, D.; Ivanauskas, A.; Sakalauskas, S.; Serbiak, P.; Svegzda, P.; Auskalniene, O.; Jedryczka, M. Weed Species within Cereal Crop Rotations Can Serve as Alternative Hosts for Fusarium graminearum Causing Fusarium Head Blight of Wheat. Fungal. Ecol. 2019, 37, 30–37. [Google Scholar] [CrossRef]
- Dong, F.; Xu, J.; Zhang, X.; Wang, S.; Xing, Y.; Mokoena, M.P.; Olaniran, A.O.; Shi, J. Gramineous Weeds near Paddy Fields Are Alternative Hosts for the Fusarium graminearum Species Complex That Causes Fusarium Head Blight in Rice. Plant Pathol. 2020, 69, 433–441. [Google Scholar] [CrossRef]
- Kang, Z.; Buchenauer, H. Immunocytochemical Localization of Fusarium Toxins in Infected Wheat Spikes by Fusarium culmorum. Physiol. Mol. Plant Pathol. 1999, 55, 275–288. [Google Scholar] [CrossRef]
- Brown, N.A.; Urban, M.; van de Meene, A.M.L.; Hammond-Kosack, K.E. The Infection Biology of Fusarium graminearum: Defining the Pathways of Spikelet to Spikelet Colonisation in Wheat Ears. Fungal. Biol. 2010, 114, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Divon, H.H.; Bøe, L.; Tveit, M.M.N.; Klemsdal, S.S. Infection Pathways and Penetration Modes of Fusarium langsethiae. Eur. J. Plant Pathol. 2019, 154, 259–271. [Google Scholar] [CrossRef]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium Head Blight of Wheat: Pathogenesis and Control Strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal Agent of Foot and Root Rot and Head Blight on Wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Stenglein, S.A. Fusarium poae a Pathogen That Needs More Attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar]
- Stenglein, S.A.; Dinolfo, M.I.; Barros, G.; Bongiorno, F.; Chulze, S.N.; Moreno, M.V. Fusarium poae Pathogenicity and Mycotoxin Accumulation on Selected Wheat and Barley Genotypes at a Single Location in Argentina. Plant Dis. 2014, 98, 1733–1738. [Google Scholar] [CrossRef] [Green Version]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of Wheat Infection Timing on Fusarium Head Blight Causal Agents and Secondary Metabolites in Grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium Species and Mycotoxins Associated with Head Blight in Small-Grain Cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Marín, P.; Moretti, A.; Ritieni, A.; Jurado, M.; Vázquez, C.; González-Jaén, M.T. Phylogenetic Analyses and Toxigenic Profiles of Fusarium equiseti and Fusarium acuminatum Isolated from Cereals from Southern Europe. Food Microbiol. 2012, 31, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Beccari, G.; Colasante, V.; Tini, F.; Senatore, M.T.; Prodi, A.; Sulyok, M.; Covarelli, L. Causal Agents of Fusarium Head Blight of Durum Wheat (Triticum durum Desf.) in Central Italy and Their in vitro Biosynthesis of Secondary Metabolites. Food Microbiol. 2018, 70, 17–27. [Google Scholar] [CrossRef]
- Glenn, A.E. Mycotoxigenic Fusarium Species in Animal Feed. Anim. Feed Sci. Technol. 2007, 137, 213–240. [Google Scholar] [CrossRef]
- Chandler, E.A.; Simpson, D.R.; Thomsett, M.A.; Nicholson, P. Development of PCR Assays to Tri7 and Tri13 Trichothecene Biosynthetic Genes, and Characterisation of Chemotypes of Fusarium graminearum, Fusarium culmorum and Fusarium cerealis. Physiol. Mol. Plant Pathol. 2003, 62, 355–367. [Google Scholar] [CrossRef]
- Vesonder, R.F.; Golifiski, P.; Plattner, R.; Zietkiewicz, D.L. Mycotoxin Formation by Different Geographic Isolates of Fusarium crookwellense. Mycopathologia 1991, 113, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Alisaac, E.; Rathgeb, A.; Karlovsky, P.; Mahlein, A.K. Fusarium Head Blight: Effect of Infection Timing on Spread of Fusarium graminearum and Spatial Distribution of Deoxynivalenol within Wheat Spikes. Microorganisms 2021, 9, 79. [Google Scholar] [CrossRef]
- Basler, R. Diversity of Fusarium Species Isolated from UK Forage Maize and the Population Structure of F. graminearum from Maize and Wheat. PeerJ 2016, 4, e2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European Database of Fusarium graminearum and F. culmorum Trichothecene Genotypes. Front Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef] [Green Version]
- Siou, D.; Gélisse, S.; Laval, V.; Elbelt, S.; Repinçay, C.; Bourdat-Deschamps, M.; Suffert, F.; Lannou, C. Interactions between Head Blight Pathogens: Consequences for Disease Development and Toxin Production in Wheat Spikes. Appl. Environ. Microbiol. 2015, 81, 957–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Meca, G.; Juan, C.; Mañes, J. Biosynthesis of Beauvericin and Enniatins Invitro by Wheat Fusarium Species and Natural Grain Contamination in an Area of Central Italy. Food Microbiol. 2015, 46, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; Mccormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New Tricks of an Old Enemy: Isolates of Fusarium graminearum Produce a Type A Trichothecene Mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Crippin, T.; Renaud, J.B.; Sumarah, M.W.; Miller, J.D. Comparing Genotype and Chemotype of Fusarium graminearum from Cereals in Ontario, Canada. PLoS ONE 2019, 14, e0216735. [Google Scholar] [CrossRef]
- Lofgren, L.; Riddle, J.; Dong, Y.; Kuhnem, P.R.; Cummings, J.A.; del Ponte, E.M.; Bergstrom, G.C.; Kistler, H.C. A High Proportion of NX-2 Genotype Strains Are Found among Fusarium graminearum Isolates from Northeastern New York State. Eur. J. Plant Pathol. 2018, 150, 791–796. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium-Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin—A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Miere, D.; Loghin, F.; Mañes, J. Presence of Enniatins and Beauvericin in Romanian Wheat Samples: From Raw Material to Products for Direct Human Consumption. Toxins 2017, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; de Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic Potentials of Fusarium Species in Various Culture Matrices Revealed by Mycotoxin Profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium Species, Chemotype Characterisation and Trichothecene Contamination of Durum and Soft Wheat in an Area of Central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Guo, Z.; DolÏl, K.; Dastjerdi, R.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Effect of Fungal Colonization of Wheat Grains with Fusarium spp. on Food Choice, Weight Gain and Mortality of Meal Beetle Larvae (Tenebrio molitor). PLoS ONE 2014, 9, e100112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Dissemination of Fusarium proliferatum by Mealworm Beetle Tenebrio molitor. PLoS ONE 2018, 13, e0204602. [Google Scholar] [CrossRef] [PubMed]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Barros, G.; Zanon, M.S.A.; Palazzini, J.M.; Haidukowski, M.; Pascale, M.; Chulze, S. Trichothecenes and Zearalenone Production by Fusarium equiseti and Fusarium semitectum Species Isolated from Argentinean Soybean. Food Addit. Contam.—Part A 2012, 29, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Ezekiel, C.N.; Odebode, A.C.; Fapohunda, S.O. Zearalenone Production by Naturally Occurring Fusarium Species on Maize, Wheat and Soybeans from Nigeria. J. Biol. Environ. Sci. 2008, 2, 77–82. [Google Scholar]
- Nazari, L.; Pattori, E.; Terzi, V.; Morcia, C.; Rossi, V. Influence of Temperature on Infection, Growth, and Mycotoxin Production by Fusarium langsethiae and F. sporotrichioides in Durum Wheat. Food Microbiol. 2014, 39, 19–26. [Google Scholar] [CrossRef]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin in Serbian Maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef] [Green Version]
- Mylona, K.; Garcia-Cela, E.; Sulyok, M.; Medina, A.; Magan, N. Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species in vitro and in Stored Cereals. Toxins 2019, 11, 495. [Google Scholar] [CrossRef] [Green Version]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal Grains—An Update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in Planta Mycotoxin Accumulation among Members of the Fusarium graminearum Species Complex on Wheat and Rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Spanic, V.; Zdunic, Z.; Drezner, G.; Sarkanj, B. The Pressure of Fusarium Disease and Its Relation with Mycotoxins in the Wheat Grain and Malt. Toxins 2019, 11, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.R. An Eight-Year Survey of Wheat Shows Distinctive Effects of Cropping Factors on Different Fusarium Species and Associated Mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Solarska, E.; Muszyńska, M. Deoxynivalenol and Zearalenone Occurence in Beers Analysed by an Enzyme-Linked Immunosorbent Assay Method. Food Control 2013, 29, 22–24. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Bemvenuti, R.H.; Zorzete, P.; de Souza Garcia, F.; Corrêa, B. Fungal Diversity and Natural Occurrence of Deoxynivalenol and Zearalenone in Freshly Harvested Wheat Grains from Brazil. Food Chem. 2016, 196, 445–450. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Berrada, H.; Miere, D.; Loghin, F.; Mañes, J. Study on Trichothecene and Zearalenone Presence in Romanian Wheat Relative to Weather Conditions. Toxins 2019, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yang, C.; Mao, Y.; Hong, X.; Du, D. Zearalenone Contamination in Corn, Corn Products, and Swine Feed in China in 2016–2018 as Assessed by Magnetic Bead Immunoassay. Toxins 2019, 11, 451. [Google Scholar] [CrossRef] [Green Version]
- McLean, M. The Phytotoxicity of Selected Mycotoxins on Mature, Germinating Zea mays Embryos. Mycopathologia 1995, 132, 173–183. [Google Scholar] [CrossRef]
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium Molds and Mycotoxins: Potential Species-Specific Effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- López-Díaz, C.; Rahjoo, V.; Sulyok, M.; Ghionna, V.; Martín-Vicente, A.; Capilla, J.; di Pietro, A.; López-Berges, M.S. Fusaric Acid Contributes to Virulence of Fusarium oxysporum on Plant and Mammalian Hosts. Mol. Plant Pathol. 2018, 19, 440–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Asta, C.; Battilani, P. Fumonisins and Their Modified Forms, a Matter of Concern in Future Scenario? World Mycotoxin J. 2016, 9, 727–739. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.S.; Wink, M. Exposure, Occurrence, and Chemistry of Fumonisins and Their Cryptic Derivatives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 769–791. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.M. Recent Research on Fumonisins: A Review. Food Addit. Contam.—Part A 2012, 29, 242–248. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-Occurrence of Regulated, Masked and Emerging Mycotoxins and Secondary Metabolites in Finished Feed and Maize–an Extensive Survey. Toxins 2016, 8, 863. [Google Scholar] [CrossRef] [Green Version]
- Stanciu, O.; Juan, C.; Miere, D.; Loghin, F.; Mañes, J. Occurrence and Co-Occurrence of Fusarium Mycotoxins in Wheat Grains and Wheat Flour from Romania. Food Control 2017, 73, 147–155. [Google Scholar] [CrossRef]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef]
- Ivanova, L.; Fæste, C.K.; van Pamel, E.; Daeseleire, E.; Callebaut, A.; Uhlig, S. Presence of Enniatin B and Its Hepatic Metabolites in Plasma and Liver Samples from Broilers and Eggs from Laying Hens. World Mycotoxin J. 2014, 7, 167–175. [Google Scholar] [CrossRef]
- Tedjiotsop Feudjio, F.; Dornetshuber, R.; Lemmens, M.; Hoffmann, O.; Lemmens-Gruber, R.; Berger, W. Beauvericin and Enniatin: Emerging Toxins and/or Remedies? World Mycotoxin J. 2010, 3, 415–430. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a Comprehensive Definition of Modified and Other Forms of Mycotoxins Including “Masked” Mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked Mycotoxins: A Review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W. Do Plant-Bound Masked Mycotoxins Contribute to Toxicity? Toxins 2017, 9, 85. [Google Scholar] [CrossRef]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A Systematic Review of Plant-Conjugated Masked Mycotoxins: Occurrence, Toxicology, and Metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef]
- Bragulat, M.R.; Martínez, E.; Castellá, G.; Cabañes, F.J. Selective Efficacy of Culture Media Recommended for Isolation and Enumeration of Fusarium spp. J. Food Prot. 2004, 67, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Thrane, U. Comparison of Three Selective Media for Detecting Fusarium Species in Foods: A Collaborative Study. Int. J. Food Microbiol. 1996, 29, 149–156. [Google Scholar] [CrossRef]
- Jung, B.; Lee, S.; Ha, J.; Park, J.C.; Han, S.S.; Hwang, I.; Lee, Y.W.; Lee, J. Development of a Selective Medium for the Fungal Pathogen Fusarium graminearum Using Toxoflavin Produced by the Bacterial Pathogen Burkholderia glumae. Plant Pathol. J. (Faisalabad) 2013, 29, 446–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, N.S.; Schwarz, P.; Dahleen, L.S.; Neate, S.M.; Horsley, R.; Glenn, A.E.; O’Donnell, K. ELISA Analysis for Fusarium in Barley: Development of Methodology and Field Assessment. Crop Sci. 2006, 46, 2636–2642. [Google Scholar] [CrossRef]
- Gan, Z.; Marquardt, R.R.; Abramson, D.; Clear, R.M. The Characterization of Chicken Antibodies Raised against Fusarium spp. by Enzyme-Linked Immunosorbent Assay and Immunoblotting. Int. J. Food Microbiol. 1997, 38, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.; Farnleitner, A.; Mach, R.L. Novel Methods for the Quantification of Pathogenic Fungi in Crop Plants: Quantitative PCR and ELISA Accurately Determine Fusarium Biomass. In Plant Pathology; Cumagun, C.J.R., Ed.; IntechOpen Limited: London, UK, 2012; pp. 203–218. [Google Scholar]
- Henson, J.M.; French, R. The Polymerase Chain Reaction and Plant Disease Diagnosis. Annu. Rev. Phytopathol. 1993, 31, 81–109. [Google Scholar] [CrossRef]
- Schaad, N.W.; Frederick, R.D. Real-Time PCR and Its Application for Rapid Plant Disease Diagnostics. Can. J. Plant Pathol. 2002, 24, 250–258. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Kot, A.; Szczerba, H.; Nowak, M.; Muszyńska, M. A Review of Conventional PCR Assays for the Detection of Selected Phytopathogens of Wheat. J. Mol. Microbiol. Biotechnol. 2017, 27, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Kang, M.R.; Cho, E.J.; Kim, H.K.; Yun, S.H. Specific PCR Detection of Four Quarantine Fusarium Species in Korea. Plant Pathol. J. (Faisalabad) 2010, 26, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.J.; Dennis, J.I.; Smyl, C.; Wallwork, H. The Application of Species-Specific Assays Based on the Polymerase Chain Reaction to Analyse Fusarium Crown Rot of Durum Wheat. Australas. Plant Pathol. 2002, 31, 119–127. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-Time PCR for Quantification of Eleven Individual Fusarium Species in Cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- Yoder, W.T.; Christianson, L.M. Species-Specific Primers Resolve Members of Fusarium Section Fusarium. Taxonomic Status of the Edible “Quorn” Fungus Reevaluated. Fungal Genet. Biol. 1998, 23, 68–80. [Google Scholar] [CrossRef]
- Schilling, A.G.; Möller, E.M.; Geiger, H.H. Polymerase Chain Reaction-Based Assays for Species-Specific Detection of Fusarium culmorum, F. graminearum and F. avenaceum. Phytopathology 1996, 86, 515–522. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Patiño, B.; González-Jaén, M.T. PCR Detection Assays for the Trichothecene-Producing Species Fusarium graminearum, Fusarium culmorum, Fusarium poae, Fusarium equiseti and Fusarium sporotrichioides. Syst. Appl. Microbiol. 2005, 28, 562–568. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and Quantification of Fusarium culmorum and Fusarium graminearum in Cereals Using PCR Assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Mishra, P.K.; Fox, R.T.V.; Culham, A. Development of a PCR-Based Assay for Rapid and Reliable Identification of Pathogenic Fusaria. FEMS Microbiol. Lett. 2003, 218, 329–332. [Google Scholar] [CrossRef]
- Parry, D.W.; Nicholson, P. Development of a PCR Assay to Detect Fusarium poae in Wheat. Plant Pathol. 1996, 45, 383–391. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Marín, S.; Sanchis, V.; Teresa González-Jaén, M. PCR-Based Strategy to Detect Contamination with Mycotoxigenic Fusarium Species in Maize. Syst. Appl. Microbiol. 2006, 29, 681–689. [Google Scholar] [CrossRef]
- Wilson, A.; Simpson, D.; Chandler, E.; Jennings, P.; Nicholson, P. Development of PCR Assays for the Detection and Differentiation of Fusarium sporotrichioides and Fusarium langsethiae. FEMS Microbiol. Lett. 2004, 233, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, E.M.; Chełkowski, J.; Geiger, H.H. Species-Specific PCR Assays for the Fungal Pathogens Fusarium moniliforme and Fusarium subglutinans and Their Application to Diagnose Maize Ear Rot Disease. J. Phytopathol. 1999, 147, 497–508. [Google Scholar] [CrossRef]
- Patiño, B.; Mirete, S.; González-Jaén, M.T.; Mulé, G.; Rodríguez, M.T.; Vázquez, C. PCR Detection Assay of Fumonisin-Producing Fusarium verticillioides Strains. J. Food Prot. 2004, 67, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, P.; Lees, A.K.; Maurin, N.; Parry, D.W.; Rezanoor, H.N. Development of a PCR Assay to Identify and Quantify Microdochium nivale var. nivale and Microdochium nivale var. majus in Wheat. Physiol. Mol. Plant Pathol. 1996, 48, 257–271. [Google Scholar] [CrossRef]
- Nelson, R. International Plant Pathology: Past and Future Contributions to Global Food Security. Phytopathology 2020, 110, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Cowger, C.; Smith, J.; Boos, D.; Bradley, C.A.; Ransom, J.; Bergstrom, G.C. Managing a Destructive, Episodic Crop Disease: A National Survey of Wheat and Barley Growers’ Experience with Fusarium Head Blight. Plant Dis. 2020, 104, 634–648. [Google Scholar] [CrossRef]
- Salgado, J.D.; Madden, L.V.; Paul, P.A. Efficacy and Economics of Integrating In-Field and Harvesting Strategies to Manage Fusarium Head Blight of Wheat. Plant Dis. 2014, 98, 1407–1421. [Google Scholar] [CrossRef] [Green Version]
- Blandino, M.; Haidukowski, M.; Pascale, M.; Plizzari, L.; Scudellari, D.; Reyneri, A. Integrated Strategies for the Control of Fusarium Head Blight and Deoxynivalenol Contamination in Winter Wheat. Field Crops Res. 2012, 133, 139–149. [Google Scholar] [CrossRef] [Green Version]
- McKee, G.; Cowger, C.; Dill-Macky, R.; Friskop, A.; Gautam, P.; Ransom, J.; Wilson, W. Disease Management and Estimated Effects on DON (Deoxynivalenol) Contamination in Fusarium Infested Barley. Agriculture 2019, 9, 155. [Google Scholar] [CrossRef] [Green Version]
- Mesterházy, Á. Types and Components of Resistance to Fusarium Head Blight of Wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding Strategies and Advances in Line Selection for Fusarium Head Blight Resistance in Wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, H.W.; Christensen, J.J. Factors Affecting Resistance of Wheat to Scab Caused by Gibberella zeae. Phytopathology 1963, 53, 831–838. [Google Scholar]
- Mesterházy, Á.; Bartók, T.; Mirocha, C.G.; Komoróczy, R. Nature of Wheat Resistance to Fusarium Head Blight and the Role of Deoxynivalenol for Breeding. Plant Breed. 1999, 118, 97–110. [Google Scholar] [CrossRef]
- Bai, G.; Su, Z.; Cai, J. Wheat Resistance to Fusarium Head Blight. Can. J. Plant Pathol. 2018, 40, 336–346. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Cloutier, S.; Brulé-Babel, A. Fine Mapping Fhb1, a Major Gene Controlling Fusarium Head Blight Resistance in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1472. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Brulé-Babel, A. Mapping of Fhb2 on Chromosome 6BS: A Gene Controlling Fusarium Head Blight Field Resistance in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Chen, P.D.; Gill, B.S. Molecular Cytogenetic Characterization of Alien Introgressions with Gene Fhb3 for Resistance to Fusarium Head Blight Disease of Wheat. Theor. Appl. Genet. 2008, 117, 1155–1166. [Google Scholar] [CrossRef]
- Xue, S.; Li, G.; Jia, H.; Xu, F.; Lin, F.; Tang, M.; Wang, Y.; An, X.; Xu, H.; Zhang, L.; et al. Fine Mapping Fhb4, a Major QTL Conditioning Resistance to Fusarium Infection in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2010, 121, 147–156. [Google Scholar] [CrossRef]
- Xue, S.; Xu, F.; Tang, M.; Zhou, Y.; Li, G.; An, X.; Lin, F.; Xu, H.; Jia, H.; Zhang, L.; et al. Precise Mapping Fhb5, a Major QTL Conditioning Resistance to Fusarium Infection in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2011, 123, 1055–1063. [Google Scholar] [CrossRef]
- Cainong, J.C.; Bockus, W.W.; Feng, Y.; Chen, P.; Qi, L.; Sehgal, S.K.; Danilova, T.V.; Koo, D.H.; Friebe, B.; Gill, B.S. Chromosome Engineering, Mapping, and Transferring of Resistance to Fusarium Head Blight Disease from Elymus tsukushiensis into Wheat. Theor. Appl. Genet. 2015, 128, 1019–1027. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.; Hou, Y.; Cai, J.; Shen, X.; Zhou, T.; Xu, H.; Ohm, H.W.; Wang, H.; Li, A.; et al. High-Density Mapping of the Major FHB Resistance Gene Fhb7 Derived from Thinopyrum ponticum and Its Pyramiding with Fhb1 by Marker-Assisted Selection. Theor. Appl. Genet. 2015, 128, 2301–2316. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium Head Blight of Wheat and Barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Groenenboom-de Haas, B.H.; Torres, A.M.; Köhl, J.; Chulze, S.N. Biocontrol and Population Dynamics of Fusarium spp. on Wheat Stubble in Argentina. Plant Pathol. 2013, 62, 859–866. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of Wheat Endophytes as Biological Control Agents against Fusarium Head Blight Using Two Different in vitro Tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Slininger, P.J.; Bothast, R.J. Selection and Evaluation of Microorganisms for Biocontrol of Fusarium Head Blight of Wheat Incited by Gibberella zeae. Plant Dis. 2001, 85, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Lipps, P.E.; Slininger, P.J. Field Testing of Antagonists of Fusarium Head Blight Incited by Gibberella zeae. Biol. Control 2004, 29, 245–255. [Google Scholar] [CrossRef]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J.; Slininger, P.J. Greenhouse and Field Evaluation of Biological Control of Fusarium Head Blight on Durum Wheat. Plant Dis. 2002, 86, 1350–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Zhu, Y.; Ma, D.; Xu, W.; Zhou, J.; Yan, H.; Yang, L.; Yin, J. Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight. Agronomy 2019, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic Action of Bacillus subtilis Strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Zachetti, V.G.L.; Cendoya, E.; Nichea, M.J.; Chulze, S.N.; Ramirez, M.L. Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens 2019, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.R.; Doohan, F.M. Comparison of the Efficacy of Chitosan with That of a Fluorescent Pseudomonad for the Control of Fusarium Head Blight Disease of Cereals and Associated Mycotoxin Contamination of Grain. Biol. Control 2009, 48, 48–54. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Luz, C.; Torrijos, R.; Meca, G.; Weber, P.; Bänziger, I.; Voegele, R.T.; Six, J.; Vogelgsang, S. Use of Botanicals to Suppress Different Stages of the Life Cycle of Fusarium graminearum. Phytopathology 2019, 109, 2116–2123. [Google Scholar] [CrossRef] [Green Version]
- Wegulo, S.N.; Bockus, W.W.; Nopsa, J.H.; de Wolf, E.D.; Eskridge, K.M.; Peiris, K.H.S.; Dowell, F.E. Effects of Integrating Cultivar Resistance and Fungicide Application on Fusarium Head Blight and Deoxynivalenol in Winter Wheat. Plant Dis. 2011, 95, 554–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, D.L.; Bradley, C.A.; Ames, K.A.; Willyerd, K.T.; Madden, L.V.; Paul, P.A. Efficacy of Fungicide Applications during and after Anthesis against Fusarium Head Blight and Deoxynivalenol in Soft Red Winter Wheat. Plant Dis. 2014, 98, 1387–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, P.A.; Bradley, C.A.; Madden, L.V.; Dalla Lana, F.; Bergstrom, G.C.; Dill-Macky, R.; Esker, P.D.; Wise, K.A.; McMullen, M.; Grybauskas, A.; et al. Meta-Analysis of the Effects of QoI and DMI Fungicide Combinations on Fusarium Head Blight and Deoxynivalenol in Wheat. Plant Dis. 2018, 102, 2602–2615. [Google Scholar] [CrossRef] [Green Version]
- Lehoczki-Krsjak, S.; Varga, M.; Mesterházy, Á. Distribution of Prothioconazole and Tebuconazole between Wheat Ears and Flag Leaves Following Fungicide Spraying with Different Nozzle Types at Flowering. Pest. Manag. Sci. 2015, 71, 105–113. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Tóth, B.; Varga, M.; Bartók, T.; Szabó-Hevér, Á.; Farády, L.; Lehoczki-Krsjak, S. Role of Fungicides, Application of Nozzle Types, and the Resistance Level of Wheat Varieties in the Control of Fusarium Head Blight and Deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef] [Green Version]
- Behmann, J.; Mahlein, A.K.; Rumpf, T.; Römer, C.; Plümer, L. A Review of Advanced Machine Learning Methods for the Detection of Biotic Stress in Precision Crop Protection. Precis. Agric. 2015, 16, 239–260. [Google Scholar] [CrossRef]
- Bauriegel, E.; Giebel, A.; Geyer, M.; Schmidt, U.; Herppich, W.B. Early Detection of Fusarium Infection in Wheat Using Hyper-Spectral Imaging. Comput. Electron. Agric. 2011, 75, 304–312. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; van den Boom, T.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A Uniform Decimal Code for Growth Stages of Crops and Weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Menesatti, P.; Antonucci, F.; Pallottino, F.; Giorgi, S.; Matere, A.; Nocente, F.; Pasquini, M.; D’Egidio, M.G.; Costa, C. Laboratory vs. in-Field Spectral Proximal Sensing for Early Detection of Fusarium Head Blight Infection in Durum Wheat. Biosyst. Eng. 2013, 114, 289–293. [Google Scholar] [CrossRef]
- Femenias, A.; Gatius, F.; Ramos, A.J.; Sanchis, V.; Marín, S. Use of Hyperspectral Imaging as a Tool for Fusarium and Deoxynivalenol Risk Management in Cereals: A Review. Food Control 2020, 108, 106819. [Google Scholar] [CrossRef]
- Ma, H.; Huang, W.; Jing, Y.; Pignatti, S.; Laneve, G.; Dong, Y.; Ye, H.; Liu, L.; Guo, A.; Jiang, J. Identification of Fusarium Head Blight in Winter Wheat Ears Using Continuous Wavelet Analysis. Sensors 2020, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Jie, L.; Wang, S.; Qi, H.J.; Li, S.W. Classifying Wheat Hyperspectral Pixels of Healthy Heads and Fusarium Head Blight Disease Using a Deep Neural Network in the Wild Field. Remote Sens. 2018, 10, 395. [Google Scholar] [CrossRef] [Green Version]
- Qiu, R.; Yang, C.; Moghimi, A.; Zhang, M.; Steffenson, B.J.; Hirsch, C.D. Detection of Fusarium Head Blight in Wheat Using a Deep Neural Network and Color Imaging. Remote Sens. 2019, 11, 2658. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Y.; Chen, G.; Yin, X.; Hu, R.J.; Gu, C.Y.; Pan, Z.G.; Zhou, X.G.; Chen, Y. Integrating Spectral and Image Data to Detect Fusarium Head Blight of Wheat. Comput. Electron. Agric. 2020, 175, 105588. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, L.; Huang, W.; Dong, Y.; Weng, S.; Zhao, J.; Ma, H.; Liu, L. Detection of Wheat Fusarium Head Blight Using UAV-Based Spectral and Image Feature Fusion. Front. Plant Sci. 2022, 13, 1004427. [Google Scholar] [CrossRef]
- Liu, L.; Dong, Y.; Huang, W.; Du, X.; Ma, H. Monitoring Wheat Fusarium Head Blight Using Unmanned Aerial Vehicle Hyperspectral Imagery. Remote Sens. 2020, 12, 3811. [Google Scholar] [CrossRef]
- Ma, H.; Huang, W.; Dong, Y.; Liu, L.; Guo, A. Using UAV-Based Hyperspectral Imagery to Detect Winter Wheat Fusarium Head Blight. Remote Sens. 2021, 13, 3024. [Google Scholar] [CrossRef]
- Oerke, E.C.; Steiner, U.; Dehne, H.W.; Lindenthal, M. Thermal Imaging of Cucumber Leaves Affected by Downy Mildew and Environmental Conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauriegel, E.; Giebel, A.; Herppich, W.B. Hyperspectral and Chlorophyll Fluorescence Imaging to Analyse the Impact of Fusarium culmorum on the Photosynthetic Integrity of Infected Wheat Ears. Sensors 2011, 11, 3765–3779. [Google Scholar] [CrossRef] [Green Version]
- Bauriegel, E.; Herppich, W.B. Hyperspectral and Chlorophyll Fluorescence Imaging for Early Detection of Plant Diseases, with Special Reference to Fusarium Spec. Infections on Wheat. Agriculture 2014, 4, 32–57. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Barbedo, J.G.A.; del Ponte, E.M.; Bohnenkamp, D.; Mahlein, A.K. From Visual Estimates to Fully Automated Sensor-Based Measurements of Plant Disease Severity: Status and Challenges for Improving Accuracy. Phytopathol. Res. 2020, 2, 9. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Taxonomical position of the genus Fusarium and Microdochium according to MycoBank database, 2022.
Figure 1.
Taxonomical position of the genus Fusarium and Microdochium according to MycoBank database, 2022.

Figure 2.
Disease cycle and symptoms of Fusarium head blight on wheat spikes and kernels.

Figure 3.
Chemical structure of the main mycotoxins produced by Fusarium species on wheat.


Figure 4.
Spectral sensors and their application in integrated disease management of Fusarium head blight on wheat.
Figure 4.
Spectral sensors and their application in integrated disease management of Fusarium head blight on wheat.


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alisaac, E.; Mahlein, A.-K. Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management. Toxins 2023, 15, 192. https://doi.org/10.3390/toxins15030192
AMA Style
Alisaac E, Mahlein A-K. Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management. Toxins. 2023; 15(3):192. https://doi.org/10.3390/toxins15030192
Chicago/Turabian StyleAlisaac, Elias, and Anne-Katrin Mahlein. 2023. "Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management" Toxins 15, no. 3: 192. https://doi.org/10.3390/toxins15030192
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.