
Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion

School of Marine Science and Technology, Harbin Institute of Technology, Huancui District, Weihai 264209, China
*
Author to whom correspondence should be addressed.
Toxics 2022, 10(6), 302; https://doi.org/10.3390/toxics10060302
Submission received: 26 April 2022
/
Revised: 29 May 2022
/
Accepted: 30 May 2022
/
Published: 2 June 2022
(This article belongs to the Section Ecotoxicology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Heavy metal pollution in the Antarctic has gone beyond our imagination. Copper toxicity is a selective pressure on Planococcus sp. O5. We observed relatively broad tolerance in the polar bacterium. The heavy metal resistance pattern is Pb2+ > Cu2+ > Cd2+ > Hg2+ > Zn2+. In the study, we combined biochemical and metabolomics approaches to investigate the Cu2+ adaptation mechanisms of the Antarctic bacterium. Biochemical analysis revealed that copper treatment elevated the activity of antioxidants and enzymes, maintaining the bacterial redox state balance and normal cell division and growth. Metabolomics analysis demonstrated that fatty acids, amino acids, and carbohydrates played dominant roles in copper stress adaptation. The findings suggested that the adaptive mechanisms of strain O5 to copper stress included protein synthesis and repair, accumulation of organic permeable substances, up-regulation of energy metabolism, and the formation of fatty acids.
1. Introduction
Over the past few years, with the high level of industrial activities and widespread pesticides and fertilizers, heavy metals have been commonly detected in diverse environments around the world and gradually accumulated [1]. Furthermore, heavy metal concentrations can be subsequently biomagnified thousands of times through biological amplification in an ecosystem [2]. Therefore, these heavy metals seriously threaten the stability of the ecological system and the health of human beings [3].
Although metals play an essential role, directly or indirectly, in vital cellular processes such as aerobic metabolism and cellular respiration, heavy metal concentrations above the maximum threshold are toxic to living things by the alterations of nucleic acids and polypeptide conformation and the disturbance of cell wall integrity, enzyme specificity, oxidative phosphorylation, and osmotic balance [4,5]. As we all know, microorganisms are found almost everywhere on earth [6]. Microbes have activated defensive strategies and evolved several adaptation mechanisms for survival [7], such as accumulation on a cell wall, transportation across the cell membrane, a permeable membrane, intracellular sequestration, and enzymatic detoxifications [5,8]. The capability of organisms to modulate their metabolism is a central characteristic required for proliferation, hibernation, and survival [9]. The metabolic mechanism of microbial resistance to heavy metal could be elucidated in detail from different perspectives, which we shall describe next. Adjusting the fatty acid composition of the cell membrane and reconfiguring energy-generating processes have been seen as the most efficient adaptation mechanisms to heavy metals [10]. Under metal stress, partial microorganisms fulfill their energy requirements through substrate-level phosphorylation rather than oxidative phosphorylation [11]. For example, the resistance of Proteobacteria to heavy metals may be because the phylum can utilize a variety of organics as carbon and energy sources [12]. The presence of large amounts of long-chain fatty and phospholipid saturation contributes to low fluidity and rigidity of the membrane to prevent the bacterial cell from contaminants [13,14]. Active efflux and precipitation are in bacterial partial resistance toward metals [15], shown in the acidophilic bacterium Acidithiobacillus ferrooxidans and Bacillus sphaericus [16,17,18]. In Escherichia coli, the addition of copper (Cu) stimulates the degradation of intracellular polyphosphate, and phosphate exportation also supports this mechanism [19]. In addition, heavy metals are precipitated and eliminated by compounds produced by microorganisms under heavy metal stress, contributing to a degree of bacterial resistance to metals [15]. To circumvent aluminum (Al) toxicity, Pseudomonas fluorescens promotes the synthesis of citrate involved in the sequestration of Al [20]. Desulfovibrio desulfuricans can regulate the precipitation of metals by forming metal sulfides [21]. The metabolic level of low molecular weight organic acids can be up-regulated to dissolve heavy metals by using them as a final electron acceptor or decreasing pH [22,23]. Oxalobacter formigens absorb as minimal as possible Pb by converting it to oxalate that can be excreted from the gut [24]. Interestingly, bacteria increase stress response-related metabolites to rebalance oxidative stress and osmotic pressure damaged by heavy metals. For instance, Scenedesmus obliquus increases lipid esters and Cys-GSH isomers for antioxidant defense mechanisms and reactive oxygen species prevention under cadmium stress [25]. In addition, ethanol tolerance involves increased glycine metabolism, which serves as protective osmolytes in Escherichia coli [26].
The Antarctic, an isolated place and often considered a clean slate, is facing the challenge of negative factors derived from human activities [27,28,29]. Unfortunately, heavy metals have been detected in abiotic samples such as surface soil, atmosphere particulate, and snow in Antarctica [30]. Furthermore, heavy metal concentrations in Southern Ocean organisms are significantly higher than in other oceans [29]. The migration and accumulation of heavy metals have become one of the severe problems in Antarctica. In fact, Cu is one of the most common sources of heavy metals contributing to contamination in Antarctica [31]. As an essential micronutrient, Cu is employed by most organisms to perform different functions, such as acting as a catalytic cofactor in cellular redox reactions and metal homeostasis [32,33,34]. However, an excessive amount of copper can be toxic.
Although the adaptation strategies to Cu have been relatively well characterized, the metabolic reprogramming leading to stress-induced lifestyle changes in polar microorganisms remains a mystery. The sensitivity of organisms to contamination may vary with latitude [35]. Polar organisms have evolved unique characteristics to adapt to severe regimes at high latitudes, including lower metabolisms, longer lifespans, and higher lipid content in tissues [36]. However, most research focuses on low-temperature enzyme production and low-temperature adaptation mechanisms, which ignores unique metabolic mechanisms that adapt to the environment [37,38]. Strain O5 isolated from Antarctic sea ice was subsequently identified as Planococcus based on 16S rDNA sequence analysis [39].
In this study, cell growth and physiological and biochemical variations of the Antarctic bacterium Planococcus sp. O5 after Cu2+ exposure were analyzed to explore the tolerance mechanism of the bacterium to Cu. The results will help elucidate the adaptation mechanism of polar microorganisms under heavy metal exposure. Meanwhile, the strain has extensive tolerance to heavy metals, which can be applied to deal with the heavy metal pollution in Antarctic in the future.
2. Materials and Methods
2.1. Bacterial Strains and Culture
The Antarctic strain Planococcus sp. O5 was isolated from Antarctic sea ice collected by the 23rd China Antarctic scientific expedition. To investigate the growth effect of Cu2+ exposure, strain O5 was cultured in 2216E liquid medium (5.0 g of peptone, 1.0 g of yeast extract, and 0.015 g of iron phosphate tetrahydrate in 1000 mL of purified and sterilized seawater) at 10 °C with the agitation of 120 rpm. Additional CuSO4 (final concentration 0.5 mmol/L) was added as copper stress.
2.2. MIC Determination of Heavy Metals
The minimum inhibitory concentration (MIC) of strain O5 was tested as described by Rajpert [40]. Various metal resistance tests were performed in log phase culture of strains that were inoculated in 2216E liquid medium supplemented with Cu2+, Cd2+, Pb2+, Zn2+, and Hg2+ in the concentration ranging from 0 mM to 1500 mM. Strain growth was monitored using OD595 measurement with a UV spectrophotometer. The lowest metal concentration that hampered growth was regarded as the MIC of the test strain against metal.
2.3. Measurements of Electrical Conductivity and Biomass
The membrane permeability was detected using a conductivity meter, and the biomass was measured using OD595 [41].
2.4. Measurements of Antioxidant System
Changes in the antioxidant enzyme activities were determined to understand the influence of Antarctic bacterium under copper stress. For this purpose, 100 mg of fresh weight (FW) bacterial strain was homogenized in 20 mL 50 mM phosphate buffer (pH 7.8) using a prechilled mortar and pestle before centrifugation at 12,000 rpm for 30 min at 4 °C. The collected supernatants were employed to determine the antioxidant enzyme activities of superoxide dismutase (SOD), glutathione reductase (GR), ascorbate peroxidase (APX), and the content of glutathione (GSH) and carotenoid.
SOD activity was assayed as described by Zhang based on the inhibition of the oxidation inhibition rate of pyrogallol reaching 50% [42], and GR was measured following the method of Pinto, Mata, and Lopezbarea. GR activity was measured in OD340/(min•g FW) [43]. APX activity was measured using the method of Nakano, Y. and Asada, K. A unit of the enzyme activity was defined as ascorbic acid consumed by the bacterium (min•g FW) [44]. GSH content was performed as per the report described by Yoon and was measured in OD400 [45]. The multiparameter flow cytometry method documented by Freitas [46] was used to assay carotenoid content.
2.5. GC-MS analysis of Metabolites
2.5.1. Sample Preparation
The bacteria supplemented with 0.5 mmol/L Cu2+ in the logarithmic and stable phases were acquired by centrifugation at 12,000 rpm at 4 °C for 5 min, respectively. Subsequently, 2 mL of 60% precooled methanol (−40 °C) were added and placed on ice for 5 min to quench the cellular reaction. After centrifugation, the collected cell pellet was introduced to 0.5 mL of methanol (50%, −40 °C), followed by rupturing with the sonication method. The broken cells were centrifuged (4 °C, 12,000 rpm, 10 min), and 10 μL of succinic-2,2,3,3-d 4 acid (0.3 mg/mL) was added to the supernatant. When the sample was dried, 100 μL of 20 mg/mL pyridine amine hydrochloride was added and subsequently oxidated at 30 °C for 1.5 h. Afterward, MSTFA (100 mL) was used to derivate the samples by incubation at 37 °C for 0.5 h.
2.5.2. GC-MS Analysis of Metabolites
The metabolites were analyzed using GC-TOF-MS (Agilent 7890A, Santa Clara, CA, USA), and 1 μL of the derivatized sample was injected into GC-MS, which was equipped with a DB-FFAP capillary column (60m × 0.25 μm × 0.25μm). The elution program setting: isothermal at 80 °C for 1 min, then an increase of 2 °C min−1 up to 100 °C, and ramped at 4 °C min−1 to 240 °C, and then held for 15 min at 240 °C. The ion source temperature was maintained at 200 °C. The mass spectrometer was set to scan a mass range of 50–800 m/z at 20 scans/s with an electron beam of 70 eV.
2.5.3. Data Processing
The peak integration and peak alignment were conducted by applying the XCMS package of R software, and components were manually identified and confirmed using the NIST library. Noise and low abundance components were eliminated from the data matrix based on a noise threshold (S/N > 10). The ultimate two-dimensional matrix consisted of retention time (RT) and mass-to-charge ratio (m/z) data pairs.
2.5.4. Statistical Analysis
The processed data matrix was submitted to the MetaboAnalyst 4.0 (http://www.metaboanalyst.ca/, accessed on 4 March 2022) to conduct data pre-processing and multivariate statistical analysis. Data were normalized to total integral normalization before being log-transformed. Principal component analysis (PCA) was performed to provide a general overview and remove irrelevant variables. Orthogonal partial least squares discriminant analysis (OPLS-DA) was conducted to further investigate the metabolic variation. Metabolites of interest were filtered using volcano plots with fold change (FC) ≥1.2 and p-value < 0.05. To explore the related metabolic pathways for differential metabolites, compounds of interest were imported into the Pathway modules of MetaboAnalyst.
3. Results
3.1. Heavy Metals Resistance Analysis
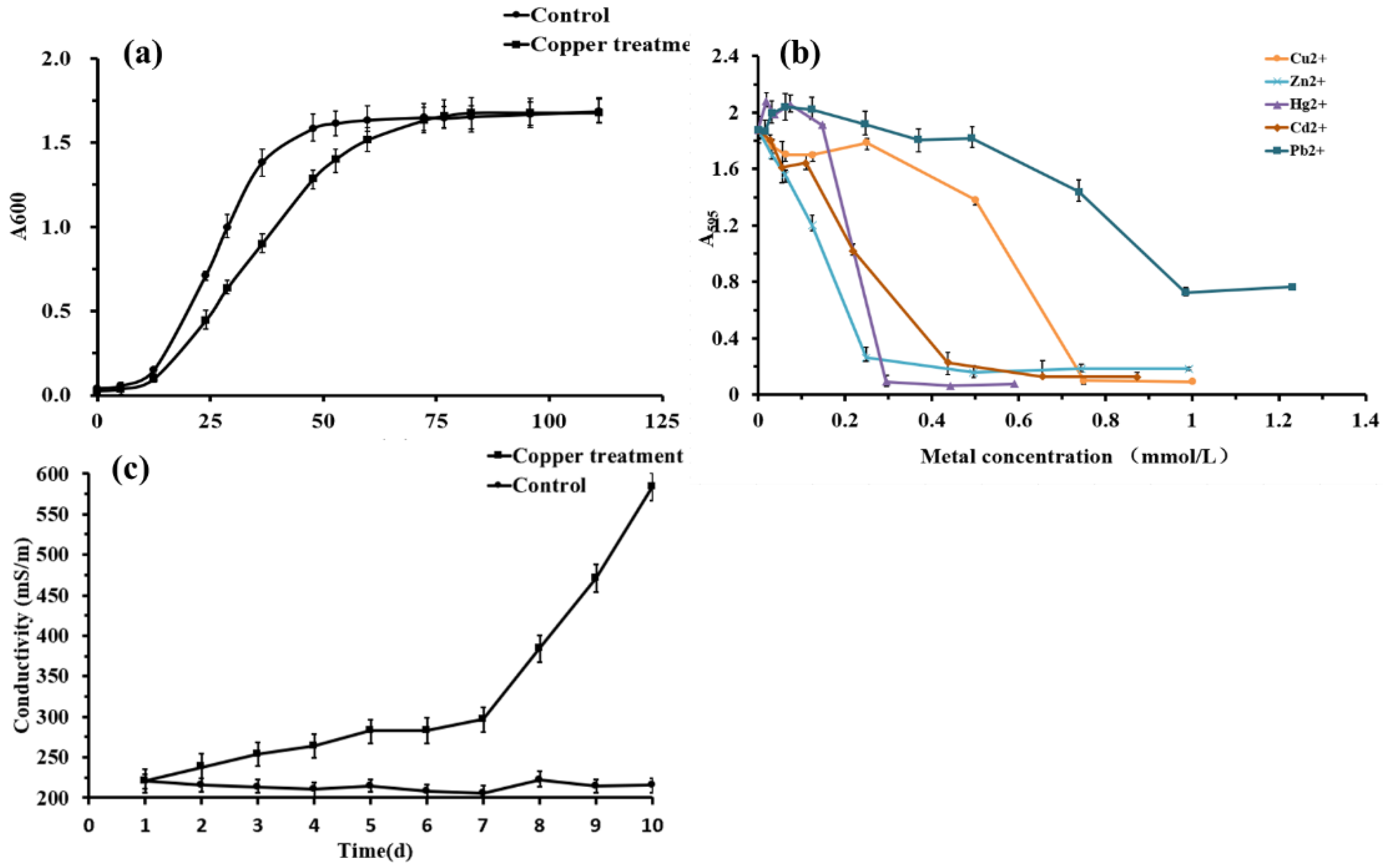
Typical bacterial growth was observed in the induced and normal groups (Figure 1a). Although the growth rate of Planococcus sp. O5 in the normal group was higher in the induced group in the first 72 h, the growth of the copper exposure group presented an equal OD595 value to the untreated group subsequently. The relative metal resistance of strain O5 was in the order of Pb2+ > Cu2+ > Cd2+ > Hg2+ > Zn2+ (Figure 1b), and the MIC reached 1.0 mmol/L, 0.8 mmol/L, 0.7 mmol/L, 0.6 mmol/L, 0.5 mmol/L, respectively. Meanwhile, the optimum concentration of copper stress was determined to be 0.5 mmol/L.
3.2. Change in the Membrane Permeability
Electrical conductivity provides an indirect indication of membrane permeability. The conductivity of the control was stably maintained between 200–230 mS/m, as shown in Figure 1c. Correspondingly, the conductivity of the bacteria-induced with Cu2+ increased slowly on days 1–7 before dramatically increasing on day 10, reaching 586 mS/m. These findings suggested that the integrity of the cell membrane was altered after stimulation with 0.5 mmol/L Cu2+.
3.3. Response of the Antioxidant System
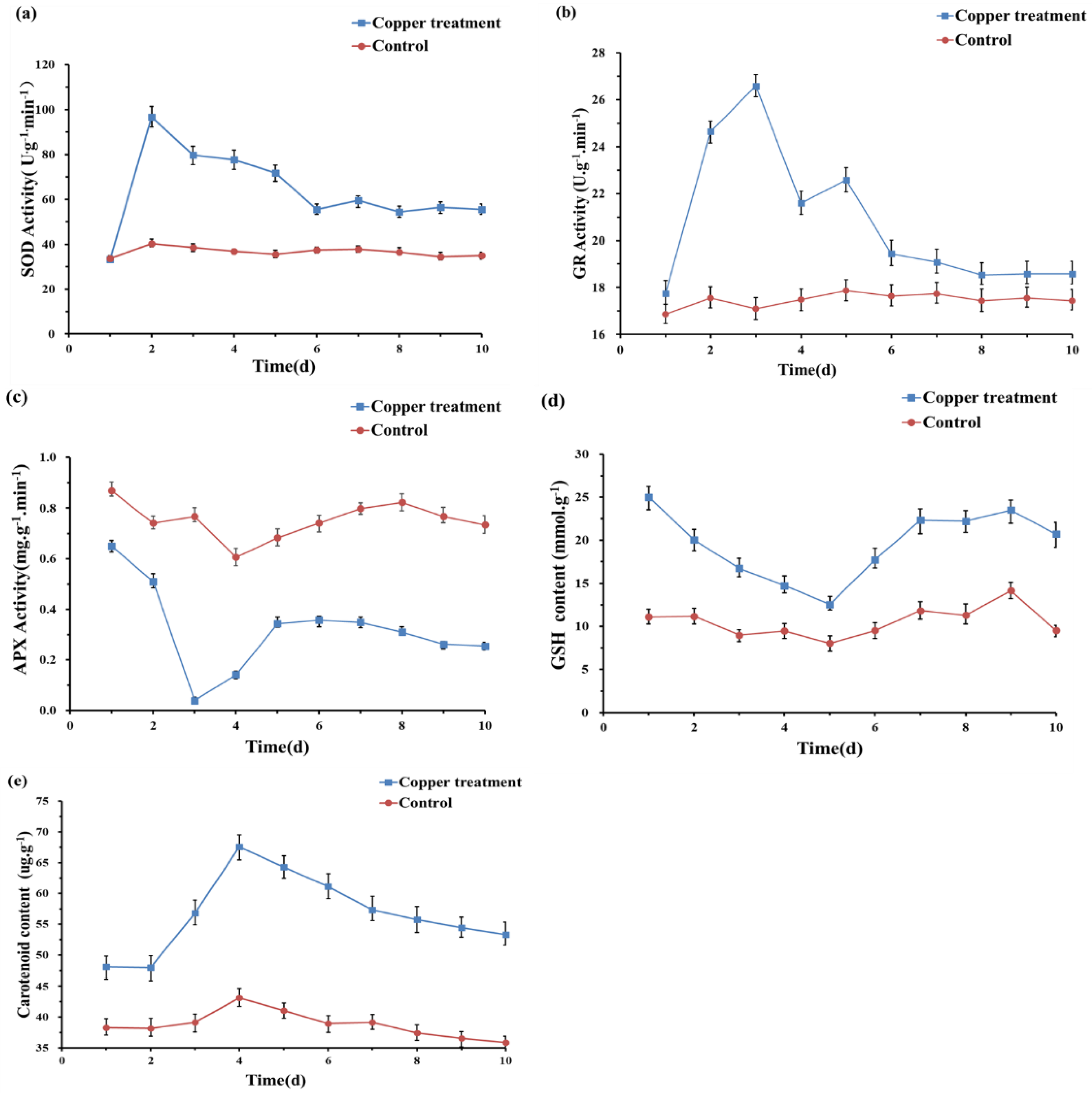
We also measured the changes in the content of antioxidant substances and the activities of antioxidant enzymes further to understand the biochemical mechanism of strain O5 to Cu, as shown in Figure 2. The enzymatic activity of SOD, GR, and APX remained almost unchanged throughout the experiment without Cu2+ induction. However, SOD and GR activities rose fast after the exposure to 0.5 mmol/L Cu2+, reaching their maximum value on day 2 and 3, respectively. In contrast, APX activity decreased and was significantly lower than the control. Carotenoid and GSH content accumulated rapidly under 0.5 mmol/L Cu2+ stress and remained considerably higher content than in the untreated group.
3.4. Metabolic Response of Strain O5 to Cu Induction
3.4.1. Metabolic Profile Analysis
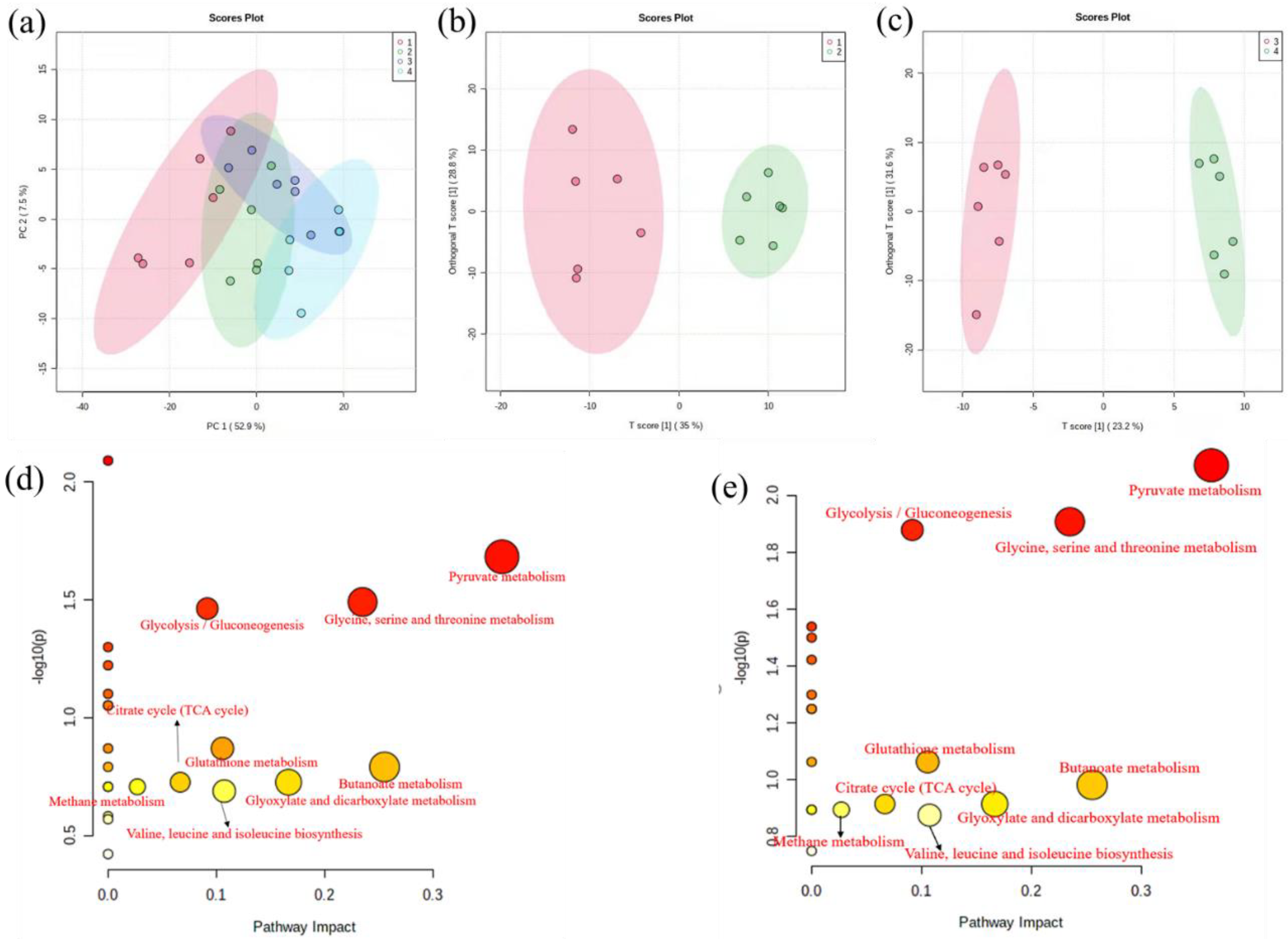
In the PCA score plot (Figure 3a), all 24 data samples were within the 95% confidence ellipse, indicating that no sample contained outliers. Although there was a noticeable separation between the Cu exposure and the control groups in the PCA, the treatment and control groups were intermixed. To further investigate the metabolic variation induced by copper stress, OPLS-DA was performed. As demonstrated in Figure 3b,c, the treatment and control groups showed a more distinct separation. The R2 values for the OPLS-DA model were satisfactory (>0.945, >0.99), explaining the majority of the variance between the samples. The Q2 values were much higher (>0.728, 0.7), indicating that the vast majority of the variation was to be expected.
3.4.2. Identification and Analysis of Differential Metabolites
To discover the significant alterations of metabolites induced by copper exposure, we evaluated the changes in metabolites abundances using the filtering function of volcano plots. In total, 13 different metabolites were filtered based on the FC and the p-value with 4 significantly reduced and 9 increased (Table S1). The differential metabolites included energy, amino acid acids, and organic acids.
3.4.3. Perturbed Biological Pathway Responded to Copper Stress
Pathway analysis was performed to investigate relevant pathways connected to Cu response, with the results shown in Figure 3d,e. The color shades and circle size were based on p-values and pathway impact values. Redder and large pathway circles indicated that the pathway was greatly perturbed. MetaboAnalyst Pathway found 7 key metabolic pathways in the logarithmic phase using pathway enrichment analysis, with pyruvate metabolism, butanoate metabolism, and glycine serine and threonine metabolism showing the most pronounced changes. Similarly, the most significant changes in the metabolic pathway also had the most critical impact during the stable growth phase. The p-value and impact factor of significant pathways are shown in Tables S2 and S3.
4. Discussion
Heavy metal ions, which are highly toxic, non-degradable, bioaccumulate, and biomagnifying as a result of the food chain, constitute a severe threat to ecological environments. Copper ions are micronutrients essential for the biological functions of living organisms. However, excess copper ions in cells are detrimental through various induced physiological, biochemical, and genotoxic effects [47]. Only copper-resistant microorganisms can strive and utilize trace metals to achieve metabolic functions while resisting or detoxifying their excesses. To discover the adaptation strategies of Antarctic microorganisms to heavy metal (Cu) stress, we combined biochemical and metabolomics methods to analyze the bacterium Planococcus sp. O5, isolated from the Antarctic sea ice sample.
4.1. Heavy Metals Resistance
In this study, the Antarctic bacterium Planococcus sp. O5 exhibited a relatively broad tolerance to Cu2+, Hg2+, Zn2+, Cd2+ and Pb2+, and especially Pb2+ and Cu2+ (Figure 1b). Similar bacterial resistance to multiple heavy metals was reported in other Antarctic strains, such as Antarctic Rhodotorula mucilaginosa resistance pattern of Cd2+ > Pb2+ = Mn2+ > Cu2+ > Cr3+ > Hg2+, and Antarctic bacteria isolated from rock lichen resistance pattern of Cr3+ > Ni3+ > Cu2+ > Co2+ > Hg2+ [48,49]. Multiple heavy metals resistance is attributed to these metals with similar toxic mechanisms and detoxifying processes [50]. In addition, the growth curve of bacteria appeared that the Antarctic sea-ice bacterium challenged by copper has adapted to the presence of 0.5 mmol/L Cu2+ (Figure 1a). In addition, the emergence of a lag phase may be due to oxidative stress induced by copper requiring the consumption of glutathione (GSH), whose synthesis requires additional energy. A similar adaptation mechanism has been observed in Aspergillus niger [51].
4.2. Effect on Redox Status
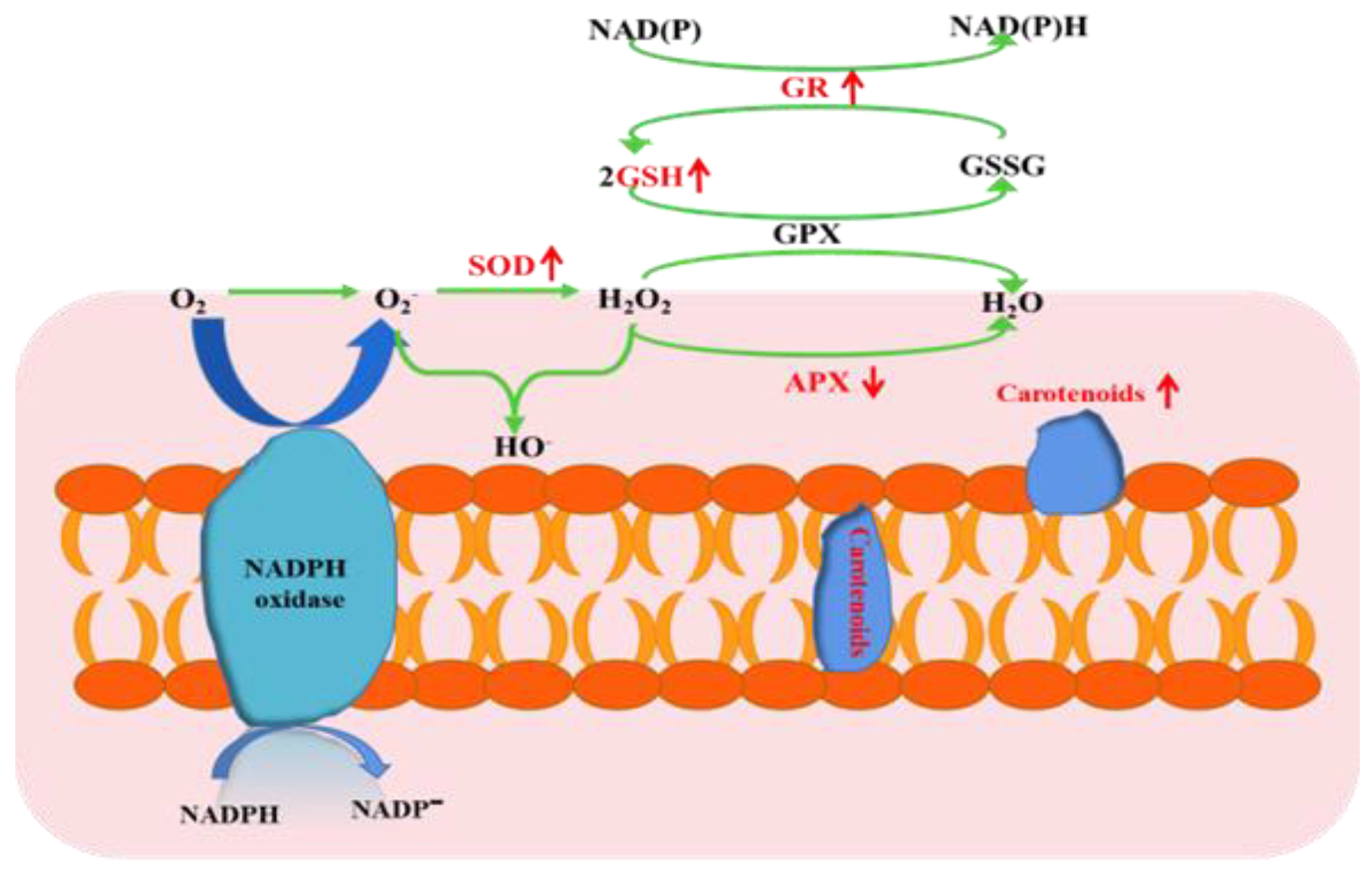
Reactive oxygen species (ROS), including superoxide anion (O2−), hydroxyl radical (∙OH), and hydrogen peroxide (H2O2), are among the significant toxicities of heavy metals to most living organisms by altering the reducing environment [52,53]. The membrane structure of polar bacterium was damaged by lipid peroxidation (Figure 1c), and similar responses occurred in other microorganisms [54]. However, strain O5 could take advantage of various antioxidative defense systems to rebalance the redox status (Figure 4) [55]. Glutathione (GSH), an essential indicator of the redox environment, plays a major role in cellular defense response against oxidative stress [56]. The copper resistance expressed in our study may also be explained by the activation of glutathione, which is rich in thiol groups and may be related to a metal complexation mechanism via a rich sulfur bond. This detoxification mechanism has been demonstrated in yeast [57]. Therefore, the increase in GR activity was entirely expected. The GR activity boosted the regeneration efficiency of GSH to maintain the intracellular redox balance. GR catalyzes GSSG into GSH in the presence of coenzyme β-nicotinamide adenine dinucleotide 2′-phosphate hydrate (NADPH) [58], which has been regarded as being related to resistance to oxidative stress in a microorganism [59]. Carotenoids are mainly located in the cell membrane, acting as antioxidant protectants for the cell membrane integrity [60]. In short, it is of great significance to regulate different antioxidant mechanisms when facing endogenous redox damage induced by copper.
4.3. Metabolic Reprogramming
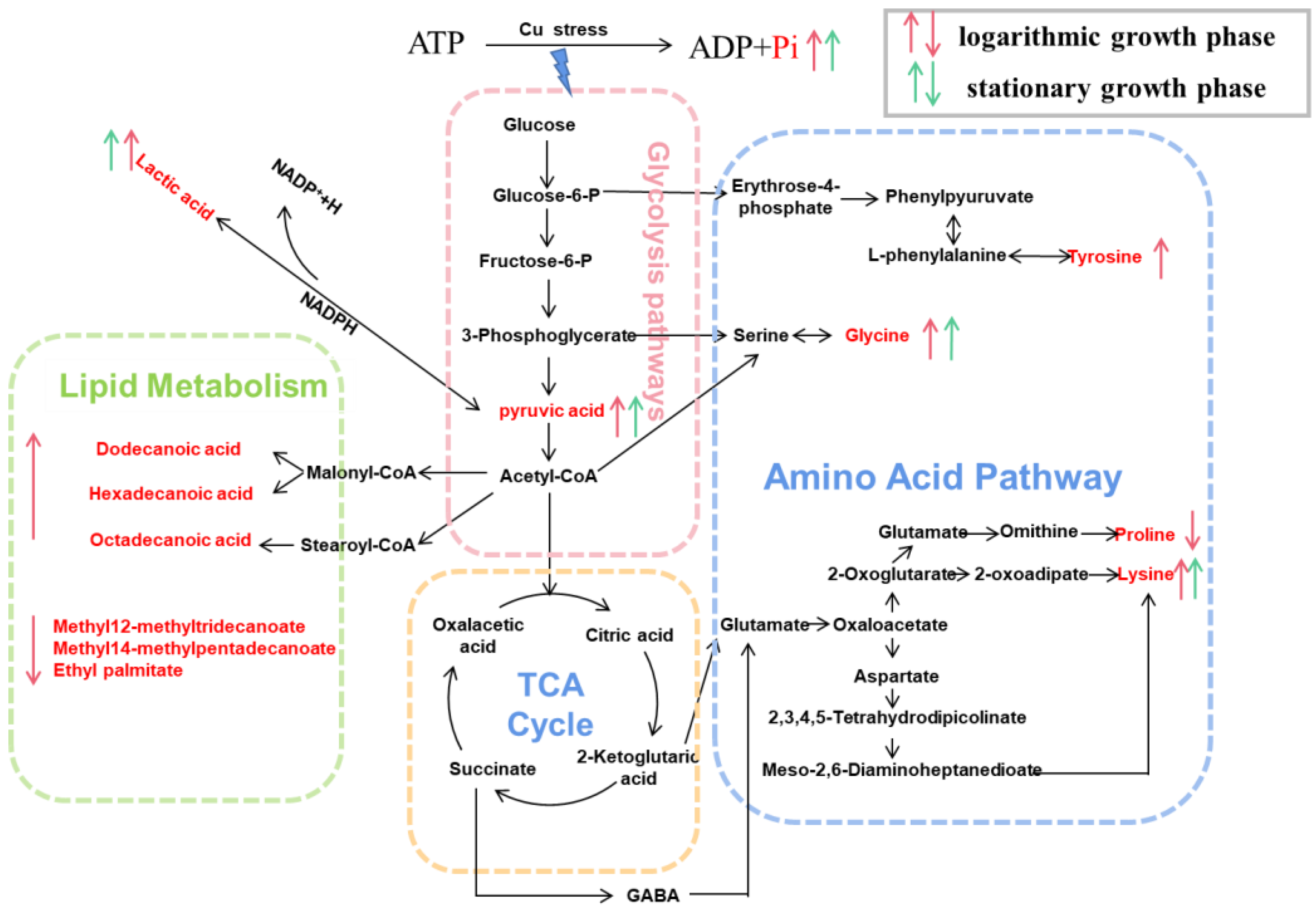
The pathway enrichment analysis indicated that copper stress could generate metabolic reprogramming, resulting in alterations in many metabolites, particularly in energy, amino acid acids, and lipid metabolism, forming a metabolic network to deal with copper stress (Figure 5).
4.3.1. Energy Metabolism
Carbohydrate and energy metabolism acted as key attributions in the adaptive responses to heavy metals [49]. In organisms, lactic acid is a byproduct of anaerobic metabolism, and pyruvic, an end product of glycolysis by the Embden Meyerhof pathway, is conversed to lactate acid by oxidizing nicotinamide adenine dinucleotide (NADH) in the presence of lactate dehydrogenase [61]. Bacteria activated the breakdown of sugars to generate more energy and amino acids as a defensive mechanism when exposed to copper. As a result, pyruvate acid and NADPH accumulated abundantly, leading to an aggregation of lactic acid. This energy model might alleviate the cytotoxicity of extreme Cu by reducing ROS levels [62].
4.3.2. Amino Acid Metabolism
Amino acids are commonly used in all living cells for osmoregulation, energy sources, protein synthesis, metabolite precursors, and signaling molecules [63]. Glycine, an organic osmolyte [64], was reported to be closely associated with the Cu2+ tolerance of Pseudomonas [26]. In addition, glycine is a precursor of glutathione [65] and can alleviate oxidative damage. A previous study demonstrated that lysine altered NADPH flux to produce glutathione, enhancing tolerance to oxidative stress [66]. Hence, in this study, lysine-overproducing was expected to exhibit higher tolerance to copper stress. Surprisingly, proline content declined, which had not been observed in other organisms responding to environmental stresses [67,68,69,70]. Proline metabolism is highly relevant to redox homeostasis, protein and nucleotide synthesis, and ATP production, and is especially closely associated with the progression of oxidative stress [71]. Consequently, in this study, the down-regulation of proline demonstrated the oxidative balance was disrupted under copper stress, leading to lipid peroxidation [72]. Tyr is susceptible to modification under conditions of cellular redox imbalance. The oxidation of phenylalanine is a marker of oxidative stress [73]. Under conditions of ROS overaccumulation, tyrosine can be formed by phenylalanine hydroxylation or oxidation [74]. Increased concentrations of Tyr were also observed in plants in response to biological stress [75]. Moreover, the upregulation of glycine, lysine, and tyrosine was considered to repair damaged proteins and activate the synthesis of newer proteins [76]. The mentioned studies implicated the protein biosynthesis mechanisms enhanced by excessive copper [77].
4.3.3. Lipid Metabolism
Stearic acid and palmitic acid were identified as quantitative markers of cellular stress, with which the overproduction and accumulation of ROS have been frequently associated [78]. This research implied the disruption of membrane integrity by lipid peroxidation (Figure 1c). This damage mechanism induced by copper has also been previously reported in filamentous fungus Paecilomyces marquandii [79]. Membranes of bacteria consist mainly of a lipid bilayer and embedded proteins, which allow solutes to selectively transport substances across the membrane to facilitate physiological processes such as respiration and signal transduction [80]. Integrity and fluidity of cell membranes influenced by lipids composition and the unsaturation degree of fatty acids are critical for organism survival in response to external stress [81]. Stress factors usually lead to lipid metabolism reconfiguration, resulting in decreased or increased membrane fluidity [82]. A low fluidity of the cell membrane can be perceived to effectively prevent copper ions from entering the bacteria cells [83,84]. As expected, saturated fatty acids have identified upregulation. In brief, the regulation of fatty acids significantly enhanced the resistance of copper and oxidative.
Copper stress resulted in metabolic reprogramming, according to metabolic profile analyses. In response to copper adaptation, bacteria activated the breakdown of intracellular sugars to generate more energy. In addition, amino acids support diverse functions, including maintaining appropriate cell status by increasing osmotic substances, producing new stress proteins, and repairing damaged or misfolded proteins. Furthermore, the accumulation of fatty acids may reduce the fluidity of the cell membrane, which can prevent copper ions from entering the bacteria cells.
5. Conclusions
Planococcus sp. O5 presented a wide range of heavy metal resistance, such as Pb, Cu, Cd, Hg, and Zn. In the study, we employed a system analysis strategy by integrating biochemical and comparative metabolomics to shed light on the adaptive mechanism of the polar bacterium to Cu2+. Our results indicated that strain O5 exhibited a relatively broad tolerance to Cu2+, Hg2+ Zn2+, Cd2+, and Pb2+, especially Pb2+ and Cu2+. Under copper pressure, strain O5 maintained intracellular redox balance by increasing antioxidant enzymes (SOD, GR) and antioxidant substances (GSH, Carotenoid). Planococcus sp. O5 in the presence of Cu stress achieved inherently different metabolite profiles, including amino acids, organic acids, and fatty acids. The adaption mechanism of strain O5 has been introduced in protein synthesis and repair, organic osmolyte accumulation, energy metabolism up-regulation, and fatty acids formation. This study laid a theoretical basis for revealing the response of biochemical and metabolomic mechanisms in polar bacterium to heavy metals, while also providing a new perspective on the bioremediation of metal-polluted environments.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/toxics10060302/s1, Table S1: The list of differential metabolites between copper exposed group and untreated group in the logarithmic phase; Table S2: The list of differential metabolites between copper exposed group and untreated group in the stable phase; Table S3: The P-value and impact factor of significant pathways in the logarithmic phase; Table S4: The P-value and impact factor of significant pathways in the stable phase; Figure S1: Permutation test chart of copper exposed group and untreated group in the stable phase; Figure S2: Permutation test chart of copper exposed group and untreated group in the logarithmic phase.
Author Contributions
Z.C., writing—original draft preparation and formal analysis; C.S. and X.G., methodology, validation and formal analysis; X.W., investigation; G.K., writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Shandong Provincial Natural Science Foundation, China [ZR2021MC180], and Natural Scientific Research Innovation Foundation in Harbin Institute of Technology [2019KYCXJJYB17].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thank you to the scientists of the 23rd China Antarctic Expedition for donating experimental samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaur, N.; Flora, G.; Yadav, M.; Tiwari, A. A review with recent advancements on bioremediation-based abolition of heavy metals. Environ. Sci. Process. Impacts 2014, 16, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Malidareh, H.B.; Mahvi, A.; Yunesian, M.; Alimohammadi, M.; Nazmara, S. Effect of fertilizer application on paddy soil heavy metals concentration and groundwater in North of Iran. Middle-East J. Sci. Res. 2014, 20, 1721–1727. [Google Scholar]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; McHenry, M.P. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Pal, A.; Bhattacharjee, S.; Saha, J.; Sarkar, M.; Mandal, P. Bacterial survival strategies and responses under heavy metal stress: A comprehensive overview. Crit. Rev. Microbiol. 2022, 48, 327–355. [Google Scholar] [CrossRef]
- Sogin, E.M.; Leisch, N.; Dubilier, N. Chemosynthetic symbioses. Curr. Biol. 2020, 30, R1137–R1142. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Zhu, J.; Rensing, C.; Liu, Y.; Gao, S.; Chen, W.; Huang, Q.; Liu, Y.R. Recent advances in exploring the heavy metal(loid) resistant microbiome. Comput. Struct. Biotechnol. J. 2021, 19, 94–109. [Google Scholar] [CrossRef]
- Nanda, M.; Kumar, V.; Sharma, D.K. Multimetal tolerance mechanisms in bacteria: The resistance strategies acquired by bacteria that can be exploited to ‘clean-up’ heavy metal contaminants from water. Aquat. Toxicol. 2019, 212, 1–10. [Google Scholar] [CrossRef]
- MacLean, A.; Bley, A.M.; Appanna, V.P.; Appanna, V.D. Metabolic manipulation by Pseudomonas fluorescens: A powerful stratagem against oxidative and mental stress. J. Med. Microbiol. 2020, 69, 339–346. [Google Scholar] [CrossRef]
- Jurkiewicz, P.; Olzynska, A.; Cwiklik, L.; Conte, E.; Jungwirth, P.; Megli, F.M.; Hof, M. Biophysics of lipid bilayers containing oxidatively modified phospholipids: Insights from fluorescence and EPR experiments and from MD simulations. Biochim. Biophys. Acta 2012, 1818, 2388–2402. [Google Scholar] [CrossRef] [Green Version]
- Alhasawi, A.; Costanzi, J.; Auger, C.; Appanna, N.D.; Appanna, V.D. Metabolic reconfigurations aimed. at the detoxification of a multi-mental stress in Pseudomonas fluorescens: Implications for the bioremediation of metal pollutants. J. Biotechnol. 2015, 200, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Barker-Finkel, J.; Galloway, T.S.; Handy, R.D.; Ford, T.E. Temporal bacterial diversity associated with metal-contaminated river sediments. Ecotoxicology 2010, 19, 317–328. [Google Scholar] [CrossRef]
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J.; Rafique, M.; Rizwan, M. Heavy metal remediation and resistance mechanism of Aeromonas, Bacillus, and Pseudomonas: A review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Murinova, S.; Dercova, K. Response mechanisms of bacterial degraders to environmental contaminants on the level of cell walls and cytoplasmic membrane. Int. J. Microbiol. 2014, 2014, 873081. [Google Scholar] [CrossRef] [Green Version]
- Dave, D.; Sarma, S.; Parmar, P.; Shukla, A.; Goswami, D.; Shukla, A.; Saraf, M. Microbes as a boon for the bane of heavy metals. Environ. Sustain. 2020, 3, 233–255. [Google Scholar] [CrossRef]
- Alvarez, S.; Jerez, C.A. Copper ions stimulate polyphosphate degradation and phosphate efflux in Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2004, 70, 5177–5182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulakovskaya, T. Inorganic polyphosphates and heavy metal resistance in microorganisms. World J. Microbiol. Biotechnol. 2018, 34, 139. [Google Scholar] [CrossRef] [PubMed]
- Merroun, M.L.; Raff, J.; Rossberg, A.; Hennig, C.; Reich, T.; Selenska-Pobell, S. Complexation of uranium by cells and S-layer sheets of Bacillus sphaericus JG-A12. Appl. Environ. Microbiol. 2005, 71, 5532–5543. [Google Scholar] [CrossRef] [Green Version]
- Grillo-Puertas, M.; Schurig-Briccio, L.A.; Rodriguez-Montelongo, L.; Rintoul, M.R.; Rapisarda, V.A. Copper tolerance mediated by polyphosphate degradation and low-affinity inorganic phosphate transport system in Escherichia coli. BMC Microbiol. 2014, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J.; Lemire, J.; Kalyuzhnyi, S.; Appanna, V. A novel metabolic network leads to enhanced citrate biogenesis in Pseudomonas fluorescens exposed to aluminum toxicity. Extremophiles 2008, 12, 451–459. [Google Scholar] [CrossRef]
- Yin, K.; Wang, Q.; Lv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Ferreira, P.A.A.; Bomfeti, C.A.; Soares, C.R.F.d.S.; Soares, B.L.; Moreira, F.M.d.S. Cupriavidus necator strains: Zinc and cadmium tolerance and bioaccumulation. Sci. Agric. 2018, 75, 452–460. [Google Scholar] [CrossRef]
- Ghavidel, A.; Naji Rad, S.; Alikhani, H.A.; Sharari, M.; Ghanbari, A. Bioleaching of heavy metals from sewage sludge, direct action of Acidithiobacillus ferrooxidans or only the impact of pH? J. Mater. Cycles Waste Manag. 2017, 20, 1179–1187. [Google Scholar] [CrossRef]
- Eggers, S.; Safdar, N.; Sethi, A.K.; Suen, G.; Peppard, P.E.; Kates, A.E.; Skarlupka, J.H.; Kanarek, M.; Malecki, K.M.C. Urinary lead concentration and composition of the adult gut microbiota in a cross-sectional population-based sample. Environ. Int. 2019, 133 Pt A, 105122. [Google Scholar] [CrossRef] [PubMed]
- Mangal, V.; Nguyen, T.Q.; Fiering, Q.; Gueguen, C. An untargeted metabolomics approach for the putative characterization of metabolites from Scenedesmus obliquus in response to cadmium stress. Environ. Pollut. 2020, 266 Pt 2, 115123. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.C.; Weljie, A.M.; Turner, R.J. Metabolomics reveals differences of metal toxicity in cultures of Pseudomonas pseudoalcaligenes KF707 grown on different carbon sources. Front. Microbiol. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargagli, R.; Agnorelli, C.; Borghini, F.; Monaci, F. Enhanced deposition and bioaccumulation of mercury in Antarctic terrestrial ecosystems facing a coastal polynya. Environ. Sci. Technol. 2005, 39, 8150–8185. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Casella, P.; Bruni, V.; Michaud, L. Response of bacterial isolates from Antarctic shallow sediments towards heavy metals, antibiotics and polychlorinated biphenyls. Ecotoxicology 2013, 22, 240–250. [Google Scholar] [CrossRef]
- Bargagli, R. Trace metals in Antarctica related to climate change and increasing human impact. Rev. Environ. Contam. Toxicol. 2000, 166, 129–173. [Google Scholar]
- Chu, W.L.; Dang, N.L.; Kok, Y.Y.; Yap, K.S.I.; Phang, S.M.; Convey, P. Heavy metal pollution in Antarctica and its potential impacts on algae. Polar Sci. 2019, 20, 75–83. [Google Scholar] [CrossRef]
- Puasa, N.A.; Zulkharnain, A.; Verasoundarapandian, G.; Wong, C.Y.; Zahri, K.N.M.; Merican, F.; Shaharuddin, N.A.; Gomez-Fuentes, C.; Ahmad, S.A. Effects of Diesel, Heavy Metals and Plastics Pollution on Penguins in Antarctica: A Review. Animals 2021, 11, 2505. [Google Scholar] [CrossRef] [PubMed]
- Ravet, K.; Pilon, M. Copper and iron homeostasis in plants: The challenges of oxidative stress. Antioxid. Redox Signal. 2013, 19, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; Sitthisak, S.; Sengupta, M.; Johnson, M.; Jayaswal, R.K.; Morrissey, J.A. Copper stress induces a global stress response in Staphylococcus aureus and represses sae and agr expression and biofilm formation. Appl. Environ. Microbiol. 2010, 76, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barwinska-Sendra, A.; Waldron, K.J. The Role of Intermetal Competition and Mis-Metalation in Metal Toxicity. Adv. Microb. Physiol. 2017, 70, 315–379. [Google Scholar] [PubMed]
- Sfiligoj, B.J.; King, C.K.; Candy, S.G.; Mondon, J.A. Determining the sensitivity of the Antarctic amphipod Orchomenella pinguides to metals using a joint. model of survival response to exposure concentration and duration. Ecotoxicology 2015, 24, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Chapman, P.M.; Riddle, M.J. Toxic effects of contaminants in polar marine environments. Environ. Sci. Technol. 2005, 39, 200A–207A. [Google Scholar] [CrossRef] [Green Version]
- Fenice, M. The Psychrotolerant Antarctic Fungus Lecanicillium muscarium CCFEE 5003: A Powerful Producer of Cold-Tolerant Chitinolytic Enzymes. Molecules 2016, 21, 447. [Google Scholar] [CrossRef] [Green Version]
- Yusof, N.A.; Hashim, N.H.F.; Bharudin, I. Cold Adaptation Strategies and the Potential of Psychrophilic Enzymes from the Antarctic Yeast, Glaciozyma antarctica PI12. J. Fungi 2021, 7, 528. [Google Scholar] [CrossRef]
- Min, W.; Guangfeng, K.; Cuijuan, S.; Qiuju, X.; Yingying, H.; Zhenhuan, L. In Heavy metal tolerance of an antarctic bacterial Strain O5 and its antioxidant enzyme activity changes induced by Cu2+. In Proceedings of the 2011 IEEE International Conference on Systems Biology (ISB), Zhuhai, China, 2–4 September 2011; pp. 303–306. [Google Scholar]
- Rajpert, L.; Sklodowska, A.; Matlakowska, R. Biotransformation of copper from Kupferschiefer black shale (Fore-Sudetic Monocline, Poland) by yeast Rhodotorula mucilaginosa LM9. Chemosphere 2013, 91, 1257–1265. [Google Scholar] [CrossRef]
- Ghaffari, S.; Sarp, A.S.K.; Lange, D.; Gulsoy, M. Potassium iodide potentiated photodynamic inactivation of Enterococcus faecalis using Toluidine Blue: Comparative analysis and post-treatment biofilm formation study. Photodiagn. Photodyn. Ther. 2018, 24, 245–249. [Google Scholar] [CrossRef]
- Zhang, H.B.; Wang, L.; Sha, T. Absorption of heavy metals by microorganisms isolated from lead-zinc tailing. J. Microbiol. 2004, 24, 34–37. [Google Scholar]
- Pinto, M.C.; Mata, A.M.; Lopez-Barea, J. Reversible inactivation of Saccharomyces cerevisiae glutathione reductase under reducing conditions. Arch. Biochem. Biophys. 1984, 228, 1–12. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through an increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.H.; Byun, J.R. Occurrence of glutathione sulphydryl (GSH) and antioxidant activities in probiotic Lactobacillus spp. Asian-Australas. J. Anim. Sci. 2004, 17, 1582–1585. [Google Scholar] [CrossRef]
- Freitas, C.; Nobre, B.; Gouveia, L.; Roseiro, J.; Reis, A.; da Silva, T.L. New at-line flow cytometric protocols for determining carotenoid content and cell viability during Rhodosporidium toruloides NCYC 921 batch growth. Process Biochem. 2014, 49, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Ojuederie, O.B.; Babalola, O.O. Microbial and Plant-Assisted Bioremediation of Heavy Metal Polluted Environments: A Review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [Green Version]
- Tashirev, A.B.; Rokitko, P.V.; Levishko, A.S.; Romanovskaia, V.A.; Tashireva, A.A. Resistance of chemoorganotrophic bacteria isolated from Antarctic cliffs to toxic metals. Mikrobiol. Z. 2012, 74, 3–7. [Google Scholar]
- Kan, G.; Wang, X.; Jiang, J.; Zhang, C.; Chi, M.; Ju, Y.; Shi, C. Copper stress response in yeast Rhodotorula mucilaginosa AN5 isolated from sea ice, Antarctic. Microbiologyopen 2019, 8, e00657. [Google Scholar] [CrossRef]
- Escamilla-Rodríguez, A.; Carlos-Hernández, S.; Díaz-Jiménez, L. Evidence of Resistance of Heavy Metals from Bacteria Isolated from Natural Waters of a Mining Area in Mexico. Water 2021, 13, 2766. [Google Scholar] [CrossRef]
- Dias, M.; Gomes de Lacerda, J.T.J.; Perdigao Cota de Almeida, S.; de Andrade, L.M.; Oller do Nascimento, C.A.; Rozas, E.E.; Mendes, M.A. Response mechanism of mine-isolated fungus Aspergillus niger IOC 4687 to copper stress determined by proteomics. Metallomics 2019, 11, 1558–1566. [Google Scholar] [CrossRef]
- Elahi, A.; Rehman, A.; Zajif Hussain, S.; Zulfiqar, S.; Shakoori, A.R. Isolation and characterization of a highly effective bacterium Bacillus cereus b-525k for hexavalent chromium detoxification. Saudi J. Biol. Sci. 2022, 29, 2878–2885. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Banthia, S.; Hazra, C.; Sen, R.; Das, S.; Das, K. Electrodeposited functionally graded coating inhibits Gram-positive and Gram-negative bacteria by a lipid peroxidation mediated membrane damage mechanism. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 102, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Bruno-Barcena, J.M.; Azcarate-Peril, M.A.; Hassan, H.M. Role of antioxidant enzymes in bacterial resistance to organic acids. Appl. Environ. Microbiol. 2010, 76, 2747–2753. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Zhou, X.; Zhang, J.; Wang, J.; Xie, H.; Wu, Z. alpha-Lipoic acid protects against the cytotoxicity and oxidative stress induced by cadmium in HepG2 cells through regeneration of glutathione by glutathione reductase via Nrf2/ARE signaling pathway. Environ. Toxicol. Pharmacol. 2016, 45, 274–281. [Google Scholar] [CrossRef]
- Maria, P.; Antonella, C.; Marianna, D.; Elisa, Z.; Mario, S.; Filomena, D.; Carmen, R.; Clara, U.; Emilio, D.; Marco, L.G.; et al. Heavy-metal resistant microorganisms in sediments from submarine canyons and the adjacent continental slope in the northeastern Ligurian margin (Western Mediterranean Sea). Prog. Oceanogr. 2018, 168, 155–168. [Google Scholar]
- Yan, X.; Zhao, X.-E.; Sun, J.; Zhu, S.; Lei, C.; Li, R.; Gong, P.; Ling, B.; Wang, R.; Wang, H. Probing glutathione reductase activity with graphene quantum dots and gold nanoparticles system. Sens. Actuators B Chem. 2018, 263, 27–35. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, A.; Visiedo, F.; Dominguez-Riscart, J.; Ruiz-Mateos, B.; Saez-Benito, A.; Lechuga-Sancho, A.M.; Mateos, R.M. Blunted Reducing Power Generation in Erythrocytes Contributes to Oxidative Stress in Prepubertal Obese Children with Insulin Resistance. Antioxidants 2021, 10, 244. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N.; Ahmad, P. The effect of NADPH oxidase inhibitor diphenyleneiodonium (DPI) and glutathione (GSH) on Isatis cappadocica, under Arsenic (As) toxicity. Int. J. Phytoremediat. 2021, 23, 945–957. [Google Scholar] [CrossRef]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free lactic acid production under acidic conditions by lactic acid bacteria strains: Challenges and future prospects. Appl. Microbiol. Biotechnol. 2018, 102, 5911–5924. [Google Scholar] [CrossRef]
- Zhai, Q.; Xiao, Y.; Narbad, A.; Chen, W. Comparative metabolomic analysis reveals global cadmium stress response of Lactobacillus plantarum strains. Metallomics 2018, 10, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Fotiadis, D.; Kanai, Y.; Palacin, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Aspects Med. 2013, 34, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Tao, H.; Purvis, J.E.; York, S.W.; Shanmugam, K.T.; Ingram, L.O. Gene array-based identification of changes that contribute to ethanol tolerance in ethanologenic Escherichia coli: Comparison of KO11 (parent) to LY01 (resistant mutant). Biotechnol. Prog. 2003, 19, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Hu, T.; Li, H.; Jin, G.; Zhang, Y. Metabonomic profiling in study hepatoprotective effect of polysaccharides from Flammulina velutipes on carbon tetrachloride-induced acute liver injury rats using GC-MS. Int. J. Biol. Macromol. 2018, 110, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Olin-Sandoval, V.; Yu, J.S.L.; Miller-Fleming, L.; Alam, M.T.; Kamrad, S.; Correia-Melo, C.; Haas, R.; Segal, J.; Pena Navarro, D.A.; Herrera-Dominguez, L.; et al. Lysine harvesting is an antioxidant strategy and triggers underground polyamine metabolism. Nature 2019, 572, 249–253. [Google Scholar] [CrossRef]
- Tian, B.; Qiao, Z.; Zhang, L.; Li, H.; Pei, Y. Hydrogen sulfide and proline cooperate to alleviate cadmium stress in foxtail millet seedlings. Plant Physiol. Biochem. 2016, 109, 293–299. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, S. Prescience of endogenous regulation in Arabidopsis thaliana by Pseudomonas putida MTCC 5279 under phosphate starved salinity stress condition. Sci. Rep. 2020, 10, 5855. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ni, S.; Cai, S.; Zhang, G. Comprehensive dissection of primary metabolites in response to diverse abiotic stress in barley at the seedling stage. Plant Physiol. Biochem. 2021, 161, 54–64. [Google Scholar] [CrossRef]
- Geng, P.; Qin, W.; Xu, G. Proline metabolism in cancer. Amino Acids 2021, 53, 1769–1777. [Google Scholar] [CrossRef]
- Shavalikohshori, O.; Zalaghi, R.; Sorkheh, K.; Enaytizamir, N. The expression of proline production/degradation genes under salinity and cadmium stresses in Triticum aestivum inoculated with Pseudomonas sp. Int. J. Environ. Sci. Technol. 2019, 17, 2233–2242. [Google Scholar] [CrossRef]
- Tyminski, M.; Ciacka, K.; Staszek, P.; Gniazdowska, A.; Krasuska, U. Toxicity of meta-Tyrosine. Plants 2021, 10, 2800. [Google Scholar] [CrossRef]
- Ipson, B.R.; Fisher, A.L. Roles of the tyrosine isomers meta-tyrosine and ortho-tyrosine in oxidative stress. Ageing Res. Rev. 2016, 27, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokvam, J.; Brenes-Arguedas, T.; Lee, J.S.; Coley, P.D.; Kursar, T.A. Allelochemic function for a primary metabolite: The case of l-tyrosine hyper-production in Inga umbellifera (Fabaceae). Am. J. Bot. 2006, 93, 1109–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. Evaluation of Daphnia magna metabolic responses to organic contaminant exposure with and without dissolved organic matter using 1H nuclear magnetic resonance (NMR)-based metabolomics. Ecotoxicol. Environ. Saf. 2018, 164, 189–200. [Google Scholar] [CrossRef]
- Li, W.H.; Shi, Y.C.; Chang, C.H.; Huang, C.W.; Hsiu-Chuan Liao, V. Selenite protects Caenorhabditis elegans from oxidative stress via DAF-16 and TRXR-1. Mol. Nutr. Food Res. 2014, 58, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Gao, Z.; Shi, T.Q.; Song, P.; Ren, L.J.; Huang, H.; Ji, X.J. Reactive Oxygen Species-Mediated Cellular Stress Response and Lipid Accumulation in Oleaginous Microorganisms: The State of the Art and Future Perspectives. Front. Microbiol. 2017, 8, 793. [Google Scholar] [CrossRef]
- Mirosława, S.; Przemysław, B.; Sylwia, R.; Justyna, N.; Jerzy, D. Comparative study of metal induced phospholipid modifications in the heavy metal tolerant filamentous fungus Paecilomyces marquandii and implications for the fungal membrane integrity. Acta Biochim. Pol. 2013, 60, 695–700. [Google Scholar]
- Baoukina, S.; Rozmanov, D.; Tieleman, D.P. Composition Fluctuations in Lipid Bilayers. Biophys. J. 2017, 113, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Mikami, K.; Murata, N. Membrane fluidity and the perception of environmental signals in cyanobacteria and plants. Prog. Lipid Res. 2003, 42, 527–543. [Google Scholar] [CrossRef]
- Ramos, J.L.; Duque, E.; Gallegos, M.T.; Godoy, P.; Ramos-Gonzalez, M.I.; Rojas, A.; Teran, W.; Segura, A. Mechanisms of solvent tolerance in gram-negative bacteria. Annu. Rev. Microbiol. 2002, 56, 743–768. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Inarrairaegui, A.; Ugalde, U. New insights into copper homeostasis in filamentous fungi. Int. Microbiol. 2020, 23, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Fu, L.; Hu, Z.; Li, Y.; Zheng, X.; Zhang, Z.; Jiang, C.; Zeng, B. Modulation of Fatty Acid Composition of Aspergillus oryzae in Response to Ethanol Stress. Microorganisms 2019, 7, 158. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(a) Growth curves of the Planococcus sp. O5 in the absence and presence of 0.5 mmol/L Cu2+. (b) Heavy metal tolerance of Planococcus sp. O5 of Cu2+, Zn2+, Pb2+, Cd2+, and Hg2+, separately. (c) Effect of 0.5 mmol/L Cu2+ on the conductivity of Planococcus sp. O5.
Figure 1.
(a) Growth curves of the Planococcus sp. O5 in the absence and presence of 0.5 mmol/L Cu2+. (b) Heavy metal tolerance of Planococcus sp. O5 of Cu2+, Zn2+, Pb2+, Cd2+, and Hg2+, separately. (c) Effect of 0.5 mmol/L Cu2+ on the conductivity of Planococcus sp. O5.

Figure 2.
Effects of 0.5 mmol/L Cu2+ on the activities of SOD (a), GR (b), APX (c), GSH (d), and Carotenoid (e) of Planococcus sp. O5.
Figure 2.
Effects of 0.5 mmol/L Cu2+ on the activities of SOD (a), GR (b), APX (c), GSH (d), and Carotenoid (e) of Planococcus sp. O5.

Figure 3.
(a) shows the PCA models for 24 samples; numbers 1,2 represent the control group and copper exposure in the logarithmic phase, and numbers 3,4 represent the control group and copper exposure in the stationary growth phase. (b,c) separation of control (red) and copper exposure (green) samples in logarithmic phase and stationary growth phase using OPLS-DA, respectively. (d,e) metabolites changes mapped to the metabolic pathways exposed to 0.5 mmol/L Cu2+ of Planococcus sp. O5 in the logarithmic and stationary growth phase.
Figure 3.
(a) shows the PCA models for 24 samples; numbers 1,2 represent the control group and copper exposure in the logarithmic phase, and numbers 3,4 represent the control group and copper exposure in the stationary growth phase. (b,c) separation of control (red) and copper exposure (green) samples in logarithmic phase and stationary growth phase using OPLS-DA, respectively. (d,e) metabolites changes mapped to the metabolic pathways exposed to 0.5 mmol/L Cu2+ of Planococcus sp. O5 in the logarithmic and stationary growth phase.

Figure 4.
Biochemical reveals Planococcus sp. O5 copper tolerance mechanisms. The down and up arrow means down-regulation and up-regulation, respectively.
Figure 4.
Biochemical reveals Planococcus sp. O5 copper tolerance mechanisms. The down and up arrow means down-regulation and up-regulation, respectively.

Figure 5.
Metabonomics reveals Planococcus sp. O5 copper tolerance mechanisms. The identified significantly different metabolites were integrated into pathways. The down and up arrow means down-regulation and up-regulation, respectively.
Figure 5.
Metabonomics reveals Planococcus sp. O5 copper tolerance mechanisms. The identified significantly different metabolites were integrated into pathways. The down and up arrow means down-regulation and up-regulation, respectively.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheng, Z.; Shi, C.; Gao, X.; Wang, X.; Kan, G. Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion. Toxics 2022, 10, 302. https://doi.org/10.3390/toxics10060302
AMA Style
Cheng Z, Shi C, Gao X, Wang X, Kan G. Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion. Toxics. 2022; 10(6):302. https://doi.org/10.3390/toxics10060302
Chicago/Turabian StyleCheng, Ziyi, Cuijuan Shi, Xiujun Gao, Xiaofei Wang, and Guangfeng Kan. 2022. "Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion" Toxics 10, no. 6: 302. https://doi.org/10.3390/toxics10060302
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.