
Ecotoxicological Effects of Four Commonly Used Organic Solvents on the Scleractinian Coral Montipora digitata

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
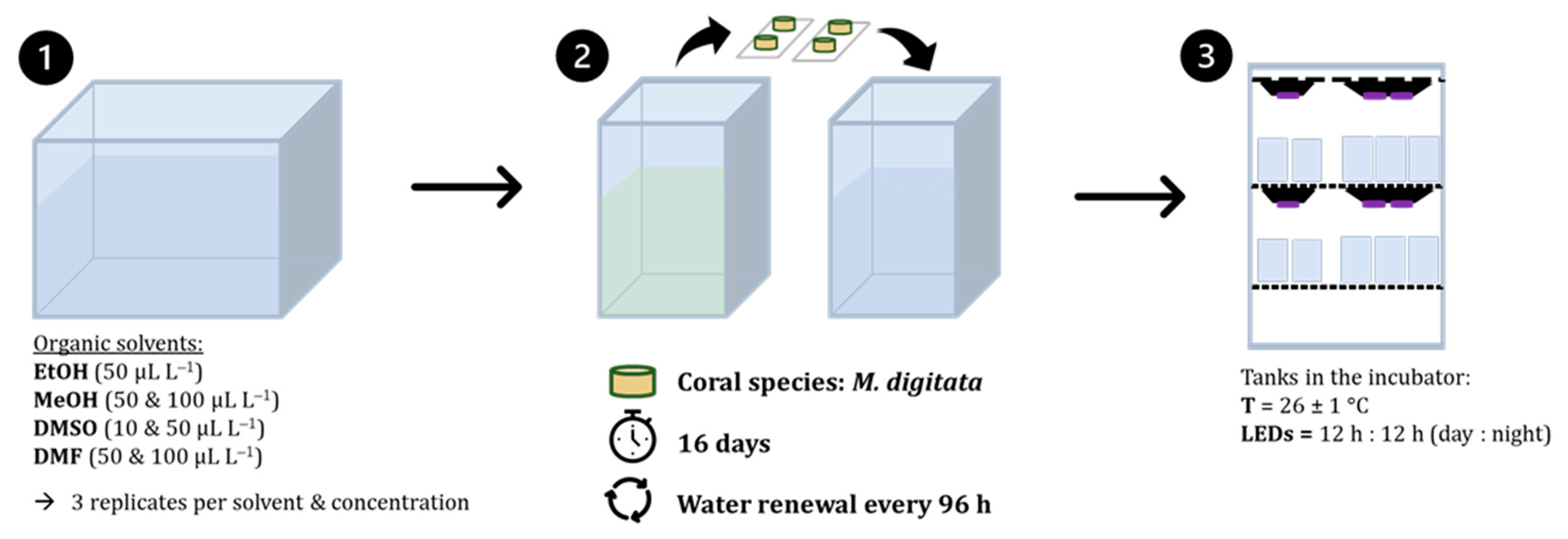
2.1. Husbandry and Supply of the Test Organism Montipora Digitata
2.2. Experimental Set Up
2.2.1. Preparation of Test Media and Controls
2.2.2. Static Renewal System
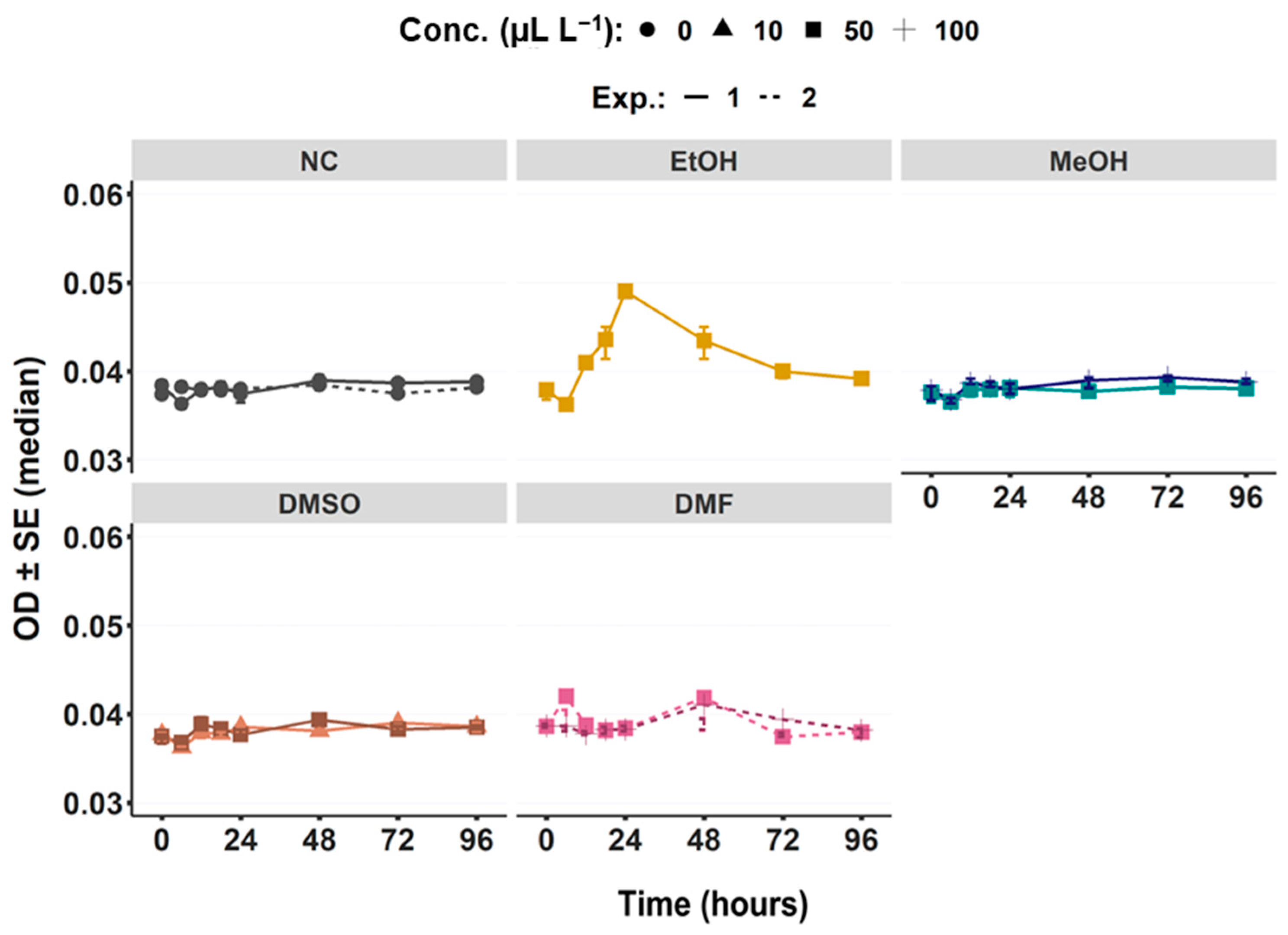
2.3. Water Quality
2.3.1. Water Parameters
2.3.2. Turbidity
2.4. Coral Health Indicators as Biological Responses at the Organismal and Cellular Levels
2.4.1. Organismal Level
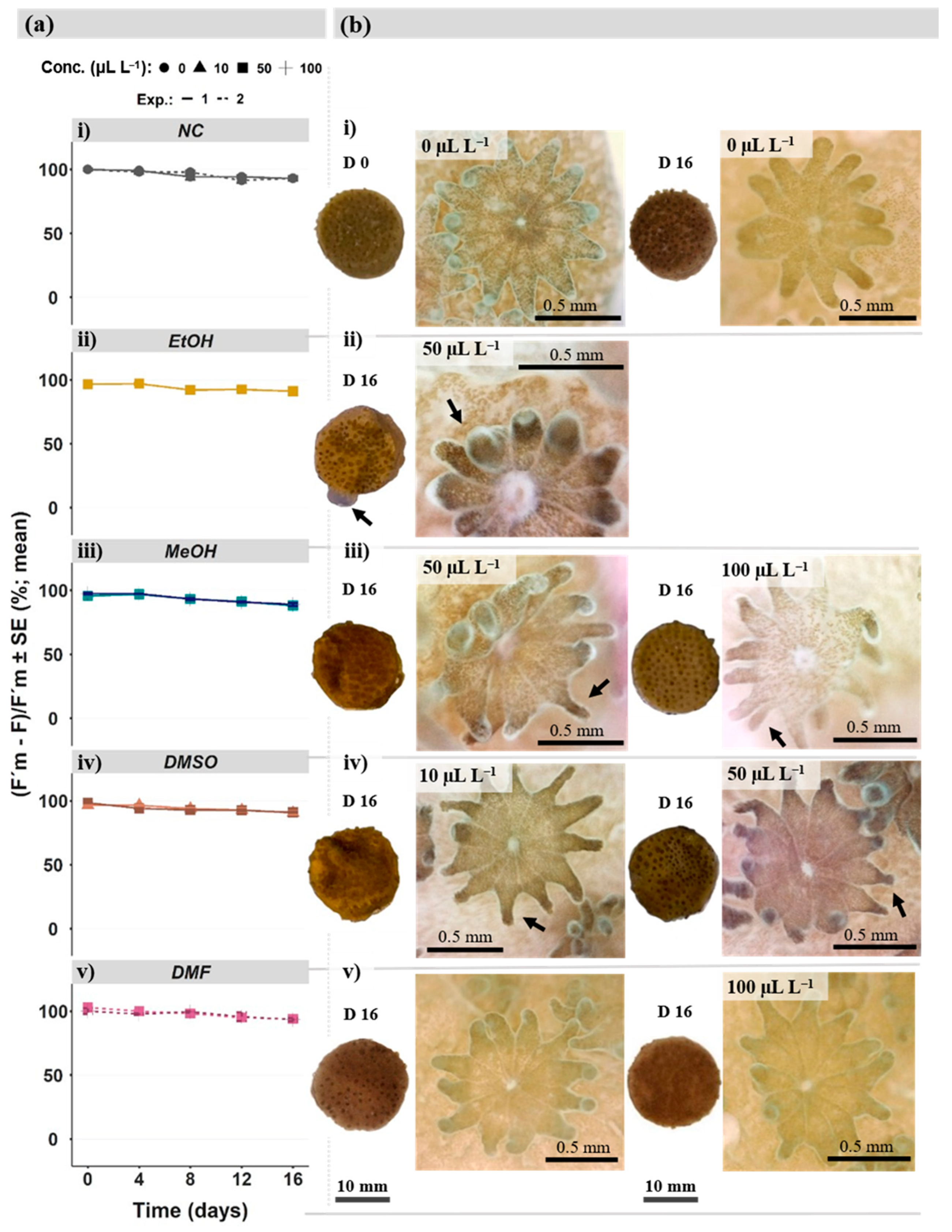
Photosynthetic Efficiency of Symbiotic Algae/Symbionts
Morphological Analysis
2.4.2. Cellular Level
Sample Preparation for Biomarker Analyses
- 150 µL for LPO, in which 2.5 µL butylated hydroxytoluene (BHT) was added.
- 170 µL for CAT, including 30 µL for total protein (Pr) quantification.
- 450 µL for CEA divided into:
- ○
- 150 µL for electron transport system (ETS) activity.
- ○
- 150 µL for carbohydrate and protein (CH/Pr) quantification.
- ○
- 150 µL for lipid (LIP) quantification.
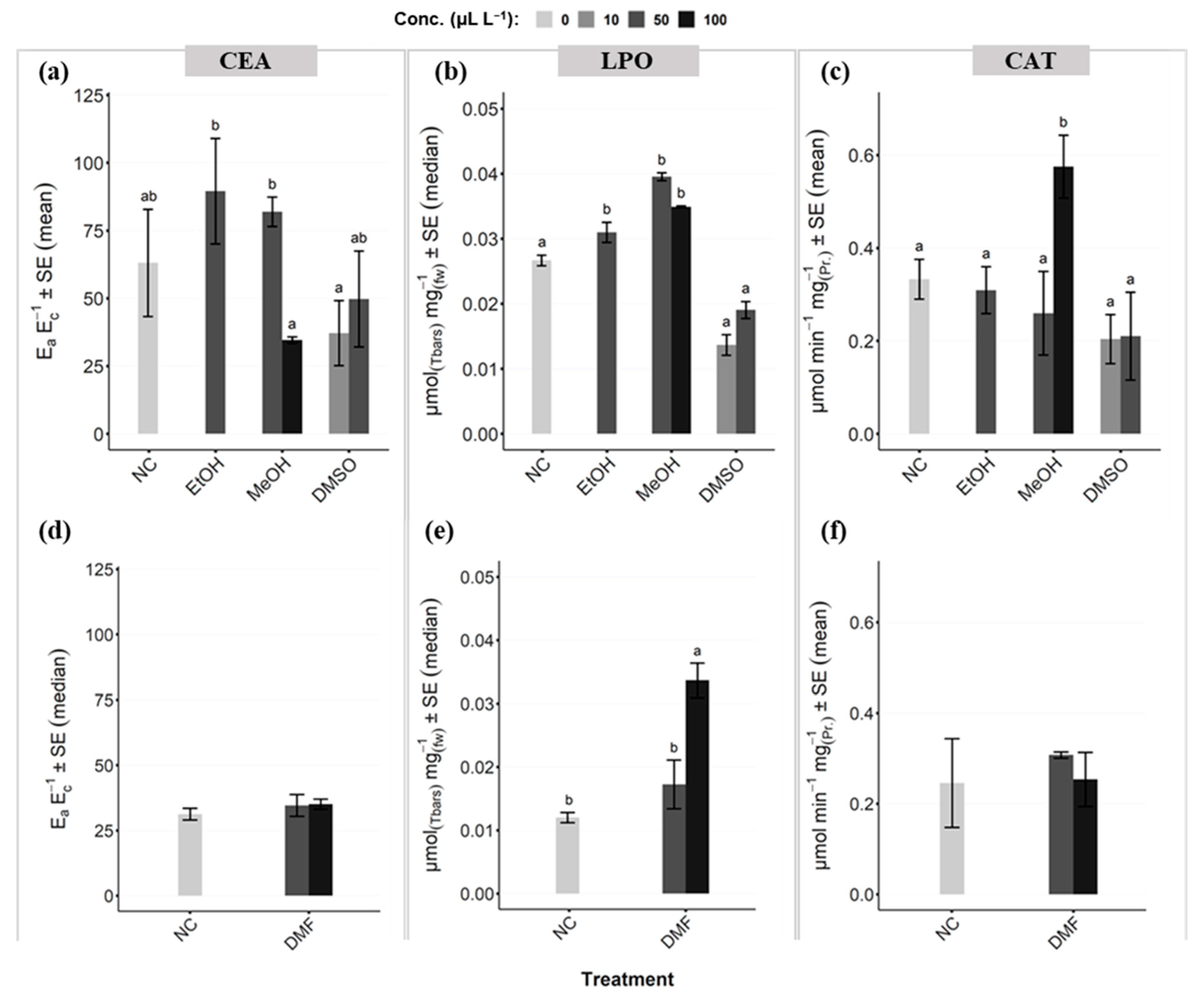
Oxidative Stress Responses (LPO; CAT)
Cellular Energy Allocation (CEA)
- Energy availability (Ea):
- Energy consumption (Ec):
2.5. Statistical Analyses
3. Results
3.1. Water Quality
3.1.1. Water Parameters
3.1.2. Water Turbidity via OD Measurement
3.2. Coral Health Indicator on Organismal Level
3.2.1. Photobiological Responses
3.2.2. Coral Fragment and Polyp Morphology
3.3. Coral Health Indicator on Cellular Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spalding, M.D.; Grenfell, A.M. New estimates of global and regional coral reef areas. Coral Reefs 1997, 16, 225–230. [Google Scholar] [CrossRef]
- Crossland, C.J.; Hatcher, B.G.; Smith, S.V. Role of coral reefs in global ocean production. Coral Reefs 1991, 10, 55–64. [Google Scholar] [CrossRef]
- Kayanne, H. Response of Coral Reefs to Global Warming. In Coral Reef Science-Strategy for Ecosystem Symbiosis and Coexistence with Humans under Multiple Stresses; Kayanne, H., Ed.; Springer: Tokyo, Japan, 2016. [Google Scholar]
- Cesar, H.S.J. Coral Reefs: Their Functions, Threats and Economic Value; Kalmar University: Kalmar, Sweden, 2002. [Google Scholar]
- Fabricius, K.; De’ath, G.; McCook, L.; Turak, E.; Williams, D.M. Changes in algal, coral and fish assemblages along water quality gradients on the inshore Great Barrier Reef. Mar. Pollut. Bull. 2005, 51, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.; Wilkinson, C. Global Threats to Coral Reefs: Coral Bleaching, Global Climate Change, Disease, Predator Plagues, and Invasive Species. In Status of Coral Reefs of the World; Wilkinson, C., Ed.; Australian Institute of Marine Science: Townsville, Australia, 2004; Volume 1. [Google Scholar]
- Hunter, C.L.; Evans, C.W. Coral Reefs in Kaneohe Bay, Hawaii: Two Centuries of Western Influence and Two Decades of Data. Bull. Mar. Sci. 1995, 57, 501–515. [Google Scholar]
- Álvarez-Muñoz, D.; Llorca, M.; Blasco, J.; Barceló, D. Contaminants in the Marine Environment. In Marine Ecotoxicology; Academic Press: Cambridge, MA, USA, 2016; pp. 1–34. [Google Scholar]
- Ouédraogo, D.-Y.; Mell, H.; Perceval, O.; Burga, K.; Domart-Coulon, I.; Hédouin, L.; Delaunay, M.; Guillaume, M.M.M.; Castelin, M.; Calvayrac, C.; et al. What are the toxicity thresholds of chemical pollutants for tropical reef-building corals? A systematic review. Environ. Evid. 2023, 12, 4. [Google Scholar] [CrossRef]
- Pawlowski, S.; Petersen-Thiery, M. Sustainable Sunscreens: A Challenge Between Performance, Animal Testing Ban, and Human and Environmental Safety. In Sunscreens in Coastal Ecosystems; The Handbook of Environmental Chemistry; Springer: Cham, Switzerland, 2020; pp. 185–207. [Google Scholar]
- Mitchelmore, C.L.; He, K.; Gonsior, M.; Hain, E.; Heyes, A.; Clark, C.; Younger, R.; Schmitt-Kopplin, P.; Feerick, A.; Conway, A.; et al. Occurrence and distribution of UV-filters and other anthropogenic contaminants in coastal surface water, sediment, and coral tissue from Hawaii. Sci. Total Environ. 2019, 670, 398–410. [Google Scholar] [CrossRef]
- Moeller, M.; Pawlowski, S.; Petersen-Thiery, M.; Miller, I.B.; Nietzer, S.; Heisel-Sure, Y.; Kellermann, M.Y.; Schupp, P.J. Challenges in Current Coral Reef Protection–Possible Impacts of UV Filters Used in Sunscreens, a Critical Review. Front. Mar. Sci. 2021, 8, 665548. [Google Scholar] [CrossRef]
- Green, J.; Wheeler, J.R. The use of carrier solvents in regulatory aquatic toxicology testing: Practical, statistical and regulatory considerations. Aquat. Toxicol. 2013, 14445, 242–249. [Google Scholar] [CrossRef]
- Hutchinson, T.H.; Shillabeer, N.; Winter, M.J.; Pickford, D.B. Acute and chronic effects of carrier solvents in aquatic organisms: A critical review. Aquat. Toxicol. 2006, 76, 69–92. [Google Scholar] [CrossRef]
- Okumura, J.K.H.T.H.S.Y. Influence of Organic Solvents on the Growth of Marine Microalgae. Arch. Environ. Contam. Toxicol. 2001, 41, 123–128. [Google Scholar] [CrossRef]
- OECD. Guidance document on aqueous-phase aquatic toxicity testing of difficult test chemicals. OECD Environ. Health Saf. Publ. 2019, 23, 3–81. [Google Scholar] [CrossRef]
- Machado, E.S.D.A.; Costa, A.P.L.; Rodrigues, A.C.M.; Bem-Haja, P.; Pires, S.S.F.; Soares, A.; Marques, C.R.; Pacheco, M.G.G.; Rocha, R.J.M. Organic solvents alter photo physiological and oxidative stress profiles of the coral Zoanthus sp.-Towards an optimization of ecotoxicological protocols. Sci. Total Environ. 2021, 777, 146072. [Google Scholar] [CrossRef]
- Roth, M.S. The engine of the reef: Photobiology of the coral-algal symbiosis. Front. Microbiol. 2014, 5, 422. [Google Scholar] [CrossRef]
- Warner, M.E.; Berry-Lowe, S. Differential xanthophyll cycling and photochemical activity in symbiotic dinoflagellates in multiple locations of three species of Caribbean coral. J. Exp. Mar. Biol. Ecol. 2006, 339, 86–95. [Google Scholar] [CrossRef]
- Jones, R.J.; Hoegh-Guldberg, O. Diurnal changes in the photochemical efficiency of the symbiotic dinoflagellates (Dinophyceae) of corals: Photoprotection, photoinactivation and the relationship to coral bleaching. Plant Cell Environ. 2001, 24, 89–99. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jones, R.J. Photoinhibition and photoprotection in symbiotic dinoflagellates from reef-building corals. Mar. Ecol. Prog. Ser. 1999, 183, 73–86. [Google Scholar] [CrossRef]
- He, T.; Tsui, M.M.P.; Tan, C.J.; Ng, K.Y.; Guo, F.W.; Wang, L.H.; Chen, T.H.; Fan, T.Y.; Lam, P.K.S.; Murphy, M.B. Comparative toxicities of four benzophenone ultraviolet filters to two life stages of two coral species. Sci. Total Environ. 2019, 651, 2391–2399. [Google Scholar] [CrossRef]
- Soffer, N.; Brandt, M.E.; Correa, A.M.; Smith, T.B.; Thurber, R.V. Potential role of viruses in white plague coral disease. ISME J. 2014, 8, 271–283. [Google Scholar] [CrossRef]
- Cervino, J.M.; Hayes, R.L.; Honovich, M.; Goreau, T.J.; Jones, S.; Rubec, P.J. Changes in zooxanthellae density, morphology, and mitotic index in hermatypic corals and anemones exposed to cyanide. Mar. Pollut. Bull. 2003, 46, 573–586. [Google Scholar] [CrossRef]
- Hidaka, M. Life History and Stress Response of Scleractinian Corals. In Coral Reef Science, 5th ed.; Kayanne, H., Ed.; Coral Reefs of the World 5; Springer: Tokyo, Japan, 2016; pp. 1–24. [Google Scholar]
- Downs, C.A.; Mueller, E.; Phillips, S.; Fauth, J.E.; Woodley, C.M. A molecular biomarker system for assessing the health of coral (Montastraea faveolata) during heat stress. Mar. Biotechnol. 2000, 2, 533–544. [Google Scholar] [CrossRef]
- Kühnhold, H.; Kamyab, E.; Novais, S.; Indriana, L.; Kunzmann, A.; Slater, M.; Lemos, M. Thermal stress effects on energy resource allocation and oxygen consumption rate in the juvenile sea cucumber, Holothuria scabra (Jaeger, 1833). Aquaculture 2017, 467, 109–117. [Google Scholar] [CrossRef]
- Rodrigues, A.P.; Oliva-Teles, T.; Mesquita, S.R.; Delerue-Matos, C.; Guimarães, L. Integrated biomarker responses of an estuarine invertebrate to high abiotic stress and decreased metal contamination. Mar. Environ. Res. 2014, 101, 101–114. [Google Scholar] [CrossRef]
- Lemos, M.F.L.; Soares, A.M.V.M.; Correia, A.C.; Esteves, A.C. Proteins in ecotoxicology-How, why and why not? Proteomics 2010, 10, 873–887. [Google Scholar] [CrossRef]
- Fel, J.-P.; Lacherez, C.; Bensetra, A.; Mezzache, S.; Béraud, E.; Léonard, M.; Allemand, D.; Ferrier-Pagès, C. Photochemical response of the scleractinian coral Stylophora pistillata to some sunscreen ingredients. Coral Reefs 2019, 38, 109–122. [Google Scholar] [CrossRef]
- Allison, N.; Cohen, I.; Finch, A.A.; Erez, J. Controls on Sr/Ca and Mg/Ca in scleractinian corals: The effects of Ca-ATPase and transcellular Ca channels on skeletal chemistry. Geochim. Et Cosmochim. Acta 2011, 75, 6350–6360. [Google Scholar] [CrossRef]
- Schnetger, B.; Lehners, C. Determination of nitrate plus nitrite in small volume marine water samples using vanadium (III) chloride as a reduction agent. Mar. Chem. 2014, 160, 91–98. [Google Scholar] [CrossRef]
- Kitchener, B.G.B.; Wainwright, J.; Parsons, A.J. A review of the principles of turbidity measurement. Prog. Phys. Geogr. Earth Environ. 2017, 41, 620–642. [Google Scholar] [CrossRef]
- Walz. MINI-PAM-II-Manual for Standalone Use; Heinz Walz GmbH: Reutlingen/Pfullingen, Germany, 2018. [Google Scholar]
- Matyssek, R.; Herppich, W.B. Experimentelle Pflanzenökologie: Chlorophyllfluoreszenzanalyse. In Handbuch der Geodäsie; Springer Reference Naturwissenschaften; Springer Spektrum: Berlin/Heidelberg, Germany, 2018; pp. 1–56. [Google Scholar]
- Casereto, B.E.; Suzuki, T.; Susuki, Y. Chemical and Biological Characteristics of Coral Reef Ecosystem at Microscale/Nanoscale: Effect of Multiple and Synergistic Stresses. In Coral Reef Science-Strategy for Ecosystem Symbiosis and Coexistence with Humans under Multiple Stresses; Kayanne, H., Ed.; Springer: Tokyo, Japan, 2016; Volume 5. [Google Scholar]
- Cooper, T.F.; Fabricius, K.E. Pigmentation of massive corals as a simple bioindicator for marine water quality. Mar. Pollut. Bull. 2012, 65, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.; Madeira, C.; Jogee, N.; Ferreira, A.; Gouveia, R.; Cabral, H.; Diniz, M.; Vinagre, C. Integrative indices for health assessment in reef corals under thermal stress. Ecol. Indic. 2020, 113, 106230. [Google Scholar] [CrossRef]
- Alves, L.M.F.; Nunes, M.; Marchand, P.; Le Bizec, B.; Mendes, S.; Correia, J.P.S.; Lemos, M.F.L.; Novais, S.C. Blue sharks (Prionace glauca) as bioindicators of pollution and health in the Atlantic Ocean: Contamination levels and biochemical stress responses. Sci. Total Environ. 2016, 563–564, 282–292. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P.; Draper, H.H. Comparative studies on different methods of malonaldehyde determination. Methods Enzymol. 1984, 105, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Aloísio Torres, M.; Pires Testa, C.; Gáspari, C.; Beatriz Masutti, M.; Maria Neves Panitz, C.; Curi-Pedrosa, R.; Alves de Almeida, E.; Di Mascio, P.; Wilhelm Filho, D. Oxidative stress in the mussel Mytella guyanensis from polluted mangroves on Santa Catarina Island, Brazil. Mar. Pollut. Bull. 2002, 44, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A. Catalase Activity, 1st ed.; CRC Press Inc: Boca Raton, FL, USA, 1984. [Google Scholar]
- Bradford, M.M. Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 24, 248–254. [Google Scholar] [CrossRef]
- DeCoen, W.M.; Janssen, C.R. The Missing Biomarker Link: Relationships between Effects on The Cellular Energy Allocation Biomarker of Toxicant-Stressed Daphnia Magna and Corresponding Population Characteristics. Environ. Toxicol. Chem. 2003, 22, 1632–1641. [Google Scholar] [CrossRef]
- Gnaiger, E. Calculation of Energetic and Biochemical Equivalents of Respiratory Oxygen Consumption. In Polarographic Oxygen Sensors; Springer: Berlin/Heidelberg, Germany, 1983; pp. 337–345. [Google Scholar]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR). R Package 2014, 27, 1–9. [Google Scholar]
- Dinno, A. Conover-Iman Test of Multiple Comparisons Using Rank Sums. R Package Version 1.1.5. 2017. Available online: https://cran.r-project.org/web/packages/conover.test/conover.test.pdf (accessed on 2 April 2023).
- Dinno, A. Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf (accessed on 2 April 2023).
- Wickham, H. An Introduction to ggplot: An Implementation of the Grammar of Graphics in R. Statistics. 2006, pp. 1–8. Available online: https://www.semanticscholar.org/paper/ggplot%3A-An-implementation-of-the-Grammar-of-in-R-Wickham/7f3e2207d2ef8fc0cee74069879c8adf35303a91 (accessed on 2 April 2023).
- Borneman, E. Introduction to the husbandry of corals in aquariums: A review. Public Aquar. Husb. Ser. 2008, 2, 3–14. [Google Scholar]
- Yan, X.; Guoxing, S.; Yu, D. Effect of N,N-Dimethyl Formamide Used as Organic Solvent on Two Species of Green Algae Chlorella. Bull. Environ. Contam. Toxicol. 2002, 68, 592–599. [Google Scholar] [CrossRef]
- Leblanc, G.A.; Surprenant, D.C. The acute and chronic toxicity of acetone, dimethyl formamide, and triethylene glycol to Daphnia magna (Straus). Arch. Environ. Contam. Toxicol. 1983, 12, 305–310. [Google Scholar] [CrossRef]
- Girotti, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- Pannunzio, T.M.; Storey, K.B. Antioxidant defenses and lipid peroxidation during anoxia stress and aerobic recovery in the marine gastropod Littorina littorea. J. Exp. Mar. Biol. Ecol. 1998, 221, 227–292. [Google Scholar] [CrossRef]
- Jyothi, K.; Kalyani, D.; Nachiappan, V. Effect of acute exposure of N, N-Dimethylformamide, an industrial solvent on lipid peroxidation and antioxidants in liver and kidney of rats. Indian J. Biochem. Biophys. 2012, 49, 279–284. [Google Scholar] [PubMed]
- Jarosz, T.; Weltje, L. Untersuchungen zur akuten und chronischen Toxizität organischer Lösemittel für die Substanztestung in der aquatischen Ökotoxikologie. In Proceedings of the 3. Gemeinsame Jahrestagung SETAC-GLB und GDCh, Frankfurt/Main, Germany, 23–26 September 2008. [Google Scholar]
- Yu, Z.W.; Quinn, P.J. Dimethyl sulphoxide: A review of its applications in cell biology. Biosci. Rep. 1994, 14, 259–281. [Google Scholar] [CrossRef] [PubMed]
- Calesnick, B.; Dinan, A. Pharmakologie und Toxikologie von Dimethylsulfoxid (DMSO). In DMSO; Jacob, S., Herschler, R.J., Schmellenkamp, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1985. [Google Scholar]
- van Hoek, A.N.; de Jong, M.D.; van Os, C.H. Effects of dimethylsulfoxide and mercurial sulfhydryl reagents on water and solute permeability of rat kidney brush border membranes. Biochim. Et Biophys. Acta (BBA) Biomembr. 1990, 1030, 203–210. [Google Scholar] [CrossRef]
- Sakharov, B.V.; Volkov, V. The effect of dimethyl sulfoxide and thiourea on the water diffusion permeability of the E. coli membrane. Biofizika 1989, 34, 464–467. [Google Scholar]
- Deschaseaux, E.S.M.; Jones, G.B.; Deseo, M.A.; Shepherd, K.M.; Kiene, R.P.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Effects of environmental factors on dimethylated sulfur compounds and their potential role in the antioxidant system of the coral holobiont. Limnol. Oceanogr. 2014, 59, 758–768. [Google Scholar] [CrossRef]
- Keller, M.D.; Kiene, R.P.; Matrai, P.A.; Bellows, W.K. Production of glycine betaine and dimethylsulfoniopropionate in marine phytoplankton. I. Batch cultures. Mar. Biol. 1999, 135, 237–248. [Google Scholar] [CrossRef]
- Klein, S.M.; Cohen, G.; Cederbaum, A.I. Production of Formaldehyde during Metabolism of Dimethyl Sulfoxide by Hydroxyl Radical Generating Systems. Biochemistry 1981, 20, 6006–6012. [Google Scholar] [CrossRef]
- Cowgill, U.M.; Milazzo, D.P. The Sensitivity of Ceriodaphnia dubia and Daphnia magna to seven chemicals utilizing the three-brood test. Arch. Environ. Contam. Toxicol. 1991, 20, 211–217. [Google Scholar] [CrossRef]
- Rayburn, J.R.; Fisher, W.S. Developmental Toxicity of Three Carrier Solvents Using Embryos of the Grass Shrimp, Palaemonetes pugio. Arch. Environ. Contam. Toxicol. 1997, 33, 217–221. [Google Scholar] [CrossRef]
- Buhl-Mortensen, P.; Braga-Henriques, A.; Stevenson, A. Polyp loss and mass occurrence of sea urchins on bamboo corals in the deep sea: An indirect effect of fishing impact? Ecology 2022, 103, e03564. [Google Scholar] [CrossRef]
- Smith, M.G.; Des Etages, S.G.; Snyder, M. Microbial Synergy via an Ethanol-Triggered Pathway. Mol. Cell. Biol. 2004, 24, 3874–3884. [Google Scholar] [CrossRef] [PubMed]
- de Bruyn, W.J.; Clark, C.D.; Senstad, M.; Toms, N.; Harrison, A.W. Biological degradation of ethanol in Southern California coastal seawater. Mar. Chem. 2020, 218, 103703. [Google Scholar] [CrossRef]
- Gluth, G.; Hanke, W. A comparison of physiological changes in carp, Cyprinus carpio, induced by several pollutants at sublethal concentrations: I. The dependency on exposure time. Ecotoxicol. Environ. Saf. 1985, 9, 179–188. [Google Scholar] [CrossRef]
- Corpe, W.A. A method for detecting methylotrophic bacteria on solid surfaces. J. Microbiol. Methods 1985, 3, 215–221. [Google Scholar] [CrossRef]
- Dinsdale, E.A.; Pantos, O.; Smriga, S.; Edwards, R.A.; Angly, F.; Wegley, L.; Hatay, M.; Hall, D.; Brown, E.; Haynes, M.; et al. Microbial Ecology of Four Coral Atolls in the Northern Line Islands. PLoS ONE 2008, 3, e1584. [Google Scholar] [CrossRef]
- Costa, A.P.L.; Silva, D.A.M.; Rodrigues, A.C.M.; Marques, C.R.; Soares, A.M.V.M.; Rocha, R.J.M. Species-specific oxidative stress responses and cellular energy allocation after coral shipping. Aquac. Rep. 2021, 19, 100623. [Google Scholar] [CrossRef]
- Cadenas, E.; Sies, H. The lag phase. Free Radic. Res. 1998, 28, 601–609. [Google Scholar] [CrossRef]
- Yakovleva, I.; Bhagooli, R.; Takemura, A.; Hidaka, M. Differential susceptibility to oxidative stress of two scleractinian corals: Antioxidant functioning of mycosporine-glycine. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 721–730. [Google Scholar] [CrossRef]
- Zhang, T.; Qu, Y.; Zhang, Q.; Tang, J.; Cao, R.; Dong, Z.; Wang, Q.; Zhao, J. Risks to the stability of coral reefs in the South China Sea: An integrated biomarker approach to assess the physiological responses of Trochus niloticus to ocean acidification and warming. Sci. Total Environ. 2021, 782, 146876. [Google Scholar] [CrossRef]
- Kim, J.; Park, Y.; Choi, K. Phototoxicity and oxidative stress responses in Daphnia magna under exposure to sulfathiazole and environmental level ultraviolet B irradiation. Aquat. Toxicol. 2009, 91, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Anithajothi, R.; Duraikannu, K.; Umagowsalya, G.; Ramakritinan, C.M. The Presence of Biomarker Enzymes of Selected Scleractinian Corals of Palk Bay, Southeast Coast of India. BioMed Res. Int. 2014, 2014, 684874. [Google Scholar] [CrossRef] [PubMed]
- Krueger, T.; Hawkins, T.D.; Becker, S.; Pontasch, S.; Dove, S.; Hoegh-Guldberg, O.; Leggat, W.; Fisher, P.L.; Davy, S.K. Differential coral bleaching—Contrasting the activity and response of enzymatic antioxidants in symbiotic partners under thermal stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 190, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Rosset, S.; Wiedenmann, J.; Reed, A.J.; D′Angelo, C. Phosphate deficiency promotes coral bleaching and is reflected by the ultrastructure of symbiotic dinoflagellates. Mar. Pollut. Bull. 2017, 118, 180–187. [Google Scholar] [CrossRef]
- Allen, A.E.; Booth, M.G.; Frischer, M.E.; Verity, P.G.; Zehr, J.P.; Zani, S. Diversity and Detection of Nitrate Assimilation Genes in Marine Bacteria. Appl. Environ. Microbiol. 2001, 67, 5343–5348. [Google Scholar] [CrossRef]
- Godinot, C.; Ferrier-Pagès, C.; Grover, R. Control of phosphate uptake by zooxanthellae and host cells in the scleractinian coral Stylophora pistillata. Limnol. Oceanogr. 2009, 54, 1627–1633. [Google Scholar] [CrossRef]
- Fosså, S.A.; Nilsen, A.J. Das Korallenriff-Aquarium; Natur und Tier Verlag GmbH: Münster, Germany, 2013. [Google Scholar]
- TropicMarin. Supply System. Available online: https://www.tropic-marin.com/versorgungssysteme?lang=en (accessed on 2 March 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Conc. | Total nr. of Fragments | Paleness | Polyp Losses | Tissue Swelling | |||
|---|---|---|---|---|---|---|---|---|
| µL L−1 | TN | n | % of TN | n | % of TN | n | % of TN | |
| NC | 0 | 24 | 7 | 30 | 1 | 4 | 0 | 0 |
| EtOH | 50 | 12 | 7 | 58 | 6 | 50 | 8 | 66 |
| MeOH | 50 | 12 | 6 | 50 | 5 | 41 | 0 | 0 |
| 100 | 12 | 9 | 75 | 5 | 41 | 4 | 33 | |
| DMSO | 10 | 12 | 9 | 75 | 3 | 25 | 1 | 8 |
| 50 | 12 | 8 | 66 | 6 | 50 | 1 | 8 | |
| DMF | 50 | 12 | 2 | 16 | 2 | 16 | 0 | 0 |
| 100 | 12 | 3 | 25 | 2 | 16 | 0 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Mauro, V.; Kamyab, E.; Kellermann, M.Y.; Moeller, M.; Nietzer, S.; Luetjens, L.H.; Pawlowski, S.; Petersen-Thiery, M.; Schupp, P.J. Ecotoxicological Effects of Four Commonly Used Organic Solvents on the Scleractinian Coral Montipora digitata. Toxics 2023, 11, 367. https://doi.org/10.3390/toxics11040367
Di Mauro V, Kamyab E, Kellermann MY, Moeller M, Nietzer S, Luetjens LH, Pawlowski S, Petersen-Thiery M, Schupp PJ. Ecotoxicological Effects of Four Commonly Used Organic Solvents on the Scleractinian Coral Montipora digitata. Toxics. 2023; 11(4):367. https://doi.org/10.3390/toxics11040367
Chicago/Turabian StyleDi Mauro, Valentina, Elham Kamyab, Matthias Y. Kellermann, Mareen Moeller, Samuel Nietzer, Laura H. Luetjens, Sascha Pawlowski, Mechtild Petersen-Thiery, and Peter J. Schupp. 2023. "Ecotoxicological Effects of Four Commonly Used Organic Solvents on the Scleractinian Coral Montipora digitata" Toxics 11, no. 4: 367. https://doi.org/10.3390/toxics11040367