
Contaminants of Emerging Concern (CECs) and Male Reproductive Health: Challenging the Future with a Double-Edged Sword
 , ,
, ,
Abstract
:1. Introduction
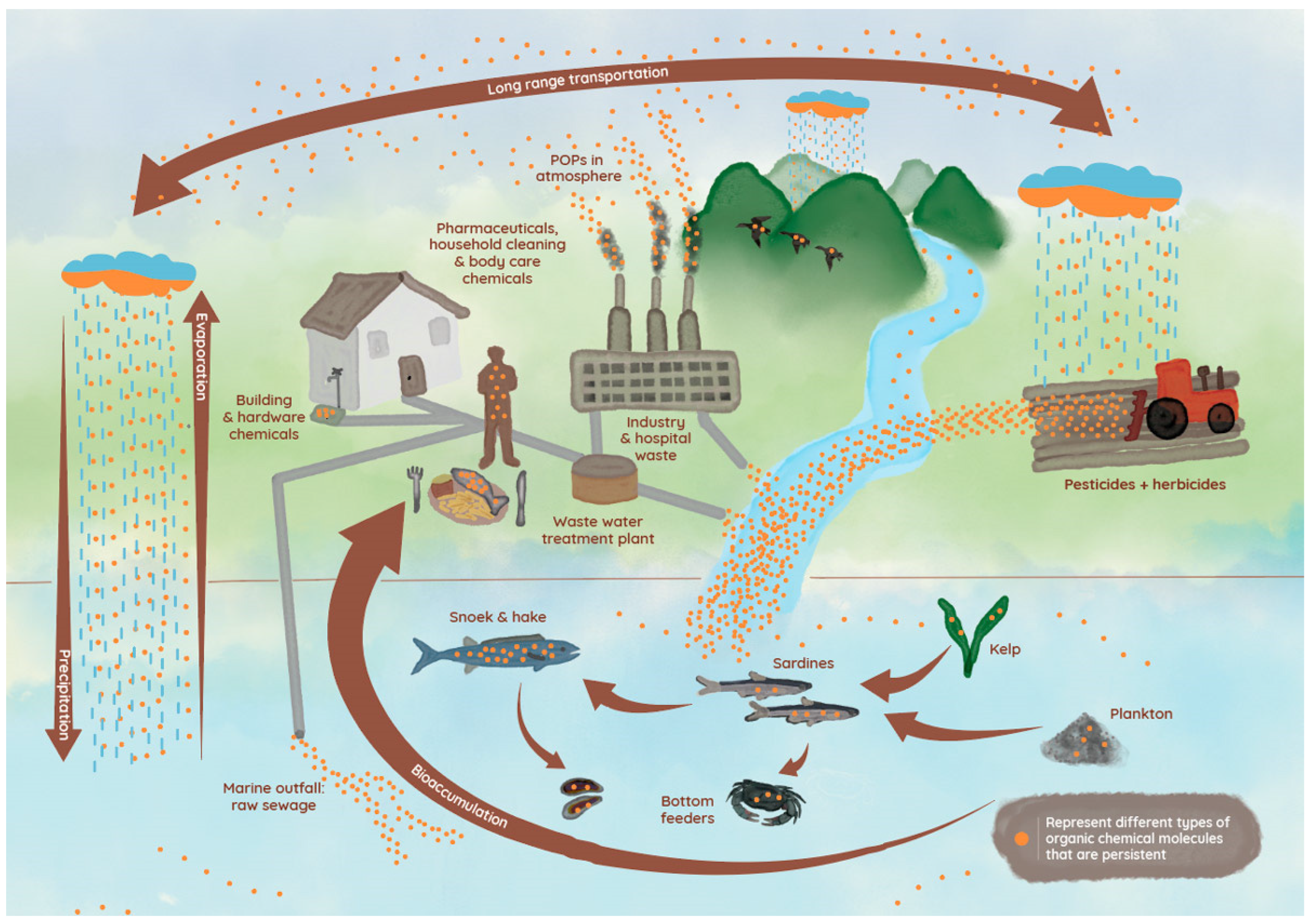
2. Origin and Distribution of CECs
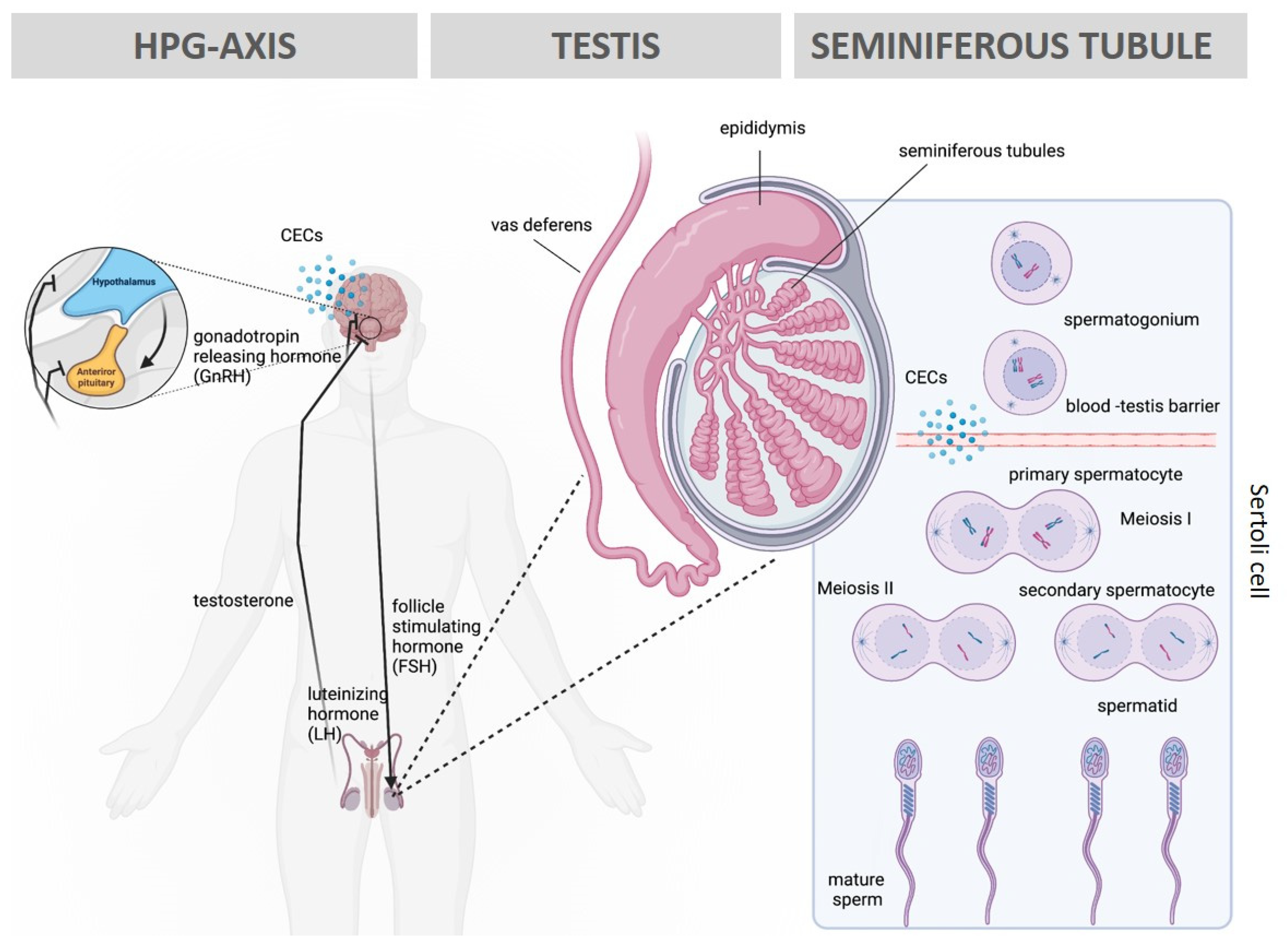
3. Impact of CECs on Male Fertility
3.1. Pesticides
3.1.1. Atrazine
3.1.2. Simazine
3.1.3. Chlorpyrifos
3.2. Pharmaceuticals
3.2.1. Sulfamethoxazole
3.2.2. Non-Steroidal Anti-Inflammatory Drugs
4. Impact of CECs on Offspring and Potential Mechanisms
5. Management, Detection, and Possible Treatment of CEC Exposures
6. Role of Spermatozoa in Toxicology Applications
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matzuk, M.M.; Lamb, D.J. The biology of infertility: Research advances and clinical challenges. Nat. Med. 2008, 14, 1197–1213. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Notariale, R.; Rabbito, D.; Ausió, J.; Olanrewaju, O.S.; Guerriero, G. Mytilus galloprovincialis (Lamarck, 1819) spermatozoa: Hsp70 expression and protamine-like protein property studies. Environ. Sci. Pollut. Res. 2018, 25, 12957–12966. [Google Scholar] [CrossRef] [PubMed]
- US Environmental Protection Agency [EPA]. Contaminants of Emerging Concern Including Pharmaceuticals and Personal Care Products. 2022. Available online: https://www.epa.gov/wqc/contaminants-emerging-concern-including-pharmaceuticals-and-personal-care-products (accessed on 23 January 2023).
- Khan, J.A.; Sayed, M.; Khan, S.; Shah, N.S.; Dionysiou, D.D.; Boczkaj, G. Advanced oxidation processes for the treatment of contaminants of emerging concern. In Contaminants of Emerging Concern in Water and Wastewater; Hernández-Maldonado, A.J., Blaney, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 299–365. [Google Scholar] [CrossRef]
- Naidu, R.; Biswas, B.; Willett, I.R.; Cribb, J.; Singh, B.K.; Nathanail, C.P.; Coulon, F.; Semple, K.T.; Jones, K.C.; Barclay, A.; et al. Chemical pollution: A growing peril and potential catastrophic risk to humanity. Environ. Int. 2021, 1, 106616. [Google Scholar] [CrossRef]
- Colborn, T.; Smolen, M.J. Epidemiological analysis of persistent organochlorine contaminants in cetaceans. In Reviews of Environmental Contamination and Toxicology; Ware, G.W., Ed.; Springer: New York, NY, USA, 1996; Volume 146, pp. 91–172. [Google Scholar] [CrossRef]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.; Overstreet, R.M.; et al. Emerging marine diseases—Climate links and anthropogenic factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef] [Green Version]
- Rivetti, I.; Fraschetti, S.; Lionello, P.; Zambianchi, E.; Boero, F. Global warming and mass mortalities of benthic invertebrates in the Mediterranean Sea. PLoS ONE 2014, 9, e115655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthilselvan, A.; Mcduffie, H.H.; Dosman, J.A. Association of asthma with use of pesticides. Am. Rev. Respir. Dis. 1992, 146, 884–887. [Google Scholar] [CrossRef]
- Vial, T.; Nicolas, B.; Descotes, J. Clinical immunotoxicity of pesticides. Toxicol. Environ. Health Sci. 1996, 48, 215–229. [Google Scholar] [CrossRef]
- McBride, M.L. Childhood cancer and environmental contaminants. Can. J. Public Health 1998, 89, S58. [Google Scholar] [CrossRef]
- Foster, W.G.; Neal, M.S.; Han, M.S.; Dominguez, M.M. Environmental contaminants and human infertility: Hypothesis or cause for concern? Toxicol. Environ. Health Sci. Part B 2008, 11, 162–176. [Google Scholar] [CrossRef]
- Nateghian, Z.; Aliabadi, E. Aspects of environmental pollutants on male fertility and sperm parameters. J. Environ. Treat. Tech. 2020, 8, 299–309. [Google Scholar]
- Fainberg, J.; Kashanian, J.A. Recent advances in understanding and managing male infertility. F1000Research 2019, 8, 670. [Google Scholar] [CrossRef] [PubMed]
- Alahmar, A.T. Role of oxidative stress in male infertility: An updated review. J. Hum. Reprod. Sci. 2019, 12, 4. [Google Scholar] [CrossRef]
- Gabrielsen, J.S.; Tanrikut, C. Chronic exposures and male fertility: The impacts of environment, diet, and drug use on spermatogenesis. Andrology 2016, 4, 648–661. [Google Scholar] [CrossRef]
- Pizzol, D.; Foresta, C.; Garolla, A.; Demurtas, J.; Trott, M.; Bertoldo, A.; Smith, L. Pollutants and sperm quality: A systematic review and meta-analysis. Environ. Sci. Pollut. Res. 2021, 28, 4095–4103. [Google Scholar] [CrossRef]
- Chen, T.; Belladelli, F.; Del Giudice, F.; Eisenberg, M.L. Male fertility as a marker for health. Reprod 2022, 44, 131–144. [Google Scholar] [CrossRef]
- García-Fernández, A.J.; Espín, S.; Gómez-Ramírez, P.; Sánchez-Virosta, P.; Navas, I. Water Quality and Contaminants of Emerging Concern (CECs). In Chemometrics and Cheminformatics in Aquatic Toxicology; Roy, K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2021; Volume 13, pp. 1–21. [Google Scholar] [CrossRef]
- Céspedes, R.; Lacorte, S.; Ginebreda, A.; Barceló, D. Chemical monitoring and occurrence of alkylphenols, alkylphenol ethoxylates, alcohol ethoxylates, phthalates and benzothiazoles in sewage treatment plants and receiving waters along the Ter River basin (Catalonia, NE Spain). Anal. Bioanal. Chem. 2006, 385, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Morteani, G.; Möller, P.; Fuganti, A.; Paces, T. Input and fate of anthropogenic estrogens and gadolinium in surface water and sewage plants in the hydrological basin of Prague (Czech Republic). Environ. Geochem. Health 2006, 28, 257–264. [Google Scholar] [CrossRef]
- Kasprzyk-Hordern, B.; Dinsdale, R.M.; Guwy, A.J. The occurrence of pharmaceuticals, personal care products, endocrine disruptors and illicit drugs in surface water in South Wales, UK. Water Res. 2008, 42, 3498–3518. [Google Scholar] [CrossRef] [PubMed]
- Kuster, M.; de Alda, M.J.; Hernando, M.D.; Petrovic, M.; Martín-Alonso, J.; Barceló, D. Analysis and occurrence of pharmaceuticals, estrogens, progestogens and polar pesticides in sewage treatment plant effluents, river water and drinking water in the Llobregat river basin (Barcelona, Spain). J. Hydrol. 2008, 358, 112–123. [Google Scholar] [CrossRef]
- Zuccato, E.; Castiglioni, S.; Bagnati, R.; Chiabrando, C.; Grassi, P.; Fanelli, R. Illicit drugs, a novel group of environmental contaminants. Water Res. 2008, 42, 961–968. [Google Scholar] [CrossRef]
- Benotti, M.J.; Trenholm, R.A.; Vanderford, B.J.; Holady, J.C.; Stanford, B.D.; Snyder, S.A. Pharmaceuticals and endocrine disrupting compounds in US drinking water. Environ. Sci. Technol. 2009, 43, 597–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojemaye, C.Y.; Onwordi, C.T.; Pampanin, D.M.; Sydnes, M.O.; Petrik, L. Presence and risk assessment of herbicides in the marine environment of Camps Bay (Cape Town, South Africa). Sci. Total Environ. 2020, 738, 140346. [Google Scholar] [CrossRef] [PubMed]
- Ojemaye, C.Y.; Onwordi, C.T.; Pampanin, D.M.; Sydnes, M.O.; Petrik, L. Herbicides in Camps Bay (Cape Town, South Africa), supplemented. Sci. Total Environ. 2021, 778, 146057. [Google Scholar] [CrossRef] [PubMed]
- Ojemaye, C.Y.; Petrik, L. Pharmaceuticals and personal care products in the marine environment around False Bay, Cape Town, South Africa: Occurrence and risk—Assessment study. Environ. Toxicol. Chem. 2022, 41, 614–634. [Google Scholar] [CrossRef]
- Petrik, L.; Green, L.; Abegunde, A.P.; Zackon, M.; Sanusi, C.Y.; Barnes, J. Desalination and seawater quality at Green Point, Cape Town: A study on the effects of marine sewage outfalls. S. Afr. J. Sci. 2017, 113, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Curchod, L.; Oltramare, C.; Junghans, M.; Stamm, C.; Dalvie, M.A.; Röösli, M.; Fuhrimann, S. Temporal variation of pesticide mixtures in rivers of three agricultural watersheds during a major drought in the Western Cape, South Africa. Water Res. X 2020, 6, 100039. [Google Scholar] [CrossRef]
- Chow, R.; Curchod, L.; Davies, E.; Veludo, A.F.; Oltramare, C.; Dalvie, M.A.; Stamm, C.; Röösli, M.; Fuhrimann, S. Seasonal drivers and risks of aquatic pesticide pollution in drought and post-drought conditions in three Mediterranean watersheds. Sci. Total Environ. 2023, 858, 159784. [Google Scholar] [CrossRef]
- Fuhrimann, S.; Mol, H.G.; Dias, J.; Dalvie, M.A.; Röösli, M.; Degrendele, C.; Figueiredo, D.M.; Huss, A.; Portengen, L.; Vermeulen, R. Quantitative assessment of multiple pesticides in silicone wristbands of children/guardian pairs living in agricultural areas in South Africa. Sci. Total Environ. 2022, 812, 152330. [Google Scholar] [CrossRef]
- Veludo, A.F.; Figueiredo, D.M.; Degrendele, C.; Masinyana, L.; Curchod, L.; Kohoutek, J.; Kukučka, P.; Martiník, J.; Přibylová, P.; Klánová, J.; et al. Seasonal variations in air concentrations of 27 organochlorine pesticides (OCPs) and 25 current-use pesticides (CUPs) across three agricultural areas of South Africa. Chemosphere 2022, 289, 133162. [Google Scholar] [CrossRef]
- Degrendele, C.; Prokeš, R.; Šenk, P.; Jílková, S.R.; Kohoutek, J.; Melymuk, L.; Přibylová, P.; Dalvie, M.A.; Röösli, M.; Klánová, J.; et al. Human exposure to pesticides in dust from two agricultural sites in South Africa. Toxics 2022, 10, 629. [Google Scholar] [CrossRef] [PubMed]
- Fuhrimann, S.; Klánová, J.; Přibylová, P.; Kohoutek, J.; Dalvie, M.A.; Röösli, M.; Degrendele, C. Qualitative assessment of 27 current-use pesticides in air at 20 sampling sites across Africa. Chemosphere 2020, 258, 127333. [Google Scholar] [CrossRef]
- Česen, M.; Ahel, M.; Terzić, S.; Heath, D.J.; Heath, E. The occurrence of contaminants of emerging concern in Slovenian and Croatian wastewaters and receiving Sava River. Sci. Total Environ. 2019, 650, 2446–2453. [Google Scholar] [CrossRef]
- K’oreje, K.O.; Okoth, M.; Van Langenhove, H.; Demeestere, K. Occurrence and treatment of contaminants of emerging concern in the African aquatic environment: Literature review and a look ahead. J. Environ. Manag. 2020, 254, 109752. [Google Scholar] [CrossRef]
- Nilsen, E.; Smalling, K.L.; Ahrens, L.; Gros, M.; Miglioranza, K.S.; Picó, Y.; Schoenfuss, H.L. Critical review: Grand challenges in assessing the adverse effects of contaminants of emerging concern on aquatic food webs. Environ. Toxicol. Chem. 2019, 38, 46–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salimi, M.; Esrafili, A.; Gholami, M.; Jonidi Jafari, A.; Rezaei Kalantary, R.; Farzadkia, M.; Kermani, M.; Sobhi, H.R. Contaminants of emerging concern: A review of new approach in AOP technologies. Environ. Monit. Assess. 2017, 189, 414. [Google Scholar] [CrossRef]
- Pico, Y.; Belenguer, V.; Corcellas, C.; Díaz-Cruz, M.S.; Eljarrat, E.; Farré, M.; Gago-Ferrero, P.; Huerta, B.; Navarro-Ortega, A.; Petrovic, M.; et al. Contaminants of emerging concern in freshwater fish from four Spanish Rivers. Sci. Total Environ. 2019, 659, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Picó, Y.; Alvarez-Ruiz, R.; Alfarhan, A.H.; El-Sheikh, M.A.; Alobaid, S.M.; Barceló, D. Uptake and accumulation of emerging contaminants in soil and plant treated with wastewater under real-world environmental conditions in the Al Hayer area (Saudi Arabia). Sci. Total Environ. 2019, 652, 562–572. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E. Bioaccumulation of non-essential hazardous heavy metals and metalloids in freshwater fish. Risk to human health. Environ. Chem. Lett. 2018, 16, 903–917. [Google Scholar] [CrossRef]
- Du, B.; Haddad, S.P.; Luek, A.; Scott, W.C.; Saari, G.N.; Burket, S.R.; Breed, C.S.; Kelly, M.; Broach, L.; Rasmussen, J.B.; et al. Bioaccumulation of human pharmaceuticals in fish across habitats of a tidally influenced urban bayou. Environ. Toxicol. Chem. 2016, 35, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.M.; Vaishnavi, C. Bioaccumulative potential and toxicity of endosulfan insecticide to non-target animals. Comp. Biochem. Physiol. Part—C Toxicol. Pharmacol. 1993, 105, 347–361. [Google Scholar] [CrossRef]
- Diamond, K.M.; Good, C.J.; Johnny, N.; Sakihara, T.S.; Edmiston, P.L.; Faust, J.A.; Schoenfuss, T.C.; Rubin, A.M.; Blob, R.W.; Schoenfuss, H.L. Assessing Occurrence and Biological Consequences of Contaminants of Emerging Concern on Oceanic Islands. Water 2022, 14, 275. [Google Scholar] [CrossRef]
- Vandermeersch, G.; Lourenço, H.M.; Alvarez-Muñoz, D.; Cunha, S.; Diogène, J.; Cano-Sancho, G.; Sloth, J.J.; Kwadijk, C.; Barcelo, D.; Allegaert, W.; et al. Environmental contaminants of emerging concern in seafood–European database on contaminant levels. Environ. Res. 2015, 143, 29–45. [Google Scholar] [CrossRef] [PubMed]
- James, C.A.; Lanksbury, J.; Khangaonkar, T.; West, J. Evaluating exposures of bay mussels (Mytilus trossulus) to contaminants of emerging concern through environmental sampling and hydrodynamic modeling. Sci. Total Environ. 2020, 709, 136098. [Google Scholar] [CrossRef]
- Meador, J.P.; Yeh, A.; Young, G.; Gallagher, E.P. Contaminants of emerging concern in a large temperate estuary. Environ. Pollut. 2016, 213, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Tototzintle, M.; Ferreira, I.J.; da Silva Duque, S.; Barrocas, P.R.G.; Saggioro, E.M. Removal of contaminants of emerging concern (CECs) and antibiotic resistant bacteria in urban wastewater using UVA/TiO2/H2O2 photocatalysis. Chemosphere 2018, 210, 449–457. [Google Scholar] [CrossRef]
- Kolesnikova, L.I.; Kolesnikov, S.I.; Kurashova, N.A.; Bairova, T.A. Causes and factors of male infertility. Ann. Russ. Acad. Med. Sci. 2015, 70, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blay, R.M.; Pinamang, A.D.; Sagoe, A.E.; Owusu, E.D.A.; Koney, N.K.K.; Arko-Boham, B. Influence of Lifestyle and Environmental Factors on Semen Quality in Ghanaian Men. Int. J. Reprod. Med. 2020, 2020, 6908458. [Google Scholar] [CrossRef] [PubMed]
- Hauser, R.; Sokol, R. Science linking environmental contaminant exposures with fertility and reproductive health impacts in the adult male. Fertil. Steril. 2008, 89, e59–e65. [Google Scholar] [CrossRef]
- Selvaraju, V.; Baskaran, S.; Agarwal, A.; Henkel, R. Environmental contaminants and male infertility: Effects and mechanisms. Andrologia 2021, 53, e13646. [Google Scholar] [CrossRef]
- Harper, A.P.; Finger, B.J.; Green, M.P. Chronic atrazine exposure beginning prenatally impacts liver function and sperm concentration with multi-generational consequences in mice. Front. Endocrinol. 2020, 11, 917. [Google Scholar] [CrossRef]
- Komsky-Elbaz, A.; Zubov, A.; Roth, Z. Effect of the herbicide atrazine and its major metabolite, DACT, on bovine sperm cryotolerance. Theriogenology 2019, 140, 117–123. [Google Scholar] [CrossRef]
- Abarikwu, S.O.; Costa, G.M.; Lacerda, S.M.; de França, L.R. Atrazine impairs testicular function in BalB/c mice by affecting Leydig cells. Toxicology 2021, 455, 152761. [Google Scholar] [CrossRef]
- Saalfeld, G.Q.; Varela Junior, A.S.; Castro, T.; Pereira, F.A.; Gheller, S.M.M.; da Silva, A.C.; Corcini, C.D.; da Rosa, C.E.; Colares, E.P. Low atrazine dosages reduce sperm quality of Calomys laucha mice. Environ. Sci. Pollut. Res. 2018, 25, 2924–2931. [Google Scholar] [CrossRef]
- Zhang, X.; Cui, W.; Wang, K.E.; Chen, R.; Chen, M.; Lan, K.; Wei, Y.; Pan, C.; Lan, X. Chlorpyrifos inhibits sperm maturation and induces a decrease in mouse male fertility. Environ. Res. 2020, 188, 109785. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razik, R.K.; Mosallam, E.M.; Hamed, N.A.; Badawy, M.E.; Abo-El-Saad, M.M. Testicular deficiency associated with exposure to cypermethrin, imidacloprid, and chlorpyrifos in adult rats. Environ. Toxicol. Pharmacol. 2021, 87, 103724. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, A.A.; Ogaly, H.A.; Ibrahim, M.A.; Abdallah, A.A.; Zaki, A.R.; Tohamy, A.F. The reproductive injury and oxidative testicular toxicity induced by chlorpyrifos can be restored by zinc in male rats. Biol. Trace Elem. Res. 2022, 200, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Babazadeh, M.; Najafi, G. Effect of chlorpyrifos on sperm characteristics and testicular tissue changes in adult male rats. Vet. Res. Forum. 2017, 8, 319–326. [Google Scholar]
- Rocco, L.; Peluso, C.; Cesaroni, F.; Morra, N.; Cesaroni, D.; Stingo, V. Genomic damage in human sperm cells exposed in vitro to environmental pollutants. J. Environ. Anal. Toxicol. 2012, 2, 1000117. [Google Scholar] [CrossRef]
- Kennedy, J.H.; Korn, N.; Thurston, R.J. Prostaglandin levels in seminal plasma and sperm extracts of the domestic turkey, and the effects of cyclooxygenase inhibitors on sperm mobility. Reprod. Biol. Endocrinol. 2003, 1, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Megharbel, S.M.; Al-Salmi, F.A.; Al-Harthi, S.; Alsolami, K.; Hamza, R.Z. Chitosan/Selenium Nanoparticles Attenuate Diclofenac Sodium-Induced Testicular Toxicity in Male Rats. Crystals 2021, 11, 1477. [Google Scholar] [CrossRef]
- Gallego-Ríos, S.E.; Atencio-García, V.J.; Peñuela, G.A. Effect of ibuprofen in vivo and in vitro on the sperm quality of the striped catfish Pseudoplatystoma magdaleniatum. Environ. Sci. Pollut. Res. 2021, 28, 36133–36141. [Google Scholar] [CrossRef]
- Uzun, B.; Atli, O.; Perk, B.O.; Burukoglu, D.İ.L.E.K.; Ilgin, S. Evaluation of the reproductive toxicity of naproxen sodium and meloxicam in male rats. Hum. Exp. Toxicol. 2015, 34, 415–429. [Google Scholar] [CrossRef]
- Tanyildizi, S.; Bozkurt, T. The effects of lincomycin-spectinomycin and sulfamethoxazole-trimethoprim on hyaluronidase activities and sperm characteristics of rams. J. Vet. Med. Sci. 2003, 65, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oputiri, D.; Elias, A. Impact of co-administered lopinavir/ritonavir and sulfamethoxazole/trimethoprim on reproductive indices of male albino rats. Am. J. Pharmacol. Sci. 2014, 2, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lu, G.; Wu, D.; Yan, Z. A multi-biomarker assessment of single and combined effects of norfloxacin and sulfamethoxazole on male goldfish (Carassius auratus). Ecotoxicol. Environ. Saf. 2014, 102, 12–17. [Google Scholar] [CrossRef]
- Mac, M.J.; Edsall, C.C. Environmental contaminants and the reproductive success of lake trout in the great lakes: An epidemiological approach. J. Toxicol. Environ. Health—A Curr. Issues 2009, 33, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Brasso, R.L.; Cristol, D.A. Effects of mercury exposure on the reproductive success of tree swallows (Tachycineta bicolor). Ecotoxicology 2008, 17, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Barraza, A.D.; Finlayson, K.A.; Leusch, F.D.L.; van de Merwe, J.P. Systematic review of reptile reproductive toxicology to inform future research directions on endangered or threatened species, such as sea turtles. Environ. Pollut. 2021, 286, 117470. [Google Scholar] [CrossRef]
- Barakat, R.; Seymore, T.; Lin, P.C.P.; Park, C.J.; Ko, C.M.J. Prenatal exposure to an environmentally relevant phthalate mixture disrupts testicular steroidogenesis in adult male mice. Environ. Res. 2019, 172, 194–201. [Google Scholar] [CrossRef]
- Facemire, C.F.; Gross, T.S.; Guillette, L.J. Reproductive impairment in the Florida panther: Nature or nurture? Environ. Health Perspect. 1995, 103, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Tosti, E. Effects of ecosystem stress on reproduction and development. Mol. Reprod. Dev. 2019, 86, 1269–1272. [Google Scholar] [CrossRef] [Green Version]
- Slutsky, M.; Levin, J.L.; Levy, B.S. Azoospermia and Oligospermia among a Large Cohort of DBCP Applicators in 12 Countries. Int. J. Occup. Environ. Health 1999, 5, 116–122. [Google Scholar] [CrossRef]
- Ojemaye, C.Y.; Onwordi, C.T.; Petrik, L. Herbicides in the tissues and organs of different fish species (Kalk Bay harbour, South Africa): Occurrence, levels and risk assessment. Int. J. Environ. Sci. Technol. 2020, 17, 1637–1648. [Google Scholar] [CrossRef]
- Krzastek, S.C.; Farhi, J.; Gray, M.; Smith, R.P. Impact of environmental toxin exposure on male fertility potential. Transl. Androl. Urol. 2020, 9, 2797. [Google Scholar] [CrossRef]
- Ghafouri-Khosrowshahi, A.; Ranjbar, A.; Mousavi, L.; Nili-Ahmadabadi, H.; Ghaffari, F.; Zeinvand-Lorestani, H.; Nili-Ahmadabadi, A. Chronic exposure to organophosphate pesticides as an important challenge in promoting reproductive health: A comparative study. J. Educ. Health Promot. 2019, 8, 149. [Google Scholar]
- Kazemi, M.; Tahmasbi, A.M.; Valizadeh, R.; Naserian, A.A.; Soni, A. Organophosphate pesticides: A general review. J. Agric. Res. 2012, 2, 512–522. Available online: https://profdoc.um.ac.ir/paper-abstract-1030335.html (accessed on 14 March 2023).
- Münze, R.; Hannemann, C.; Orlinskiy, P.; Gunold, R.; Paschke, A.; Foit, K.; Becker, J.; Kaske, O.; Paulsson, E.; Peterson, M.; et al. Pesticides from wastewater treatment plant effluents affect invertebrate communities. Sci. Total Environ. 2017, 599, 387–399. [Google Scholar] [CrossRef]
- Gonsioroski, A.; Mourikes, V.E.; Flaws, J.A. Endocrine disruptors in water and their effects on the reproductive system. Int. J. Mol. Sci. 2020, 21, 1929. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Yu, Y.; Zhang, X.; Zhang, D.; Zhang, S.; Wu, M. Organochlorine pesticides in fish from Taihu Lake, China, and associated human health risk assessment. Ecotoxicol. Environ. Saf. 2013, 98, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Toms, L.M.L.; He, C.; Hobson, P.; Sly, P.D.; Aylward, L.L.; Mueller, J.F. Pesticide metabolite concentrations in Queensland pre-schoolers—Exposure trends related to age and sex using urinary biomarkers. Environ. Res. 2019, 176, 108532. [Google Scholar] [CrossRef] [PubMed]
- Chetty-Mhlanga, S.; Basera, W.; Fuhrimann, S.; Probst-Hensch, N.; Delport, S.; Mugari, M.; van Wyk, J.; Röösli, M.; Dalvie, M.A. A prospective cohort study of school-going children investigating reproductive and neurobehavioral health effects due to environmental pesticide exposure in the Western Cape, South Africa: Study protocol. BMC Public Health 2018, 18, 857. [Google Scholar] [CrossRef] [Green Version]
- Chetty-Mhlanga, S.; Fuhrimann, S.; Basera, W.; Eeftens, M.; Röösli, M.; Dalvie, M.A. Association of activities related to pesticide exposure on headache severity and neurodevelopment of school-children in the rural agricultural farmlands of the Western Cape of South Africa. Environ. Int. 2021, 146, 106237. [Google Scholar] [CrossRef]
- Piel, C.; Pouchieu, C.; Carles, C.; Béziat, B.; Boulanger, M.; Bureau, M.; Busson, A.; Grüber, A.; Lecluse, Y.; Migault, L.; et al. Agricultural exposures to carbamate herbicides and fungicides and central nervous system tumour incidence in the cohort AGRICAN. Environ. Int. 2019, 130, 104876. [Google Scholar] [CrossRef] [PubMed]
- Matos, B.; Publicover, S.J.; Castro, L.F.C.; Esteves, P.J.; Fardilha, M. Brain and testis: More alike than previously thought? Open Biol. 2021, 11, 200322. [Google Scholar] [CrossRef] [PubMed]
- Njoku, K.L.; Ezeh, C.V.; Obidi, F.O.; Akinola, M.O. Assessment of Pesticide Residue Levels in Vegetables sold in some Markets in Lagos State, Nigeria. Niger. J. Biotechnol. 2017, 32, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Al-ShamarY, N.M.; Al-Ghouti, M.A.; Al-Shaikh, I.; Al-Meer, S.H.; Ahmad, T.A. Evaluation of pesticide residues of organochlorine in vegetables and fruits in Qatar: Statistical analysis. Environ. Monit. Assess. 2016, 188, 198. [Google Scholar] [CrossRef]
- Centner, T.J. Cancelling pesticide registrations and revoking tolerances: The case of chlorpyrifos. Environ. Toxicol. Pharmacol. 2018, 57, 53–61. [Google Scholar] [CrossRef]
- Knapke, E.T.; Magalhaes, D.D.P.; Dalvie, M.A.; Mandrioli, D.; Perry, M.J. Environmental and occupational pesticide exposure and human sperm parameters: A Navigation Guide review. Toxicology 2022, 465, 153017. [Google Scholar] [CrossRef]
- Sadeghnia, H.; Shahba, S.; Ebrahimzadeh-Bideskan, A.; Mohammadi, S.; Malvandi, A.M.; Mohammadipour, A. Atrazine neural and reproductive toxicity. Toxin Rev. 2021, 41, 1290–1303. [Google Scholar] [CrossRef]
- Shang, J.Z.; Li, S.R.; Li, X.Q.; Zhou, Y.T.; Ma, X.; Liu, L.; Niu, D.; Duan, X. Simazine perturbs the maturational competency of mouse oocyte through inducing oxidative stress and DNA damage. Ecotoxicol. Environ. Saf. 2022, 230, 113105. [Google Scholar] [CrossRef]
- Eshete, M.; Bowleg, J.; Perales, S.G.; Okunrobo, M.; Watkins, D.; Spencer, H. Adsorption of propazine, simazine and bisphenol A on the surface of nanoparticles of iron oxide nanoparticles of carbon and metallic oxides. J. Environ. Prot. 2018, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Park, H.O.; Bae, J. Disturbed relaxin signaling pathway and testicular dysfunction in mouse offspring upon maternal exposure to simazine. PLoS ONE 2012, 7, e44856. [Google Scholar] [CrossRef] [PubMed]
- Dantas, T.A.; Cancian, G.; Neodini, D.N.R.; Mano, D.R.S.; Capucho, C.; Predes, F.S.; Pulz, R.B.; Pigoso, A.A.; Dolder, H.; Severi-Aguiar, G.D.C. Leydig cell number and sperm production decrease induced by chronic ametryn exposure: A negative impact on animal reproductive health. Environ. Sci. Pollut. Res. 2015, 22, 8526–8535. [Google Scholar] [CrossRef]
- Park, H.O.; Ko, J.J.; Bae, J.H. Simazine-induced Alteration of the Expression Levels of Apoptosis-and Steroidogenesis-regulating Genes in Testicular Cells. Dev. Reprod. 2011, 15, 159–166. [Google Scholar]
- Sang, C.; Sørensen, P.B.; An, W.; Andersen, J.H.; Yang, M. Chronic health risk comparison between China and Denmark on dietary exposure to chlorpyrifos. Environ. Pollut. 2020, 257, 113590. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Chlorpyrifos; Tolerance revocations: Proposed rule. Fed. Regist. 2015, 80, 69080–69110. [Google Scholar]
- US EPA. Chlorpyrifos; Order denying PANNA and NRDC’s petition to revoke tolerances. Fed. Regist. 2017, 82, 16581–16592. [Google Scholar]
- Li, J.; Pang, G.; Ren, F.; Fang, B. Chlorpyrifos-induced reproductive toxicity in rats could be partly relieved under high-fat diet. Chemosphere 2019, 229, 94–102. [Google Scholar] [CrossRef]
- Archer, E.; Petrie, B.; Kasprzyk-Hordern, B.; Wolfaardt, G.M. The fate of pharmaceuticals and personal care products (PPCPs), endocrine disrupting contaminants (EDCs), metabolites and illicit drugs in a WWTW and environmental waters. Chemosphere 2017, 174, 437–446. [Google Scholar] [CrossRef]
- Ojemaye, C.Y.; Pampanin, D.M.; Sydnes, M.O.; Green, L.; Petrik, L. The burden of emerging contaminants upon an Atlantic Ocean marine protected reserve adjacent to Camps Bay, Cape Town, South Africa. Heliyon 2022, 8, e12625. [Google Scholar] [CrossRef]
- Ruan, Y.; Lin, H.; Zhang, X.; Wu, R.; Zhang, K.; Leung, K.M.; Lam, J.C.; Lam, P.K. Enantiomer-specific bioaccumulation and distribution of chiral pharmaceuticals in a subtropical marine food web. J. Hazard. Mater. 2020, 394, 122589. [Google Scholar] [CrossRef] [PubMed]
- Ojemaye, C.Y.; Petrik, L. Occurrences, levels and risk assessment studies of emerging pollutants (pharmaceuticals, perfluoroalkyl and endocrine disrupting compounds) in fish samples from Kalk Bay harbour, South Africa. Environ. Pollut. 2019, 252, 562–572. [Google Scholar] [CrossRef]
- Brown, A.R.; Gunnarsson, L.; Kristiansson, E.; Tyler, C.R. Assessing variation in the potential susceptibility of fish to pharmaceuticals, considering evolutionary differences in their physiology and ecology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibamureke, U.M.C.; Wagenaar, G.M. Histopathological changes in Oreochromis mossambicus (Peters, 1852) ovaries after a chronic exposure to a mixture of the HIV drug nevirapine and the antibiotics sulfamethoxazole and trimethoprim. Chemosphere 2021, 274, 129900. [Google Scholar] [CrossRef]
- Arshad, M.; Ara, C.; Andleeb, S.; Ahmad, N. Teratogenesis Induced by Trimethoprim Sulfamethoxazole in Mice. Pak. J. Zool. 2018, 50, 5. [Google Scholar] [CrossRef]
- Bedford, S.J.; McDonnell, S.M. Measurements of reproductive function in stallions treated with trimethoprim-sulfamethoxazole and pyrimethamine. J. Am. Vet. Med. Assoc. 1999, 215, 1317–1319. [Google Scholar]
- Salarkia, E.; Sepehri, G.; Torabzadeh, P.; Abshenas, J.; Saberi, A. Effects of administration of co-trimoxazole and folic acid on sperm quality and histological changes of testes in male rats. Int. J. Reprod. BioMed. 2017, 15, 625. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, C.A.; Rogers, S.; Hills, F.; Rahman, F.; Howell, R.J.; Homa, S.T. Effects of co-trimoxazole, erythromycin, amoxycillin, tetracycline and chloroquine on sperm function in vitro. Hum. Reprod. 1998, 13, 1878–1886. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.J.; Sharma, S. Effect Of Antibiotic Therapy On Sperm Quality. Eur. J. Mol. Clin. Med. 2020, 7, 2020. [Google Scholar]
- Madikizela, L.M.; Ncube, S. Occurrence and ecotoxicological risk assessment of non-steroidal anti-inflammatory drugs in South African aquatic environment: What is known and the missing information? Chemosphere 2021, 280, 130688. [Google Scholar] [CrossRef]
- Gonzalez-Rey, M.; Bebianno, M.J. Does non-steroidal anti-inflammatory (NSAID) ibuprofen induce antioxidant stress and endocrine disruption in mussel Mytilus galloprovincialis? Environ. Toxicol. Pharmacol. 2012, 33, 361–371. [Google Scholar] [CrossRef]
- Kwak, K.; Ji, K.; Kho, Y.; Kim, P.; Lee, J.; Ryu, J.; Choi, K. Chronic toxicity and endocrine disruption of naproxen in freshwater waterfleas and fish, and steroidogenic alteration using H295R cell assay. Chemosphere 2018, 204, 156–162. [Google Scholar] [CrossRef]
- Tiwary, E.; Hu, M.; Prasain, J.K. Sperm-Guiding Unconventional Prostaglandins in C. elegans: Synthesis and Signaling. Metabolites 2021, 11, 853. [Google Scholar] [CrossRef]
- Wojcieszyńska, D.; Guzik, U. Naproxen in the environment: Its occurrence, toxicity to nontarget organisms and biodegradation. Appl. Microbiol. Biotechnol. 2020, 104, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Qiu, B.; Dang, Y.; Sun, D. Enhanced adsorption of naproxen from aquatic environments by β-cyclodextrin-immobilized reduced graphene oxide. Chem. Eng. J. 2021, 412, 128710. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Fatima, M.; Hossain, M.; Mondal, A.C. Evaluation of naproxen-induced oxidative stress, hepatotoxicity and in-vivo genotoxicity in male Wistar rats. J. Pharm. Anal. 2018, 8, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A.; Purohit, A.; Ram, H. Assessment of dose-dependent reproductive toxicity of diclofenac sodium in male rats. Drug Chem. Toxicol. 2019, 42, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A. Effect of diclofenac on semen quality: A review. Andrologia 2021, 53, e14021. [Google Scholar] [CrossRef]
- Olayaki, L.A.; Adeyemi, W.J.; Yinusa, J.S.; Adedayo, G.A. Omega-3 fatty acids moderate oxidative and proinflammatory events in experimental hepatotoxicity in Wistar rats: Comparison with livolin. Synergy 2018, 7, 17–24. [Google Scholar] [CrossRef]
- Mousa, A.A.; Elweza, A.E.; Elbaz, H.T.; Tahoun, E.A.E.A.; Shoghy, K.M.; Elsayed, I. Eucalyptus Globulus protects against diclofenac sodium induced hepatorenal and testicular toxicity in male rats. J. Tradit. Complement. Med. 2020, 10, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Martien, K.M.; Gagnidze, K.; Pfaff, D. Implications of prenatal steroid perturbations for neurodevelopment, behavior, and autism. Endocr. Rev. 2014, 35, 961–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, J.F.; Geraghty, E.M.; Tancredi, D.J.; Delwiche, L.D.; Schmidt, R.J.; Ritz, B.; Hansen, R.L.; Hertz-Picciotto, I. Neurodevelopmental disorders and prenatal residential proximity to agricultural pesticides: The charge study. Environ. Health Perspect. 2014, 122, 1103–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schug, T.T.; Blawas, A.M.; Gray, K.; Heindel, J.J.; Lawler, C.P. Elucidating the links between endocrine disruptors and neurodevelopment. Endocrinology 2015, 156, 1941–1951. [Google Scholar] [CrossRef] [Green Version]
- Immler, S. The sperm factor: Paternal impact beyond genes. Heredity 2018, 121, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Champroux, A.; Cocquet, J.; Henry-Berger, J.; Drevet, J.R.; Kocer, A. A decade of exploring the mammalian sperm epigenome: Paternal epigenetic and transgenerational inheritance. Front. Cell Dev. Biol. 2018, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Blecharz-Klin, K.; Sznejder-Pachołek, A.; Wawer, A.; Pyrzanowska, J.; Piechal, A.; Joniec-Maciejak, I.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Early exposure to paracetamol reduces level of testicular testosterone and changes gonadal expression of genes relevant for steroidogenesis in rats offspring. Drug Chem. Toxicol. 2021, 45, 1862–1869. [Google Scholar] [CrossRef]
- Aleixo, J.F.; Pereira, M.R.F.; Montagnini, B.G.; Pereira, M.J.D.; Forcato, S.; Moreira, E.G.; Ceravolo, G.S.; Vieira, M.L.; Kiss, A.C.I.; Gerardin, D.C.C. Effect of paracetamol treatment on maternal care and reproductive outcomes in female rat offspring. Reprod. Fertil. Dev. 2021, 32, 1311–1325. [Google Scholar] [CrossRef]
- Altindağ, F.; Rağbetli, M.Ç. The effect of maternal treatment with diclofenac sodium and thymoquinone on testicular parameters in rat offspring. Rev. Int. Androl. 2021, 19, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Manikkam, M.; Guerrero-Bosagna, C.; Tracey, R.; Haque, M.M.; Skinner, M.K. Transgenerational Actions of Environmental Compounds on Reproductive Disease and Identification of Epigenetic Biomarkers of Ancestral Exposures. PLoS ONE 2012, 7, e31901. [Google Scholar] [CrossRef] [Green Version]
- McBirney, M.; King, S.E.; Pappalardo, M.; Houser, E.; Unkefer, M.; Nilsson, E.; Sadler-Riggleman, I.; Beck, D.; Winchester, P.; Skinner, M.K. Atrazine induced epigenetic transgenerational inheritance of disease, lean phenotype and sperm epimutation pathology biomarkers. PLoS ONE 2017, 12, e0184306. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Gonzalez, P.; Anderson, R.A.; Macdonald, J.; van den Driesche, S.; Kilcoyne, K.; Jørgensen, A.; McKinnell, C.; Macpherson, S.; Sharpe, R.M.; Mitchell, R.T. Effects of exposure to acetaminophen and ibuprofen on fetal germ cell development in both sexes in rodent and human using multiple experimental systems. Environ. Health Perspect. 2018, 126, 047006. [Google Scholar] [CrossRef]
- Albert, O.; Desdoits-Lethimonier, C.; Lesné, L.; Legrand, A.; Guillé, F.; Bensalah, K.; Dejucq-Rainsford, N.; Jégou, B. Paracetamol, aspirin and indomethacin display endocrine disrupting properties in the adult human testis in vitro. Hum. Reprod. 2013, 28, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Van den Driesche, S.; Macdonald, J.; Anderson, R.A.; Johnston, Z.C.; Chetty, T.; Smith, L.B.; McKinnell, C.; Dean, A.; Homer, N.Z.; Jorgensen, A.; et al. Prolonged exposure to acetaminophen reduces testosterone production by the human fetal testis in a xenograft model. Sci. Transl. Med. 2015, 7, 288ra80. [Google Scholar] [CrossRef] [Green Version]
- Gervin, K.; Nordeng, H.; Ystrom, E.; Reichborn-Kjennerud, T.; Lyle, R. Long-term prenatal exposure to paracetamol is associated with DNA methylation differences in children diagnosed with ADHD. Clin. Epigenetics 2017, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.; Rangabhashiyam, S.; Verma, P.; Singh, P.; Devi, P.; Kumar, P.; Hussain, C.M.; Gaurav, G.K.; Kumar, K.S. Environmental and health impacts of contaminants of emerging concerns: Recent treatment challenges and approaches. Chemosphere 2021, 272, 129492. [Google Scholar] [CrossRef]
- Warren-Vega, W.M.; Campos-Rodríguez, A.; Zárate-Guzmán, A.I.; Romero-Cano, L.A. A Current Review of Water Pollutants in American Continent: Trends and Perspectives in Detection, Health Risks, and Treatment Technologies. Int. J. Environ. Res. Public Health 2023, 20, 4499. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.I.; Dar, M.U.; Bhat, R.A.; Singh, J.P.; Singh, K.; Bhat, S.A. Prospectives and challenges of wastewater treatment technologies to combat contaminants of emerging concerns. Ecol. Eng. 2020, 152, 105882. [Google Scholar] [CrossRef]
- Rizzo, L.; Malato, S.; Antakyali, D.; Beretsou, V.G.; Đolić, M.B.; Gernjak, W.; Heath, E.; Ivancev-Tumbas, I.; Karaolia, P.; Ribeiro, A.R.; et al. Consolidated vs new advanced treatment methods for the removal of contaminants of emerging concern from urban wastewater. Sci. Total Environ. 2019, 655, 986–1008. [Google Scholar] [CrossRef] [PubMed]
- Yokota, H.; Higashi, K.; Hanada, E.; Matsuzaki, E.; Tsuruda, Y.; Suzuki, T.; Nakano, E.; Eguchi, S. Recovery from reproductive and morphological abnormalities in medaka (Oryzias latipes) following a 14-day exposure to diclofenac. Environ. Toxicol. Chem. 2017, 36, 3277–3283. [Google Scholar] [CrossRef]
- Østensen, M.; Khamashta, M.; Lockshin, M.; Parke, A.; Brucato, A.; Carp, H.; Doria, A.; Rai, R.; Meroni, P.; Cetin, I.; et al. Anti-inflammatory and immunosuppressive drugs and reproduction. Arthritis Res. 2006, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Lueders, I.; Young, D.; Maree, L.; Van der Horst, G.; Luther, I.; Botha, S.; Tindall, B.; Fosgate, G.; Ganswindt, A.; Bertschinger, H.J. Effects of GnRH vaccination in wild and captive African Elephant bulls (Loxodonta africana) on reproductive organs and semen quality. PLoS ONE 2017, 12, e0178270. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Kaur, P.; Ghanghas, P.; Kaur, J.; Kaushal, N. Selenium ameliorates ibuprofen induced testicular toxicity by redox regulation: Running head: Se protects against NSAID induced testicular toxicity. Reprod. Toxicol. 2020, 96, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. The effect of antioxidants on male factor infertility: The Males, Antioxidants, and Infertility (MOXI) randomized clinical trial. Fertil. Steril. 2020, 113, 552–560. [Google Scholar] [CrossRef]
- Vasilevskaya, N.V.; Sikalyuk, A.I. Bioindication of toxicity and mutagenicity of the environment. IOP Conf. Ser. Earth Environ. Sci. 2021, 677, 052067. [Google Scholar] [CrossRef]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The natural indicator of environmental pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Fabbrocini, A.; Di Stasio, M.; D’Adamo, R. Computerized sperm motility analysis in toxicity bioassays: A new approach to pore water quality assessment. Ecotoxicol. Environ. Saf. 2010, 73, 1588–1595. [Google Scholar] [CrossRef]
- Shaliutina, O.; Materiienko, A.; Shaliutina-Kolešová, A.; Gazo, I. Using fish spermatozoa in in vitro toxicity tests: A potential toxicology tool. Aquaculture 2021, 539, 736647. [Google Scholar] [CrossRef]
- Jain, A.K.; Singh, D.; Dubey, K.; Maurya, R.; Mittal, S.; Pandey, A.K. Models and methods for in vitro toxicity. In In Vitro Toxicol; Academic Press: Cambridge, MA, USA, 2018; pp. 45–46. [Google Scholar] [CrossRef]
- Kollár, T.; Kása, E.; Ferincz, Á.; Urbányi, B.; Csenki-Bakos, Z.; Horváth, Á. Development of an in vitro toxicological test system based on zebrafish (Danio rerio) sperm analysis. Environ. Sci. Pollut. Res. Int. 2018, 25, 14426–14436. [Google Scholar] [CrossRef]
- Aitken, R.J. Development of in vitro tests of human sperm function: A diagnostic tool and model system for toxicological analyses. Toxicol. Vitr. 1990, 4, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.; Binet, M.T.; Styan, C.A. Importance of sperm density in assessing the toxicity of metals to the fertilization of broadcast spawners. Ecotoxicol Environ Saf. 2019, 172, 547–555. [Google Scholar] [CrossRef]
- Dietrich, G.J.; Dietrich, M.; Kowalski, R.K.; Dobosz, S.; Karol, H.; Demianowicz, W.; Glogowski, J. Exposure of rainbow trout milt to mercury and cadmium alters sperm motility parameters and reproductive success. Aquat Toxicol. 2010, 97, 277–284. [Google Scholar] [CrossRef] [PubMed]
- De Assis, C.B.; Branco, G.S.; Moreira, R.G.; Pinheiro, J.P.S. Aluminum at environmental concentrations affects the sperm quality of the freshwater teleost Astyanax altiparanae: An in vitro study. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 243, 108983. [Google Scholar] [CrossRef]
- Kim, S.H.; Bae, J.W.; Kim, D.H.; Jeong, D.J.; Ha, J.J.; Yi, J.K.; Kwon, W.S. Detrimental effects of temephos on male fertility: An in vitro study on a mouse model. Reprod. Toxicol. 2020, 96, 150–155. [Google Scholar] [CrossRef]
- Selvaraju, S.; Nandi, S.; Gupta, P.S.P.; Ravindra, J.P. Effects of heavy metals and pesticides on buffalo (Bubalus bubalis) spermatozoa functions in vitro. Reprod. Domest. Anim. 2011, 46, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Betancourt, M.; Reséndiz, A. Effect of two insecticides and two herbicides on the porcine sperm motility patterns using computer-assisted semen analysis (CASA) in vitro. Reprod. Toxicol. 2006, 22, 508–512. [Google Scholar] [CrossRef]
- Pflieger-Bruss, S.; Hagemann, S.; Körner, W.; Hanf, V.; Köhn, F.M.; Müller, C.; Schill, W.B. Effects of single non-ortho, mono-ortho, and di-ortho chlorinated biphenyls on human sperm functions in vitro. Reprod. Toxicol. 2006, 21, 280–284. [Google Scholar] [CrossRef]
- Anifandis, G.; Amiridis, G.; Dafopoulos, K.; Daponte, A.; Dovolou, E.; Gavriil, E.; Gorgogietas, V.; Kachpani, E.; Mamuris, Z.; Messini, C.I.; et al. The in vitro impact of the herbicide roundup on human sperm motility and sperm mitochondria. Toxics 2017, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Montano, L.; Donato, F.; Bianco, P.M.; Lettieri, G.; Guglielmino, A.; Motta, O.; Bonapace, I.M.; Piscopo, M. Semen quality as a potential susceptibility indicator to SARS-CoV-2 insults in polluted areas. Environ. Sci. Pollut. Res. Int. 2021, 28, 37031–37040. [Google Scholar] [CrossRef]
- Hardneck, F.; Israel, G.; Pool, E.; Maree, L. Quantitative assessment of heavy metal effects on sperm function using computer-aided sperm analysis and cytotoxicity assays. Andrologia 2018, 50, e13141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmer, T.; Ljungberg, B.; Jankowski, V.; Jankowski, J.; Glorieux, G.; Stegmayr, B.G. An in-vitro assay using human spermatozoa to detect toxicity of biologically active substances. Sci. Rep. 2019, 9, 14525. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, R.C.; Queiroz, S.C.D.N.; da Luz, C.F.P.; Porto, R.S.; Rath, S. Bee pollen as a bioindicator of environmental pesticide contamination. Chemosphere 2016, 163, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Ottaviano, E.; Mulcahy, D.L. Gametophytic Selection as a Factor of Crop Plant Evolution. In Developments in Agricultural and Managed Forest Ecology; Barigozzi, C., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 1986; Volume 16, pp. 101–120. [Google Scholar] [CrossRef]
- Herrero, M.; Hormaza, J.I. Pistil strategies controlling pollen tube growth. Sex. Plant Reprod. 1996, 9, 343–347. [Google Scholar] [CrossRef]
- Gerber, M.D.; Junior, A.S.V.; Caldas, J.S.; Corcini, C.D.; Lucia, T., Jr.; Corrêa, L.B.; Corrêa, É.K. Toxicity evaluation of parboiled rice effluent using sperm quality of zebrafish as bioindicator. Ecol. Indic. 2016, 61, 214–218. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Chemical | Concentrations | In Vivo/In Vitro | Study Population | Duration | Effects | Study | |
|---|---|---|---|---|---|---|---|
| Pesticides | Atrazine | 5 mg/kg bw/day | In vivo | Pregnant female mice and male pups after gestation | Early gestation day 9.5 until 12 or 26 weeks of age | ↓ Epididymal sperm concentration | Harper et al. (2020) [54] |
| ↑ Altered steroidogenic gene expression | |||||||
| ↓ Cells within the preimplantation embryo | |||||||
| Atrazine and diamino chlorotriazine | ATZ (0.1 or 1 μM) and DACT (1 or 10 μM) | In vitro | Bull semen | During cryopreservation (3 hrs) and during capacitation (4 hrs) | ↓ Sperm vitality | Komsky-Elbaz et al. (2019) [55] | |
| ↑ MMP | |||||||
| ↓ Ca++ ionophore-induced AR | |||||||
| Atrazine | 0.5, 25, and 50 mg/kg bw | In vivo | Young adult male mice | 3 days | ↓ Testis weight and gonadosomatic index | Abarikwu et al. (2021) [56] | |
| ↑ Abnormal histology of gonads | |||||||
| ↓ Testosterone levels and production | |||||||
| ↑ Impaired spermatogenesis | |||||||
| ↓ Leydig cell viability | |||||||
| Atrazine | 0.1 mg/kg, 1 mg/kg, and 10 mg/kg of bw | In vivo | Adult male rodents | 21 days | ↓ Total and prog mot | Saalfeld et al. (2018) [57] | |
| ↓ Sperm membrane integrity | |||||||
| ↑ Membrane fluidity | |||||||
| ↓ Mitochondrial functionality and acrosome integrity | |||||||
| Chlorpyrifos | Dietary CPF: 1 mg/kg/day, 3 mg/kg/day, or 12 mg/kg/day. Intraperitoneal CPF injection: 3 mg/kg/day, 6 mg/kg/day, or 12 mg/kg/day. Gavage CPF: 12 mg/kg/day and 25 mg/kg/day. In vitro: 25 μg/mL for sperm and 50 μM or 25 μM for cell lines | In vivo and in vitro | Male mice and germ cell culture | Dietary CPF: 80 days. Intraperitoneal CPF injection: 15 days. Gavage CPF: 35 or 70 days. In vitro: sperm 1 hrs, cell line 12 or 24 hrs | ↓ Expression steroid hormone synthesis-related genes. | Zhang et al. (2020) [58] | |
| ↓ Weight of gonads and associated structures | |||||||
| ↑ Protein expression of Caspase3 | |||||||
| ↓ Sperm density and prog mot and linear movement | |||||||
| ↑ ROS | |||||||
| ↓ MMP | |||||||
| ↓ Cell line normal morphology and viability | |||||||
| Chlorpyrifos, imidacloprid, and cypermethrin | 5 mg/kg bw CYP, 9 mg/kg bw IMC, and 1.9 mg/kg bw CPF | In vivo | Male rats | 5 times per week for 1 month | ↑ Testis weight | Abdel-Razik et al. (2021) [59] | |
| ↓ Epididymis and prostate gland weights | |||||||
| ↓ Sperm counts, moti, viability and normality | |||||||
| ↑ ROS, lipid peroxidation and testis protein carbonyl content | |||||||
| ↓ Serum testosterone, LH and FSH levels | |||||||
| ↓ Spermatogenesis | |||||||
| ↑ Abnormal histology of gonads and associated cells | |||||||
| Chlorpyrifos | 1 mg/kg | In vivo | Adult male rats | 60 days | ↓ Weight of gonads and associated structures | Khalaf et al. (2022) [60] | |
| ↓ Sperm count, sperm mot and prog mot | |||||||
| ↑ Dead and abnormal sperm | |||||||
| ↓ Serum testosterone, FSH, and LH levels | |||||||
| ↑ DNA laddering | |||||||
| Chlorpyrifos | 37 mg/kg/bw | In vivo | Male rats | Sampling on days 5, 15, 30, and 45 | ↓ Testicular weight | Babazadeh and Najafi (2017) [61] | |
| ↓ Sperm count, viability and mot | |||||||
| ↓ Leydig cells | |||||||
| ↑ Abnormal histology of gonads and associated cells | |||||||
| ↑ Immature sperm and DNA damage | |||||||
| Pharmaceuticals | Atorvastatin, sildenafil citrate, gemfibrozil, ibuprofen, atenolol, ofloxacin, carbamazepine, bezafibrate, and diclofenac | Atorvastatin (13 ng/mL), sildenafil citrate (26–25 ng/mL), gemfibrozil (380 ng/mL), ibuprofen (92 ng/mL), atenolol (241 ng/mL), ofloxacin (50 ng/mL), carbamazepine (310 ng/mL), bezafibrate (57 ng/mL), and diclofenac (180 ng/mL) | In vitro | Men aged 20–30 years | 15, 30, and 45 min | ↓ Sperm mot | Rocco et al. (2012) [62] |
| ↑ Genomic damage | |||||||
| ↑ Apoptotic cells and DNA fragmentation | |||||||
| Indomethacin, diclofenac sodium, tolmetin, acetylsalicylic acid, resveratrol, and NS-398 | 0 to 15 mM | In vitro | Turkey toms | 5 min | ↓ Sperm mot | Kennedy et al. (2003) [63] | |
| Diclofenac | 10 mg/kg | In vivo | Male rats | 30 days | ↓ Serum testosterone, LH and FSH | El-Megharbel et al. (2021) [64] | |
| ↓ Sperm mot and count | |||||||
| ↓ Testicular tissue antioxidant defence enzymes | |||||||
| Ibuprofen | 25 and 50 μg/L | In vivo and in vitro | Mature male striped catfish | 4 months | ↓ Prog and total mot, rapid and medium speeds | Gallego-Ríos et al. (2021) [65] | |
| ↑ Slow speeds and immotile sperm | |||||||
| ↓ VCL, VSL and VAP, LIN and STR, ALH, WOB and BCF | |||||||
| Naproxen and meloxicam | Naproxen (10 mg/kg) and meloxicam (1 mg/kg) | In vivo | Male rats | 35 days | ↓ Sperm mot and count | Uzun et al. (2015) [66] | |
| ↓ Prostaglandins and ROS defence enzymes in testis | |||||||
| ↑ Abnormal histology of gonads and associated cells | |||||||
| Lincomycin-spectinomycin and sulfamethoxazole-trimethoprim | Lincomycin-spectinomycin injected 0.1 mL/kg bw and sulfamethoxazole-trimethoprim orally administered at 0.12 mL/kg bw | In vivo | Rams | Intramuscular injections once daily for 3 days and oral administration twice daily for 3 days | ↑ Serum and semen hyaluronidase activity | Tanyildizi et al. (2003) [67] | |
| ↓ Sperm count | |||||||
| ↑ Sperm mot | |||||||
| Lopinavir-ritonavir and sulfamethoxazole-trimethoprim | 22.4/4.6 mg/kg of sulfamethoxazole-trimethoprim, 22.8/5.8 mg/kg of lopinavir-ritonavir, and combined doses of sulfamethoxazole-trimethoprim + lopinavir-ritonavir | In vivo | Male rats | 2–8 weeks | ↑ Testicular MDA | Oputiri and Elias (2014) [68] | |
| ↓ SOD | |||||||
| ↓ Sperm mot and count | |||||||
| ↑ Abnormal sperm morphology | |||||||
| ↑ Abnormal testicular histology | |||||||
| Norfloxacin and sulfamethoxazole | Norfloxacin: 0.0032, 0.016, 0.08, 0.4, 2, and 10 mg/L. Mixture of norfloxacin and sulfamethoxazole: 0.0016 + 0.008, 0.008 + 0.04, 0.04 + 0.2, 0.2 + 1.0, 1.0 + 5.0, and 5.0 + 25.0 mg/L, respectively. | In vivo | One year old male goldfish | 7 days | ↑ DNA damage of the gonads | Liu et al. (2014) [69] | |
| Species | Endogenous/Exogenous | Environmental Contaminant | Concentrations | Duration | Effects | Study |
|---|---|---|---|---|---|---|
| Sydney worm (Galeolaria caespitosa) | Exogenous | Heavy metals (Cu, Zn, Pb, and Cd) | Cu (12–33) Zn (160–550), Pb (560–1500), and Cd (4900–6100) μg/L | 30 min | ↓ Fertilization rate | Lockyer et al., 2019 [157] |
| Rainbow trout (Oncorhynchus mykiss) | Exogenous | Hg2+ and Cd2 | 1, 10, 100 mg Hg2+/L and 10, 100, 500 mg Cd2 | 4 and 24 h | ↓ Viability | Dietrich et al., 2010 [158] |
| ↓ Kinematics | ||||||
| ↑ DNA fragmentation | ||||||
| Yellow-tailed lambari (Astyanax altiparanae) | Exogenous | Aluminium | 0.05, 0.1, 0.3, and 0.5 mg/L | 50 sec, 10 and 30 min | ↓ Membrane vitality | de Assis et al., 2021 [159] |
| ↓ Mitochondrial activity | ||||||
| ↓ Mot and kinematics | ||||||
| Mouse (Mus musculus) | Endogenous | Temephos | 0.1, 1, 10, and 100 mM | 90 min | ↓ Sperm mot, medium and rapid sperm mot, prog mot, kinematics | Kim et al., 2020 [160] |
| ↑ Live AR | ||||||
| ↓ Live Capacitated | ||||||
| ↓ ATP levels | ||||||
| ↓ PKA activity and tyrosine phosphorylation | ||||||
| ↓ Fertilization rate | ||||||
| Buffalo (Bubalus bubalis) | Endogenous | Cadmium, lead, chlorpyrifos, and endosulfan | 0.005, 0.05, 0.02, 0.1, 0.5, 1.0, 2.0, and 4.0 μg/mL | 1 h | ↓ Prog and total mot | Selvaraju et al., 2011 [161] |
| ↓ Kinematics | ||||||
| ↑ Tail abnormality | ||||||
| ↓ Plasmalemma, functional membrane integrities and acrosomal integrities | ||||||
| ↓ Normal nuclear morphology | ||||||
| ↑ Nuclear chromatin decondensation | ||||||
| ↓ MMP | ||||||
| ↓ Sperm–zona binding and in vitro | ||||||
| Duroc boar (Sus scrofa) | Endogenous | Atrazine, fenoxaprop-ethyl, malathion, and diazinon | 50, 100, and 500 μM | 1 h | ↓ Viability | Betancourt and Reséndiz, 2006 [162] |
| ↓ Prog mot | ||||||
| ↓ Kinematics | ||||||
| Human (Homo sapiens) | Endogenous | PCB126, PCB118, and PCB153 | 2–20 μg PCB/mL | 5 hr | No effects observed on sperm mot, vitality, spontaneous AR, or inducibility of the AR. | Pflieger-Bruss et al., 2006 [164] |
| Human (Homo sapiens) | Endogenous | Roundup | 1 mg/L | 1 and 3 hr | ↓ Prog and total mot | Anifandis et al., 2017 [164] |
| ↓ MMP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcu, D.; Keyser, S.; Petrik, L.; Fuhrimann, S.; Maree, L. Contaminants of Emerging Concern (CECs) and Male Reproductive Health: Challenging the Future with a Double-Edged Sword. Toxics 2023, 11, 330. https://doi.org/10.3390/toxics11040330
Marcu D, Keyser S, Petrik L, Fuhrimann S, Maree L. Contaminants of Emerging Concern (CECs) and Male Reproductive Health: Challenging the Future with a Double-Edged Sword. Toxics. 2023; 11(4):330. https://doi.org/10.3390/toxics11040330
Chicago/Turabian StyleMarcu, Daniel, Shannen Keyser, Leslie Petrik, Samuel Fuhrimann, and Liana Maree. 2023. "Contaminants of Emerging Concern (CECs) and Male Reproductive Health: Challenging the Future with a Double-Edged Sword" Toxics 11, no. 4: 330. https://doi.org/10.3390/toxics11040330