
South American National Contributions to Knowledge of the Effects of Endocrine Disrupting Chemicals in Wild Animals: Current and Future Directions
 , , ,
, , ,
Abstract
:1. Introduction
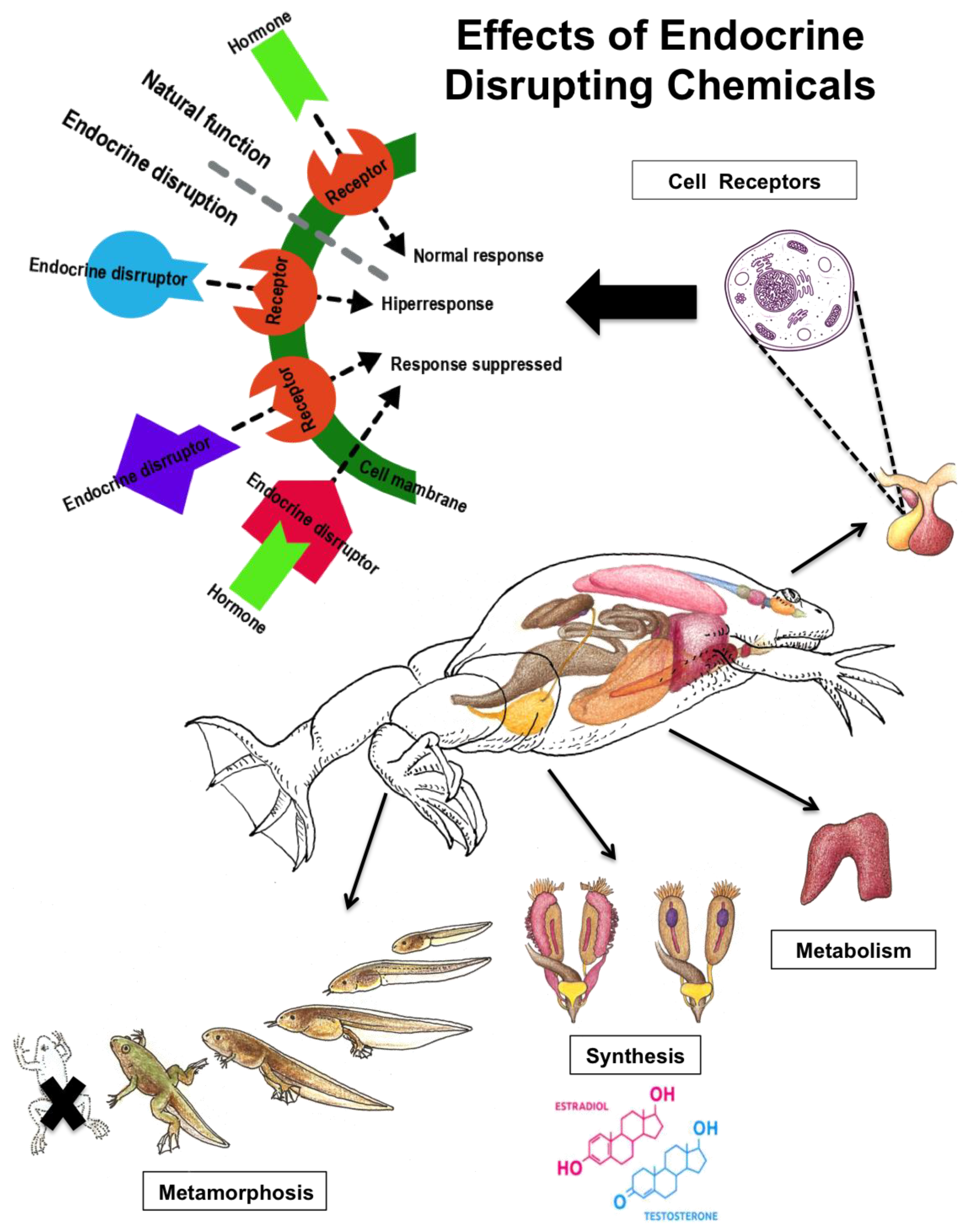
1.1. Endocrine Regulation and Effects of Xenobioc Chemicals
1.1.1. The Hypothalamic-Pituitary-Gonadal (HPG) Axis
1.1.2. The Hypothalamic-Pituitary-Thyroid Axis
1.2. The Use of Biomarkers of Endocrine Disruptors
2. Materials and Methods
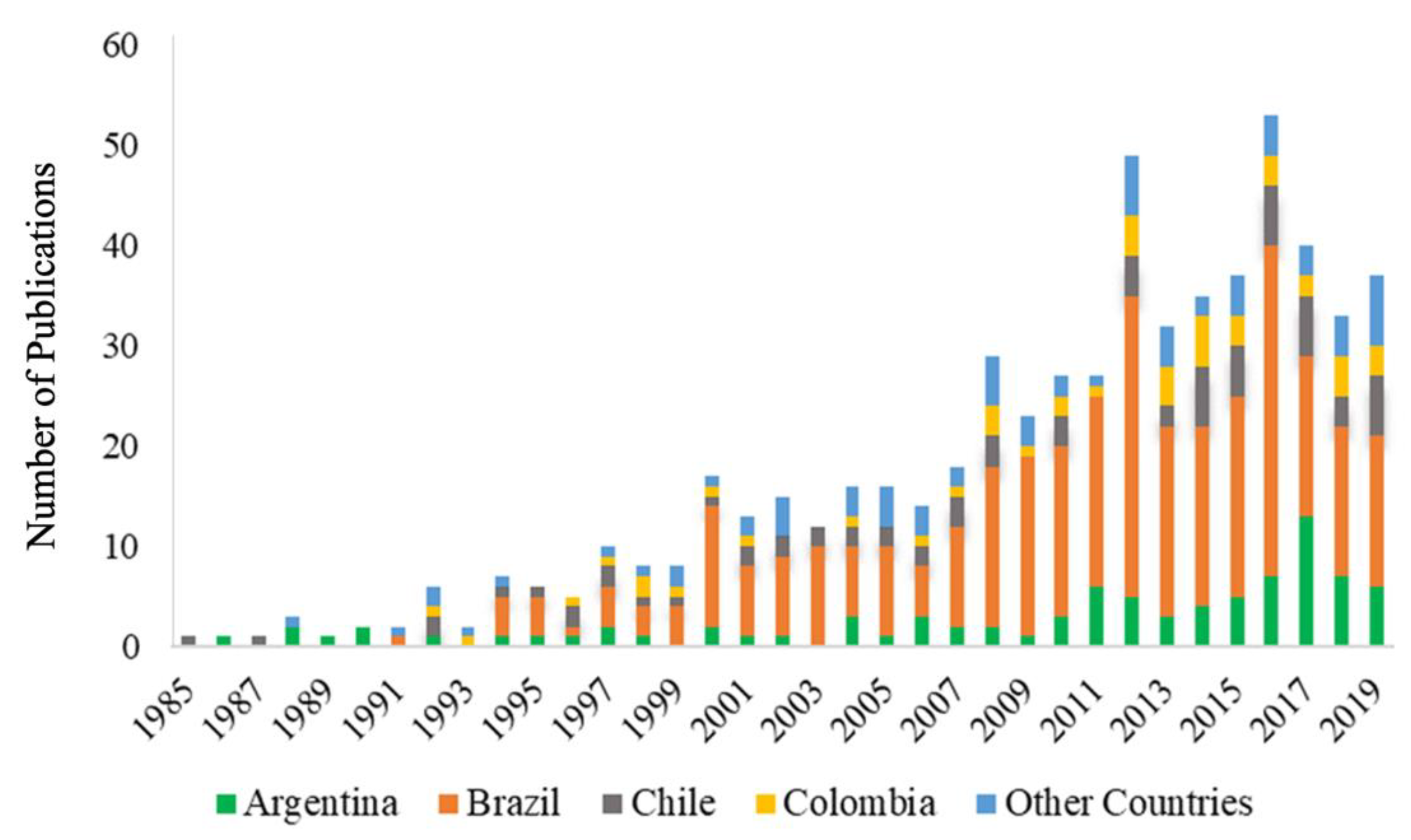
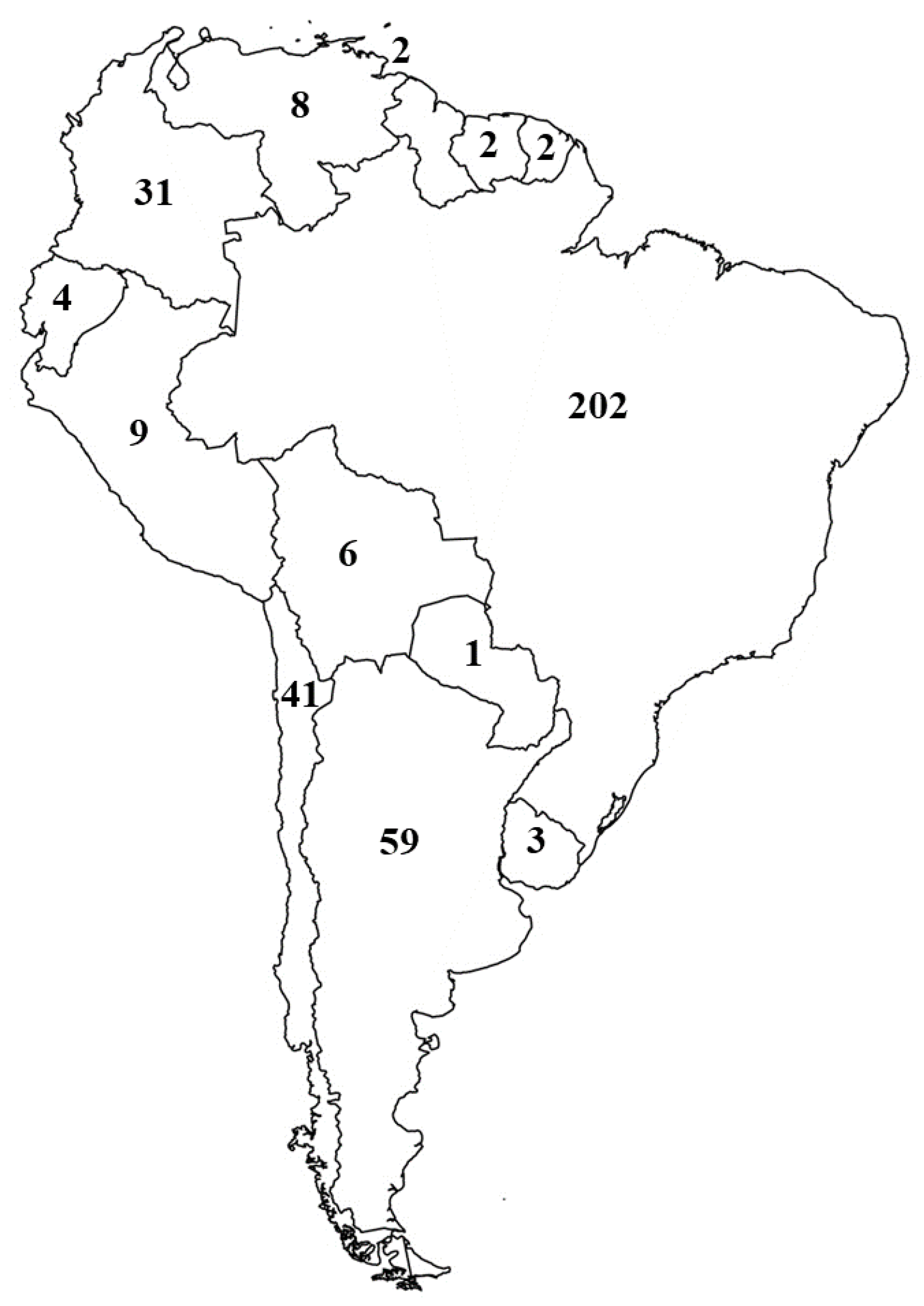
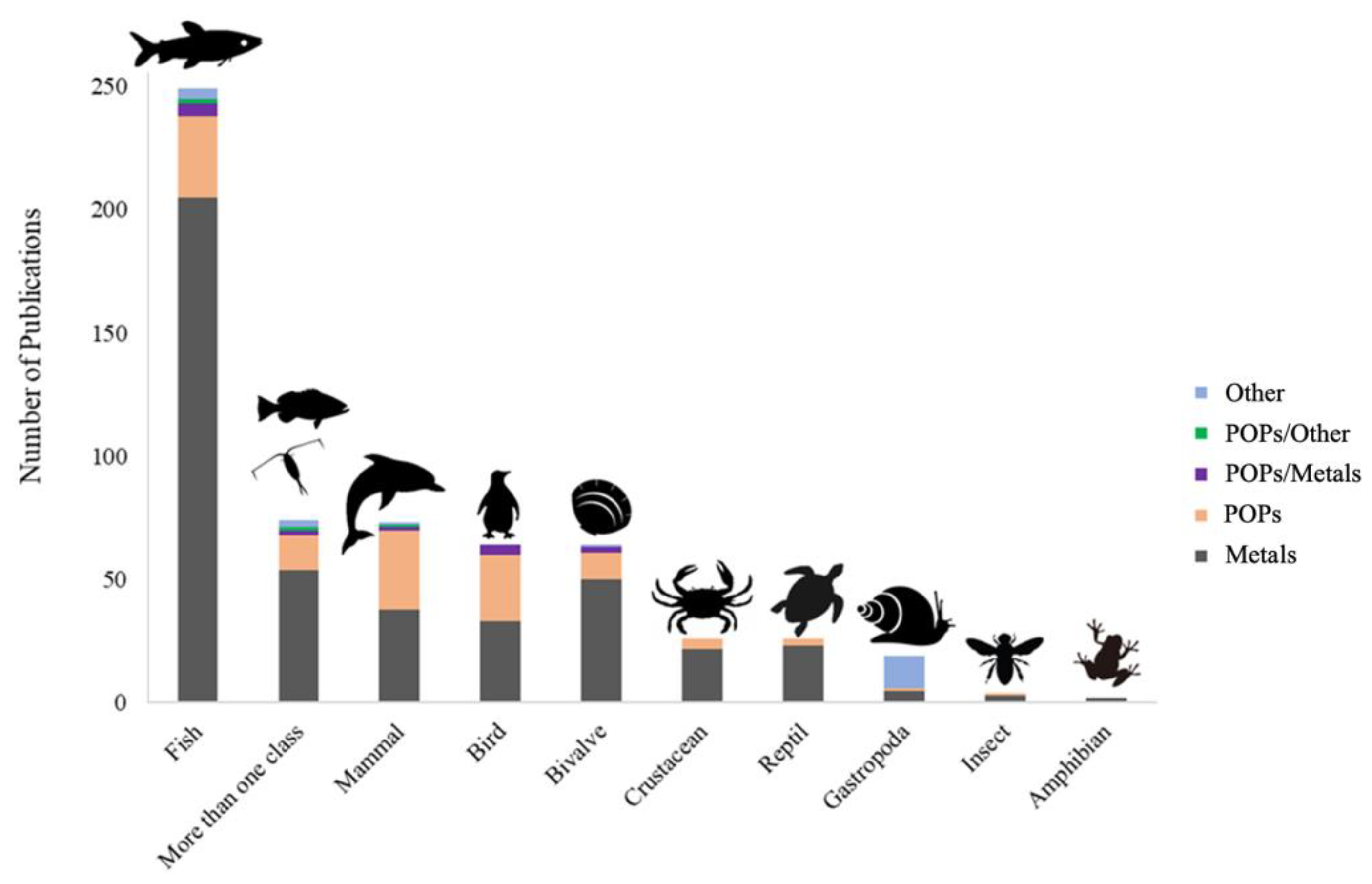
3. Results and Discussion
3.1. Argentina
3.2. Brazil
3.3. Chile
3.4. Colombia
3.5. Venezuela
3.6. Other Countries
4. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diamanti-Kandarakis, E.; Bourguignon, J.-P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-Disrupting Chemicals: An Endocrine Society Scientific Statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic Transgenerational Actions of Endocrine Disruptors. Reprod. Toxicol. 2011, 31, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Inostroza, L.; Baur, R.; Csaplovics, E. Urban Sprawl and Fragmentation in Latin America: A Dynamic Quantification and Characterization of Spatial Patterns. J. Environ. Manag. 2013, 115, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Ceddia, M.G.; Bardsley, N.O.; Gomez-y-Paloma, S.; Sedlacek, S. Governance, Agricultural Intensification, and Land Sparing in Tropical South America. Proc. Natl. Acad. Sci. USA 2014, 111, 7242–7247. [Google Scholar] [CrossRef] [Green Version]
- De Sy, V.; Herold, M.; Achard, F.; Beuchle, R.; Clevers, J.; Lindquist, E.; Verchot, L. Land Use Patterns and Related Carbon Losses Following Deforestation in South America. Environ. Res. Lett. 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Jara-Samaniego, J.; Moral, M.R.; Perez-Murcia, D.; Paredes, C.; Gálvez-Sola, L.; Gavilanes-Terán, I.; Bustamante, M.Á. Urban Waste Management and Potential Agricultural Use in South American Developing Countries: A Case Study of Chimborazo Region (Ecuador). Commun. Soil Sci. Plant. Anal. 2015, 46, 157–169. [Google Scholar] [CrossRef]
- Laurance, W.F.; Sayer, J.; Cassman, K.G. Agricultural Expansion and Its Impacts on Tropical Nature. Trends Ecol. Evol. 2014, 29, 107–116. [Google Scholar] [CrossRef]
- Barra, R.; Colombo, J.C.; Eguren, G.; Gamboa, N.; Jardim, W.F. Persistent Organic Pollutants (POPs) in Eastern and Western South American Countries. Rev. Environ. Contam. Toxicol. 2006, 185, 1–33. [Google Scholar] [CrossRef]
- Furley, T.H.; Brodeur, J.; Silva de Assis, H.C.; Carriquiriborde, P.; Chagas, K.R.; Corrales, J.; Denadai, M.; Fuchs, J.; Mascarenhas, R.; Miglioranza, K.S.; et al. Toward Sustainable Environmental Quality: Identifying Priority Research Questions for Latin America. Integr. Environ. Assess. Manag. 2018, 14, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Harfoot, M.B.J.; Tittensor, D.P.; Knight, S.; Arnell, A.P.; Blyth, S.; Brooks, S.; Butchart, S.H.M.; Hutton, J.; Jones, M.I.; Kapos, V.; et al. Present and Future Biodiversity Risks from Fossil Fuel Exploitation. Conserv. Lett. 2018, 11, e12448. [Google Scholar] [CrossRef]
- Hunt, L.; Bonetto, C.; Resh, V.H.; Buss, D.F.; Fanelli, S.; Marrochi, N.; Lydy, M.J. Insecticide Concentrations in Stream Sediments of Soy Production Regions of South America. Sci. Total Environ. 2016, 547, 114–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uglietti, C.; Gabrielli, P.; Cooke, C.A.; Vallelonga, P.; Thompson, L.G. Widespread Pollution of the South American Atmosphere Predates the Industrial Revolution by 240 Y. Proc. Natl. Acad. Sci. USA 2015, 112, 2349–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassman, K.G.; Dobermann, A.R.; Walters, D.T. Agroecosystems, Nitrogen-Use Efficiency, and Nitrogen Management. AMBIO J. Hum. Environ. 2002, 31, 132–140. [Google Scholar] [CrossRef]
- Fasola, E.; Ribeiro, R.; Lopes, I. Microevolution Due to Pollution in Amphibians: A Review on the Genetic Erosion Hypothesis. Environ. Pollut. 2015, 204, 181–190. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, X.; Chen, Y.; Fu, R.; Du, X.; Chen, X.; Zhan, A. Zooplankton Biodiversity Monitoring in Polluted Freshwater Ecosystems: A Technical Review. Environ. Sci. Ecotechnol. 2020, 1, 663–679. [Google Scholar] [CrossRef]
- Galli, A.; Wackernagel, M.; Iha, K.; Lazarus, E. Ecological Footprint: Implications for Biodiversity. Biol. Conserv. 2014, 173, 121–132. [Google Scholar] [CrossRef]
- Naidu, R.; Arias Espana, V.A.; Liu, Y.; Jit, J. Emerging Contaminants in the Environment: Risk-Based Analysis for Better Management. Chemosphere 2016, 154, 350–357. [Google Scholar] [CrossRef]
- Norris, D.O.; Carr, J.A. Endocrine Disruption: Biological Bases for Health Effects in Wildlife and Humans; Oxford University Press, Inc.: New York, NY, USA, 2006; ISBN 9780195137491. [Google Scholar]
- Yurdakök, K. Environmental Pollution and the Fetus. J. Pediatr. Neonatal Individ. Med. 2012, 1, 33–42. [Google Scholar] [CrossRef]
- Gavrilescu, M.; Demnerová, K.; Aamand, J.; Agathos, S.; Fava, F. Emerging Pollutants in the Environment: Present and Future Challenges in Biomonitoring, Ecological Risks and Bioremediation. Nat. Biotechnol. 2014, 32, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, S.M.; Gore, A.C. Estrogenic Environmental Endocrine-Disrupting Chemical Effects on Reproductive Neuroendocrine Function and Dysfunction across the Life Cycle. Rev. Endocr. Metab. Disord. 2007, 8, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, H.J.; Guillette, L.J. Birth Defects in Wildlife: The Role of Environmental Contaminants as Inducers of Reproductive and Developmental Dysfunction. Syst. Biol. Reprod. Med. 2010, 56, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Rodriguez-Jorquera, I.A.; Silva-Sanchez, C.; Strynar, M.; Denslow, N.D.; Toor, G.S. Footprints of Urban Micro-Pollution in Protected Areas: Investigating the Longitudinal Distribution of Perfluoroalkyl Acids in Wildlife Preserves. PLoS ONE 2016, 11, e0148654. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jorquera, I.A.; Siroski, P.; Espejo, W.; Nimptsch, J.; Choueri, P.G.; Choueri, R.B.; Moraga, C.A.; Mora, M.; Toor, G.S. Latin American Protected Areas: Protected from Chemical Pollution? Integr. Environ. Assess. Manag. 2016, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Kim, J.; Kim, Y. Identification of Linkages between EDCs in Personal Care Products and Breast Cancer through Data Integration Combined with Gene Network Analysis. Int. J. Environ. Res. Public Health 2017, 14, 1158. [Google Scholar] [CrossRef] [Green Version]
- Tubbs, C.W.; McDonough, C.E. Reproductive Impacts of Endocrine-Disrupting Chemicals on Wildlife Species: Implications for Conservation of Endangered Species. Annu. Rev. Anim. Biosci. 2018, 6, 287–304. [Google Scholar] [CrossRef]
- Hamlin, H.J.; Guillette, L.J. Embryos as Targets of Endocrine Disrupting Contaminants in Wildlife. Birth Defects Res. Part. C Embryo Today Rev. 2011, 93, 19–33. [Google Scholar] [CrossRef]
- Ternes, T.A.; Joss, A.; Siegrist, H. Peer Reviewed: Scrutinizing Pharmaceuticals and Personal Care Products in Wastewater Treatment. Environ. Sci. Technol. 2004, 38, 392A–399A. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajda, A.M.; Norris, D.O. Endocrine-active chemicals (EACs) in fishes. In Hormones and Reproduction of Vertebrates; Norris, D.O., Lopez, K.H., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 1, pp. 245–264. [Google Scholar]
- Iavicoli, I.; Fontana, L.; Bergamaschi, A. The Effects of Metals as Endocrine Disruptors. J. Toxicol. Environ. Health Part B Crit. Rev. 2009, 12, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Schug, T.T.; Johnson, A.F.; Birnbaum, L.S.; Colborn, T.; Guillette, L.J.; Crews, D.P.; Collins, T.; Soto, A.M.; vom Saal, F.S.; McLachlan, J.A.; et al. Minireview: Endocrine Disruptors: Past Lessons and Future Directions. Mol. Endocrinol. 2016, 30, 833–847. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Palioura, E.; Kandarakis, S.A.; Koutsilieris, M. The Impact of Endocrine Disruptors on Endocrine Targets. Horm. Metab. Res. 2010, 42, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Denoncourt, J.; Wallace, S.J.; De Solla, S.R.; Langlois, V.S. Plasticizer Endocrine Disruption: Highlighting Developmental and Reproductive Effects in Mammals and Non-Mammalian Aquatic Species. Gen. Comp. Endocrinol. 2015, 219, 74–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnif, W.; Hassine, A.I.H.; Bouaziz, A.; Bartegi, A.; Thomas, O.; Roig, B. Effect of Endocrine Disruptor Pesticides: A Review. Int. J. Environ. Res. Public Health 2011, 8, 2265–2303. [Google Scholar] [CrossRef] [Green Version]
- Schug, T.T.; Janesick, A.; Blumberg, B.; Heindel, J.J. Endocrine Disrupting Chemicals and Disease Susceptibility. J. Steroid Biochem. Mol. Biol. 2011, 127, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Dyer, C.A. Heavy metals as endocrine-disrupting chemicals. In Endocrine-Disrupting Chemicals: From Basic Research to Clinical Practice; Gore, A.C., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 111–133. [Google Scholar]
- Meyer, E.; Eagles-Smith, C.A.; Sparling, D.; Blumenshine, S. Mercury Exposure Associated with Altered Plasma Thyroid Hormones in the Declining Western Pond Turtle (Emys Marmorata) from California Mountain Streams. Environ. Sci. Technol. 2014, 48, 2989–2996. [Google Scholar] [CrossRef]
- Casals-Casas, C.; Desvergne, B. Endocrine Disruptors: From Endocrine to Metabolic Disruption. Annu. Rev. Physiol. 2011, 73, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, X.; Zhang, J.; Wu, K. Impact of Endocrine-Disrupting Chemicals on Reproductive Function in Zebrafish (Danio Rerio). Reprod. Domest. Anim. 2015, 50, 1–6. [Google Scholar] [CrossRef]
- Sumpter, J.P. Endocrine Disrupters in the Aquatic Environment: An Overview. Acta Hydrochim. Hydrobiol. 2005, 33, 9–16. [Google Scholar] [CrossRef]
- Argemi, F.; Cianni, N.; Porta, A. Disrupción Endocrina: Perspectivas Ambientales y Salud Pública. Acta Bioquím. Clín. Latinoam. 2005, 39, 291–300. [Google Scholar]
- Chichizola, C.; Scaglia, H. Disruptores Endócrinos y El Sistema Reproductivo. Bioquím. Patol. Clin. 2009, 73, 9–23. [Google Scholar]
- De Falco, M.; Forte, M.; Laforgia, V. Estrogenic and Anti-Androgenic Endocrine Disrupting Chemicals and Their Impact on the Male Reproductive System. Front. Environ. Sci. 2015, 3, 1–27. [Google Scholar] [CrossRef]
- Edwards, T.M.; Myers, J.P. Environmental Exposures and Gene Regulation in Disease Etiology. Environ. Health Perspect. 2007, 115, 1264–1270. [Google Scholar] [CrossRef]
- Gonzalez-Mille, D.J.; Espinosa-Reyes, G.; Rivero-Pérez, N.E.; Trejo-Acevedo, A.; Nava-Montes, A.D.; Ilizaliturri-Hernández, C.A. Persistent Organochlorine Pollutants (POPs) and DNA Damage in Giant Toads (Rhinella Marina) from an Industrial Area at Coatzacoalcos, Mexico. Water Air Soil Pollut. 2013, 224, 1–14. [Google Scholar] [CrossRef]
- Hoffmann, F.; Kloas, W. Estrogens Can Disrupt Amphibian Mating Behavior. PLoS ONE 2012, 7, e32097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waye, A.; Trudeau, V.L. Neuroendocrine Disruption: More than Hormones Are Upset. J. Toxicol. Environ. Health Part B Crit. Rev. 2011, 14, 270–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organisation for Economic Cooperation and Development. Detailed Review Paper State of the Science on Novel In Vitro and In Vivo Screening and Testing Methods and Endpoints for Evaluating Endocrine Disruptors; OECD Publishing: Paris, France, 2012. [Google Scholar]
- Nabi, G.; Hao, Y.; Liu, X.; Sun, Y.; Wang, Y.; Jiang, C. Hypothalamic—Pituitary—Thyroid Axis Crosstalk With the Hypothalamic—Pituitary—Gonadal Axis and Metabolic Regulation in the Eurasian Tree Sparrow During Mating and Non-Mating Periods. Front. Endocrinol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Castañeda Cortés, D.C.; Langlois, V.S.; Fernandino, J.I. Crossover of the Hypothalamic Pituitary-Adrenal/Interrenal, -Thyroid, and -Gonadal Axes in Testicular Development. Front. Endocrinol. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.Y.; Zhang, W.; Zhuo, Z.J.; He, J.Y.; Yin, Z. Neuroendocrine Regulation of Somatic Growth in Fishes. Sci. China Life Sci. 2015, 58, 137–147. [Google Scholar] [CrossRef]
- Duarte-Guterman, P.; Navarro-Martín, L.; Trudeau, V.L. Mechanisms of Crosstalk between Endocrine Systems: Regulation of Sex Steroid Hormone Synthesis and Action by Thyroid Hormones. Gen. Comp. Endocrinol. 2014, 203, 69–85. [Google Scholar] [CrossRef]
- Huang, W.T.; Weng, C.F. Roles of Hepatocyte Nuclear Factors (HNF) in the Regulation of Reproduction in Teleosts. J. Fish. Biol. 2010, 76, 225–239. [Google Scholar] [CrossRef]
- Kloas, W.; Urbatzka, R.; Opitz, R.; Würtz, S.; Behrends, T.; Hermelink, B.; Hofmann, F.; Jagnytsch, O.; Kroupova, H.; Lorenz, C.; et al. Endocrine Disruption in Aquatic Vertebrates. Ann. N. Y. Acad. Sci. 2009, 1163, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Viau, V. Functional Cross-Talk Between the Hypothalamic-Pituitary-Gonadal and -Adrenal Axes. J. Neuroendocrinol. 2002, 14, 506–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khetan, S.K. Endocrine Disruptors in the Environment; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; ISBN 9781118852934. [Google Scholar]
- Kloas, W.; Lutz, I. Amphibians as Model to Study Endocrine Disrupters. J. Chromatogr. A 2006, 1130, 16–27. [Google Scholar] [CrossRef]
- León-Olea, M.; Martyniuk, C.J.; Orlando, E.F.; Ottinger, M.A.; Rosenfeld, C.S.; Wolstenholme, J.T.; Trudeau, V.L. Current Concepts in Neuroendocrine Disruption. Gen. Comp. Endocrinol. 2014, 203, 158–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golshan, M.; Mohammad, S.; Alavi, H. Androgen Signaling in Male Fishes: Examples of Anti-Androgenic Chemicals That Cause Reproductive Disorders. Theriogenology 2019, 139, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, T.D.; Boone, M.D. Atrazine Feminizes Sex Ratio in Blanchard’s Cricket Frogs (Acris Blanchardi) at Concentrations as Low as 0.1 Μg/L. Environ. Toxicol. Chem. 2017, 9999, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.A. Populations Collapses in Marine Invertebrates Due to Endocrine Disruption: A Cause for Concern? Front. Endocrinol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Soler, P.; Bañón, R.; García-Galea, E. Effects of Industrial Pollution on the Reproductive Biology of Squalius Laietanus (Actinopterygii, Cyprinidae) in a Mediterranean Stream (NE Iberian Peninsula). Fish Physiol. Biochem. 2019, 46, 247–264. [Google Scholar] [CrossRef]
- Gore, A.C. Neuroendocrine Targets of Endocrine Disruptors. Hormones 2010, 9, 16–27. [Google Scholar] [CrossRef]
- McCoy, K.A.; Bortnick, L.J.; Campbell, C.M.; Hamlin, H.J.; Guillette, L.J.; St. Mary, C.M. Agriculture Alters Gonadal Form and Function in the Toad Bufo Marinus. Environ. Health Perspect. 2008, 116, 1526–1532. [Google Scholar] [CrossRef] [Green Version]
- Guillette, L.J.; Crain, D.A.; Gunderson, M.P.; Kools, S.A.; Milnes, M.R. Alligators and Endocrine Disrupting Contaminants: A Current Perspective. Am. Zool. 2000, 40, 438–452. [Google Scholar]
- Moore, B.C.; Roark, A.M.; Kohno, S.; Hamlin, H.J.; Guillette, L.J. Gene-Environment Interactions: The Potential Role of Contaminants in Somatic Growth and the Development of the Reproductive System of the American Alligator. Mol. Cell. Endocrinol. 2012, 354, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwindt, A.R. Parental Effects of Endocrine Disrupting Compounds in Aquatic Wildlife: Is There Evidence of Transgenerational Inheritance? Gen. Comp. Endocrinol. 2015, 219, 152–164. [Google Scholar] [CrossRef]
- Bahamonde, P.A.; Fuzzen, M.L.; Bennett, C.J.; Tetreault, G.R.; Mcmaster, M.E.; Servos, M.R.; Martyniuk, C.J.; Munkittrick, K.R. Whole Organism Responses and Intersex Severity in Rainbow Darter (Etheostoma Caeruleum) Following Exposures to Municipal Wastewater in the Grand River Basin, ON, Canada. Part A. Aquat. Toxicol. 2015, 159, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, T. Masculinization of Female Gastropod Mollusks Induced by Organotin Compounds, Focusing on Mechanism of Actions of Tributyltin and Triphenyltin for Development of Imposex. Environ. Sci. 2006, 13, 77–87. [Google Scholar]
- Iguchi, T.; Katsu, Y.; Horiguchi, T.; Watanabe, H.; Blumberg, B.; Ohta, Y. Endocrine Disrupting Organotin Compounds Are Potent Inducers of Imposex in Gastropods and Adipogenesis in Vertebrates. Mol. Cell. Toxicol. 2007, 3, 1–10. [Google Scholar]
- Brown, D.D.; Cai, L. Amphibian Metamorphosis. Dev. Biol. 2007, 306, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Brucker-Davis, F. Effects of Environmental Synthetic Chemicals on Thyroid Function. Thyroid 1998, 8, 827–856. [Google Scholar] [CrossRef]
- Sachs, L.M.; Buchholz, D.R.; Forrest, D. Insufficiency of Thyroid Hormone in Frog Metamorphosis and the Role of Glucocorticoids. Front. Endocrinol. 2019, 10, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.A.; Patiño, R. The Hypothalamus-Pituitary-Thyroid Axis in Teleosts and Amphibians: Endocrine Disruption and Its Consequences to Natural Populations. Gen. Comp. Endocrinol. 2011, 170, 299–312. [Google Scholar] [CrossRef]
- Hapon, M.B.; Gamarra-Luques, C.; Jahn, G.A. Short Term Hypothyroidism Affects Ovarian Function in the Cycling Rat. Reprod. Biol. Endocrinol. 2010, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogan, N.S.; Crump, K.L.; Duarte, P.; Lean, D.R.S.; Trudeau, V.L. Hormone Cross-Regulation in the Tadpole Brain: Developmental Expression Profiles and Effect of T3 Exposure on Thyroid Hormone- and Estrogen-Responsive Genes in Rana Pipiens. Gen. Comp. Endocrinol. 2007, 154, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Mukhi, S.; Patiño, R. Effects of Prolonged Exposure to Perchlorate on Thyroid and Reproductive Function in Zebrafish. Toxicol. Sci. 2007, 96, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.E.D.H.; Mahmoud, U.M.; Mekkawy, I.A. Reproductive Biomarkers to Identify Endocrine Disruption in Clarias Gariepinus Exposed to 4-Nonylphenol. Ecotoxicol. Environ. Saf. 2012, 78, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Xu, W.; Chen, X.R.; Lou, Q.Q.; Wei, W.J.; Qin, Z.F. Low Concentrations of 17β-Trenbolone Induce Female-to-Male Reversal and Mortality in the Frog Pelophylax Nigromaculatus. Aquat. Toxicol. 2015, 158, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H. Chemical Contamination and the Thyroid. Endocrine 2014, 48, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.N.; Braverman, L.E. Environmental Pollutants and the Thyroid. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 801–813. [Google Scholar] [CrossRef]
- Mann, R.M.; Hyne, R.V.; Choung, C.B.; Wilson, S.P. Amphibians and Agricultural Chemicals: Review of the Risks in a Complex Environment. Environ. Pollut. 2009, 157, 2903–2927. [Google Scholar] [CrossRef]
- Veldhoen, N.; Propper, C.R.; Helbing, C.C. Enabling Comparative Gene Expression Studies of Thyroid Hormone Action through the Development of a Flexible Real-Time Quantitative PCR Assay for Use across Multiple Anuran Indicator and Sentinel Species. Aquat. Toxicol. 2014, 148, 162–173. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Wu, D.; Liu, G.; Tao, L.; Fu, W.; Hou, J. Teratogenic Effects of Organic Extracts from the Pearl River Sediments on Xenopus Laevis Embryos. Environ. Toxicol. Pharmacol. 2014, 37, 202–209. [Google Scholar] [CrossRef]
- Tan, S.W.; Meiller, J.C.; Mahaffey, K.R. The Endocrine Effects of Mercury in Humans and Wildlife. Crit. Rev. Toxicol. 2009, 39, 228–269. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kashida, Y.; Yoneda, K.; Iwata, H.; Watanabe, M.; Tanabe, S.; Fukatsu, H.; Machida, N.; Mitsumori, K. Thyroid Lesions and Dioxin Accumulation in the Livers of Jungle Crows (Corvus Macrorhynchos) in Urban and Suburban Tokyo. Arch. Environ. Contam. Toxicol. 2005, 48, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Silins, I.; Högberg, J. Combined Toxic Exposures and Human Health: Biomarkers of Exposure and Effect. Int. J. Environ. Res. Public Health 2011, 8, 629–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fort, D.J.; Degitz, S.; Tietge, J.; Touart, L.W. The Hypothalamic-Pituitary-Thyroid (HPT) Axis in Frogs and Its Role in Frog Development and Reproduction. Crit. Rev. Toxicol. 2007, 37, 117–161. [Google Scholar] [CrossRef]
- Hansson, T.; Baršiene, J.; Tjärnlund, U.; Åkerman, G.; Linderoth, M.; Zebühr, Y.; Sternbeck, J.; Järnberg, U.; Balk, L. Cytological and Biochemical Biomarkers in Adult Female Perch (Perca Fluviatilis) in a Chronically Polluted Gradient in the Stockholm Recipient (Sweden). Mar. Pollut. Bull. 2014, 81, 27–40. [Google Scholar] [CrossRef]
- Orton, F.; Baynes, A.; Clare, F.; Duffus, A.L.J.; Larroze, S.; Scholze, M.; Garner, T.W.J. Body Size, Nuptial Pad Size and Hormone Levels: Potential Non-Destructive Biomarkers of Reproductive Health in Wild Toads (Bufo Bufo). Ecotoxicology 2014, 23, 1359–1365. [Google Scholar] [CrossRef]
- Nordberg, G.F. Biomarkers of Exposure, Effects and Susceptibility in Humans and Their Application in Studies of Interactions among Metals in China. Toxicol. Lett. 2010, 192, 45–49. [Google Scholar] [CrossRef]
- Ryan, P.B.; Burke, T.A.; Cohen Hubal, E.A.; Cura, J.J.; McKone, T.E. Using Biomarkers to Inform Cumulative Risk Assessment. Environ. Health Perspect. 2007, 115, 833–840. [Google Scholar] [CrossRef]
- Santos-Neto, E.B.; Azevedo-Silva, C.E.; Bisi, T.L.; Santos, J.; Meirelles, A.C.O.; Carvalho, V.L.; Azevedo, A.F.; Guimarães, J.E.; Lailson-Brito, J. Organochlorine Concentrations (PCBs, DDTs, HCHs, HCB and MIREX) in Delphinids Stranded at the Northeastern Brazil. Sci. Total Environ. 2014, 472, 194–203. [Google Scholar] [CrossRef]
- Mela, M.; Randi, M.A.F.; Ventura, D.F.; Carvalho, C.E.V.; Pelletier, E.; Oliveira Ribeiro, C.A. Effects of Dietary Methylmercury on Liver and Kidney Histology in the Neotropical Fish Hoplias Malabaricus. Ecotoxicol. Environ. Saf. 2007, 68, 426–435. [Google Scholar] [CrossRef]
- Basu, N.; Goodrich, J.M.; Head, J. Ecogenetics of Mercury: From Genetic Polymorphisms and Epigenetics to Risk Assessment and Decision-Making. Environ. Toxicol. Chem. 2014, 33, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Gutleb, A.C.; Schenck, C.; Staib, E. Giant Otter (Pteronura Brasiliensis) at Risk? Total Mercury and Methylmercury Levels in Fish and Otter Scats, Peru. Ambio 1997, 26, 511–514. [Google Scholar]
- Curi, N.H.D.A.; Brait, C.H.H.; Filho, N.R.A.; Talamoni, S.A. Heavy Metals in Hair of Wild Canids from the Brazilian Cerrado. Biol. Trace Elem. Res. 2012, 147, 97–102. [Google Scholar] [CrossRef]
- Dias Fonseca, F.R.; Malm, O.; Waldemarin, H.F. Mercury Levels in Tissues of Giant Otters (Pteronura Brasiliensis) from the Rio Negro, Pantanal, Brazil. Environ. Res. 2005, 98, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Josef, C.F.; Adriano, L.R.; De França, E.J.; Arantes de Carvalho, G.G.; Ferreira, J.R. Determination of Hg and Diet Identification in Otter (Lontra Longicaudis) Feces. Environ. Pollut. 2008, 152, 592–596. [Google Scholar] [CrossRef] [PubMed]
- May Junior, J.; Quigley, H.; Hoogenstein, R.; Tortato, F.; Devlin, A.; Carvahlo, R., Jr.; Morato, R.; Sartorello, L.; Rampin, L.; Haberfeld, M.; et al. Mercury Content in the Fur of Jaguars (Panthera Onca) from Two Areas under Different Levels of Gold Mining Impact in the Brazilian Pantanal. An. Acad. Bras. Cienc. 2017, 90, 1–11. [Google Scholar] [CrossRef]
- Zocche, J.J.; Dimer Leffa, D.; Paganini Damiani, A.; Carvalho, F.; Ávila Mendonça, R.; dos Santos, C.E.I.; Appel Boufleur, L.; Ferraz Dias, J.; de Andrade, V.M. Heavy Metals and DNA Damage in Blood Cells of Insectivore Bats in Coal Mining Areas of Catarinense Coal Basin, Brazil. Environ. Res. 2010, 110, 684–691. [Google Scholar] [CrossRef]
- Guerrero-Castilla, A.; Olivero-Verbel, J.; Marrugo-Negrete, J. Heavy Metals in Wild House Mice from Coal-Mining Areas of Colombia and Expression of Genes Related to Oxidative Stress, DNA Damage and Exposure to Metals. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 762, 24–29. [Google Scholar] [CrossRef]
- Racero-Casarrubia, J.A.; Marrugo-Negrete, J.L.; Pinedo-Hernández, J.J. Hallazgo de Mercurio En Piezas Dentales de Jaguares (Panthera Onca) Provenientes De La Zona Amortiguadora Del Parque Nacional Natural Paramillo, Cordoba, Colombia. Lat. Am. J. Conserv. 2012, 2, 87–92. [Google Scholar]
- Lajmanovich, R.; La Sierra, P.D.; Marino, F.; Peltzer, P.; Lenardón, A.; Lorenzatti, E. Determinación de Residuos de Organoclorados En Vertebrados Silvestres Del Litoral Fluvial de Argentina. Miscelánea 2005, 14, 389–398. [Google Scholar]
- Alava, J.J.; Ross, P.S.; Ikonomou, M.G.; Cruz, M.; Jimenez-Uzcátegui, G.; Dubetz, C.; Salazar, S.; Costa, D.P.; Villegas-Amtmann, S.; Howorth, P.; et al. DDT in Endangered Galapagos Sea Lions (Zalophus Wollebaeki). Mar. Pollut. Bull. 2011, 62, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Cortes, S.; Fortt, A. Mercury Content in Chilean Fish and Estimated Intake Levels. Food Addit. Contam. 2007, 24, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, E.; Espín, S.; Barbar, F.; Lambertucci, S.A.; Gómez-Ramírez, P.; García-Fernández, A. Contaminants in the Southern Tip of South America: Analysis of Organochlorine Compounds in Feathers of Avian Scavengers from Argentinean Patagonia. Ecotoxicol. Environ. Saf. 2015, 115, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Muto, E.; Soares, L.; Sarkis, J.; Hortellani, M.; Petti, M.; Corbisier, T. Biomagnification of Mercury through the Food Web of the Santos Continental Shelf, Subtropical Brazil. Mar. Ecol. Prog. Ser. 2014, 512, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Zapata, L.M.; Bock, B.C.; Palacio, J.A. Mercury Concentrations in Tissues of Colombian Slider Turtles, Trachemys Callirostris, from Northern Colombia. Bull. Environ. Contam. Toxicol. 2014, 92, 562–566. [Google Scholar] [CrossRef]
- Cledon, M.; Theobald, N.; Gerwinski, W.; Penchaszadeh, P.E. Imposex and Organotin Compounds in Marine Gastropods and Sediments from the Mar Del Plata Coast, Argentina. J. Mar. Biol. Assoc. 2006, 86, 751–755. [Google Scholar] [CrossRef]
- Goldberg, R.N.; Averbuj, A.; Cledón, M.; Luzzatto, D.; Sbarbati Nudelman, N. Search for Triorganotins along the Mar Del Plata (Argentina) Marine Coast: Finding of Tributyltin in Egg Capsules of a Snail Adelomelon Brasiliana (Lamarck, 1822) Population Showing Imposex Effects. Appl. Organomet. Chem. 2004, 18, 117–123. [Google Scholar] [CrossRef]
- Martínez, M.L.; Piol, M.N.; Sbarbati Nudelman, N.; Verrengia Guerrero, N.R. Tributyltin Bioaccumulation and Toxic Effects in Freshwater Gastropods Pomacea Canaliculata after a Chronic Exposure: Field and Laboratory Studies. Ecotoxicology 2017, 26, 691–701. [Google Scholar] [CrossRef]
- Commendatore, M.; Franco, M.; Gomes Costa, P.; Castro, I.; Fillman, G.; Bigatti, G.; Esteves, J.; Nievas, M. Butyltins, Polyaromatic Hydrocarbons, Organochlorine Pesticides, and Polychlorinated Biphenyls in Sediments and Bivalve Mollusks in a Mid-Latitude Environment From the Patagonian Coastal Zone. Environ. Toxicol. Chem. 2015, 34, 2750–2763. [Google Scholar] [CrossRef] [Green Version]
- Penchaszadeh, P.E.; Antelo, C.S.; Zabala, S.; Bigatti, G. Reproduction and Imposex in the Edible Snail Adelomelon Ancilla from Northern Patagonia, Argentina. Mar. Biol. 2009, 156, 1929–1939. [Google Scholar] [CrossRef]
- Penchaszadeh, P.E.; Averbuj, A.; Cledón, M. Imposex in Gastropods from Argentina (South -Western Atlantic). Mar. Pollut. Bull. 2001, 42, 790–791. [Google Scholar] [CrossRef] [PubMed]
- Bigatti, G.; Primost, M.A.; Cledón, M.; Averbuj, A.; Theobald, N.; Gerwinski, W.; Arntz, W.; Morriconi, E.; Penchaszadeh, P.E. Biomonitoring of TBT Contamination and Imposex Incidence along 4700 Km of Argentinean Shoreline (SW Atlantic: From 38S to 54S). Mar. Pollut. Bull. 2009, 58, 695–701. [Google Scholar] [CrossRef]
- Primost, M.A.; Bigatti, G.; Márquez, F. Shell Shape as Indicator of Pollution in Marine Gastropods Affected by Imposex. Mar. Freshw. Res. 2016, 67, 1948–1954. [Google Scholar] [CrossRef]
- Márquez, F.; Primost, M.A.; Bigatti, G. Shell Shape as a Biomarker of Marine Pollution Historic Increase. Mar. Pollut. Bull. 2017, 114, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Cid, F.D.; Antón, R.I.; Caviedes-Vidal, E. Organochlorine Pesticide Contamination in Three Bird Species of the Embalse La Florida Water Reservoir in the Semiarid Midwest of Argentina. Sci. Total Environ. 2007, 385, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Jofré, M.B.; Antón, R.I.; Caviedes-Vidal, E. Organochlorine Contamination in Anuran Amphibians of an Artificial Lake in the Semiarid Midwest of Argentina. Arch. Environ. Contam. Toxicol. 2008, 55, 471–480. [Google Scholar] [CrossRef]
- Rautenberg, G.E.; Amé, M.V.; Monferrán, M.V.; Bonansea, R.I.; Hued, A.C. A Multi-Level Approach Using Gambusia Affinis as a Bioindicator of Environmental Pollution in the Middle-Lower Basin of Suquía River. Ecol. Indic. 2015, 48, 706–720. [Google Scholar] [CrossRef]
- Miranda, A.L.; Roche, H.; Randi, M.A.F.; Menezes, M.L.; Oliveira Ribeiro, C.A. Bioaccumulation of Chlorinated Pesticides and PCBs in the Tropical Freshwater Fish Hoplias Malabaricus: Histopathological, Physiological, and Immunological Findings. Environ. Int. 2008, 34, 939–949. [Google Scholar] [CrossRef]
- Castro, I.B.; Fillmann, G. High Tributyltin and Imposex Levels in the Commercial Muricid Thais Chocolata from Two Peruvian Harbor Areas. Environ. Toxicol. Chem. 2012, 31, 955–960. [Google Scholar] [CrossRef]
- Dorneles, P.R.; Lailson-Brito, J.; Fernandez, M.A.S.; Vidal, L.G.; Barbosa, L.A.; Azevedo, A.F.; Fragoso, A.B.L.; Torres, J.P.M.; Malm, O. Evaluation of Cetacean Exposure to Organotin Compounds in Brazilian Waters through Hepatic Total Tin Concentrations. Environ. Pollut. 2008, 156, 1268–1276. [Google Scholar] [CrossRef]
- Dos Santos, D.M.; Turra, A.; de Marchi, M.R.R.; Montone, R.C. Distribution of Butyltin Compounds in Brazil’s Southern and Southeastern Estuarine Ecosystems: Assessment of Spatial Scale and Compartments. Environ. Sci. Pollut. Res. 2016, 23, 16152–16163. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, D.M.; Santos, G.S.; Cestari, M.M.; de Oliveira Ribeiro, C.A.; de Assis, H.C.S.; Yamamoto, F.; Guiloski, I.C.; de Marchi, M.R.R.; Montone, R.C. Bioaccumulation of Butyltins and Liver Damage in the Demersal Fish Cathorops Spixii (Siluriformes, Ariidae). Environ. Sci. Pollut. Res. 2014, 21, 3166–3174. [Google Scholar] [CrossRef] [PubMed]
- Pletsch, A.L.; Beretta, M.; Tavares, T.M. Spatial Distribution of Organic Tin Compounds in Coastal Sediment and Phallusia Nigra of the Todos Os Santos Bay and Northern Coast of Bahia-Brazil. Quim. Nova 2010, 33, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Rossato, M.; Castro, I.B.; Paganini, C.L.; Colares, E.P.; Fillmann, G.; Pinho, G.L.L. Sex Steroid Imbalances in the Muricid Stramonita Haemastoma from TBT Contaminated Sites. Environ. Sci. Pollut. Res. 2016, 23, 7861–7868. [Google Scholar] [CrossRef]
- Sant’Anna, B.S.; Santos, D.M.; Marchi, M.R.R.; Zara, F.J.; Turra, A. Surface-Sediment and Hermit-Crab Contamination by Butyltins in Southeastern Atlantic Estuaries after Ban of TBT-Based Antifouling Paints. Environ. Sci. Pollut. Res. 2014, 21, 6516–6524. [Google Scholar] [CrossRef]
- Maciel, D.; Castro, Í.; de Souza, J.; Yogui, G.; Fillmann, G.; Zanardi-Lamardo, E.; Zanardi-lamardo, E. Assessment of Organotins and Imposex in Two Estuaries of the Northeastern Brazilian Coast. Mar. Pollut. Bull. 2018, 126, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Prado, P.S.; Souza, C.C.; Bazzoli, N.; Rizzo, E. Reproductive Disruption in Lambari Astyanax Fasciatus from a Southeastern Brazilian Reservoir. Ecotoxicol. Environ. Saf. 2011, 74, 1879–1887. [Google Scholar] [CrossRef]
- Paschoalini, A.L.; Savassi, L.A.; Arantes, F.P.; Rizzo, E.; Bazzoli, N. Heavy Metals Accumulation and Endocrine Disruption in Prochilodus Argenteus from a Polluted Neotropical River. Ecotoxicol. Environ. Saf. 2019, 169, 539–550. [Google Scholar] [CrossRef]
- Tolussi, C.E.; Olio, A.D.; Kumar, A.; Ribeiro, C.S.; Lo, F.L.; Bain, P.A.; De Souza, G.B.; Da, R.; Honji, R.M.; Moreira, R.G. Environmental Pollution Affects Molecular and Biochemical Responses during Gonadal Maturation of Astyanax Fasciatus (Teleostei: Characiformes: Characidae). Ecotoxicol. Environ. Saf. 2018, 147, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Prado, P.S.; Pinheiro, A.P.B.; Bazzoli, N.; Rizzo, E. Reproductive Biomarkers Responses Induced by Xenoestrogens in the Characid Fish Astyanax Fasciatus Inhabiting a South American Reservoir: An Integrated Field and Laboratory Approach. Environ. Res. 2014, 131, 165–173. [Google Scholar] [CrossRef]
- Moresco, R.M.; Margarido, V.P.; de Oliveira, C. A Persistent Organic Pollutant Related with Unusual High Frequency of Hermaphroditism in the Neotropical Anuran Physalaemus Cuvieri Fitzinger, 1826. Environ. Res. 2014, 132, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.F.; Damasceno, É.P.; Jimenez, P.C.; Araújo, P.F.R.; Bezerra, M.F.; de Morais, P.C.V.; Cavalcante, R.M.; Loureiro, S.; Lotufo, L.V.C. Endocrine Disruption in Sphoeroides Testudineus Tissues and Sediments Highlights Contamination in a Northeastern Brazilian Estuary. Environ. Monit. Assess. 2016, 188, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.A.; Moreira, D.P.; Melo, R.M.C.; Vieira, A.B.C.; Prado, P.S.; da Silva, M.A.N.; Bazzoli, N.; Rizzo, E. Reproductive Effects of Oestrogenic Endocrine Disrupting Chemicals in Astyanax Rivularis Inhabiting Headwaters of the Velhas River, Brazil. Sci. Total Environ. 2017, 592, 693–703. [Google Scholar] [CrossRef]
- Weber, A.A.; Moreira, D.P.; Magno, R.; Melo, C.; Ribeiro, Y.M.; Bazzoli, N.; Rizzo, E. Environmental Exposure to Oestrogenic Endocrine Disruptors Mixtures Reflecting on Gonadal Sex Steroids and Gametogenesis of the Neotropical Fish Astyanax Rivularis. Gen. Comp. Endocrinol. 2019, 279, 99–108. [Google Scholar] [CrossRef]
- Yamamoto, F.Y.; Garcia, J.R.E.; Kupsco, A.; Oliveira Ribeiro, C.A. Vitellogenin Levels and Others Biomarkers Show Evidences of Endocrine Disruption in Fish Species from Iguaçu River-Southern Brazil. Chemosphere 2017, 186, 88–99. [Google Scholar] [CrossRef]
- Simões, T.; Silva, A.; Santos, A.; Chagas, C. Heavy Metals in Blood and in Nests Affect Reproduction Parameters in Eretmochelys Imbricata, Linnaeus, 1766 (Testudines: Cryptodira). Ecotoxicol. Environ. Contam. 2019, 14, 65–72. [Google Scholar] [CrossRef]
- Deliberalli, W.; Cansian, R.L.; Pereira, A.A.M.; Loureiro, R.C.; Hepp, L.U.; Restello, R.M. The Effects of Heavy Metals on the Incidence of Morphological Deformities in Chironomidae (Diptera). Zoologia 2018, 35, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Otegui, M.B.P.; Zamprogno, G.C.; França, M.A.; Daros, B.N.; Albino, J.; Costa, M.B. Imposex Response in Shell Sizes of Intertidal Snails in Multiple Environments. J. Sea Res. 2019, 147, 10–18. [Google Scholar] [CrossRef]
- Gooding, M.; Gallardo, C.; Leblanc, G. Imposex in Three Marine Gastropod Species in Chile and Potential Impact on Muriciculture. Mar. Pollut. Bull. 1999, 38, 1227–1231. [Google Scholar] [CrossRef]
- Huaquín, L.G.; Osorio, C.; Verdugo, R.; Collado, G. Morphological Changes in the Reproductive System of Females Acanthina Monodon (Pallas, 1774) (Gastropoda: Muricidae) Asffected by Imposex from the Coast of Central Chile. Invertebr. Reprod. Dev. 2004, 46, 111–117. [Google Scholar] [CrossRef]
- Mattos, Y.; Romero, M.S. Imposex in Thaisella Chocolata (Duclos, 1832) (Gastropoda: Muricidae) Caldera Bay, Chile. Lat. Am. J. Aquat. Res. 2016, 44, 825–834. [Google Scholar] [CrossRef]
- Batista, R.M.; Castro, I.B.; Fillmann, G. Imposex and Butyltin Contamination Still Evident in Chile after TBT Global Ban. Sci. Total Environ. 2016, 566–567, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Mattos, Y.; Stotz, W.B.; Romero, M.S.; Bravo, M.; Fillmann, G.; Castro, Í.B. Butyltin Contamination in Northern Chilean Coast: Is There a Potential Risk for Consumers? Sci. Total Environ. 2017, 595, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Chiang, G.; McMaster, M.E.; Urrutia, R.; Saavedra, M.F.; Gavilán, J.F.; Tucca, F.; Barra, R.; Munkittrick, K.R. Health Status of Native Fish (Percilia Gillissi and Trichomycterus Areolatus) Downstream of the Discharge of Effluent from a Tertiary-Treated Elemental Chlorine-Free Pulp Mill in Chile. Environ. Toxicol. Chem. 2011, 30, 1793–1809. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, M.O.; Puchi, M.; Bustos, P.; Romo, X.; Morín, V. Vitellogenin Induction and Reproductive Status in Wild Chilean Flounder Paralichthys Adspersus (Steindachner, 1867) as Biomarkers of Endocrine Disruption along the Marine Coast of the South Pacific. Arch. Environ. Contam. Toxicol. 2012, 62, 314–322. [Google Scholar] [CrossRef]
- Leonardi, M.; Tarifeño, E.; Vera, J. Diseases of the Chilean Flounder, Paralichthys Adspersus (Steindachner, 1867), as a Biomarker of Marine Coastal Pollution near the Itata River (Chile): Part II. Histopathological Lesions. Arch. Environ. Contam. Toxicol. 2009, 56, 546–556. [Google Scholar] [CrossRef]
- Orrego, R.; Hewitt, L.M.; Mcmaster, M.; Chiang, G.; Quiroz, M.; Munkittrick, K.; Gavilán, J.F.; Barra, R. Safety Assessing Wild Fish Exposure to Ligands for Sex Steroid Receptors from Pulp and Paper Mill Effluents in the Biobio River Basin, Central Chile. Ecotoxicol. Environ. Saf. 2019, 171, 256–263. [Google Scholar] [CrossRef]
- Rojas-Hucks, S.; Gutleb, A.C.; González, C.M.; Contal, S.; Mehennaoui, K.; Jacobs, A.; Witters, H.E.; Pulgar, J. Xenopus Laevis as a Bioindicator of Endocrine Disruptors in the Region of Central Chile. Arch. Environ. Contam. Toxicol. 2019, 77, 390–408. [Google Scholar] [CrossRef]
- Olivero-Verbel, J.; Tao, L.; Johnson-Restrepo, B.; Guette-Fernandez, J.; Baldiris-Avila, R.; O´byrne-Hoyos, I.; Kannan, K. Perfluorooctanesulfonate and Related Fluorochemicals in Biological Samples from the North Coast of Colombia. Environ. Pollut. 2006, 142, 367–372. [Google Scholar] [CrossRef]
- Monsalve, A.S.; Criollo, S.M.D.; Uribe, M.E.V.; Mantilla, J.F.G.; Forero, A.R. Exposure to Pesticides in Residents or the Banks of the Río Bogotá (Suesca) and the Capitán Fish. Rev. Ciencias Salud 2012, 10, 29–41. [Google Scholar]
- Sierra-Marquez, L.; Sierra-Marquez, J.; De La Rosa, J.; Olivero-Verbel, J. Imposex in Stramonita Haemastoma from Coastal Sites of Cartagena, Colombia. Braz. J. Biol. 2017, 78, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivero-Verbel, J.; Agudelo-Frias, D.; Caballero-Gallardo, K. Morphometric Parameters and Total Mercury in Eggs of Snowy Egret (Egretta Thula) from Cartagena Bay and Totumo Marsh, North of Colombia. Mar. Pollut. Bull. 2013, 69, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Rubí, J.; Luna-Acosta, A.; Ortiz-Zarragoitia, M.; Zaldibar, B.; Izagirre, U.; Ahrens, M.J. Assessment of Ecosystem Health Disturbance in Mangrove-Lined Caribbean Coastal Systems Using the Oyster Crassostrea Rhizophorae as Sentinel Species. Sci. Total Environ. 2018, 618, 718–735. [Google Scholar] [CrossRef]
- Sierra-Marquez, L.; Espinosa-Araujo, J.; Atencio-Garcia, V.; Olivero-Verbel, J. Effects of Cadmium Exposure on Sperm and Larvae of the Neotropical Fish Prochilodus Magdalenae. Comp. Biochem. Physiol. Part C 2019, 225, 108577. [Google Scholar] [CrossRef] [PubMed]
- Moncaleano-Niño, A.M.; Barrrios-Latorre, S.; Poloche-Hernández, J.F.; Becquet, V.; Huet, V.; Villamil, L.; Thomas-Guyon, H.; Ahrens, M.J.; Luna-Acosta, A. Alterations of Tissue Metallothionein and Vitellogenin Concentrations in Tropical Cup Oysters (Saccostrea Sp.) Following Short-Term (96 h) Exposure to Cadmium. Aquat. Toxicol. 2017, 185, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Montaño-Campaz, M.; Gomes-Dias, L.; Toro Restrepo, B.; García-Merchán, V. Incidence of Deformities and Variation in Shape of Mentum and Wing of Chironomus Columbiensis (Diptera, Chironomidae) as Tools to Assess Aquatic Contamination. PLoS ONE 2019, 14, e0210348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdaneta, V.; Camafeita, E.; Poleo, G.; Guerrero, H.; Bernal, C.; Galindo-Castro, I.; Diez, N.; Iii, C. Proteomic Characterization of Vitellogenins from Three Species of South American Fresh Water Fish. Lat. Am. J. Aquat. Res. 2018, 46, 187–196. [Google Scholar] [CrossRef]
- Paz-Villarraga, C.A.; Castro, I.B.; Miloslavich, P.; Fillmann, G. Venezuelan Caribbean Sea under the Threat of TBT. Chemosphere 2015, 119, 704–710. [Google Scholar] [CrossRef]
- Peralta, A.C.; Miloslavich, P.; Bigatti, G. Imposex En Voluta Musica (Caenogastropoda: Volutidae) En El Noreste de La Península de Araya, Venezuela. Rev. Biol. Trop. 2014, 62, 523–532. [Google Scholar] [CrossRef]
- Miloslavich, P.; Penchaszadeh, P.E.; Bigatti, G. Imposex En Gastrópodos de Venezuela. Ciencias Mar. 2007, 33, 319–324. [Google Scholar] [CrossRef]
- Castro, Í.B.; Arroyo, M.F.; Costa, P.G.; Fillmann, G. Butyltin Compounds and Imposex Levels in Ecuador. Arch. Environ. Contam. Toxicol. 2012, 62, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Chumbimune-Ilizarbe, L.M.; Ponce-Mora, Z.J. Monitoring of Pollution Tributyltin (TBT) in Port of Paracas Lima (Peru). Científica 2015, 12, 222–230. [Google Scholar]
- Guabloche, A.; Alvarez, J.; Rivas, R.; Hurtado, S.; Pradel, R.; Iannacone, J. Imposex in the Marine Snail Xanthochorus Buxea (Broderip, 1833) (Muricidae) From the South American Pacific. Biology 2013, 11, 237–249. [Google Scholar]
- Rodríguez Grimón, R.O.; Arroyo Osorio, M.F.; de Freitas, D.M.; Castro, Í.B. Tributyltin Impacts in Galapagos Islands and Ecuadorian Shore: Marine Protected Areas under Threat. Mar. Policy 2016, 69, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Enrique, S.; Guabloche, A.; Tuesta, E.; Iannacone, J. Imposex Responses in Thaisella Chocolata and Xanthochorus Buxeus from Callao Harbor, Peru. Reg. Stud. Mar. Sci. 2019, 26, 100510. [Google Scholar] [CrossRef]
- Carnikian, A.; Miguez, D.; Vizziano-Cantonnet, D. Histomorphological Evaluation of Pimephales Promelas Male Gonads after Exposure to Pulp Mill and Domestic Discharges into the Uruguay River (Fray Bentos-Uruguay). Indian J. Sci. Technol 2011, 4, 268–269. [Google Scholar]
- Rivas-Rivera, N.; Eguren, G.; Carrasco-Letelier, L.; Munkittrick, K.R. Screening of Endocrine Disruption Activity in Sediments from the Uruguay River. Ecotoxicology 2014, 23, 1137–1142. [Google Scholar] [CrossRef]
- Vidal, N.; Loureiro, M.; Hued, A.C.; Eguren, G.; de Mello, F.T. Female Masculinization and Reproductive Success in Cnesterodon Decemmaculatus (Jenyns, 1842) (Cyprinodontiforme: Poeciliidae) under Anthropogenic Impact. Ecotoxicology 2018, 27, 1331–1340. [Google Scholar] [CrossRef]
- Arbeláez-Cortés, E. Knowledge of Colombian Biodiversity: Published and Indexed. Biodivers. Conserv. 2013, 22, 2875–2906. [Google Scholar] [CrossRef]
- Armenteras, D.; Gast, F.; Villareal, H. Andean Forest Fragmentation and the Representativeness of Protected Natural Areas in the Eastern Andes, Colombia. Biol. Conserv. 2003, 113, 245–256. [Google Scholar] [CrossRef]
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined Use of Systematic Conservation Planning, Species Distribution Modelling, and Connectivity Analysis Reveals Severe Conservation Gaps in a Megadiverse Country (Peru). PLoS ONE 2014, 9, e114367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrea, C.; Warnars, L. Ecuador’s Yasuni-ITT Initiative: Avoiding Emissions by Keeping Petroleum Underground. Energy Sustain. Dev. 2009, 13, 219–223. [Google Scholar] [CrossRef]
- Lewinsohn, T.M.; Prado, P. How Many Species Are There in Brazil? Conserv. Biol. 2005, 19, 619–624. [Google Scholar] [CrossRef]
- Mittermeier, R.; da Fonseca, G.; Rylands, A.; Brandon, K. A Brief History of Biodiversity Conservation in Brazil. Conserv. Biol. 2005, 19, 601–607. [Google Scholar] [CrossRef]
- Cordy, P.; Veiga, M.M.; Salih, I.; Al-Saadi, S.; Console, S.; Garcia, O.; Mesa, L.A.; Velásquez-López, P.C.; Roeser, M. Mercury Contamination from Artisanal Gold Mining in Antioquia, Colombia: The World’s Highest per Capita Mercury Pollution. Sci. Total Environ. 2011, 410–411, 154–160. [Google Scholar] [CrossRef]
- Fernández, F. The Greatest Impediment to the Study of Biodiversity in Colombia. Caldasia 2011, 33, 1–5. [Google Scholar]
- Santos, B.S.; Silva, L.C.N.; Silva, T.D.; Rodrigues, J.F.S. Application of Omics Technologies for Evaluation of Antibacterial Mechanisms of Action of Plant-Derived Products. Front. Microbiol. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Beldomenico, P.M.; Rey, F.; Prado, W.S.; Villarreal, J.C.; Muñoz-de-Toro, M.; Luque, E.H. In Ovum Exposure to Pesticides Increases the Egg Weight Loss and Decreases Hatchlings Weight of Caiman Latirostris (Crocodylia: Alligatoridae). Ecotoxicol. Environ. Saf. 2007, 68, 246–251. [Google Scholar] [CrossRef]
- Durando, M.; Cocito, L.; Rodríguez, H.A.; Varayoud, J.; Ramos, J.G.; Luque, E.H.; Muñoz-de-Toro, M. Neonatal Expression of Amh, Sox9 and Sf-1 MRNA in Caiman Latirostris and Effects of in Ovo Exposure to Endocrine Disrupting Chemicals. Gen. Comp. Endocrinol. 2013, 191, 31–38. [Google Scholar] [CrossRef]
- Rey, F.; González, M.; Zayas, M.A.; Stoker, C.; Durando, M.; Luque, E.H.; Muñoz-de-Toro, M. Prenatal Exposure to Pesticides Disrupts Testicular Histoarchitecture and Alters Testosterone Levels in Male Caiman Latirostris. Gen. Comp. Endocrinol. 2009, 162, 286–292. [Google Scholar] [CrossRef]
- Stoker, C.; Beldoménico, P.M.; Bosquiazzo, V.L.; Zayas, M.A.; Rey, F.; Rodríguez, H.; Muñoz-de-Toro, M.; Luque, E.H. Developmental Exposure to Endocrine Disruptor Chemicals Alters Follicular Dynamics and Steroid Levels in Caiman Latirostris. Gen. Comp. Endocrinol. 2008, 156, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Stoker, C.; Rey, F.; Rodriguez, H.; Ramos, J.G.; Sirosky, P.; Larriera, A.; Luque, E.H.; Muñoz-De-Toro, M. Sex Reversal Effects on Caiman Latirostris Exposed to Environmentally Relevant Doses of the Xenoestrogen Bisphenol A. Gen. Comp. Endocrinol. 2003, 133, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Stoker, C.; Repetti, M.R.; García, S.R.; Zayas, M.A.; Galoppo, G.H.; Beldoménico, H.R.; Luque, E.H.; Muñoz-de-Toro, M. Organochlorine Compound Residues in the Eggs of Broad-Snouted Caimans (Caiman Latirostris) and Correlation with Measures of Reproductive Performance. Chemosphere 2011, 84, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Stoker, C.; Zayas, M.A.; Ferreira, M.A.; Durando, M.; Galoppo, G.H.; Rodríguez, H.A.; Repetti, M.R.; Beldoménico, H.R.; Caldini, E.G.; Luque, E.H.; et al. The Eggshell Features and Clutch Viability of the Broad-Snouted Caiman (Caiman Latirostris) Are Associated with the Egg Burden of Organochlorine Compounds. Ecotoxicol. Environ. Saf. 2013, 98, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Durando, M.; Canesini, G.; Cocito, L.L.; Galoppo, G.H.; Zayas, M.A.; Luque, E.H.; Muñoz-de-Toro, M. Histomorphological Changes in Testes of Broad-Snouted Caimans (Caiman Latirostris) Associated with in Ovo Exposure to Endocrine-Disrupting Chemicals. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2016, 325, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Da Cuña, R.H.; Pandolfi, M.; Genovese, G.; Piazza, Y.; Ansaldo, M.; Lo Nostro, F.L. Endocrine Disruptive Potential of Endosulfan on the Reproductive Axis of Cichlasoma Dimerus (Perciformes, Cichlidae). Aquat. Toxicol. 2013, 126, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Genovese, G.; Da Cuña, R.; Towle, D.W.; Maggese, M.C.; Lo Nostro, F. Early Expression of Zona Pellucida Proteins under Octylphenol Exposure in Cichlasoma Dimerus (Perciformes, Cichlidae). Aquat. Toxicol. 2011, 101, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Rey Vázquez, G.; Meijide, F.J.; Da Cuña, R.H.; Lo Nostro, F.L.; Piazza, Y.G.; Babay, P.A.; Trudeau, V.L.; Maggese, M.C.; Guerrero, G.A. Exposure to Waterborne 4-Tert-Octylphenol Induces Vitellogenin Synthesis and Disrupts Testis Morphology in the South American Freshwater Fish Cichlasoma Dimerus (Teleostei, Perciformes). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 150, 298–306. [Google Scholar] [CrossRef]
- Roggio, M.A.; Hued, A.C.; Roggio, M.A.; Hued, A.C. Effects of 4 N-Nonylphenol Exposure on the Reproductive Behavior and Testis Histology of Jenynsia Multidentata (Anablepidae: Cyprinodontiformes). Nat. Neotrop. 2012, 1–2, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Roggio, M.A.; Guyón, N.F.; Hued, A.C.; Amé, M.V.; Valdés, M.E.; Giojalas, L.C.; Wunderlin, D.A.; Bistoni, M.A. Effects of the Synthetic Estrogen 17α-Ethinylestradiol on Aromatase Expression, Reproductive Behavior and Sperm Quality in the Fish Jenynsia Multidentata. Bull. Environ. Contam. Toxicol. 2014, 92, 579–584. [Google Scholar] [CrossRef]
- Guyón, N.F.; Roggio, M.A.; Amé, M.V.; Hued, A.C.; Valdés, M.E.; Giojalas, L.C.; Wunderlin, D.A.; Bistoni, M.A. Impairments in Aromatase Expression, Reproductive Behavior, and Sperm Quality of Male Fish Exposed to 17β-Estradiol. Environ. Toxicol. Chem. 2012, 31, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.R.; Fernandino, J.I.; Carriquiriborde, P.; Somoza, G.M. Feminization and Altered Gonadal Gene Expression Profile by Ethinylestradiol Exposure to Pejerrey, Odontesthes Bonariensis, a South American Teleost Fish. Environ. Toxicol. Chem. 2012, 31, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Meijide, F.J.; Rey Vázquez, G.; Piazza, Y.G.; Babay, P.A.; Itria, R.F.; Lo Nostro, F.L. Effects of Waterborne Exposure to 17β-Estradiol and 4-Tert-Octylphenol on Early Life Stages of the South American Cichlid Fish Cichlasoma Dimerus. Ecotoxicol. Environ. Saf. 2016, 124, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Rey Vázquez, G.; Meijide, F.J.; Lo Nostro, F.L. Recovery of the Reproductive Capability Following Exposure to 4-Tert-Octylphenol in the Neotropical Cichlid Fish Cichlasoma Dimerus. Bull. Environ. Contam. Toxicol. 2016, 96, 585–590. [Google Scholar] [CrossRef]
- Da Cuña, R.H.; Rey Vázquez, G.; Dorelle, L.; Rodríguez, E.M.; Guimarães Moreira, R.; Lo Nostro, F.L. Mechanism of Action of Endosulfan as Disruptor of Gonadal Steroidogenesis in the Cichlid Fish Cichlasoma Dimerus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 187, 74–80. [Google Scholar] [CrossRef]
- Gárriz, Á.; Pamela, S.; Carriquiriborde, P.; Miranda, L.A. Effects of Heavy Metals Identified in Chascomús Shallow Lake on the Endocrine-Reproductive Axis of Pejerrey Fish (Odontesthes Bonariensis). Gen. Comp. Endocrinol. 2018, 273, 1–10. [Google Scholar] [CrossRef]
- Negro, C.L. Histopathological Effects of Endosulfan to Hepatopancreas, Gills and Ovary of the Freshwater Crab Zilchiopsis Collastinensis (Decapoda: Trichodactylidae). Ecotoxicol. Environ. Saf. 2015, 113, 87–94. [Google Scholar] [CrossRef]
- Mac Loughlin, C.; Canosa, I.S.; Silveyra, G.R.; López Greco, L.S.; Rodríguez, E.M. Effects of Atrazine on Growth and Sex Differentiation, in Juveniles of the Freshwater Crayfish Cherax Quadricarinatus. Ecotoxicol. Environ. Saf. 2016, 131, 96–103. [Google Scholar] [CrossRef]
- Silveyra, G.R.; Canosa, I.S.; Rodríguez, E.M.; Medesani, D.A. Effects of Atrazine on Ovarian Growth, in the Estuarine Crab Neohelice Granulata. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 192, 1–6. [Google Scholar] [CrossRef]
- Berasategui, A.A.; Biancalana, F.; Fricke, A.; Fernandez -Severini, M.D.; Uibrig, R.; Dutto, M.S.; Marcovecchio, J.; Calliari, D.; Hoffmeyer, M.S. The Impact of Sewage Effluents on the Fecundity and Survival of Eurytemora Americana in a Eutrophic Estuary of Argentina. Estuar. Coast. Shelf Sci. 2017, 211, 1–9. [Google Scholar] [CrossRef]
- Medina, M.F.; Cosci, A.; Cisint, S.; Crespo, C.A.; Ramos, I.; Iruzubieta Villagra, A.L.; Fernández, S.N. Histopathological and Biological Studies of the Effect of Cadmium on Rhinella Arenarum Gonads. Tissue Cell 2012, 44, 418–426. [Google Scholar] [CrossRef]
- Svartz, G.; Meijide, F.; Pérez Coll, C. Effects of a Fungicide Formulation on Embryo-Larval Development, Metamorphosis, and Gonadogenesis of the South American Toad Rhinella Arenarum. Environ. Toxicol. Pharmacol. 2016, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bach, N.C.; Natale, G.S.; Somoza, G.M.; Ronco, A.E. Effect on the Growth and Development and Induction of Abnormalities by a Glyphosate Commercial Formulation and Its Active Ingredient during Two Developmental Stages of the South-American Creole Frog, Leptodactylus Latrans. Environ. Sci. Pollut. Res. 2016, 23, 23959–23971. [Google Scholar] [CrossRef] [PubMed]
- Seabra Pereira, C.D.; Abessa, D.M.S.; Choueri, R.B.; Almagro-Pastor, V.; Cesar, A.; Maranho, L.A.; Martín-Díaz, M.L.; Torres, R.J.; Gusso-Choueri, P.K.; Almeida, J.E.; et al. Ecological Relevance of Sentinels’ Biomarker Responses: A Multi-Level Approach. Mar. Environ. Res. 2014, 96, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Vergílio, C.S.; Moreira, R.V.; Carvalho, C.E.V.; Melo, E.J.T. Effects of in Vitro Exposure to Mercury on Male Gonads and Sperm Structure of the Tropical Fish Tuvira Gymnotus Carapo (L.). J. Fish Dis. 2014, 37, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Boscolo Pereira, T.S.; Pereira Boscolo, C.N.; Felício, A.A.; Batlouni, S.R.; Schlenk, D.; Alves de Almeida, E. Estrogenic Activities of Diuron Metabolites in Female Nile Tilapia (Oreochromis Niloticus). Chemosphere 2016, 146, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcon, L.; Thomé, R.G.; Mounteer, A.H.; Bazzoli, N.; Rizzo, E.; dos Anjos Benjamin, L. Immunohistochemical, Morphological and Histometrical Analyses of Follicular Development in Astyanax Bimaculatus (Teleostei: Characidae) Exposed to an Organochlorine Insecticide. Ecotoxicol. Environ. Saf. 2017, 143, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Guiloski, I.C.; Ribas, J.L.C.; Piancini, L.D.S.; Dagostim, A.C.; Cirio, S.M.; Fávaro, L.F.; Boschen, S.L.; Cestari, M.M.; da Cunha, C.; Silva de Assis, H.C. Paracetamol Causes Endocrine Disruption and Hepatotoxicity in Male Fish Rhamdia Quelen after Subchronic Exposure. Environ. Toxicol. Pharmacol. 2017, 53, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Zebral, Y.D.; Costa, P.G.; de Castro Knopp, B.; Lansini, L.R.; Zafalon-Silva, B.; Bianchini, A.; Robaldo, R.B. Effects of a Glyphosate-Based Herbicide in Pejerrey Odontesthes Humensis Embryonic Development. Chemosphere 2017, 185, 860–867. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, I.; Ramon, J.; Garcia, E.; Castro, D.; Neto, A.C.; Gusso-choueri, P.K.; Brasil, R.; Borges, S.; Araujo, L.; Alberto, C.; et al. Embryo Toxicity Assay in the Fish Species Rhamdia Quelen (Teleostei, Heptaridae) to Assess Water Quality in the Upper Iguaçu Basin (Parana, Brazil). Chemosphere 2018, 208, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Brinati, A.; Oliveira, J.M.; Oliveira, V.S.; Barros, M.S.; Carvalho, B.M.; Oliveira, L.S.; Queiroz, M.E.L.; Matta, S.L.P.; Freitas, M.B. Low, Chronic Exposure to Endosulfan Induces Bioaccumulation and Decreased Carcass Total Fatty Acids in Neotropical Fruit Bats. Bull. Environ. Contam. Toxicol. 2016, 97, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, L.; Leonardi, M.; Morin, V. Induction of Vitellogenin-like Lipoproteins in the Mussel Aulacomya Ater under Exposure to 17 β -Estradiol. Rev. Biol. Mar. Oceanogr. 2012, 47, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Chiang, G.; Barra, R.; Díaz-Jaramillo, M.; Rivas, M.; Bahamonde, P.; Munkittrick, K.R. Estrogenicity and Intersex in Juvenile Rainbow Trout (Oncorhynchus Mykiss) Exposed to Pine/Eucalyptus Pulp and Paper Production Effluent in Chile. Aquat. Toxicol. 2015, 164, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Orrego, R.; Moraga-Cid, G.; González, M.; Barra, R.; Valenzuela, A.; Burgos, A.; Gavilán, J.F. Reproductive, Physiological, and Biochemical Responses in Juvenile Female Rainbow Trout (Oncorhynchus Mykiss) Exposed to Sediment from Pulp and Paper Mill Industrial Discharge Areas. Environ. Toxicol. Chem. 2005, 24, 1935–1943. [Google Scholar] [CrossRef] [Green Version]
- Orrego, R.; Burgos, A.; Moraga-Cid, G.; Inzunza, B.; Gonzalez, M.; Valenzuela, A.; Barra, R.; Gavilán, J.F. Effects of Pulp and Paper Mill Discharges on Caged Rainbow Trout (Oncorhynchus Mykiss): Biomarker Responses along a Pollution Gradient in the Biobio River, Chile. Environ. Toxicol. Chem. 2006, 25, 2280–2287. [Google Scholar] [CrossRef]
- Orrego, R.; Guchardi, J.; Hernandez, V.; Krause, R.; Roti, L.; Armour, J.; Ganeshakumar, M.; Holdway, D. Pulp and Paper Mill Effluent Treatments Have Differential Endocrine-Disrupting Effects on Rainbow Trout. Environ. Toxicol. Chem. 2009, 28, 181–188. [Google Scholar] [CrossRef]
- Bahamonde, P.; Berrocal, C.; Barra, R.; Mcmaster, M.E.; Munkittrick, K.R.; Chiang, G. Mucus Phosphoproteins as an Indirect Measure of Endocrine Disruption in Native Small-Bodied Freshwater Fish, Exposed to Wastewater Treatment Plant and Pulp and Paper Mill Effluents. Gayana 2019, 83, 10–20. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | M N(%) | POPs N(%) | POPs/M N(%) | POPs/Other N(%) | Others N(%) | Total N |
|---|---|---|---|---|---|---|
| Argentina | 58 (66) | 22 (25) | 1 (1.1) | 1 (1.1) | 6 (6.8) | 88 |
| Bolivia | 7 (100) | 0 | 0 | 0 | 0 | 7 |
| Brazil | 247 (76) | 59 (18.2) | 7 (2.2) | 2 (0.6) | 10 (3) | 325 |
| Chile | 34 (47.2) | 33 (45.8) | 3 (4.2) | 0 | 2 (2.8) | 72 |
| Colombia | 36 (76.6) | 7 (14.9) | 2 (4.3) | 1 (2.1) | 1 (2.1) | 47 |
| Ecuador | 4 (66.7) | 2 (33.3) | 0 | 0 | 0 | 6 |
| French Guyana | 9 (90) | 0 | 1 (10) | 0 | 0 | 10 |
| Paraguay | 1 (100) | 0 | 0 | 0 | 0 | 1 |
| Peru | 11 (78.7) | 1 (7.0) | 0 | 0 | 2 (14.2) | 14 |
| Surinam | 3 (100) | 0 | 0 | 0 | 0 | 3 |
| Trinidad and Tobago | 3 (100) | 0 | 0 | 0 | 0 | 3 |
| Uruguay | 3 (60) | 0 | 0 | 0 | 2 (40) | 5 |
| Venezuela | 22 (88) | 2 (8) | 0 | 0 | 1 (4) | 25 |
| TOTAL | 438 (72.3) | 126 (20.8) | 14 (2.3) | 4 (0.6) | 24 (4) | 606 |
| Species | State of Development | Contaminant | Concentration | Biomarkers or Biological Alterations | Reference |
|---|---|---|---|---|---|
| Reptile Caiman latirostris | Egg | Endosulfan Atrazine DDT Oxyclordan PCB Bisfenol A 17 β-estradiol | 2/20 ppm 0.2/2 ppm Range BDL −153.0 ng g−1 lipid 52.0 ± 710.5 ng g g−1 lipid Range BDL −34.3 ng g−1 lipid 17.8 ± 73.9 ng g−1 lipid Range BDL −136.6 ng g−1 lipid 23.0 ± 74.0 ng g−1 lipid 1.4/140 ppm 0.014/1.4 ppm | Egg weight loss, reduction in hatchling fractional weight, ltered levels of steroid hormones, follicular dynamics, decreased shell porosity and number of eggs per clutch, reduced weight, and size in the young, indirect effect on survival, alteration in gene expression, impaired gonadal histoarchitecture. | [123,124,125,126,127,128,129] |
| Reptile Caiman latirostris | Egg | Endosulfan, Bisphenol A | 20 ppm 1.4 ppm | Tortuous seminiferous tubules with empty tubular lumens. BPA: Relative seminiferous tubular area was decreased. | [130] |
| Fish Cichlasoma dimerus | Adult (males, females) | Endosulfan, 17β-estradiol Octylphenol 4-tert-octylphenol | 0.1, 0.3, 1 μL L−1 10 μg g−1 body weight dose 50 μg g−1 body weight dose 30, 150 and 300 µgL−1 | Increased synthesis of vitellogenin and zona pellucida proteins, impaired testicular structure. | [131,132,133] |
| Fish Jenynsia multidentate | Adult (males) | 4n–nonylphenol | 0.20 and 40 µg L−1 | Gonadsomatic index decreased, multiple apoptotic bodies in Sertoli cells, loss of testicular cystic structure. | [134] |
| Fish Jenynsia multidentate | Adult (males) | 17α-ethinylestradiol | 10, 75 and 150 ngL−1 | Reduction in live and motile spermatozoa, increase in dead and immotile spermatozoa and sperm speed, gonadsomatic index decreased. | [135] |
| Fish Jenynsia multidentate | Adult (males) | 17β-estradiol | 50, 100, and 250 ngL−1 | Reproductive behavioral: Sexual activity increased at 50 ng L−1 E2., but not at other concentrations. No modification in gonadsomatic index and sperm quality. | [136] |
| Fish Odontesthes bonariensis | Larvae | 17α-ethinylestradiol | 0.1 and 1 µg g−1 food | Altered sex ratio, expression of cyp19a1a gen increased, expression of hsd11b2 decreased. | [137] |
| Fish Cichlasoma dimerus | Larvae | 17β-estradiol, 4-tert-octylphenol | 1 and 10 μgL−1 10 μgL−1 | High concentrations of E2: feminizing effect directing sex differentiation towards ovarian development. Lower concentration of E2: testis development was inhibited. Exposure, no impairment of male gonad development and functionality. | [138] |
| Fish Cichlasoma dimerus | Adult (male) | 4-tert-octylphenol | 150 and 300 μg L−1 | High concentration of OP: Impairment of testis architecture. Fishes were transferred to OP-free water after 60 days of exposure: at day 28 testicular functionality was recovered. | [139] |
| Fish Cichlasoma dimerus | Adult (males, females) | Endosulfan | ES-AI: 100 μM | ES by itself did not affect testosterone and estradiol levels. ES with an active ingredient caused steroidogenesis disruption. | [140] |
| Fish Odontesthes bonariensis | Adult (male) | Metals | Cd 0.25 μgL−1 Cr 4 μgL−1 Cu 22 μgL−1 Zn 211 μgL−1 | Laboratory exposure to environmental concentrations of Cd, Cr, Cu and Zn. Gonads of the fish exposed to all the tested metals suffered structural damages showing shortness of the spermatic lobules, fibrosis, testis ova and the presence of yknotic cells. With Cd: increased expression gnrh, Cd and Cr: decrease of fshb. | [141] |
| Crustacea Zilchiopsis collastinensis | Adult (females) | Endosulfan | 94 ± 6; 192 ± 10 and 360 ± 15 μg endosulfan L−1 | Changes in volume of oocytes in a certain period without change in the gonadsomatic index. | [142] |
| Crustacea Cherax quadricarinatus (invasive species) | Juvenile | Atrazine | 0.1, 0.5 and 2.5 mgL−1 | Weight gain decreased. At higher atrazine concentration the proportion of females increased gradually. | [143] |
| Crustacea Neohelice granulata | Adult (female) | Atrazine | 0.03, 0.3 and 3mgL−1 | Higher proportion of previtellogenic oocytes, reduction, and delay in the ovarian growth, vitellogenin decreases. | [144] |
| Crustacea Eurytemora americana | Adult (females) | Sewage effluents (4 different water qualities) | Fertility was reduced at bioavailable contaminants from dissolved phase of the sewage effluent. | [145] | |
| Amphibian Rhinella arenarum | Adult (males, females) | Cadmium | 0.5 and 5 mg kg−1 | Ovary: nuclear and cytoplasmic pleomorphism, vacuolization of oocytes in the early stages of development. Higher dose: increase in the proportion of atretic oocytes. Testes: seminiferous tubules dilated, disappearance of cysts, leukocyte infiltration. Decreased concentration, viability, and progressive motility of sperm | [146] |
| Amphibian Rhinella arenarum | Larvae | Fludioxonil, Metalaxyl-M | 0.25 and 2 mg L−1 | General underdevelopment, gonadal development and differentiation were impaired. | [147] |
| Amphibian Leptodactylus latrans | Larvae | Glyphosate Roundup ® | 3–300 mgL−1 0.0007–9.62 mg of acid equivalentsL−1 | Oral abnormalities and edema. Swimming activity affected. | [148] |
| Species | State of Development | Contaminant | Concentrations | Biomarkers or Alterations | Reference |
|---|---|---|---|---|---|
| Mussel Perna perna | Adult (males, females) | Coastal area of São Paulo | Reduction in embryonic development, negative impact on the community structure at one study site. | [169] | |
| Fish Gymnotus carapo | Adult (males) | Mercury chloride | 5–30 μM | Reduction in sperm count and impaired sperm morphology. Direct correlation between the accumulation of Hg and severity of lesions. | [170] |
| Fish Oreochromis niloticus (invasive species) | Adult (females) | Diuron Diuron metabolites | 100 ng L−1 | Diuron metabolites: gonadosomatic indices, percentage of vitellogenic oocytes and E2 plasma levels improved. Diuron and its metabolites: germinative cells reduction. | [171] |
| Fish Astyanax bimaculatus | Adult (females) | Endosulfan | 1.15, 2.30, and 5.60 μgL−1 | Increase in diameters of secondary follicles. Secondary follicles: increased expression of integrin β1 and collagen type IV in cytoplasm of follicular cells. | [172] |
| Fish Rhamdia quelen | Adult (males) | Paracetamol | 0.25 and 2.5 µgL−1 | Reduced testosterone levels. High concentration of paracetamol induces estradiol levels. | [173] |
| Fish Odontesthes humensis | Embryos | Glyphosate-based herbicide | 0.36 mg a.e.L−1 | Reduced eye size and distance between eyes after 96 h of exposure. | [174] |
| Fish Rhamdia quelen | Larvae | Water (polluted with PAHs and toxic metals) of Iguaçu River | Skeleton deformities such as lordosis, scoliosis, and kinks in tails. Cranial abnormalities. Thorax injuries. | [175] | |
| Mammal Artibeus lituratus | Adult (males) | Endosulfan | 1.05; 0.015 (E1) gL−1 2.1; 0.015 (E2) gL−1 | Decreased plasma glucose concentration and carcass fatty acids. | [176] |
| Species | State of Development | Contaminant | Concentrations | Biomarkers or Alterations | Reference |
|---|---|---|---|---|---|
| Mussel Aulacomya ater | Adult (males, females) | 17β-estradiol | 1 and 100 μg L−1 | Increased vitellogenin and some differences in reproductive parameters. | [188] |
| Fish Oncorhynchus mykiss (invasive species) | Juvenile (males, females) | Laboratory exposures to pulp and paper mill effluents and in situ bioassay downstream of the combined discharge of the same pulp mill | 10, 35, 60 and 85% [v/v] | Higher concentrations of plasma vitellogenin. Male fish revealed intersex characteristics in all the laboratory assays and in caged fish. Increase in the average gonadosomatic index in exposed fish. | [189] |
| Fish Oncorhynchus mykiss (invasive species) | Juveniles (females) | Sediments of different gradients of contamination from the Biobio river impacted by the pulp mill Caged trout exposure to different pollution gradients in the Biobio River Intraperitoneal injection of effluent of a cellulose plant extract | Sediment from the three sampling areas (PRE, IMP, POST), in a 1:10 w/v proportion | Increase in vitellogenin and gonadosomatic index, presence of vitellogenic oocytes, inhibition of acetylcholinesterase activity and induction of 7-ethoxyresorufin O-deethylase (EROD). | [190,191,192] |
| Fish Percilia irwini | Adults (male, females) | Laboratory exposures to wastewater treatment plant and pulp and paper mill effluents | Increased VTG-like phosphoproteins and hepatic ethoxyresorufi n-o-deethylase induction levels were detected in effluent-exposed individuals. | [193] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas-Hucks, S.; Rodriguez-Jorquera, I.A.; Nimpstch, J.; Bahamonde, P.; Benavides, J.A.; Chiang, G.; Pulgar, J.; Galbán-Malagón, C.J. South American National Contributions to Knowledge of the Effects of Endocrine Disrupting Chemicals in Wild Animals: Current and Future Directions. Toxics 2022, 10, 735. https://doi.org/10.3390/toxics10120735
Rojas-Hucks S, Rodriguez-Jorquera IA, Nimpstch J, Bahamonde P, Benavides JA, Chiang G, Pulgar J, Galbán-Malagón CJ. South American National Contributions to Knowledge of the Effects of Endocrine Disrupting Chemicals in Wild Animals: Current and Future Directions. Toxics. 2022; 10(12):735. https://doi.org/10.3390/toxics10120735
Chicago/Turabian StyleRojas-Hucks, Sylvia, Ignacio A. Rodriguez-Jorquera, Jorge Nimpstch, Paulina Bahamonde, Julio A. Benavides, Gustavo Chiang, José Pulgar, and Cristóbal J. Galbán-Malagón. 2022. "South American National Contributions to Knowledge of the Effects of Endocrine Disrupting Chemicals in Wild Animals: Current and Future Directions" Toxics 10, no. 12: 735. https://doi.org/10.3390/toxics10120735