The Role of Nitrogen Fertilization on the Occurrence of Regulated, Modified and Emerging Mycotoxins and Fungal Metabolites in Maize Kernels

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Metereological Trends
2.2. Grain Yield
2.3. Fungal Ear Rot and European Corn Borer (ECB) Symptoms
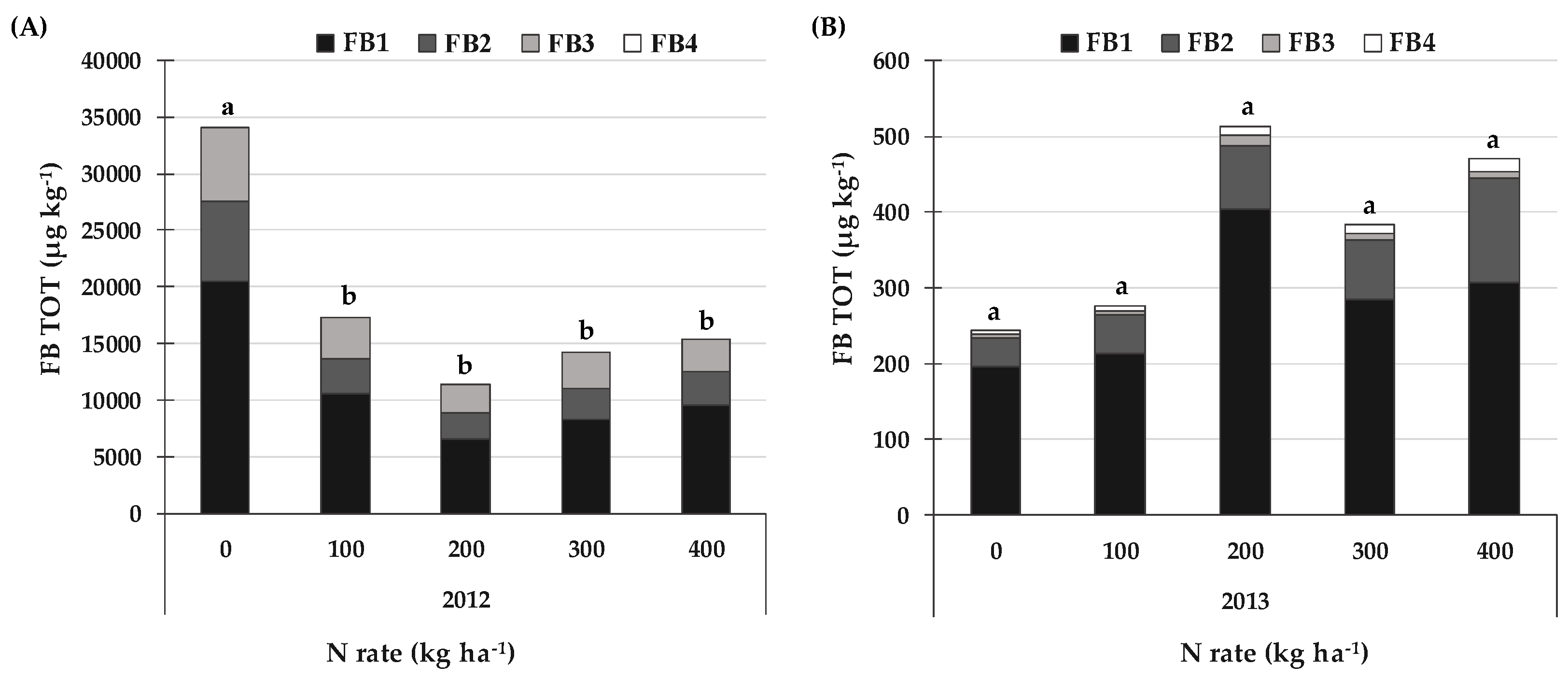
2.4. FBs and the Related Emerging Mycotoxins and Fungal Metabolites Produced by Fusarium spp. Section Liseola
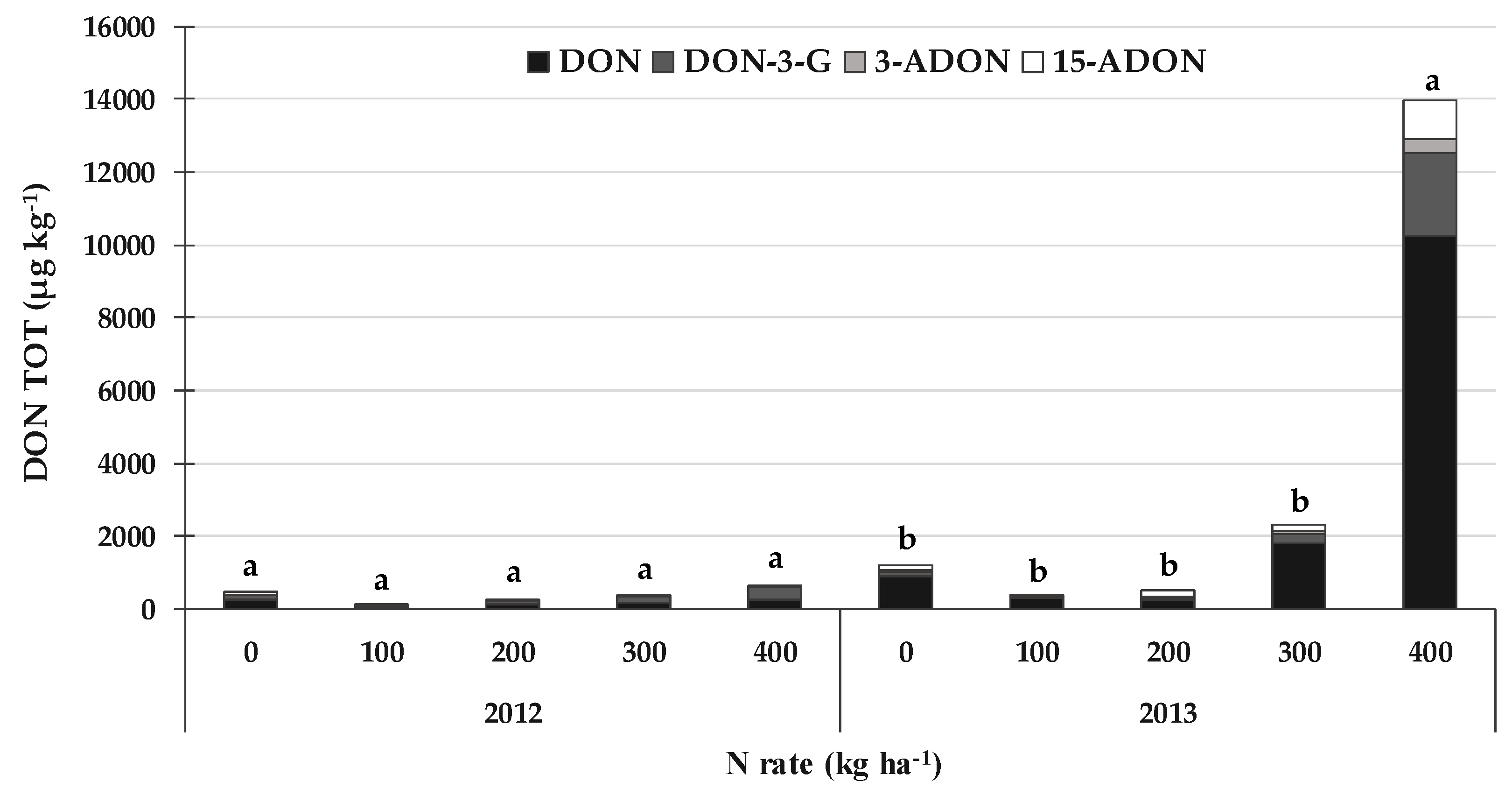
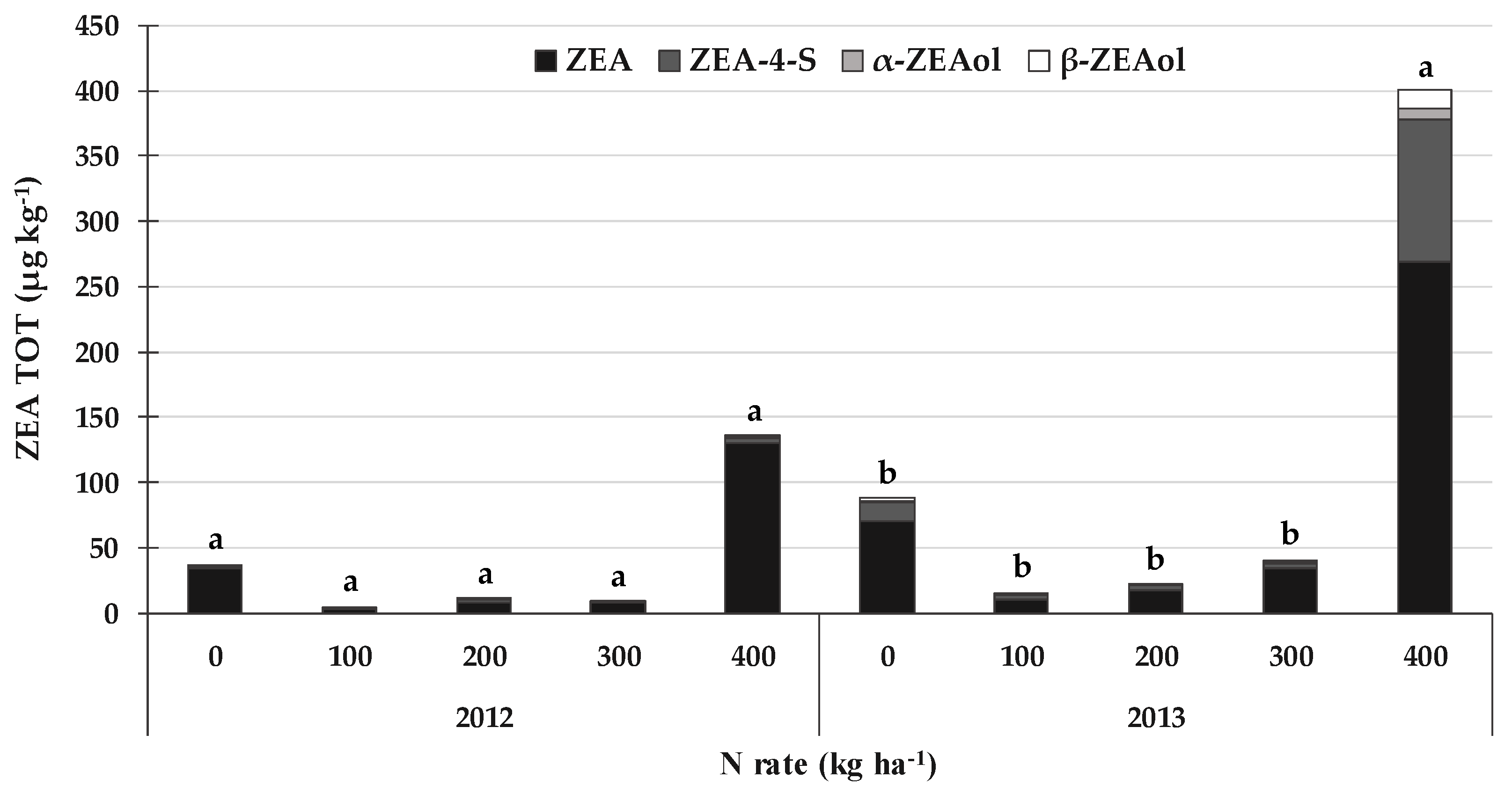
2.5. DON, ZEA and the Related Emerging Mycotoxins and Fungal Metabolites Produced by Fusarium spp. Sections Discolor, Roseum and Sporotrichiella
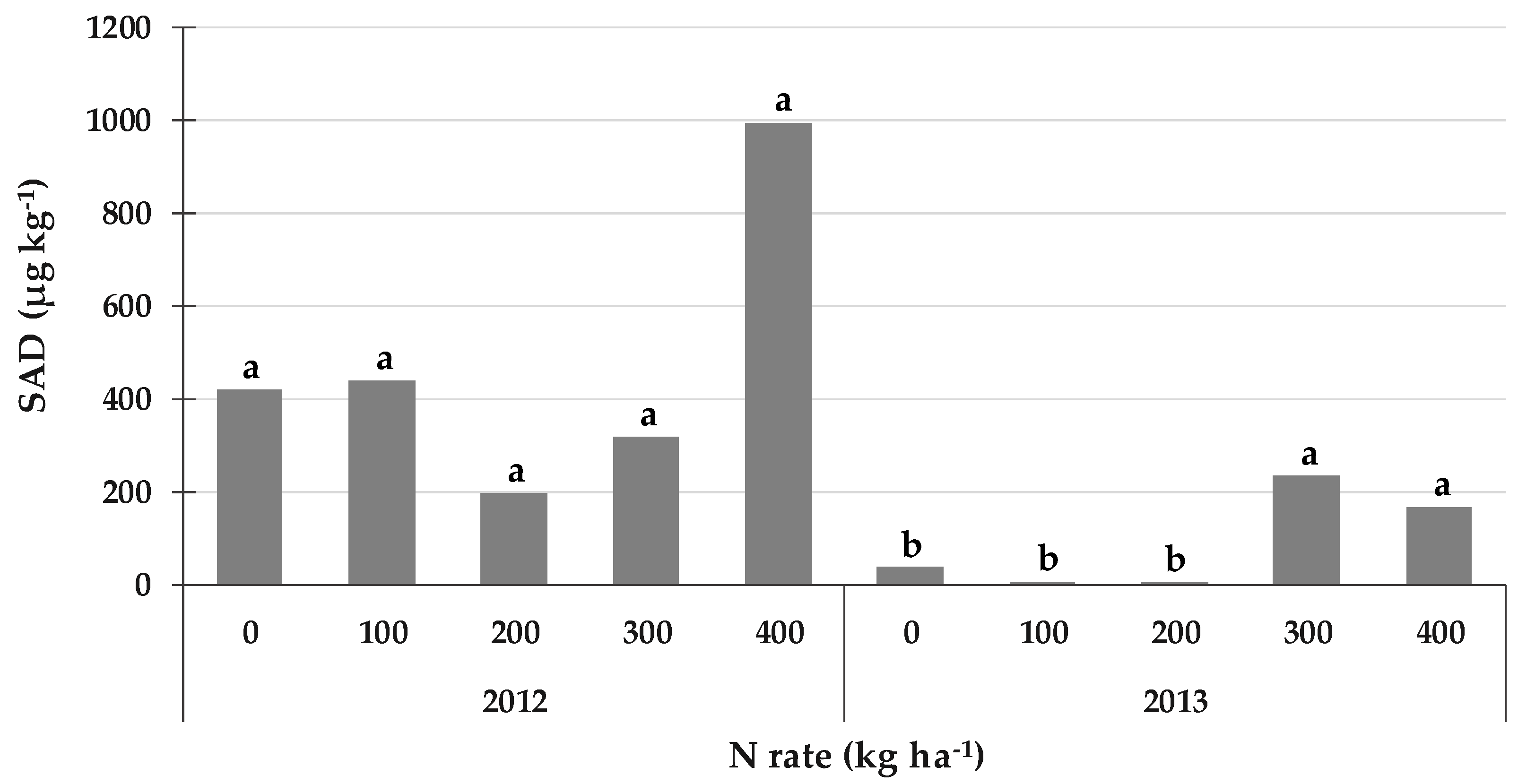
2.6. Secalonic Acid D (SAD): An Aspergillus- and Penicillium-Derived Mycotoxin
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Grain Yield
4.3. ECB and Fungal Ear Rot Symptoms and Fungal Infection
4.4. Multi-Mycotoxin LC-MS/MS Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rao, V.K.; Shilpa, P.; Girisham, S.; Reddy, S.M. Incidence of mycotoxigenic penicillia in feeds of Andhra Pradesh, India. Int. J. Biotechnol. Mol. Biol. Res. 2016, 2, 46–50. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization). Worldwide Regulations for Mycotoxins in Food and Feed in 2003; Information Division; FAO: Rome, Italy, 2003. [Google Scholar]
- Grenier, B.; Oswald, I.P. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Binder, E.M. Managing the risk of mycotoxins in modern feed production. Anim. Feed Sci. Technol. 2007, 133, 149–166. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1126/2007 of 28 September 2007 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union L 2007, 55, 14–17. [Google Scholar]
- European Commission. COMMISSION REGULATION (EC) No 165/2010 of 26 February 2010 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards aflatoxins. Off. J. Eur. Union L 2010, 50, 8–12. [Google Scholar]
- Berthiller, F.; Sulyok, M.; Krska, R.; Schuhmacher, R. Chromatographic methods for the simultaneous determination of mycotoxins and their conjugates in cereals. Int. J. Food Microbiol. 2007, 119, 33–37. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [Green Version]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain). Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar] [CrossRef]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain). Scientific Opinion on the risks to human and animal health related to the presence of moniliformin in food and feed. EFSA J. 2018, 16, 5082. [Google Scholar] [CrossRef]
- European Commission. Commission recommendation (2006/583/EC) of 17 August 2006 on the prevention and reduction of Fusarium toxins in cereals and cereal products. Off. J. Eur. Union l 2006, 234, 35–40. [Google Scholar]
- Ariño, A.; Herrera, M.; Juan, T.; Estopañan, G.; Carramiñana, J.J.; Rota, C.; Herrera, A. Influence of Agricultural Practices on the Contamination of Maize by Fumonisin Mycotoxins. J. Food Prot. 2009, 72, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.M.; Bernd, L.P.; Okumura, R.S.; Takahashi, H.W.; Ono, E.Y.S.; Hirooka, E.Y. Nitrogen fertilization effect on chemical composition and contamination by fungal-fumonisin of maize kernels. Rev. Bras. Cienc. Agrar. 2016, 11, 218–223. [Google Scholar] [CrossRef]
- Blandino, M.; Reyneri, A.; Vanara, F. Influence of nitrogen fertilization on mycotoxin contamination of maize kernels. Crop Prot. 2008, 27, 222–230. [Google Scholar] [CrossRef]
- Marocco, A.; Gavazi, C.; Pietri, A.; Tabaglio, V. On fumonisin incidence in monoculture maize under no-till, conventional tillage and two nitrogen fertilisation levels. J. Sci. Food Agric. 2008, 88, 1217–1221. [Google Scholar] [CrossRef]
- Bocianowski, J.; Szulc, P.; Wáskiewicz, A.; Cyplik, A. The Effect of Agrotechnical Factors on Fusarium Mycotoxins Level in Maize. Agriculture 2020, 10, 528. [Google Scholar] [CrossRef]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current Situation of Mycotoxin Contamination and Co-occurrence in Animal Feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [Green Version]
- Tolosa, J.; Rodríguez-Carrasco, Y.; Ruiz, M.J.; Vila-Donat, P. Multi-mycotoxin occurrence in feed, metabolism and carry-over to animal-derived food products: A review. Food Chem. Toxicol. 2021, 158, 112661. [Google Scholar] [CrossRef]
- Mihalcea, A.; Amariei, S. Study on Contamination with Some Mycotoxins in Maize and Maize-Derived Foods. Appl. Sci. 2022, 12, 2579. [Google Scholar] [CrossRef]
- Mcmahon, P. Effect of nutrition and soil function on pathogens of tropical tree crops. In Plant Pathology; Cumagun, C.J., Ed.; InTech: Rijeka, Croatian, 2012; Chapter 10; pp. 241–272. [Google Scholar] [CrossRef] [Green Version]
- Scarpino, V.; Reyneri, A.; Vanara, F.; Scopel, C.; Causin, R.; Blandino, M. Relationship between European Corn Borer injury, Fusarium proliferatum and F. subglutinans infection and moniliformin contamination in maize. Field Crops Res. 2015, 183, 69–78. [Google Scholar] [CrossRef]
- Scarpino, V.; Reyneri, A.; Sulyok, M.; Krska, R.; Blandino, M. Impact of the insecticide application to maize cultivated in different environmental conditions on emerging mycotoxins. Field Crops Res. 2018, 217, 188–198. [Google Scholar] [CrossRef]
- Blandino, M.; Reyneri, A.; Vanara, F.; Pascale, M.; Haidukowski, M.; Campagna, C. Management of fumonisin contamination in maize kernels through the timing of insecticide application against the European corn borer Ostrinia nubilalis Hübner. Food Addit. Contam. Part A 2009, 26, 1501–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Agency for Research on Cancer (IARC). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Geneva, Switzerland, 1993; Volume 56, pp. 1–599. [Google Scholar] [CrossRef]
- Jonsson, M.; Atosuo, J.; Jestoi, M.; Nathanail, A.V.; Kokkonen, U.-M.; Anttila, M.; Koivisto, P.; Lilius, E.-M.; Peltonen, K. Repeated dose 28-day oral toxicity study of moniliformin in rats. Toxicol. Lett. 2015, 233, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Fremy, J.-M.; Alassane-Kpembi, I.; Oswald, I.P.; Cottrill, B.; Van Egmond, H.P. A review on combined effects of moniliformin and co-occurring Fusarium toxins in farm animals. World Mycotoxin J. 2019, 12, 281–291. [Google Scholar] [CrossRef]
- Dhani, S.; Nagiah, S.; Naidoo, D.B.; Chuturgoon, A.A. Fusaric acid immunotoxicity and MAPK activation in normal peripheral blood mononuclear cells andThp-1 cells. Sci. Rep. 2017, 7, 3051. [Google Scholar] [CrossRef]
- Mamur, S.; Unal, F.; Yilmaz, S.; Erikel, E.; Yuzbasioglu, D. Evaluation of the cytotoxic and genotoxic effects of mycotoxin fusaric acid. Drug Chem. Toxicol. 2018, 43, 149–157. [Google Scholar] [CrossRef]
- Ojcius, D.M.; Zychlinsky, A.; Zheng, L.M.; Young, J.D.-E. Ionophore-induced apoptosis: Role of DNA fragmentation and calcium fluxes. Exp. Cell Res. 1991, 197, 43–49. [Google Scholar] [CrossRef]
- Logrieco, A.; Mulè, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Fornelli, F.; Fogliano, V.; Ritieni, A.; Caiaffa, M.F.; Randazzo, G.; Bottalico, A.; Macchia, L. Fusaproliferin production by Fusarium subglutinans and its toxicity to Artemia salina, SF-9 insect cells, and IARC/LCL 171 human B lymphocytes. Appl. Environ. Microbiol. 1996, 62, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Ritieni, A.; Monti, S.M.; Randazzo, G.; Logrieco, A.; Moretti, A.; Peluso, G.; Ferracane, R.; Fogliano, V. Teratogenic effects of fusaproliferin on chicken embryos. J. Agric. Food Chem. 1997, 45, 3039–3043. [Google Scholar] [CrossRef]
- Santos, M.C.; de Lima Mendonça, M.; Bicas, J.L. Modeling bikaverin production by Fusarium oxysporum CCT7620 in shake flask cultures. Bioresour. Bioprocess. 2020, 7, 13. [Google Scholar] [CrossRef]
- Piacentini, K.C.; Rocha, L.O.; Savi, G.D.; Carnielli-Queiroz, L.; Fontes, L.D.C.; Corrêa, B. Assessment of Toxigenic Fusarium Species and Their Mycotoxins in Brewing Barley Grains. Toxins 2019, 11, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blandino, M.; Scarpino, V.; Giordano, D.; Sulyok, M.; Krska, R.; Vanara, F.; Reyneri, A. Impact of Sowing Time, Hybrid and Environmental Conditions on the Contamination of Maize by Emerging Mycotoxins and Fungal Metabolites. Ital. J. Agron. 2017, 12, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked Mycotoxins Are Efficiently Hydrolyzed by Human Colonic Microbiota Releasing Their Aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [Green Version]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Walburga, S.; Gerrit, S.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Lorenz, N.; Dänicke, S.; Edler, L.; Gottschalk, C.; Lassek, E.; Marko, D.; Rychlik, M.; Mally, A. A critical evaluation of health risk assessment of modified mycotoxins with a special focus on zearalenone. Mycotoxin Res. 2019, 35, 27–46. [Google Scholar] [CrossRef] [Green Version]
- Woelflingseder, L.; Warth, B.; Vierheilig, I.; Schwartz-Zimmermann, H.; Hametner, C.; Nagl, V.; Novak, B.; Šarkanj, B.; Berthiller, F.; Adam, G.; et al. The Fusarium metabolite culmorin suppresses the in vitro glucuronidation of deoxynivalenol. Arch. Toxicol. 2019, 93, 1729–1743. [Google Scholar] [CrossRef] [Green Version]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef]
- Liu, J.-B.; Wang, Y.-M.; Peng, S.-Q.; Han, G.; Dong, Y.-S.; Yang, H.-Y.; Yan, C.-H.; Wang, G.-Q. Toxic effects of Fusarium mycotoxin butenolide on rat myocardium and primary culture of cardiac myocytes. Toxicon 2007, 50, 357–364. [Google Scholar] [CrossRef]
- Scarpino, V.; Blandino, M. Effects of Durum Wheat Cultivars with Different Degrees of FHB Susceptibility Grown under Different Meteorological Conditions on the Contamination of Regulated, Modified and Emerging Mycotoxins. Microorganisms 2021, 9, 408. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, B.; Li, X.; Liu, Q.; Feng, D.; Lan, T.; Kong, F.; Li, Q.; Yuan, J. Nitrogen application affects maize grain filling by regulating grain water relations. J. Integr. Agric. 2022, 21, 977–994. [Google Scholar] [CrossRef]
- Navale, V.; Vamkudoth, K.R.; Ajmera, S.; Dhuri, V. Aspergillus derived mycotoxins in food and the environment: Prevalence, detection, and toxicity. Toxicol. Rep. 2021, 8, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Giese, H. Influence of Carbohydrates on Secondary Metabolism in Fusarium avenaceum. Toxins 2013, 5, 1655–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Zavattaro, L.; Assandri, D.; Grignani, C. Achieving legislation requirements with different nitrogen fertilization strategies: Results from a long term experiment. Eur. J. Agron. 2016, 77, 199–208. [Google Scholar] [CrossRef]
- Grignani, C.; Zavattaro, L.; Sacco, D.; Monaco, S. Production, nitrogen and carbon balance of maize-based forage systems. Eur. J. Agron. 2007, 26, 442–453. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- Malachova, A.; Sulyok, M.; Beltran, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all relevant mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Month | Rainfall | Rainy Days n° | GDDs a |
|---|---|---|---|---|
| mm | Σ °C-day | |||
| 2012 | April | 148 | 13 | 112 |
| May | 147 | 6 | 242 | |
| June | 19 | 2 | 374 | |
| July | 37 | 4 | 415 | |
| August | 46 | 2 | 424 | |
| September | 50 | 5 | 264 | |
| October | 53 | 4 | 169 | |
| Sowing–Harvest b | 322 | 19 | 1673 | |
| 2013 | April | 144 | 9 | 145 |
| May | 147 | 11 | 210 | |
| June | 35 | 2 | 325 | |
| July | 137 | 7 | 430 | |
| August | 59 | 3 | 390 | |
| September | 14 | 1 | 271 | |
| October | 71 | 9 | 145 | |
| Sowing–Harvest b | 426 | 27 | 1657 |
| Year | N Rate | Grain Yield | Ear Rot Incidence a | Ear Rot Severity b | ECB Incidence c | ECB Severity d |
|---|---|---|---|---|---|---|
| (kg ha−1) | (t ha−1) | (%) | (%) | (%) | (%) | |
| 2012 | 0 | 6.7 c | 96.7 a | 18.6 a | 96.7 a | 17.3 a |
| 100 | 9.3 b | 81.7 a | 5.3 b | 80.0 a | 10.2 b | |
| 200 | 11.6 a | 95.0 a | 5.3 b | 85.0 a | 8.6 b | |
| 300 | 12.4 a | 95.0 a | 6.0 b | 91.7 a | 10.0 b | |
| 400 | 13.1 a | 91.7 a | 6.4 b | 86.7 a | 9.5 b | |
| p-value | <0.001 | 0.575 | 0.002 | 0.215 | 0.010 | |
| 2013 | 0 | 10.9 c | 25.0 a | 5.9 a | 6.7 a | 1.0 a |
| 100 | 13.3 b | 26.9 a | 2.9 a | 23.6 a | 1.6 a | |
| 200 | 14.8 a | 22.1 a | 2.0 a | 11.9 a | 1.3 a | |
| 300 | 14.5 ab | 43.3 a | 3.7 a | 31.7 a | 2.3 a | |
| 400 | 14.8 a | 42.9 a | 9.8 a | 27.5 a | 3.0 a | |
| p-value | <0.001 | 0.490 | 0.302 | 0.473 | 0.701 |
| Year | N Rate | BEA | BIK | FA | FUS | Fusarin C | MON |
|---|---|---|---|---|---|---|---|
| (kg ha−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | |
| 2012 | 0 | 891 a | 969 a | 1305 a | 5810 a | <LODa | 1937 a |
| 100 | 280 a | 486 ab | 266 b | 2409 b | <LOD | 1554 ab | |
| 200 | 347 a | 360 b | 245 b | 2563 b | <LOD | 1061 b | |
| 300 | 688 a | 466 b | 323 b | 3183 b | <LOD | 1575 ab | |
| 400 | 1093 a | 496 ab | 246 b | 2583 b | <LOD | 1926 a | |
| p-value | 0.067 | 0.040 | 0.001 | 0.005 | - | 0.023 | |
| 2013 | 0 | 9 a | 47 a | 207 a | 409 a | <LOD a | 75 a |
| 100 | 3 a | 62 a | 140 a | 26 a | <LOD a | 18 a | |
| 200 | 14 a | 97 a | 277 a | 221 a | 39 a | 27 a | |
| 300 | 19 a | 193 a | 247 a | 73 a | 91 a | 99 a | |
| 400 | 9 a | 109 a | 269 a | 92 a | 142 a | 53 a | |
| p-value | 0.714 | 0.095 | 0.777 | 0.345 | 0.134 | 0.427 |
| Year | N Rate | AUR | BUT | CULM | EQU | NIV | T-2 Toxin | HT-2 Toxin |
|---|---|---|---|---|---|---|---|---|
| (kg ha−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | (µg kg−1) | |
| 2012 | 0 | 468 a | 56 a | 171 a | 7 a | <LOD a a | 4 a | 12 a |
| 100 | 78 a | 53 a | 22 a | 205 a | 2 a | <LOD b a | <LOD c a | |
| 200 | 58 a | 33 a | 72 a | 42 a | 3 a | 3 a | 9 a | |
| 300 | 265 a | 39 a | 39 a | 171 a | <LOD a | 5 a | 9 a | |
| 400 | 606 a | 36 a | 73 a | 209 a | <LOD a | 9 a | 7 a | |
| p-value | 0.251 | 0.791 | 0.107 | 0.204 | 0.555 | 0.767 | 0.873 | |
| 2013 | 0 | 1171 b | 131 b | 885 b | 3 a | 3 b | 0.9 a | <LOD a |
| 100 | 588 b | 70 b | 1910 b | 5 a | <LOD b | <LOD a | <LOD a | |
| 200 | 2822 b | 111 b | 1713 b | 1 a | 3 b | <LOD a | <LOD a | |
| 300 | 1469 b | 119 b | 1970 b | 32 a | 2 b | <LOD a | <LOD a | |
| 400 | 13876 a | 656 a | 6004 a | 41 a | 23 a | 3 a | 4 a | |
| p-value | <0.001 | 0.039 | 0.038 | 0.490 | <0.001 | 0.507 | 0.237 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarpino, V.; Sulyok, M.; Krska, R.; Reyneri, A.; Blandino, M. The Role of Nitrogen Fertilization on the Occurrence of Regulated, Modified and Emerging Mycotoxins and Fungal Metabolites in Maize Kernels. Toxins 2022, 14, 448. https://doi.org/10.3390/toxins14070448
Scarpino V, Sulyok M, Krska R, Reyneri A, Blandino M. The Role of Nitrogen Fertilization on the Occurrence of Regulated, Modified and Emerging Mycotoxins and Fungal Metabolites in Maize Kernels. Toxins. 2022; 14(7):448. https://doi.org/10.3390/toxins14070448
Chicago/Turabian StyleScarpino, Valentina, Michael Sulyok, Rudolf Krska, Amedeo Reyneri, and Massimo Blandino. 2022. "The Role of Nitrogen Fertilization on the Occurrence of Regulated, Modified and Emerging Mycotoxins and Fungal Metabolites in Maize Kernels" Toxins 14, no. 7: 448. https://doi.org/10.3390/toxins14070448