
Reproductive Effects of S. boulardii on Sub-Chronic Acetamiprid and Imidacloprid Toxicity in Male Rats
 , , , ,
, , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Collection of Samples
2.3. Histopathological Examination
2.4. Immunohistochemical Examination
2.5. Collection of Samples, Sperm Evaluation and Epididymal Sperm Parameters
2.6. Statistic Analyses
3. Results
3.1. Pathologic Analyses
3.1.1. Histopathological Findings
3.1.2. Immunohystochemical Findings
3.2. Reproductive Organ Weights and Epididymal Sperm Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Huang, Y.; Chen, W.-J.; Wu, S.; Lei, Q.; Zhou, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Environmental occurrence, toxicity concerns, and biodegradation of neonicotinoid insecticides. Environ. Res. 2023, 218, 114953. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. An overview of the environmental risks posed by neonicotinoid insecticides. J. Appl. Ecol. 2013, 50, 977–987. [Google Scholar] [CrossRef]
- Tokumoto, J.; Danjo, M.; Kobayashi, Y.; Kinoshita, K.; Omotehara, T.; Tatsumi, A.; Hashiguchi, M.; Sekijima, T.; Kamisoyama, H.; Yokoyama, T.; et al. Effects of exposure to clothianidin on the reproductive system of male quails. J. Vet. Med. Sci. 2013, 75, 755–760. [Google Scholar] [CrossRef]
- Su, Y.; Ren, X.; Ma, X.; Wang, D.; Hu, H.; Song, X.; Cui, J.; Ma, Y.; Yao, Y. Evaluation of the Toxicity and Sublethal Effects of Acetamiprid and Dinotefuran on the Predator Chrysopa pallens (Rambur) (Neuroptera: Chrysopidae). Toxics 2022, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Arıcan, E.Y.; Kayalı, D.G.; Karaca, B.U.; Boran, T.; Öztürk, N.; Okyar, A.; Ercan, F.; Özhan, G. Reproductive effects of subchronic exposure to acetamiprid in male rats. Sci. Rep. 2020, 10, 8985. [Google Scholar] [CrossRef]
- Bal, R.; Türk, G.; Tuzcu, M.; Yilmaz, O.; Kuloglu, T.; Gundogdu, R.; Gür, S.; Agca, A.; Ulas, M.; Çambay, Z.; et al. Assessment of imidacloprid toxicity on reproductive organ system of adult male rats. J. Environ. Sci. Health Part B 2012, 47, 434–444. [Google Scholar] [CrossRef]
- Mehrpour, O.; Karrari, P.; Zamani, N.; Tsatsakis, A.M.; Abdollahi, M. Occupational exposure to pesticides and consequences on male semen and fertility: A review. Toxicol. Lett. 2014, 230, 146–156. [Google Scholar] [CrossRef]
- Gu, Y.-H.; Li, Y.; Huang, X.-F.; Zheng, J.-F.; Yang, J.; Diao, H.; Yuan, Y.; Xu, Y.; Liu, M.; Shi, H.-J.; et al. Reproductive effects of two neonicotinoid insecticides on mouse sperm function and early embryonic development in vitro. PLoS ONE 2013, 8, e70112. [Google Scholar] [CrossRef]
- Wang, A.; Wan, Y.; Zhou, L.; Xia, W.; Guo, Y.; Mahai, G.; Yang, Z.; Xu, S.; Zhang, R. Neonicotinoid insecticide metabolites in seminal plasma: Associations with semen quality. Sci. Total Environ. 2022, 811, 151407. [Google Scholar] [CrossRef] [PubMed]
- Mikolić, A.; Karačonji, I.B. Imidacloprid as reproductive toxicant and endocrine disruptor: Investigations in laboratory animals. Arch. Ind. Hyg. Toxicol. 2018, 69, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, J.; Ren, J.; Hou, Y.; Han, Z.; Xiao, J.; Li, Y. Exposure level of neonicotinoid insecticides in the food chain and the evaluation of their human health impact and environmental risk: An overview. Sustainability 2020, 12, 7523. [Google Scholar] [CrossRef]
- Hoshi, N.; Hirano, T.; Omotehara, T.; Tokumoto, J.; Umemura, Y.; Mantani, Y.; Tanida, T.; Warita, K.; Tabuchi, Y.; Yokoyama, T.; et al. Insight into the mechanism of reproductive dysfunction caused by neonicotinoid pesticides. Biol. Pharm. Bull. 2014, 37, 1439–1443. [Google Scholar] [CrossRef]
- Mosbah, R.; Djerrou, Z.; Mantovani, A. Protective effect of Nigella sativa oil against acetamiprid induced reproductive toxicity in male rats. Drug Chem. Toxicol. 2018, 41, 206–212. [Google Scholar] [CrossRef]
- Humann-Guilleminot, S.; de Montaigu, C.T.; Sire, J.; Grünig, S.; Gning, O.; Glauser, G.; Vallat, A.; Helfenstein, F. A sublethal dose of the neonicotinoid insecticide acetamiprid reduces sperm density in a songbird. Environ. Res. 2019, 177, 108589. [Google Scholar] [CrossRef]
- Mesnage, R.; Biserni, M.; Genkova, D.; Wesolowski, L.; Antoniou, M.N. Evaluation of neonicotinoid insecticides for oestrogenic, thyroidogenic and adipogenic activity reveals imidacloprid causes lipid accumulation. J. Appl. Toxicol. 2018, 38, 1483–1491. [Google Scholar] [CrossRef]
- Koliarakis, I.; Psaroulaki, A.; Nikolouzakis, T.K.; Kokkinakis, M.; Sgantzos, M.; Goulielmos, G.; Androutsopoulos, V.; Tsatsakis, A.; Tsiaoussis, J. Intestinal microbiota and colorectal cancer: A new aspect of research. J. BUON 2018, 23, 1216–1234. [Google Scholar] [PubMed]
- Mesnage, R.; Antoniou, M.N.; Tsoukalas, D.; Goulielmos, G.N.; Tsatsakis, A. Gut microbiome metagenomics to understand how xenobiotics impact human health. Curr. Opin. Toxicol. 2018, 11, 51–58. [Google Scholar] [CrossRef]
- Tsiaoussis, J.; Antoniou, M.N.; Koliarakis, I.; Mesnage, R.; Vardavas, C.I.; Izotov, B.N.; Psaroulaki, A.; Tsatsakis, A. Effects of single and combined toxic exposures on the gut microbiome: Current knowledge and future directions. Toxicol. Lett. 2019, 312, 72–97. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, R.; Calatayud, M.; Duysburgh, C.; Marzorati, M.; Antoniou, M.N. Alterations in infant gut microbiome composition and metabolism after exposure to glyphosate and Roundup and/or a spore-based formulation using the SHIME technology. Gut Microbiome 2022, 3, e6. [Google Scholar] [CrossRef]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2019, 73, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Tomaiuolo, R.; Veneruso, I.; Cariati, F.; D’Argenio, V. Microbiota and human reproduction: The case of male infertility. High-Throughput 2020, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, Z. Exploring the role of gut microbiome in male reproduction. Andrology 2022, 10, 441–450. [Google Scholar] [CrossRef]
- Średnicka, P.; Juszczuk-Kubiak, E.; Wójcicki, M.; Akimowicz, M.; Roszko, M.Ł. Probiotics as a biological detoxification tool of food chemical contamination: A review. Food Chem. Toxicol. 2021, 153, 112306. [Google Scholar] [CrossRef]
- Devan, R.S.; Mishra, A.; Prabu, P.; Mandal, T.; Panchapakesan, S. Sub-chronic oral toxicity of acetamiprid in Wistar rats. Toxicol. Environ. Chem. 2015, 97, 1236–1252. [Google Scholar] [CrossRef]
- Kobir, M.A.; Akter, L.; Sultana, N.; Pervin, M.; Awal, M.A.; Karim, M.R. Effects of imidacloprid-contaminated feed exposure on spermatogenic cells and Leydig cells in testes of adult male rabbits (Oryctolagus cuniculus). Saudi J. Biol. Sci. 2023, 30, 103541. [Google Scholar] [CrossRef]
- Tsatsakis, A.; Tyshko, N.V.; Goumenou, M.; Shestakova, S.I.; Sadykova, E.O.; Zhminchenko, V.M.; Zlatian, O.; Calina, D.; Pashorina, V.A.; Nikitin, N.S.; et al. Detrimental effects of 6 months exposure to very low doses of a mixture of six pesticides associated with chronic vitamin deficiency on rats. Food Chem. Toxicol. 2021, 152, 112188. [Google Scholar] [CrossRef]
- Committee, E.S. Guidance on conducting repeated-dose 90-day oral toxicity study in rodents on whole food/feed. EFSA J. 2011, 9, 2438. [Google Scholar] [CrossRef]
- Sevim, C.; Akpinar, E.; Tsatsakis, A.; Yildirim, S.; Tzatzarakis, M.; Vardavas, A.I.; Vardavas, C.I.; Kara, M.; Gul, Z. Investigation of the effects of probiotics on sub-chronic neonicotinoid toxicity in rats. Agronomy 2021, 11, 2003. [Google Scholar] [CrossRef]
- Sulukan, E.; Baran, A.; Şenol, O.; Kankaynar, M.; Yıldırım, S.; Bolat, I.; Ceyhun, H.A.; Toraman, E.; Ceyhun, S.B. Global warming and glyphosate toxicity (I): Adult zebrafish modelling with behavioural, immunohistochemical and metabolomic approaches. Sci. Total Environ. 2023, 858, 160086. [Google Scholar] [CrossRef] [PubMed]
- Sulukan, E.; Baran, A.; Kankaynar, M.; Kızıltan, T.; Bolat, I.; Yıldırım, S.; Ceyhun, H.A.; Ceyhun, S.B. Global warming and glyphosate toxicity (II): Offspring zebrafish modelling with behavioral, morphological and immunohistochemical approaches. Sci. Total Environ. 2023, 856, 158903. [Google Scholar] [CrossRef]
- Karaman, M.; Toraman, E.; Sulukan, E.; Baran, A.; Bolat, I.; Yıldırım, S.; Kankaynar, M.; Ghosigharehagaji, A.; Budak, H.; Ceyhun, S.B. Fluoride exposure causes behavioral, molecular and physiological changes in adult zebrafish (Danio rerio) and their offspring. Environ. Toxicol. Pharmacol. 2023, 97, 104044. [Google Scholar] [CrossRef]
- Cicek, B.; Genc, S.; Yeni, Y.; Kuzucu, M.; Cetin, A.; Yildirim, S.; Bolat, I.; Kantarci, M.; Hacimuftuoglu, A.; Lazopoulos, G.; et al. Artichoke (Cynara Scolymus) Methanolic Leaf Extract Alleviates Diethylnitrosamine-Induced Toxicity in BALB/c Mouse Brain: Involvement of Oxidative Stress and Apoptotically Related Klotho/PPARγ Signaling. J. Pers. Med. 2022, 12, 2012. [Google Scholar] [CrossRef]
- Türk, G.; Sönmez, M.; Aydin, M.; Yüce, A.; Gür, S.; Yüksel, M.; Aksu, E.H.; Aksoy, H. Effects of pomegranate juice consumption on sperm quality, spermatogenic cell density, antioxidant activity and testosterone level in male rats. Clin. Nutr. 2008, 27, 289–296. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Aksu, E.H.; Akman, O.; Özkaraca, M.; Ömür, A.D.; Uçar, Ö. Effect of Maclura Pomifera extract on cisplatin-induced damages in reproductive system of male rats. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 397–403. [Google Scholar]
- Türk, G.; Ateşşahin, A.; Sönmez, M.; Ceribaşi, A.O.; Yüce, A. Improvement of cisplatin-induced injuries to sperm quality, the oxidant-antioxidant system, and the histologic structure of the rat testis by ellagic acid. Fertil. Steril. 2008, 89, 1474–1481. [Google Scholar] [CrossRef]
- Li, Y.-J.; Song, T.-B.; Cai, Y.-Y.; Zhou, J.-S.; Song, X.; Zhao, X.; Wu, X.-L. Bisphenol A exposure induces apoptosis and upregulation of Fas/FasL and caspase-3 expression in the testes of mice. Toxicol. Sci. 2009, 108, 427–436. [Google Scholar] [CrossRef]
- Kim, J.M.; Ghosh, S.R.; Weil, A.C.P.; Zirkin, B.R. Caspase-3 and Caspaseactivated deoxyribonuclease are associated with testicular germ cell apoptosis resulting from reduced intratesticular testosterone. Endocrinology 2001, 142, 3809–3816. [Google Scholar] [CrossRef]
- Hifnawy, M.S.; Aboseada, M.A.; Hassan, H.M.; AboulMagd, A.M.; Tohamy, A.F.; Abdel-Kawi, S.H.; Rateb, M.E.; El Naggar, E.M.B.; Liu, M.; Quinn, R.J.; et al. Testicular caspase-3 and β-Catenin regulators predicted via comparative metabolomics and docking studies. Metabolites 2020, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Hartman, C.; Legoff, L.; Capriati, M.; Lecuyer, G.; Kernanec, P.-Y.; Tevosian, S.; D’Cruz, S.C.; Smagulova, F. Epigenetic Effects Promoted by Neonicotinoid Thiacloprid Exposure. Front. Cell Dev. Biol. 2021, 9, 1673. [Google Scholar] [CrossRef]
- Saber, T.M.; Arisha, A.H.; Abo-Elmaaty, A.M.A.; Abdelgawad, F.; Metwally, M.M.M.; Saber, T.; Mansour, M.F. Thymol alleviates imidacloprid-induced testicular toxicity by modulating oxidative stress and expression of steroidogenesis and apoptosis-related genes in adult male rats. Ecotoxicol. Environ. Saf. 2021, 221, 112435. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Anadón, A.; Wu, Q.; Qiao, F.; Ares, I.; Martínez-Larrañaga, M.-R.; Yuan, Z.; Martínez, M.-A. Mechanism of neonicotinoid toxicity: Impact on oxidative stress and metabolism. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 471–507. [Google Scholar] [CrossRef]
- Hirano, T.; Yanai, S.; Omotehara, T.; Hashimoto, R.; Umemura, Y.; Kubota, N.; Minami, K.; Nagahara, D.; Matsuo, E.; Aihara, Y.; et al. The combined effect of clothianidin and environmental stress on the behavioral and reproductive function in male mice. J. Vet. Med. Sci. 2015, 77, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Cardone, A. Imidacloprid induces morphological and molecular damages on testis of lizard (Podarcis sicula). Ecotoxicology 2015, 24, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Terayama, H.; Qu, N.; Endo, H.; Ito, M.; Tsukamoto, H.; Umemoto, K.; Kawakami, S.; Fujino, Y.; Tatemichi, M.; Sakabe, K. Effect of acetamiprid on the immature murine testes. Int. J. Environ. Health Res. 2018, 28, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Liu, Y. Microorganisms in the reproductive system and probiotic’s regulatory effects on reproductive health. Comput. Struct. Biotechnol. J. 2022, 20, 1541–1553. [Google Scholar] [CrossRef]
- Guo, Y.; Du, X.; Bian, Y.; Wang, S. Chronic unpredictable stress-induced reproductive deficits were prevented by probiotics. Reprod. Biol. 2020, 20, 175–183. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-induced pathogen susceptibility is mitigated by Lactobacillus plantarum immune stimulation in a Drosophila melanogaster model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef]
- El Euony, O.I.; Elblehi, S.; Abdel-Latif, H.M.; Abdel-Daim, M.M.; El-Sayed, Y.S. Modulatory role of dietary Thymus vulgaris essential oil and Bacillus subtilis against thiamethoxam-induced hepatorenal damage, oxidative stress, and immunotoxicity in African catfish (Clarias garipenus). Environ. Sci. Pollut. Res. 2020, 27, 23108–23128. [Google Scholar] [CrossRef]
- Chmiel, J.A.; Daisley, B.A.; Burton, J.P.; Reid, G. Deleterious effects of neonicotinoid pesticides on Drosophila melanogaster immune pathways. MBio 2019, 10, e01395-19. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Animal Groups | Findings |
|---|---|
| Control | Normal morphology data |
| Probiotics | Normal morphology compared to control group |
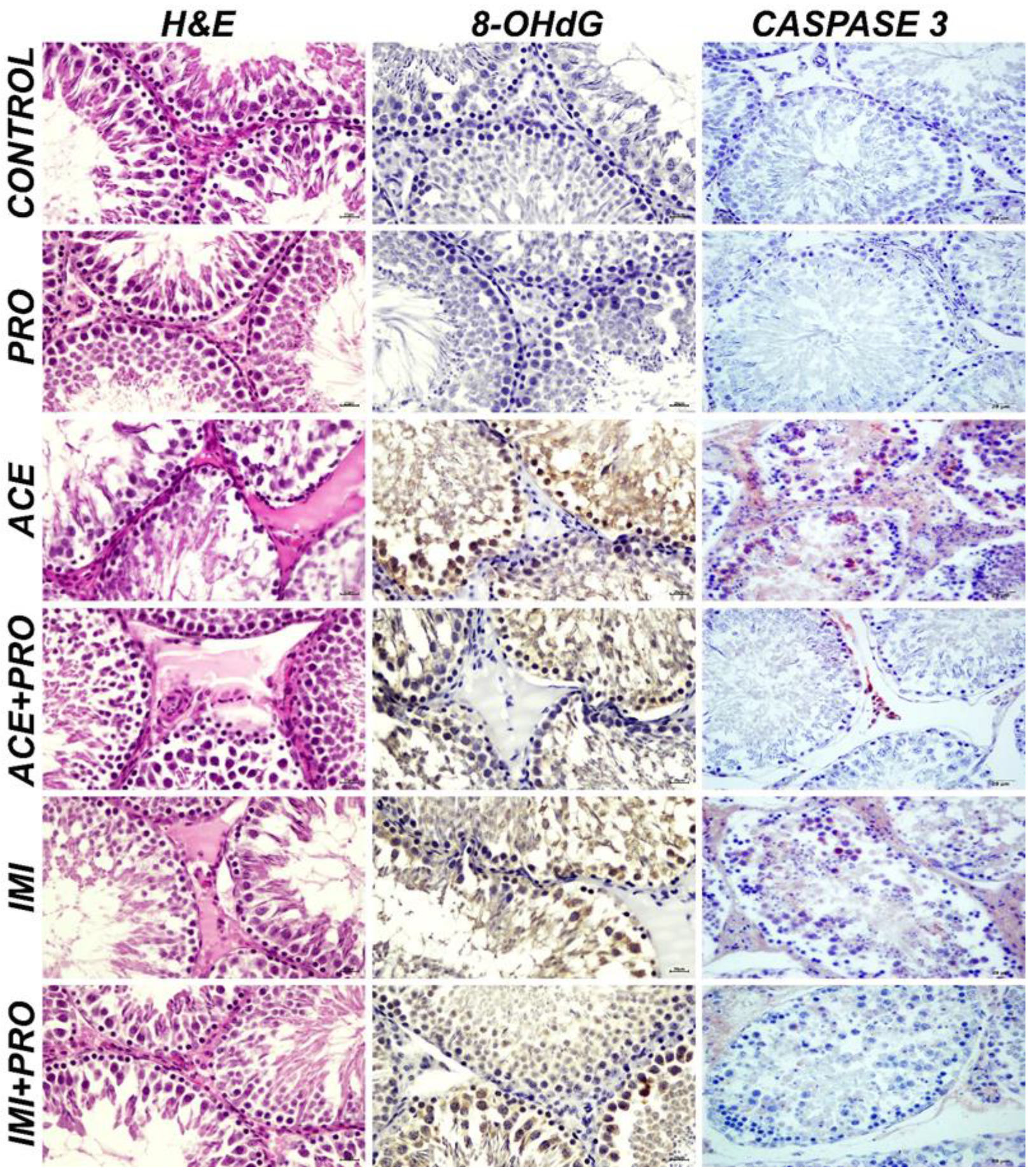
| Acetamiprid | Severe degeneration and necrosis has been detected/observed in spermatocytes at tubular wall; severe edema detected at intertubular region and hyperemia detected in blood vessels. |
| Acetamiprid + Probiotics | Slight degeneration and necrosis has been detected/observed in spermatocytes at tubular wall; edema detected at intertubular region and slight hyperemia detected in blood vessels. These results are significantly different compared to Acetamiprid group (p < 0.05). |
| Imidacloprid | Severe degeneration and necrosis has been detected/observed in spermatocytes at tubular wall and related with necrosis thinning of tubular wall detected; severe edema detected at intertubular region and severe hyperemia detected in blood vessels. |
| Imidacloprid + Probiotics | Slight hyperemia was detected in tubular wall and at vessels. These results are significantly different compared to Imidacloprid group (p < 0.05). |
| Control | Probiotics | Acetamiprid | Acetamiprid + Probiotics | Imidacloprid | Imidacloprid + Probiotics | |
|---|---|---|---|---|---|---|
| Degeneration at spermatocytes | − | − | +++ | + | +++ | + |
| Spermatocyte necrosis | − | − | +++ | + | +++ | − |
| Hyperemia in vessels | − | − | +++ | + | +++ | + |
| Tubular wall thinning | − | − | +++ | + | +++ | + |
| Edema at intertubular gap | − | − | +++ | + | +++ | − |
| Animal Groups | Findings |
|---|---|
| Control | When testicular tissues are examined immunohistochemically; Expressions of 8-OHdG and Caspase-3 were evaluated as negative |
| Probiotics | Expressions of 8-OHdG and Caspase-3 were evaluated as negative |
| Acetamiprid | Severe intracytoplasmic 8-OHdG and Caspase-3 expressions were detected. |
| Acetamiprid + Probiotics | Intermediate intracytoplasmic 8-OHdG and Caspase-3 expressions were detected. These results are significantly different compared to Acetamiprid group (p < 0.05). |
| Imidacloprid | Severe intracytoplasmic 8-OHdG and Caspase-3 expressions were detected. |
| Imidacloprid + Probiotics | Slight intracytoplasmic 8-OHdG and Caspase-3 expressions were detected. These results are significantly different compared to Imidacloprid group (p < 0.05). |
| Animal Groups | 8-OHdG | Caspase 3 |
|---|---|---|
| Control | 19.82 ± 0.18 | 20.33 ± 0.7 |
| Probiotics | 19.13 ± 0.56 | 19.76 ± 0.53 |
| Acetamiprid | 69.57 ± 1.18 * | 60.38 ± 0.92 * |
| Acetamiprid + Probiotics | 42.95 ± 1.09 * | 37.55 ± 0.81 * |
| Imidacloprid | 58.12 ± 1.26 * | 50.69 ± 1.23 * |
| Imidacloprid + Probiotics | 30.06 ± 0.85 * | 25.48 ± 0.79 * |
| Groups | Total Testis Weight (TTW) (g) | Total Cauda Epididymis Weights (TCEW) (g) | Motility (%) | Density (103/mL) |
|---|---|---|---|---|
| Control | 3.68 ± 0.14 | 0.56 ± 0.01 | 58.12 ± 2.66 | 60,000 ± 2500 |
| Probiotics | 3.86 ± 0.16 | 0.59 ± 0.03 | 58.75 ± 3.62 | 42,500 ± 1336.30 |
| Acetamiprid | 3.85 ± 0.15 | 0.57 ± 0.01 | 62.50 ± 2.83 | 50,625 ± 1132.90 |
| Acetamiprid + Probiotic | 3.93 ± 0.15 | 0.59 ± 0.01 | 53.75 ± 2.63 | 43,125 ± 1619.49 |
| Imidacloprid | 3.65 ± 0.11 | 0.57 ± 0.01 | 62.12 ± 2.28 | 59,687 ± 2929.12 |
| Imidacloprid + Probiotic | 3.73 ± 0.08 | 0.58 ± 0.02 | 51.25 ± 3.09 | 49,375 ± 2902.81 |
| Groups | Dead Sperm (%) | Head Abnormality (%) | Mid-Piece Abnormality (%) | Tail Abnormality (%) |
|---|---|---|---|---|
| Control | 14.57 ± 2.36 | 2.07 ± 0.46 | 0.14 ± 0.09 | 0.42 ± 0.13 |
| Probiotics | 20.00 ± 1.77 | 2.50 ± 0.72 | 0.62 ± 0.20 | 1.00 ± 0.40 |
| Acetamiprid | 14.64 ± 1.01 | 2.07 ± 0.57 | 0.14 ± 0.09 | 0.57 ± 0.25 |
| Acetamiprid + Probiotic | 16.18 ± 0.85 | 2.56 ± 0.30 | 0.37 ± 0.15 | 0.93 ± 0.34 |
| Imidacloprid | 14.75 ± 0.79 | 1.12 ± 0.22 | 0.18 ± 0.13 | 0.62 ± 0.22 |
| Imidacloprid + Probiotic | 14.43 ± 1.43 | 1.75 ± 0.26 | 0.18 ± 0.09 | 0.31 ± 0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevim, Ç.; Akpınar, E.; Aksu, E.H.; Ömür, A.D.; Yıldırım, S.; Kara, M.; Bolat, İ.; Tsatsakis, A.; Mesnage, R.; Golokhvast, K.S.; et al. Reproductive Effects of S. boulardii on Sub-Chronic Acetamiprid and Imidacloprid Toxicity in Male Rats. Toxics 2023, 11, 170. https://doi.org/10.3390/toxics11020170
Sevim Ç, Akpınar E, Aksu EH, Ömür AD, Yıldırım S, Kara M, Bolat İ, Tsatsakis A, Mesnage R, Golokhvast KS, et al. Reproductive Effects of S. boulardii on Sub-Chronic Acetamiprid and Imidacloprid Toxicity in Male Rats. Toxics. 2023; 11(2):170. https://doi.org/10.3390/toxics11020170
Chicago/Turabian StyleSevim, Çiğdem, Erol Akpınar, Emrah Hicazi Aksu, Ali Doğan Ömür, Serkan Yıldırım, Mehtap Kara, İsmail Bolat, Aristides Tsatsakis, Robin Mesnage, Kirill S. Golokhvast, and et al. 2023. "Reproductive Effects of S. boulardii on Sub-Chronic Acetamiprid and Imidacloprid Toxicity in Male Rats" Toxics 11, no. 2: 170. https://doi.org/10.3390/toxics11020170