Efficacy of Fumonisin Esterase in Piglets as Animal Model for Fumonisin Detoxification in Humans: Pilot Study Comparing Intraoral to Intragastric Administration


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
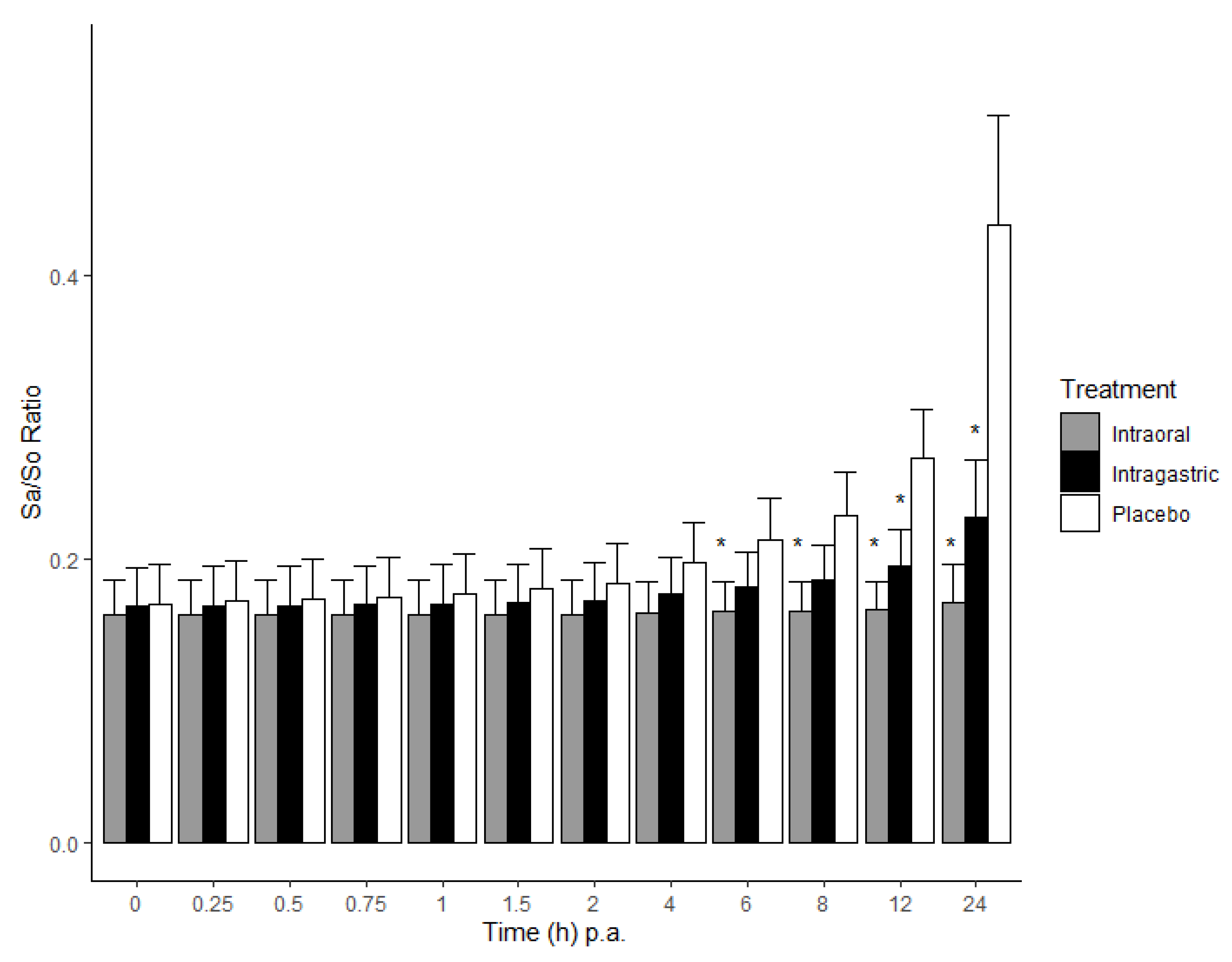
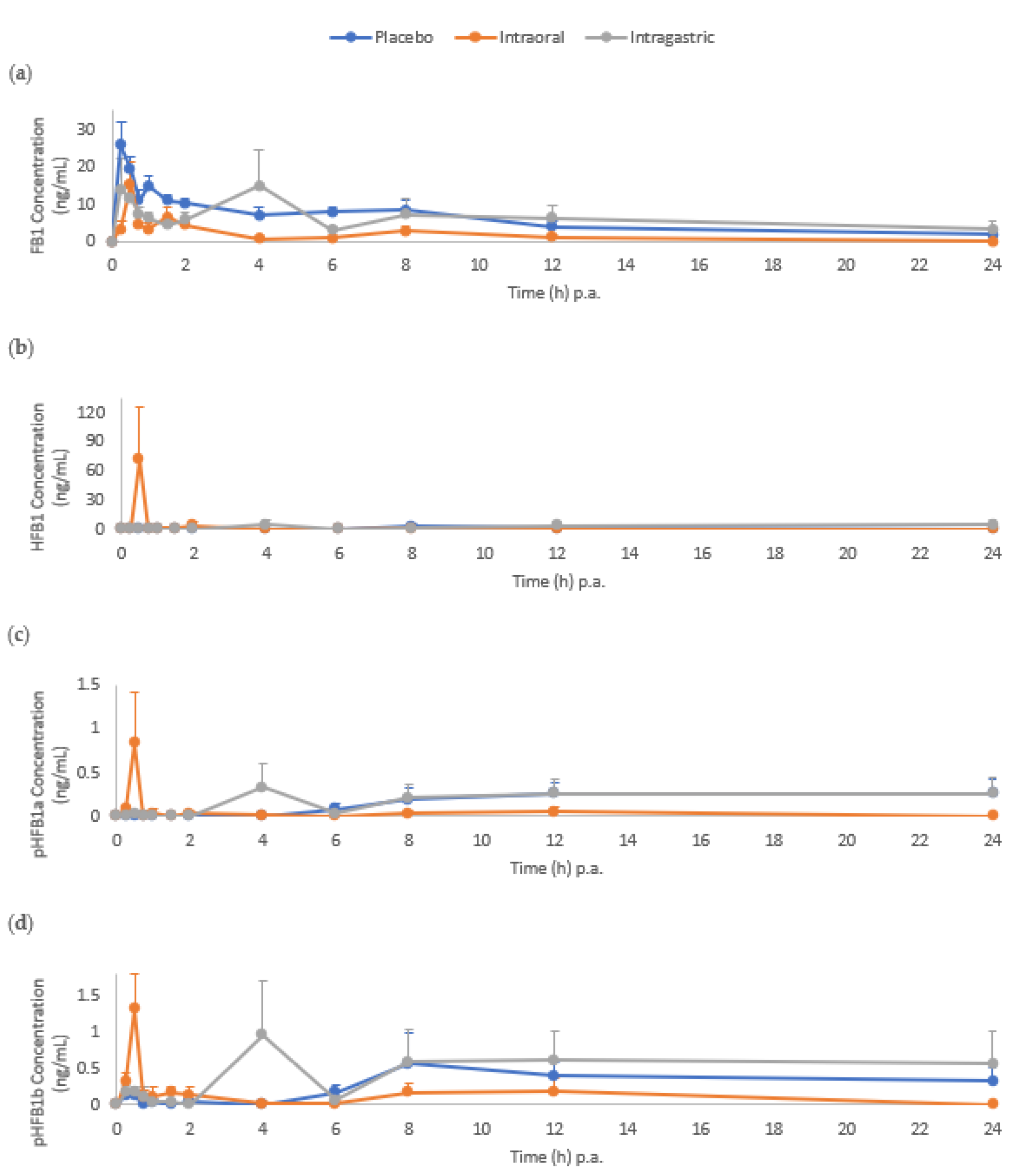
2.1. Biomarkers in Serum
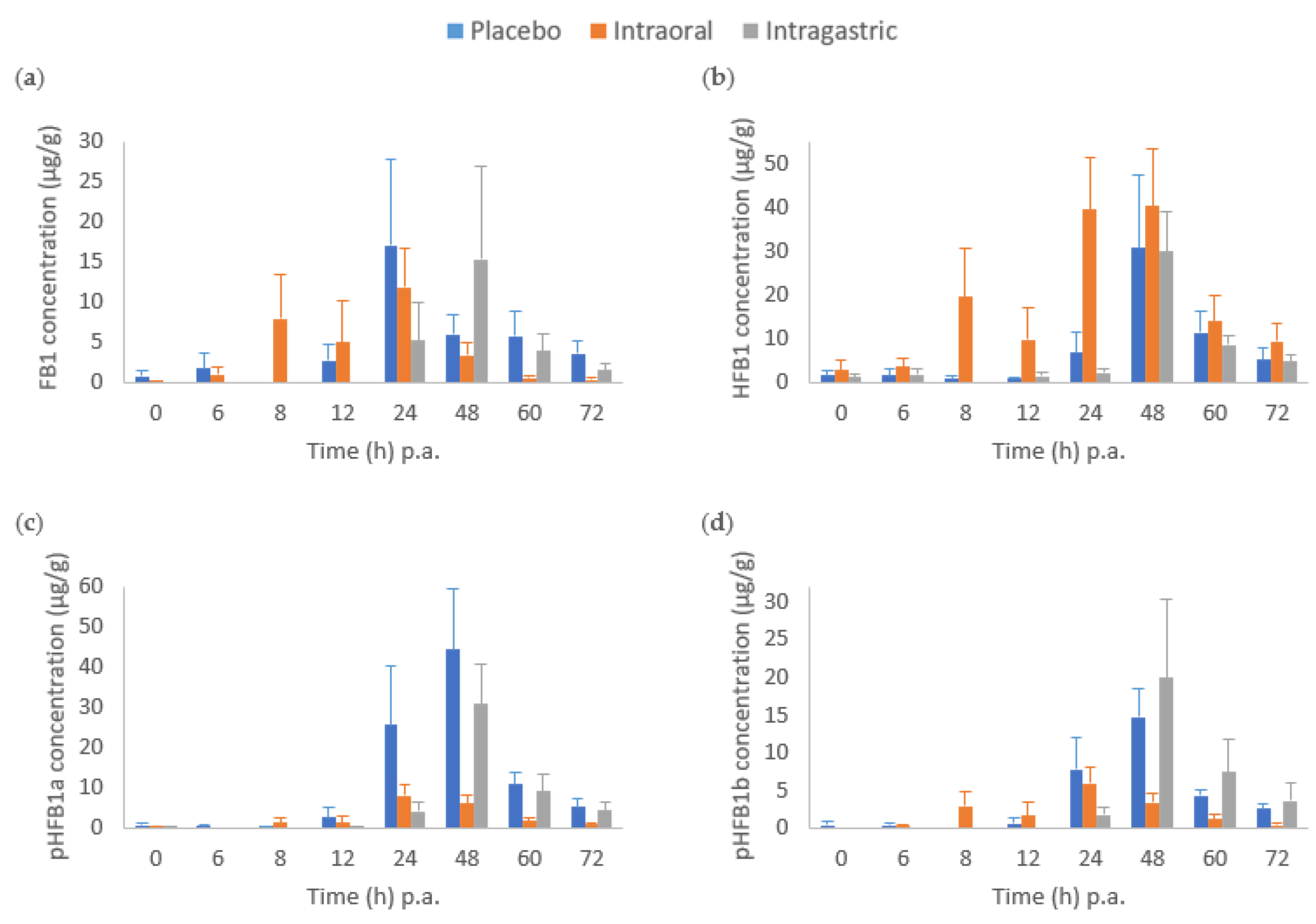
2.2. Biomarkers in Faeces
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
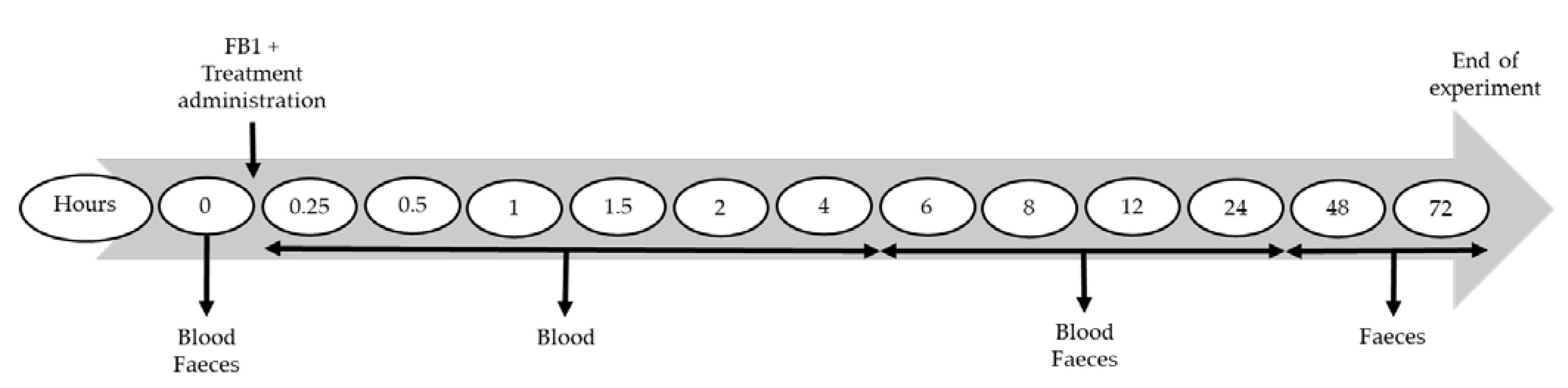
5.2. Experimental Design
5.3. Products, Treatment Preparation and Administration
5.4. Biomarker Analysis
5.5. Toxicokinetic and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef] [PubMed]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2010, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Dowswell, C.R.; Paliwal, R.L.; Cantrell, R.P. Maize in the World Economy. In Maize in the Third World; Taylor & Francis: Abingdon, UK, 1996; ISBN 9780367012311. [Google Scholar]
- Marasas, W.F.O.; Miller, J.D.; Riley, R.T.; Visconti, A. Fumonisin B1; World Health Organisation: Geneva, CH, 2000; ISBN 9241572191. [Google Scholar]
- Voss, K.A.; Riley, R.T.; Snook, M.E.; Gelineau-van Waes, J. Reproductive and Sphingolipid Metabolic Effects of Fumonisin B1 and its Alkaline Hydrolysis Product in LM/Bc Mice: Hydrolyzed Fumonisin B1 Did Not Cause Neural Tube Defects. Toxicol. Sci. 2009, 112, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef]
- WHO. Food Safety Digest—Fumonisins; World Health Organisation: Geneva, CH, 2018. [Google Scholar]
- Gelineau-van Waes, J.; Voss, K.A.; Stevens, V.L.; Speer, M.C.; Riley, R.T. Maternal Fumonisin Exposure as a Risk Factor for Neural Tube Defects. Adv. Food Nutr. Res. 2009, 56, 145–181. [Google Scholar] [CrossRef]
- Moretti, A.; Logrieco, A.F.; Susca, A. Mycotoxins: An Underhand Food Problem. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Humana Press: Totowa, NJ, USA, 2017; pp. 3–12. ISBN 9781493967056. [Google Scholar]
- Chen, C.; Mitchell, N.J.; Gratz, J.; Houpt, E.R.; Gong, Y.; Egner, P.A.; Groopman, J.D.; Riley, R.T.; Showker, J.L.; Svensen, E.; et al. Exposure to aflatoxin and fumonisin in children at risk for growth impairment in rural Tanzania. Environ. Int. 2018, 115, 29–37. [Google Scholar] [CrossRef]
- Lyon, F.R.; Wild, C.; Miller, J.; Groopman, J. (Eds.) IARC Effects of aflatoxins and fumonisins on child growth. In Mycotoxin Control in Low- and Middle- Income Countries; International Agency for Research on Cancer: Lyon, France, 2015; pp. 17–22. [Google Scholar]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef]
- Smith, G.W. Fumonisins. In Veterinary Toxicology; Gupta, R.C., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 1003–1018. ISBN 9780128114100. [Google Scholar]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, S91–S96. [Google Scholar] [CrossRef] [Green Version]
- Lowry, L.K. Role of biomarkers of exposure in the assessment of health risks. Toxicol. Lett. 1995, 77, 31–38. [Google Scholar] [CrossRef]
- Schertz, H.; Dänicke, S.; Frahm, J.; Schatzmayr, D.; Dohnal, I.; Bichl, G.; Schwartz-Zimmermann, H.E.; Colicchia, S.; Breves, G.; Teifke, J.P.; et al. Biomarker Evaluation and Toxic Effects of an Acute Oral and Systemic Fumonisin Exposure of Pigs with a Special Focus on Dietary Fumonisin Esterase Supplementation. Toxins 2018, 10, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masching, S.; Naehrer, K.; Schartz-Zimmermann, H.-E.; Sărăndan, M.; Schaumberger, S.; Dohnal, I.; Nagl, V.; Schatzmayr, D. Gastrointestinal Degradation of Fumonisin B1 by Carboxylesterase FumD Prevents Fumonisin Induced Alteration of Sphingolipid Metabolism in Turkey and Swine. Toxins 2016, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Schwartz-Zimmermann, H.E.; Hartinger, D.; Doupovec, B.; Gruber-Dorninger, C.; Aleschko, M.; Schaumberger, S.; Nagl, V.; Hahn, I.; Berthiller, F.; Schatzmayr, D.; et al. Application of biomarker methods to investigate FUMzyme mediated gastrointestinal hydrolysis of fumonisins in pigs. World Mycotoxin J. 2018, 11, 201–214. [Google Scholar] [CrossRef]
- Shephard, G.S.; van der Westhuizen, L.; Sewram, V. Biomarkers of exposure to fumonisin mycotoxins: A review. Food Addit. Contam. 2007, 24, 1196–1201. [Google Scholar] [CrossRef]
- Qiu, M.; Liu, X. Determination of sphinganine, sphingosine and Sa/So ratio in urine of humans exposed to dietary fumonisin B1. Food Addit. Contam. 2001, 18, 263–269. [Google Scholar] [CrossRef]
- van der Westhuizen, L.; Shephard, G.S.; Gelderblom, W.C.A.; Torres, O.; Riley, R.T. Fumonisin biomarkers in maize eaters and implications for human disease. World Mycotoxin J. 2013, 6, 223–232. [Google Scholar] [CrossRef]
- Van Der Westhuizen, L.; Shephard, G.S.; Burger, H.M.; Rheeder, J.P.; Gelderblom, W.C.A.; Wild, C.P.; Gong, Y.Y. Fumonisin B1 as a Urinary Biomarker of Exposure in a Maize Intervention Study Among South African Subsistence Farmers. Cancer Epidemiol. Biomark. Prev. 2011, 20, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Schelstraete, W.; Devreese, M.; Croubels, S. Comparative toxicokinetics of Fusarium mycotoxins in pigs and humans. Food Chem. Toxicol. 2020, 137, 111140. [Google Scholar] [CrossRef]
- Rose, L.J.; Okoth, S.; Rose, J.; Flett, B.C.; van Rensburg, B.J.; Viljoen, A. Preharvest Management Strategies and Their Impact on Mycotoxigenic Fungi and Associated Mycotoxins. In Mycotoxins—Impact and Management Strategies; IntechOpen: London, UK, 2018; pp. 41–57. [Google Scholar]
- Kamala, A.; Kimanya, M.; Haesaert, G.; Tiisekwa, B.; Madege, R.; Degraeve, S.; Cyprian, C.; De Meulenaer, B. Local post-harvest practices associated with aflatoxin and fumonisin contamination of maize in three agro ecological zones of Tanzania. Food Addit. Contam. Part A 2016, 33, 551–559. [Google Scholar] [CrossRef]
- Kagot, V.; Okoth, S.; De Boevre, M.; De Saeger, S. Biocontrol of Aspergillus and Fusarium Mycotoxins in Africa: Benefits and Limitations. Toxins 2019, 11, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Liu, X.; Li, J. Updating techniques on controlling mycotoxins—A review. Food Control 2018, 89, 123–132. [Google Scholar] [CrossRef]
- Odukoya, J.O.; De Saeger, S.; De Boevre, M.; Adegoke, G.O.; Audenaert, K.; Croubels, S.; Antonissen, G.; Vermeulen, K.; Gbashi, S.; Njobeh, P.B. Effect of Selected Cooking Ingredients for Nixtamalization on the Reduction of Fusarium Mycotoxins in Maize and Sorghum. Toxins 2021, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, L.W. Evaluation of Mycotoxin Binders. In Proceedings of the 4th Mid-Atlantic Nutrition Conference, Timonium, MD, USA; 2006; pp. 132–143. [Google Scholar]
- Dell’Orto, V.; Baldi, G.; Cheli, F. Mycotoxins in silage: Checkpoints for effective management and control. World Mycotoxin J. 2015, 8, 603–617. [Google Scholar] [CrossRef]
- EFSA Scientific Opinion on the safety and efficacy of fumonisin esterase (FUMzyme®) as a technological feed additive for pigs. EFSA J. 2014, 12, 3667. [CrossRef]
- Grenier, B.; Bracarense, A.F.L.; Schwartz, H.E.; Trumel, C.; Cossalter, A.-M.; Schatzmayr, G.; Kolf-Clauw, M.; Moll, W.-D.; Oswald, I.P. The low intestinal and hepatic toxicity of hydrolyzed fumonisin B1 correlates with its inability to alter the metabolism of sphingolipids. Biochem. Pharmacol. 2012, 83, 1465–1473. [Google Scholar] [CrossRef]
- Hahn, I.; Nagl, V.; Schwartz-zimmermann, H.E.; Varga, E.; Schwarz, C.; Slavik, V.; Reisinger, N.; Malachová, A.; Cirlini, M.; Generotti, S.; et al. Effects of orally administered fumonisin B1 (FB1), partially hydrolysed FB1, hydrolysed FB1 and N-(1-deoxy-D-fructos-1-yl) FB1 on the sphingolipid metabolism in rats. Food Chem. Toxicol. 2015, 76, 11–18. [Google Scholar] [CrossRef]
- Hoag, S.W. Capsules Dosage Form: Formulation and Manufacturing Considerations. In Developing Solid Oral Dosage Forms, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 723–747. ISBN 9780128024478. [Google Scholar]
- Gasthuys, E.; Vandecasteele, T.; De Bruyne, P.; Vande Walle, J.; De Backer, P.; Cornillie, P.; Devreese, M.; Croubels, S. The Potential Use of Piglets as Human Pediatric Surrogate for Preclinical Pharmacokinetic and Pharmacodynamic Drug Testing. Curr. Pharm. Des. 2016, 22, 4069–4085. [Google Scholar] [CrossRef]
- UNEP; UNICEF; WHO. Children in the New Millennium: Environmental Impact on Health; World Health Organisation: Geneva, CH, 2002; ISBN 92-807-2065-1. [Google Scholar]
- Catteuw, A.; Devreese, M.; De Baere, S.; Antonissen, G.; Huybrechts, B.; Ivanova, L.; Uhlig, S.; Martens, A.; De Saeger, S.; De Boevre, M.; et al. Toxicokinetic Studies in Piglets Reveal Age-Related Differences in Systemic Exposure to Zearalenone, Zearalenone-14-Glucoside, and Zearalenone-14-Sulfate. J. Agric. Food Chem. 2020, 68, 7757–7764. [Google Scholar] [CrossRef]
- Raiola, A.; Tenore, G.C.; Manyes, L.; Meca, G.; Ritieni, A. Risk analysis of main mycotoxins occurring in food for children: An overview. Food Chem. Toxicol. 2015, 84, 169–180. [Google Scholar] [CrossRef]
- Neckermann, K.; Claus, G.; De Baere, S.; Antonissen, G.; Lebrun, S.; Gemmi, C.; Taminiau, B.; Douny, C.; Scippo, M.L.; Schatzmayr, D.; et al. The efficacy and effect on gut microbiota of an aflatoxin binder and a fumonisin esterase using an in vitro simulator of the human intestinal microbial ecosystem (SHIME®). Food Res. Int. 2021, 145, 110395. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Mengelers, M.; De Saeger, S.; De Boevre, M. Mycotoxin Biomarkers of Exposure: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yayeh, T.; Jeong, H.R.; Park, Y.S.; Moon, S.; Sur, B.; Yoo, H.-S.; Oh, S. Fumonisin B1-Induced Toxicity Was Not Exacerbated in Glutathione Peroxidase-1/Catalase Double Knock Out Mice. Biomol. Ther. 2021, 29, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Norred, W.P.; Bacon, C.W.; Riley, R.T.; Merrill, A.H.J. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with Fusarium moniliforme. J. Biol. Chem. 1991, 266, 14486–14490. [Google Scholar] [CrossRef]
- van der Westhuizen, L.; Brown, N.L.; Marasas, W.F.O.; Swanevelder, S.; Shephard, G.S. Sphinganine/sphingosine ratio in plasma and urine as a possible biomarker for fumonisin exposure in humans in rural areas of Africa. Food Chem. Toxicol. 1999, 37, 1153–1158. [Google Scholar] [CrossRef]
- Davis, S.S.; Illum, L.; Hinchcliffe, M. Gastrointestinal transit of dosage forms in the pig. J. Pharm. Pharmacol. 2001, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Schop, M.; Jansman, A.J.M.; De Vries, S.; Gerrits, W.J.J. Increasing intake of dietary soluble nutrients affects digesta passage rate in the stomach of growing pigs. Br. J. Nutr. 2019, 121, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Schertz, H.; Kluess, J.; Frahm, J.; Schatzmayr, D.; Dohnal, I.; Bichl, G.; Schwartz-zimmermann, H.; Breves, G.; Dänicke, S. Oral and Intravenous Fumonisin Exposure in Pigs—A Single-Dose Treatment Experiment Evaluating Toxicokinetics and Detoxification. Toxins 2018, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Meyer, K.; Gottschalk, C.; Mamet, R.; Kametler, L.; Bauer, J.; Horn, P.; Kovacs, F.; Kovacs, M. In vitro microbial metabolism of fumonisin B1. Food Addit. Contam. 2007, 24, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Balogh, K.; Weber, M.; Mézes, M.; Kametler, L.; Pósa, R.; Mamet, R.; Bauer, J.; Horn, P.; Kovács, F.; et al. Absorption, distribution and elimination of fumonisin B1 metabolites in weaned piglets. Food Addit. Contam. 2008, 25, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.A.; Zsolnai, A.; Kovacs, M.; Bors, I.; Bonai, A.; Bota, B.; Szabó-Fodor, J. In vitro interaction between fumonisin B1 and the intestinal microflora of pigs. Pol. J. Microbiol. 2017, 66, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prelusky, D.B.; Trenhoim, H.L.; Marc, E.S. Pharmacokinetic fate of 14C-labelled fumonisin B1 in swine. Nat. Toxins 1994, 80, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dilkin, P.; Direito, G.; Simas, M.M.S.; Mallmann, C.A.; Corrêa, B. Toxicokinetics and toxicological effects of single oral dose of fumonisin B1 containing Fusarium verticillioides culture material in weaned piglets. Chem. -Biol. Interact. 2010, 185, 157–162. [Google Scholar] [CrossRef]
- EC Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L 229, 7–9.
- Grenier, B.; Schwartz-Zimmermann, H.E.; Gruber-Dorninger, C.; Dohnal, I.; Aleschko, M.; Schatzmayr, G.; Moll, W.D.; Applegate, T.J. Enzymatic hydrolysis of fumonisins in the gastrointestinal tract of broiler chickens. Poult. Sci. 2017, 96, 4342–4351. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D.; Shackelford, D.D.; Leslie, J.F.; Nelson, P.E. Heritability of Fumonisin B1 Production in Gibberella fujikuroi Mating Population A. Appl. Environ. Microbiol. 1992, 58, 2799–2805. [Google Scholar] [CrossRef] [Green Version]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.; Grabherr, R. Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing 2020. Available online: http://www.r-project.org/index.html (accessed on 2 February 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum | Faeces |
|---|---|
| Sa/So ratio FB1 HFB1, pHFB1a, pHFB1b | FB1 HFB1, pHFB1a, pHFB1b |
| Sa/So (sphinganine/sphingosine) ratio, FB1 (fumonisin B1), HFB1 (hydrolysed fumonisin B1 or aminopentol), pHFB1a and pHFB1b (partially hydrolysed fumonisin B1a and B1b) | |
| Mycotoxin | Treatment | Maximum Observed Concentration (Cmax) (ng/mL) ± SEM | Time of Observed Maximum Concentration (Tmax) (h) ± SEM | Area under the Concentration-Time Curve Time Zero to Time Last Measurable Concentration (AUC0→t) (h × ng/mL) ± SEM | Difference in AUC0→t between Control (Placebo) and Treated (Intraoral or Intragastric) Groups in % |
|---|---|---|---|---|---|
| FB1 | Placebo | 23 ± 3.8 | 2.2 ± 1.1 | 128 ± 15.5 | |
| Intraoral | 10 ± 2.8 | 1.9 ± 0.9 | 51.4 ± 17.0 ** | −59.8 ** | |
| Intragastric | 22 ± 7.8 | 7.0 ± 3.1 | 148 ± 51.7 | +15.6 | |
| HFB1 | Placebo | 6.4 ± 4.8 | 5.6 ± 2.7 | 36.5 ± 31.0 | |
| Intraoral | 33 ± 21 | 3.2 ± 1.2 | 49.8 ± 24.3 | +36.4 | |
| Intragastric | 23 ± 6.9 | 11 ± 4.7 | 141 ± 54.4 | +286 | |
| pHFB1a | Placebo | 0.7 ± 0.2 | 18 ± 3.5 | 6.80 ± 2.18 | |
| Intraoral | 0.6 ± 0.3 | 2.3 ± 1.5 | 2.12 ± 1.04 | −68.8 | |
| Intragastric | 0.8 ± 0.3 | 13 ± 3.7 | 7.33 ± 2.82 | +7.79 | |
| pHFB1b | Placebo | 0.9 ± 0.4 | 9.9 ± 4.0 | 7.60 ± 2.43 | |
| Intraoral | 0.9 ± 0.2 | 2.5 ± 1.2 | 3.07 ± 1.14 | −59.6 | |
| Intragastric | 1.8 ± 0.7 | 11 ± 3.4 | 15.7 ± 6.60 | +107 |
| Mycotoxin | Treatment | Maximum Observed Concentration (Cmax) (µg/g) ± SEM | Time of Observed Maximum Concentration (Tmax) (h) ± SEM | Area under the Concentration-Time Curve Time Zero to Time Last Measurable Concentration (AUC0→t) (h × µg/g) ± SEM | Difference in AUC0→t between Control (Placebo) and Treated (Intraoral or Intragastric) Groups in % |
|---|---|---|---|---|---|
| FB1 | Placebo | 20 ± 10 | 39 ± 5.7 | 617 ± 305 | |
| Intraoral | 16 ± 4.5 | 31 ± 4.4 | 391 ± 125 | −36.6 | |
| Intragastric | 18 ± 13 | 52 ± 2.5 | 531 ± 384 | −13.9 | |
| HFB1 | Placebo | 28 ± 15 | 33 ± 7.7 | 795 ± 323 | |
| Intraoral | 66 ± 11 | 34 ± 5.6 | 1745 ± 289 * | +119 * | |
| Intragastric | 30 ± 9.2 | 48 ± 0.0 | 796 ± 185 ∆ | +0.13 ∆ | |
| pHFB1a | Placebo | 56 ± 15 | 34 ± 4.8 | 1512 ± 333 | |
| Intraoral | 11 ± 2.1 | 38 ± 5.3 | 318 ± 73.0 * | −79.0 * | |
| Intragastric | 31 ± 9.9 | 48 ± 0.0 | 977 ± 335 a | −35.4 a | |
| pHFB1b | Placebo | 17 ± 3.9 | 38 ± 4.8 | 552 ± 109 | |
| Intraoral | 8.2 ± 1.7 | 29 ± 6.0 | 221 ± 43.8 * | −60.0 * | |
| Intragastric | 23 ± 12 | 48 ± 0.0 | 777 ± 420 | +40.8 |
| Treatment Groups | Products Administered | Dose (/kg BW) | Administration Route |
|---|---|---|---|
| Placebo (n = 7) | Fumonisin B1 | 2 mg | Intraoral |
| Fumonisin esterase | - | - | |
| Maltodextrin | 300 mg | Intraoral + Intragastric | |
| Intraoral (n = 8) | Fumonisin B1 | 2 mg | Intraoral |
| Fumonisin esterase | 3 U | Intraoral | |
| Maltodextrin | 300 mg | Intragastric | |
| Intragastric (n = 8) | Fumonisin B1 | 2 mg | Intraoral |
| Fumonisin esterase | 3 U | Intragastric | |
| Maltodextrin | 300 mg | Intraoral |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neckermann, K.; Antonissen, G.; Doupovec, B.; Schatzmayr, D.; Gathumbi, J.; Delcenserie, V.; Uhlig, S.; Croubels, S. Efficacy of Fumonisin Esterase in Piglets as Animal Model for Fumonisin Detoxification in Humans: Pilot Study Comparing Intraoral to Intragastric Administration. Toxins 2022, 14, 136. https://doi.org/10.3390/toxins14020136
Neckermann K, Antonissen G, Doupovec B, Schatzmayr D, Gathumbi J, Delcenserie V, Uhlig S, Croubels S. Efficacy of Fumonisin Esterase in Piglets as Animal Model for Fumonisin Detoxification in Humans: Pilot Study Comparing Intraoral to Intragastric Administration. Toxins. 2022; 14(2):136. https://doi.org/10.3390/toxins14020136
Chicago/Turabian StyleNeckermann, Kaat, Gunther Antonissen, Barbara Doupovec, Dian Schatzmayr, James Gathumbi, Véronique Delcenserie, Silvio Uhlig, and Siska Croubels. 2022. "Efficacy of Fumonisin Esterase in Piglets as Animal Model for Fumonisin Detoxification in Humans: Pilot Study Comparing Intraoral to Intragastric Administration" Toxins 14, no. 2: 136. https://doi.org/10.3390/toxins14020136