

Evaluation of the Toxicity of Microcyclamide Produced by Microcystis aeruginosa in Danio rerio Embryos
 , and
, and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Solvents, Reagents, and Standards
2.2. Cyanobacterial Strain
2.3. Microcyclamide Extraction for Isolation and Purification
2.4. Isolation and Purification of Microcyclamide
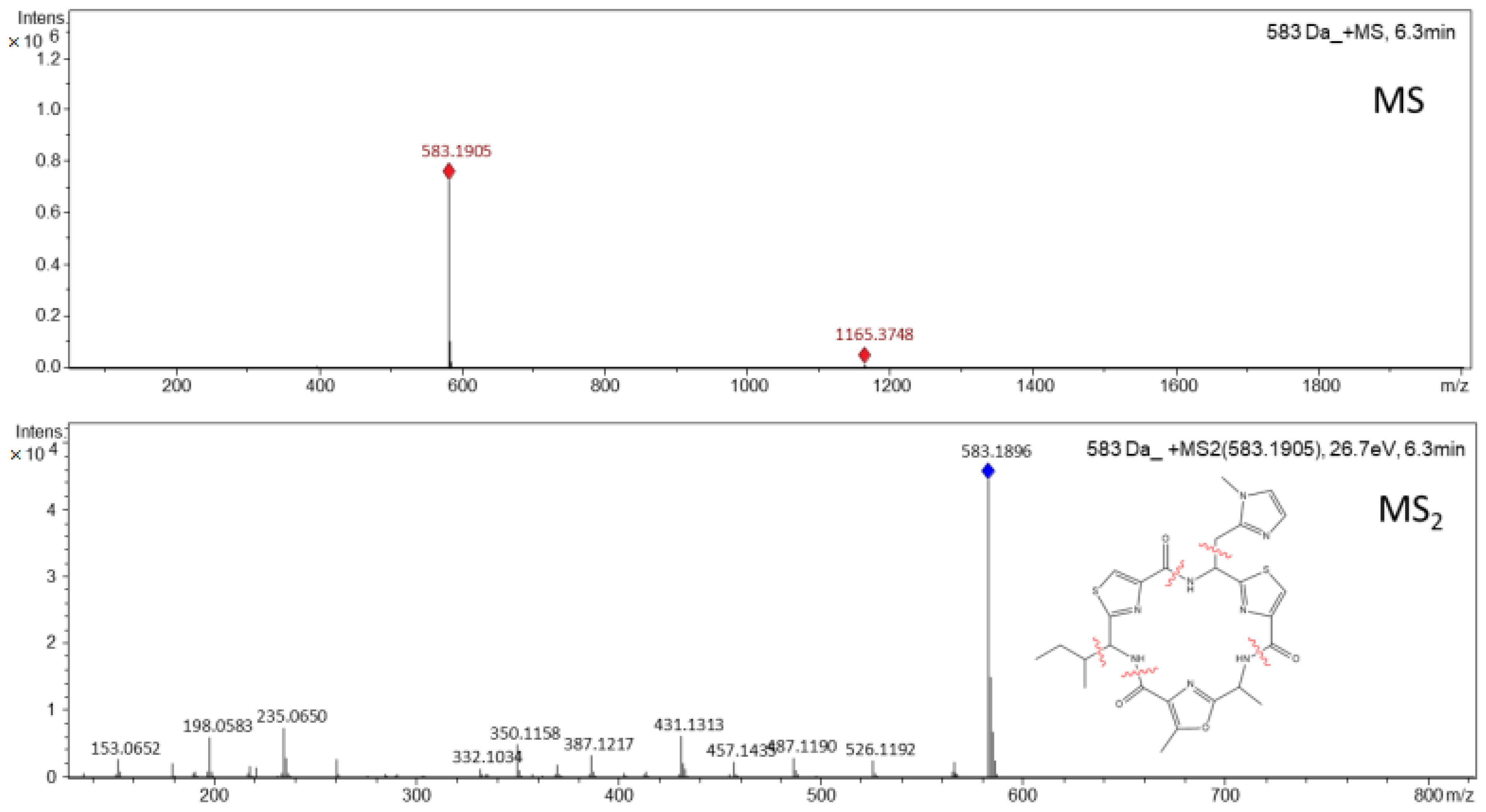
2.5. Detection and Structural Characterization of Microcyclamide Using Mass Spectrometry
2.6. Evaluation of the Acute Toxicity of Microcyclamide with D. rerio Embryos
2.7. Statistical Analysis
3. Results and Discussion
3.1. Isolation and Purification of Microcyclamide
3.2. Evaluation of the Toxicity of Microcyclamide Produced by M. aeruginosa (LTPNA 08) in D. rerio
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and threats of contamination on aquatic ecosystems. In Bioremediation and Biotechnology—Sustainable Approaches to Pollution Degradation; Hakeem, K.R., Bhat, R.A., Qadri, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–26. [Google Scholar]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Reinl, K.L.; Brookes, J.D.; Carey, C.C.; Harris, T.D.; Ibeling, B.W.; Morales-Williams, A.M.; Domis, L.N.S.; Atkins, K.S.; Isles, P.D.F.; Mesman, J.P.; et al. Cyanobacterial blooms in oligotrophic lakes: Shifting the high-nutrient paradigma. Freshw. Biol. 2021, 66, 1846–1859. [Google Scholar] [CrossRef]
- Cook, K.V.; Li, C.; Cai, H.; Krumholz, L.R.; Hambright, K.D.; Paerl, H.W.; Steffen, M.M.; Wilson, A.E.; Burford, M.A.; Grossart, H.; et al. The global Microcystis interactome. Limnol. Oceanogr. 2020, 65, S194–S207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Giesy, J.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovick, B.; Shi, T.; Tuo, X.; Li, S.C.; et al. Challenges of using blooms of Microcystis spp. in animal feeds: A comprehensive review of nutritional, toxicological and microbial health evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.M.L. Cyanobacterial peptides beyond microcystins—A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Boas, L.A.V.; Senra, M.V.X.; Fernandes, K.; Gomes, A.M.A.; Dias, R.J.P.; Pinto, E.; Fonseca, A.L. In vitro toxicity of isolated strains and cyanobacterial bloom biomasses over Paramecium caudatum (ciliophora): Lessons from a non-metazoan model organism. Ecotoxicol. Environ. Saf. 2020, 202, 110937. [Google Scholar] [CrossRef]
- Fernandes, K.; Gomes, A.; Calado, L.; Yasui, G.; Assis, D.; Henrique, T.; Fonseca, A.; Pinto, E. Toxicity of cyanopeptides from two Microcystis strains on larval development of Astyanax altiparanae. Toxins 2019, 11, 220. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Nakagawa, H.E.; Murakami, M. Microcyclamide, a cytotoxic cyclic hexapeptide from the cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2000, 63, 1315–1317. [Google Scholar] [CrossRef]
- Ziemert, N.; Ishida, K.; Quillardet, P.; Bouchier, C.; Hertweck, C.; De Marsac, N.T.E.; Dittmann, E. Microcyclamide biosynthesis in two strains of Microcystis aeruginosa: From structure to genes and vice versa. Appl. Environ. Microbiol. 2008, 74, 1791–1797. [Google Scholar] [CrossRef] [Green Version]
- Portmann, C.; Blom, J.F.; Gademann, K.E.; Jüttner, F. Aerucyclamides A and B: Isolation and synthesis of toxic ribosomal heterocyclic peptides from the cyanobacterium Microcystis aeruginosa PCC 7806. J. Nat. Prod. 2008, 71, 1193–1196. [Google Scholar] [CrossRef]
- Wang, Z.; Akbar, S.; Sun, Y.; Gu, L.; Zhang, L.; Lyu, K.; Huang, Y.; Yang, Z. Cyanobacterial dominance and succession: Factors, mechanisms, predictions, and managements. J. Environ. Manag. 2021, 297, 113281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, J.; Xiao, Y.; Zhang, Y.; Yu, Y.; Zheng, Z.; Liu, Y.; Li, Q. The impact of cyanobacteria blooms on the aquatic environment and human health. Toxins 2022, 14, 658. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.L.; Dörr, F.A.; Dörr, F.; Bortoli, S.; Delherbe, N.; Vásquez, M.; Pinto, E. Co-occurrence of microcystin na microginin congeners in Brazilian strains of Microcystis sp. FEMS Microbiol. 2012, 82, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Gorham, P.R.; Mclachlan, J.; Hammer, U.T.; Kim, W.K. Isolation and culture of toxic strains of Anabaena Flo-aquae (Lyngb.) de Bréb. Verh. Int. Ver. Theor. Angew. Limnol. 1964, 15, 796–804. [Google Scholar] [CrossRef]
- Sanz, M.; Salina, R.K.; Pinto, E. Namalides B and C and spumigins K-N from the culturee freshwater cyanobacterium Sphaerospermopsis torques-reginae. J. Nat. Prod. 2017, 80, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Paiva, C.R.F.; Ferreira, M.G.; Trossini, H.G.G.; Pinto, E. Identification, in vitro testing and molecular docking studies of microginins’ mechanism of angiotensin—Coverting enzyme inhibition. Molecules 2017, 22, 1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, M.; Andreote, P.A.; Fiore, F.M.; Dörr, A.F.; Pinto, E. Structural characterization of new peptide variants produced by cyanobacteria from the Brazilian Atlantic Coastal Forest using liquid chromatography coupled to quadrupole time-of-flight tandem mass spectrometry. Mar. Drugs 2015, 13, 3892–3919. [Google Scholar] [CrossRef]
- Paizs, M.; Suhai, S. Fragmentation pathways of protonated peptides. Mass Spectrom. Rev. 2004, 24, 508–548. [Google Scholar] [CrossRef]
- Cantú, M.D.; Carrilho, E.; Wulff, N.A.; Palmas, M.S. Sequenciamento de peptídeos usando espectrometria de massas: Um guia prático. Quím. Nova 2008, 17, 669–675. [Google Scholar] [CrossRef]
- OECD Guidelines for the Testing of Chemicals, Section 2. Test No. 236: Fish Embryo Acute Toxicity (FET) Test. Available online: https://www.oecd-ilibrary.org/environment/test-no-236-fish-embryo-acute-toxicity-fet-test_9789264203709-en (accessed on 10 November 2022).
- Raveh, A.; Moshe, S.; Evron, Z.; Flescher, E.E.; Carmeli, S. Novel thiazole and oxazole containing cyclic hexapeptides from a waterbloom of the cyanobacterium Microcystis sp. Tetrahedron 2010, 66, 2705–2712. [Google Scholar] [CrossRef]
- Le Manach, S.; Duval, C.; Marie, A.; Djediat, C.; Catherine, A.; Edery, M.; Bernard, C.E.; Marie, B. Global metabolomic characterizations of Microcystis spp. highlights clonal diversity in natural bloom-forming populations and expands metabolite structural diversity. Front. Microbiol. 2019, 10, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorova, A.K.; Juettner, F.; Linden, A.; Pluess, T.E.; Von Philipsborn, W. Nostocyclamide: A new macrocyclic, thiazole-containing allelochemical from Nostoc sp. 31 (cyanobacteria). J. Org. Chem. 1995, 60, 7891–7895. [Google Scholar] [CrossRef]
- Jüttner, F.; Todorova, A.K.; Walch, N.E.; Von Philipsborn, W. Nostocyclamide M: A cyanobacterial cyclic peptide with allelopathic activity from Nostoc 31. Phytochemistry 2001, 57, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Portmann, C.; Blom, J.F.; Kaiser, M.; Brun, R.; Jüttner, F.E.; Gademann, K. Isolation of aerucyclamides C and D and structure revision of microcyclamide 7806A: Heterocyclic ribosomal peptides from Microcystis aeruginosa PCC 7806 and their antiparasite evaluation. J. Nat. Prod. 2008, 71, 1891–1896. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Nelson, J.T.; Rasko, D.A.; Sudek, S.; Eisen, J.A.; Haygood, M.G.E.; Ravel, J. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc. Natl. Acad. Sci. USA 2005, 102, 7315–7320. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Kato, T.; Murakami, M.; Watanabe, M.; Watanabe, M.F. Microginins, zinc metalloproteases inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron 2000, 56, 8643–8656. [Google Scholar] [CrossRef]
- Sadler, T.E.; Von Elert, E. Physiological interaction of Daphnia and Microcystis with regard to cyanobacterial secondary metabolites. Aquat. Toxicol. 2014, 156, 96–105. [Google Scholar] [CrossRef]
- Shahmohamadloo, R.S.; Poirier, D.G.; Almirall, X.O.; Bhavsar, S.P.; Sibley, P.K. Assessing the toxicity of cell-bound microcystins on freshwater pelagic and benthic invertebrates. Ecotoxicol. Environ. Saf. 2020, 188, 109945. [Google Scholar] [CrossRef]
- Gademann, K.; Portmann, C.; Blom, J.F.; Zeder, M.; Jüttner, F. Multiple toxin production in the cyanobacterium Microcystis: Isolation of the toxic protease inhibitor cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [Green Version]
- Bober, B.; Bialczyk, J. Determination of the toxicity of the freshwater cyanobacterium Woronichinia naegeliana (Unger) Elenkin. J. Appl. Phycol. 2017, 29, 1355–1362. [Google Scholar] [CrossRef]
- Kohler, E.; Grundler, V.; Häussinger, D.; Kurmayer, R.; Gademann, K.; Pernthaler, J.; Blom, J.F. The toxicity and enzyme activity of a chlorine and sulfate containing aeruginosin isolated from a non-microcystin-producing Planktothrix strain. Harmful Algae 2014, 39, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujvárosi, A.Z.; Hercog, K.; Riba, M.; Gonda, S.; Filipič, M.; Vasas, G.E.; Žegura, B. The cyanobacterial oligopeptides microginins induce DNA damage in the human hepatocellular carcinoma (HepG2) cell line. Chemosphere 2020, 240, 124880. [Google Scholar] [CrossRef] [PubMed]
- Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L.; Miller, T.R. Variable cyanobacterial toxin and metabolite profiles across six eutrophic lakes of differing physiochemical characteristics. Toxins 2017, 9, 62. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Organism | Cyanobacteria (Dry Weight) * or Compound | Value of LC50 | Reference |
|---|---|---|---|
| Thamnocephalus platyurus | Cyanopeptolin 1020 | 8.8 μM | [31] |
| A. altiparanae | M. panniformis MIRS-04 (producer of microcystins and micropeptins) * | 0.24 mg mL−1 | [8] |
| A. altiparanae | M. aeruginosa NPCD-01 (producer of microginins) * | 0.32 mg mL−1 | [8] |
| C. dubia | Microcystin-LR and [D-Asp3]-microcystin-LR | 5.53 μg L−1 | [30] |
| D. magna | Microcystin-LR and [D-Asp3]-microcystin-LR | 85.72 μg L−1 | [30] |
| Plasmodium falciparum | Aerucyclamides A, B, C and D) | 5.0, 0.7, 2.3 and 6.3 μM, respectively | [26] |
| T. platyurus | Woronichinia naegeliana (producer of microginin MG-FR3) * | 7.78 μg mL−1 | [32] |
| T. platyurus | Aeruginosin 828A | 22.4 μM | [33] |
| D. rerio embryos | Microcyclamide | 42.98 (with a concentration range of 37.79–48.89) µg mL−1 | [This study] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nathane Nunes de Freitas, P.; Kinoshita Teramoto, K.; Ossanes de Souza, A.; Pinto, E. Evaluation of the Toxicity of Microcyclamide Produced by Microcystis aeruginosa in Danio rerio Embryos. Toxics 2023, 11, 128. https://doi.org/10.3390/toxics11020128
Nathane Nunes de Freitas P, Kinoshita Teramoto K, Ossanes de Souza A, Pinto E. Evaluation of the Toxicity of Microcyclamide Produced by Microcystis aeruginosa in Danio rerio Embryos. Toxics. 2023; 11(2):128. https://doi.org/10.3390/toxics11020128
Chicago/Turabian StyleNathane Nunes de Freitas, Paloma, Kazumi Kinoshita Teramoto, Alexander Ossanes de Souza, and Ernani Pinto. 2023. "Evaluation of the Toxicity of Microcyclamide Produced by Microcystis aeruginosa in Danio rerio Embryos" Toxics 11, no. 2: 128. https://doi.org/10.3390/toxics11020128