
Transgenerational Effects of Prenatal Endocrine Disruption on Reproductive and Sociosexual Behaviors in Sprague Dawley Male and Female Rats

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
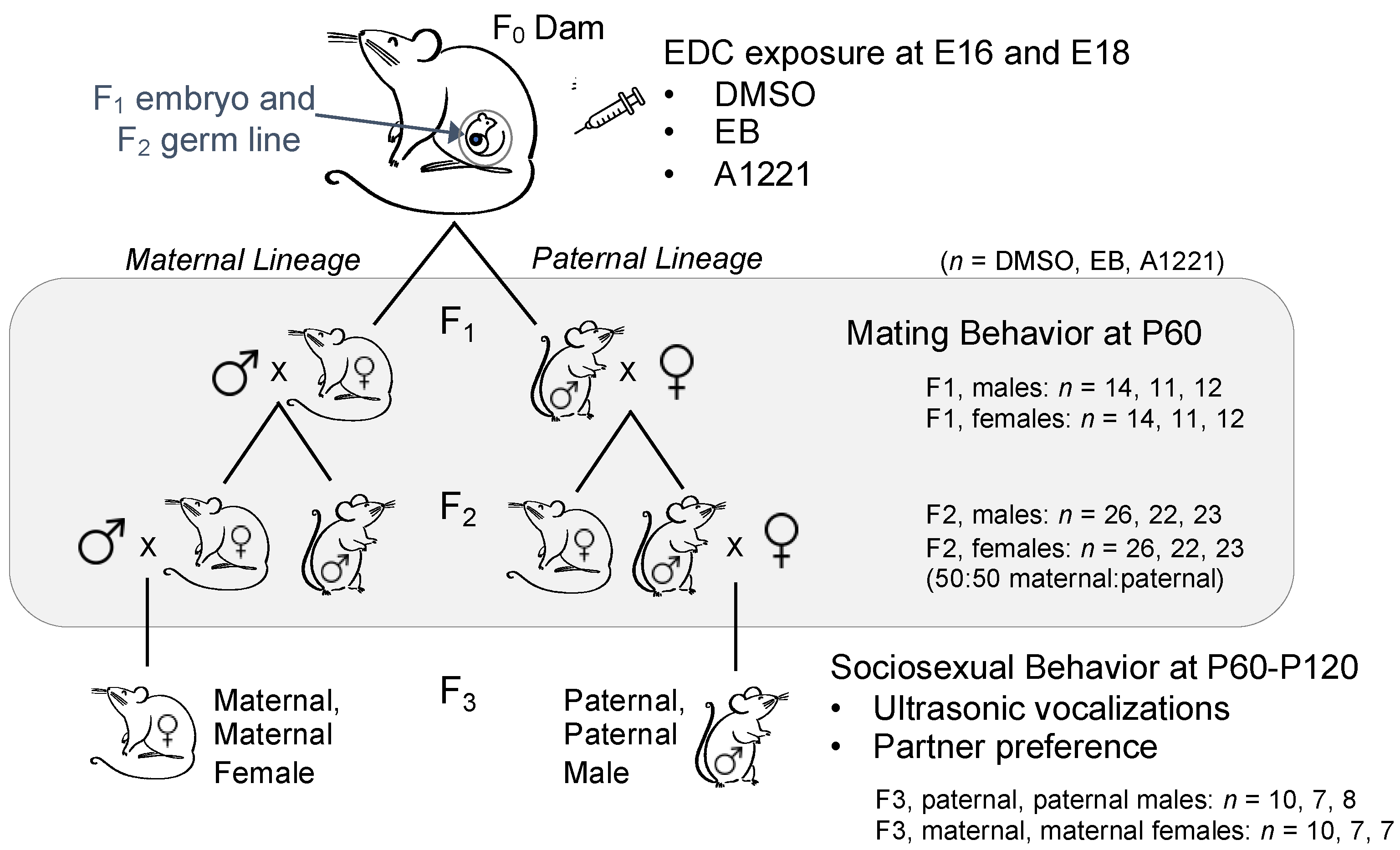
2.1. Experimental Design and Animal Husbandry
2.2. Tissue Collection
2.3. Serum Hormone Assays
2.4. Ovariectomy and Hormone Priming for Sociosexual Experiments
2.5. Reproductive Behavior and Fertility in F1 and F2 Rats
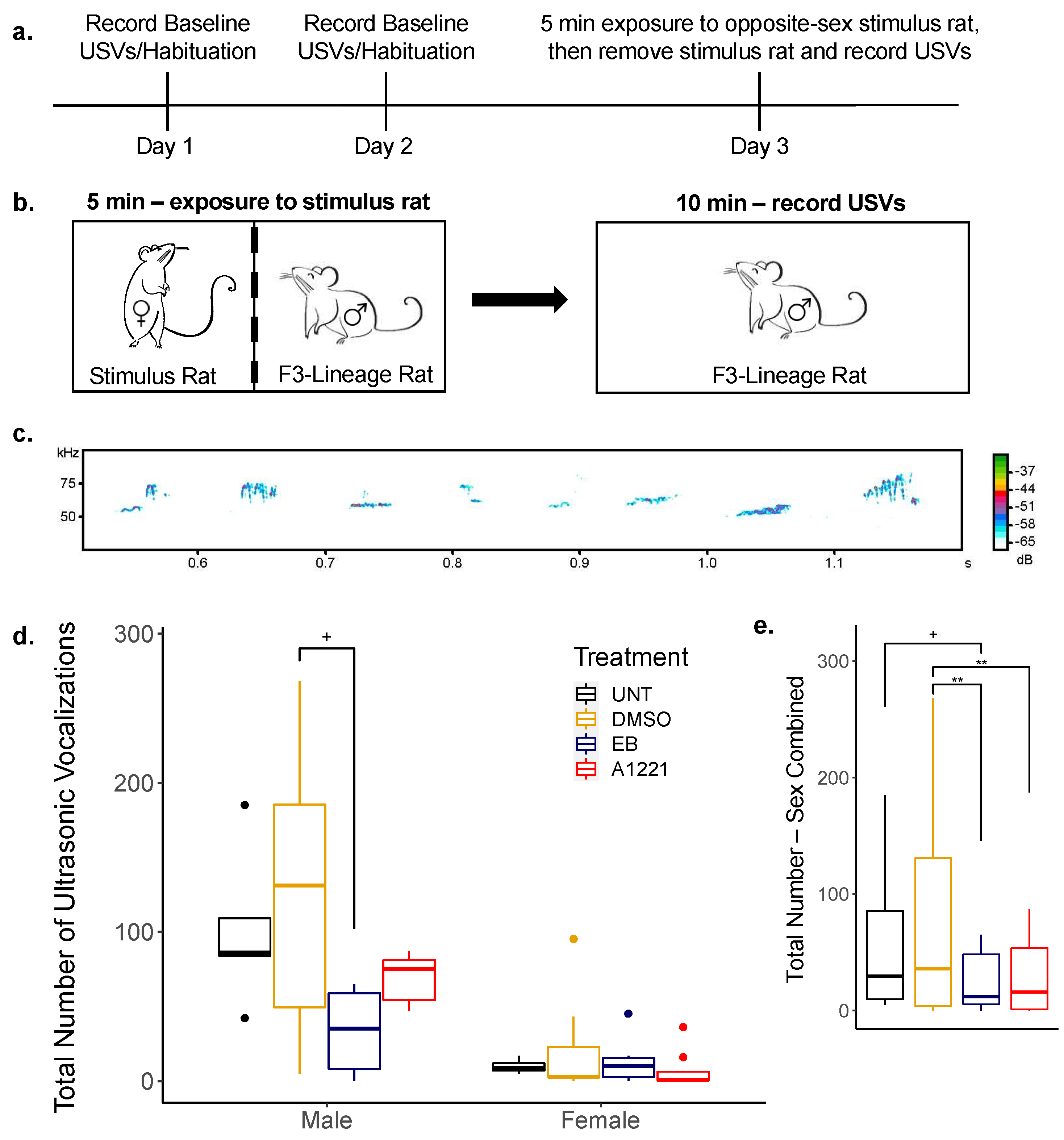
2.6. USV Recording in Sociosexual Context in F3 Rats
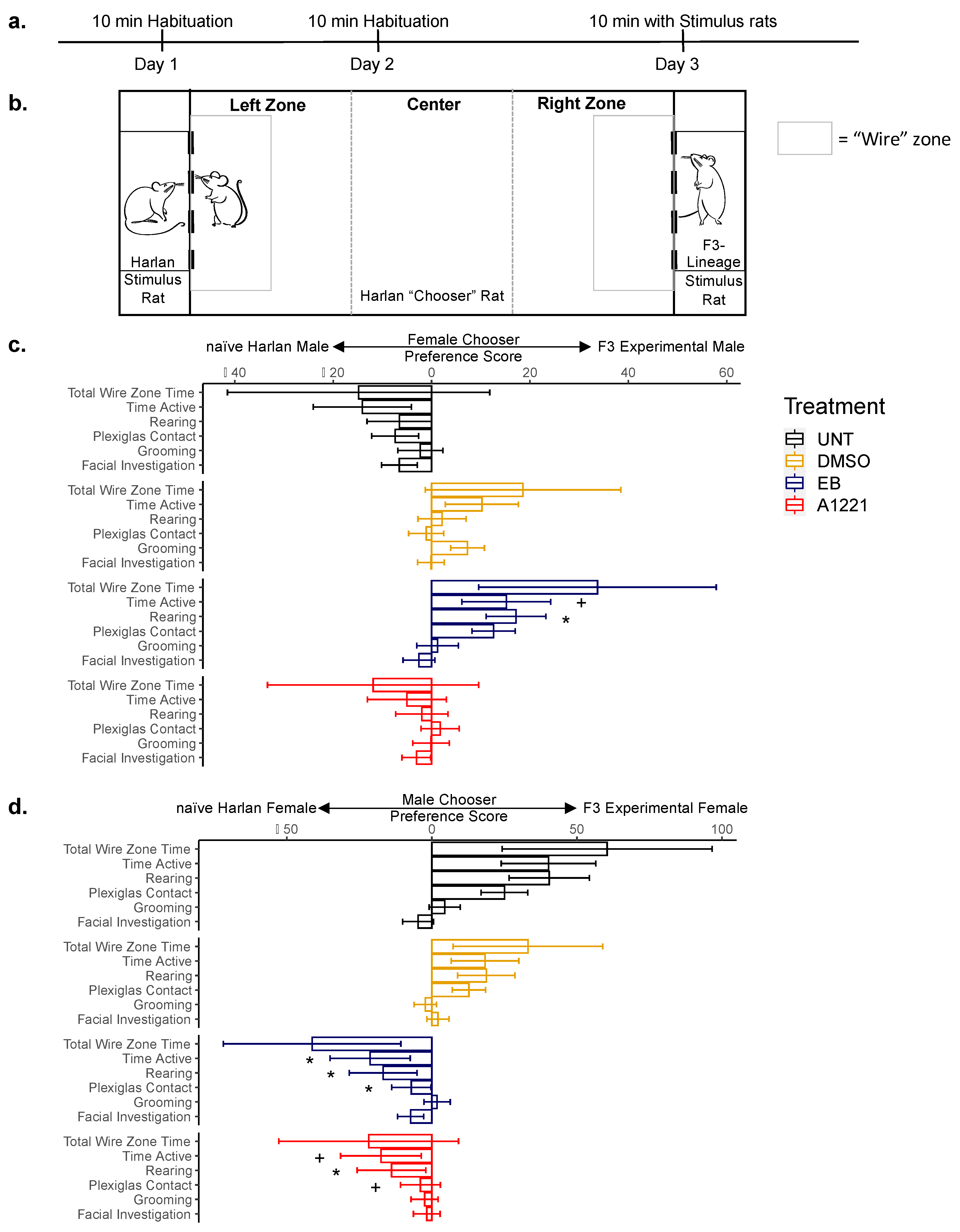
2.7. Partner Preference in F3 Rats
2.8. Statistical Analysis
3. Results
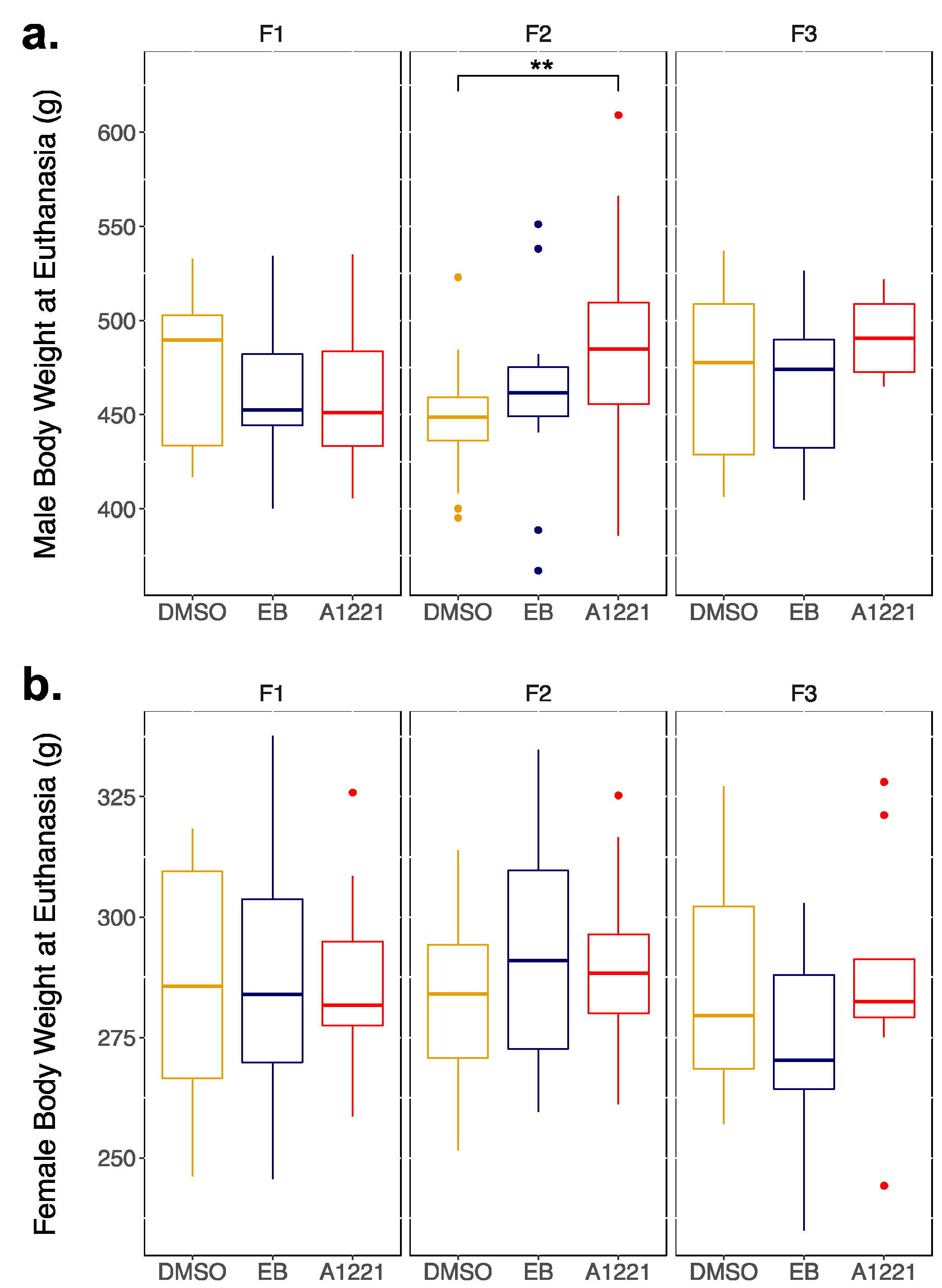
3.1. Transgenerational Somatic Changes
3.1.1. Males
3.1.2. Females
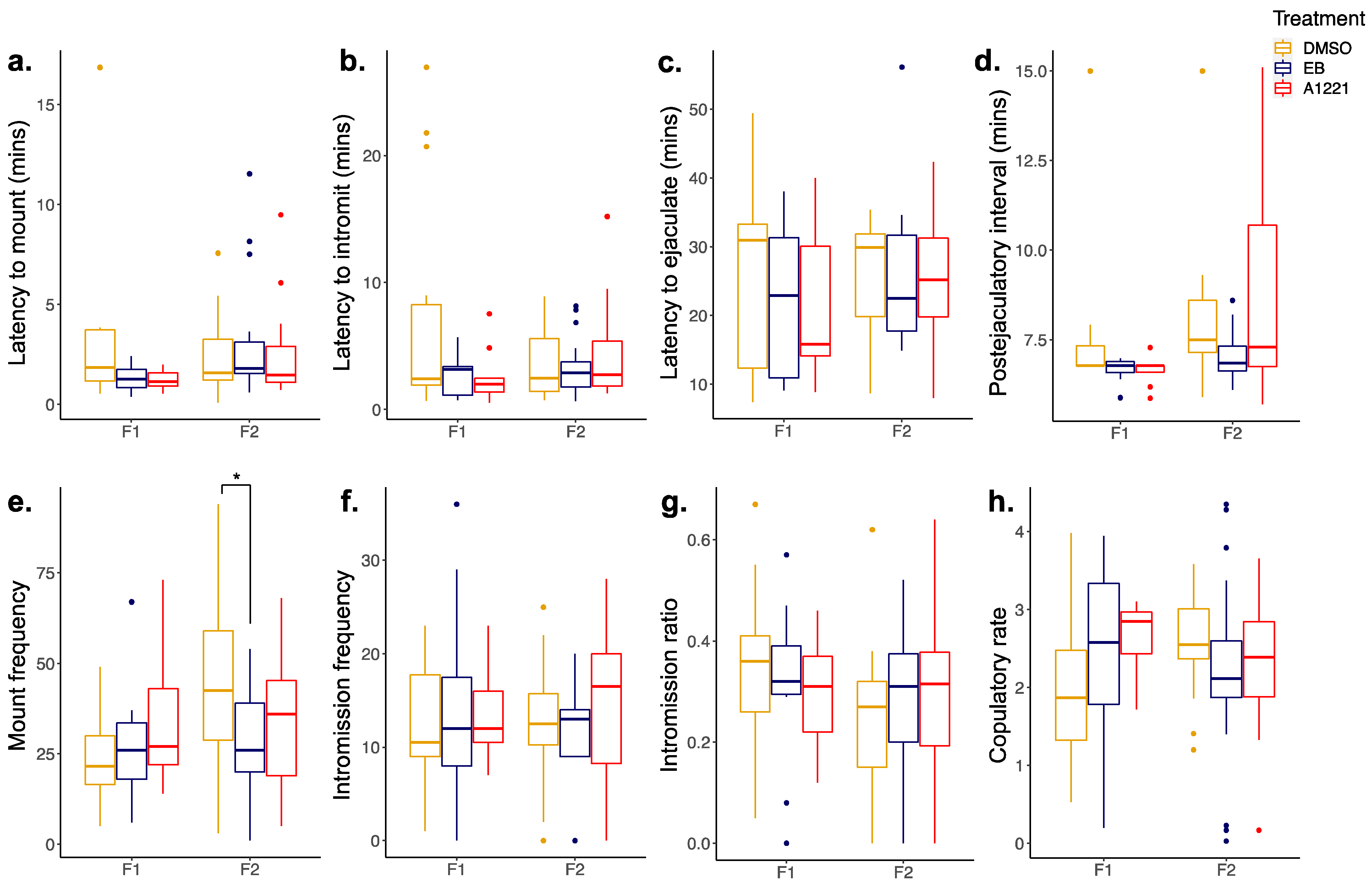
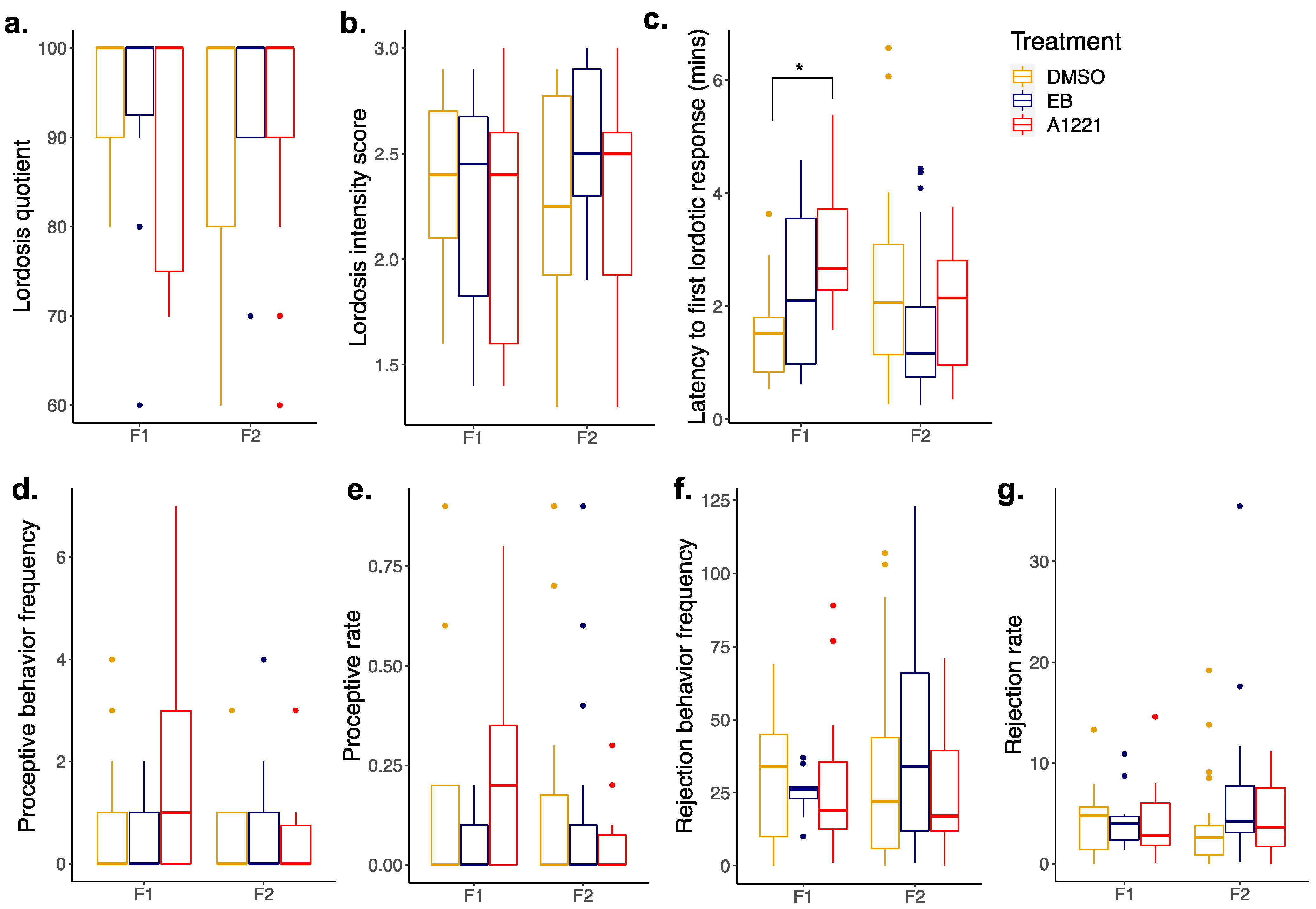
3.2. Reproductive Behavior in the F1 and F2 Generations
3.2.1. Males
3.2.2. Females
3.3. Sociosexual Behaviors in the F3 Generation
3.3.1. Ultrasonic Vocalizations (USVs)
3.3.2. Partner Preference
4. Discussion
4.1. Transgenerational Somatic Endpoints
4.2. F1 and F2 Generation Adult Mating Behavior
4.3. F3 Generation Adult Sociosexual Behaviors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoeller, R.T.; Brown, T.R.; Doan, L.L.; Gore, A.C.; Skakkebaek, N.E.; Soto, A.M.; Woodruff, T.J.; Vom Saal, F.S. Endocrine-Disrupting Chemicals and Public Health Protection: A Statement of Principles from The Endocrine Society. Endocrinology 2012, 153, 4097–4110. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. The developmental origins of adult disease. Eur. J. Epidemiol. 2003, 18, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.P.; Weeks, C.D.; Crews, D.; Gore, A.C. Application of a Novel Social Choice Paradigm to Assess Effects of Prenatal Endocrine-Disrupting Chemical Exposure in Rats (Rattus norvegicus). J. Comp. Psychol. 2018, 132, 253–267. [Google Scholar] [CrossRef]
- Reilly, M.P.; Weeks, C.D.; Topper, V.Y.; Thompson, L.M.; Crews, D.; Gore, A.C. The effects of prenatal PCBs on adult social behavior in rats. Horm. Behav. 2015, 73, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Topper, V.Y.; Reilly, M.P.; Wagner, L.M.; Thompson, L.M.; Gillette, R.; Crews, D.; Gore, A.C. Social and neuromolecular phenotypes are programmed by prenatal exposuRes to endocrine-disrupting chemicals. Mol. Cell Endocrinol. 2019, 479, 133–146. [Google Scholar] [CrossRef]
- Jones, B.A.; Watson, N.V. Perinatal BPA exposure demasculinizes males in measuRes of affect but has no effect on water maze learning in adulthood. Horm. Behav. 2012, 61, 605–610. [Google Scholar]
- Tian, Y.H.; Baek, J.H.; Lee, S.Y.; Jang, C.G. Prenatal and postnatal exposure to bisphenol a induces anxiolytic behaviors and cognitive deficits in mice. Synapse 2010, 64, 432–439. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S.; Belzunces, L.P.; Canivenc, M.C.; Schroeder, H.; Chevrier, C.; Pasquier, E. Impairment of learning and memory performances induced by BPA: Evidences from the literature of a MoA mediated through an ED. Mol. Cell Endocrinol. 2018, 475, 54–73. [Google Scholar] [CrossRef] [PubMed]
- Jasarevic, E.; Williams, S.A.; Vandas, G.M.; Ellersieck, M.R.; Liao, C.; Kannan, K.; Roberts, R.M.; Geary, D.C.; Rosenfeld, C.S. Sex and dose-dependent effects of developmental exposure to bisphenol A on anxiety and spatial learning in deer mice (Peromyscus maniculatus bairdii) offspring. Horm. Behav. 2013, 63, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, S.; Matsuzawa, D.; Ishii, D.; Tomizawa, H.; Sutoh, C.; Nakazawa, K.; Amano, K.; Sajiki, J.; Shimizu, E. Effects of perinatal exposure to low dose of bisphenol A on anxiety like behavior and dopamine metabolites in brain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 273–279. [Google Scholar] [CrossRef]
- Patisaul, H.B.; Bateman, H.L. Neonatal exposure to endocrine active compounds or an ERbeta agonist increases adult anxiety and aggression in gonadally intact male rats. Horm. Behav. 2008, 53, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hong, X.; Xie, L.; Li, T.; Yang, Y.; Zhang, Q.; Zhang, G.; Liu, X. Gestational and lactational exposure to bisphenol-A affects anxiety- and depression-like behaviors in mice. Horm. Behav. 2012, 62, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.R.; Thompson, L.M.; Rodriguez, K.; Gore, A.C. Two-hit exposure to polychlorinated biphenyls at gestational and juvenile life stages: 1. Sexually dimorphic effects on social and anxiety-like behaviors. Horm. Behav. 2016, 78, 168–177. [Google Scholar]
- Gillette, R.; Reilly, M.P.; Topper, V.Y.; Thompson, L.M.; Crews, D.; Gore, A.C. Anxiety-like behaviors in adulthood are altered in male but not female rats exposed to low dosages of polychlorinated biphenyls in utero. Horm. Behav. 2017, 87, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Scacchi, P.; Moguilevsky, J.A.; Cutrera, R. Antiandrogenic effect of perinatal exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate increases anxiety-like behavior in male rats during sexual maturation. Horm. Behav. 2013, 63, 692–699. [Google Scholar] [CrossRef]
- Kobrosly, R.W.; Evans, S.; Miodovnik, A.; Barrett, E.S.; Thurston, S.W.; Calafat, A.M.; Swan, S.H. Prenatal phthalate exposuRes and neurobehavioral development scoRes in boys and girls at 6–10 years of age. Environ. Health Perspect. 2014, 122, 521–528. [Google Scholar]
- England-Mason, G.; Martin, J.W.; MacDonald, A.; Kinniburgh, D.; Giesbrecht, G.F.; Letourneau, N.; Dewey, D. Similar names, different results: Consistency of the associations between prenatal exposure to phthalates and parent-ratings of behavior problems in preschool children. Environ. Int. 2020, 142, 105892. [Google Scholar] [CrossRef]
- Yoo, S.J.; Joo, H.; Kim, D.; Lim, M.H.; Kim, E.; Ha, M.; Kwon, H.J.; Paik, K.C.; Kim, K.M. Associations between Exposure to Bisphenol A and Behavioral and Cognitive Function in Children with Attention-deficit/Hyperactivity Disorder: A Case-control Study. Clin. Psychopharmacol. Neurosci. 2020, 18, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, F.; Nolte, E.L.R.; Wang, Y.; Margolis, A.E.; Calafat, A.M.; Wang, S.; Garcia, W.; Hoepner, L.A.; Peterson, B.S.; Rauh, V.; et al. Bisphenol A exposure and symptoms of anxiety and depression among inner city children at 10–12 years of age. Environ. Res. 2016, 151, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Vermeir, G.; Covaci, A.; Van Larebeke, N.; Schoeters, G.; Nelen, V.; Koppen, G.; Viaene, M. Neurobehavioural and cognitive effects of prenatal exposure to organochlorine compounds in three year old children. BMC Pediatr. 2021, 21, 99. [Google Scholar] [CrossRef] [PubMed]
- Anway, M.D.; Cupp, A.S.; Uzumcu, M.; Skinner, M.K. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005, 308, 1466–1469. [Google Scholar] [CrossRef] [Green Version]
- Gillette, R.; Son, M.J.; Ton, L.; Gore, A.C.; Crews, D. Passing experiences on to future generations: Endocrine disruptors and transgenerational inheritance of epimutations in brain and sperm. Epigenetics 2018, 13, 1106–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesan, D.; Feighan, K.M.; Antle, M.C.; Kurrasch, D.M. Gestational low-dose BPA exposure impacts suprachiasmatic nucleus neurogenesis and circadian activity with transgenerational effects. Sci. Adv. 2021, 7, eabd1159. [Google Scholar] [CrossRef]
- Quinnies, K.M.; Doyle, T.J.; Kim, K.H.; Rissman, E.F. Transgenerational Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Stress Hormones and Behavior. Endocrinology 2015, 156, 3077–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.T.; Edwards, M.; Shetty, S.R.; Gatewood, J.D.; Taylor, J.A.; Rissman, E.F.; Connelly, J.J. Gestational exposure to bisphenol A produces transgenerational changes in behaviors and gene expression. Endocrinology 2012, 153, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, J.T.; Goldsby, J.A.; Rissman, E.F. Transgenerational effects of prenatal bisphenol A on social recognition. Horm. Behav. 2013, 64, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, K.; Mittal, N.; Thompson Lindsay, M.; Rodriguez-Santiago, M.; Duvauchelle Christine, L.; Crews, D.; Gore Andrea, C. Effects of the endocrine-disrupting chemicals, vinclozolin and polychlorinated biphenyls, on physiological and sociosexual phenotypes in F2 generation Sprague-Dawley rats. Environ. Health Perspect. 2018, 126, 097005. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Rahman, S.; Hasbum, A.; Morales, D.; Thompson, L.M.; Crews, D.; Gore, A.C. Maternal care modulates transgenerational effects of endocrine-disrupting chemicals on offspring pup vocalizations and adult behaviors. Horm. Behav. 2019, 107, 96–109. [Google Scholar] [CrossRef]
- Crews, D.; Gillette, R.; Scarpino, S.V.; Manikkam, M.; Savenkova, M.I.; Skinner, M.K. Epigenetic transgenerational inheritance of altered stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 9143–9148. [Google Scholar] [CrossRef] [Green Version]
- Crews, D.; Gore, A.C.; Hsu, T.S.; Dangleben, N.L.; Spinetta, M.; Schallert, T.; Anway, M.D.; Skinner, M.K. Transgenerational epigenetic imprints on mate preference. Proc. Natl. Acad. Sci. USA 2007, 104, 5942–5946. [Google Scholar] [CrossRef] [Green Version]
- Robaire, B.; Delbes, G.; Head, J.A.; Marlatt, V.L.; Martyniuk, C.J.; Reynaud, S.; Trudeau, V.L.; Mennigen, J.A. A cross-species comparative approach to assessing multi- and transgenerational effects of endocrine disrupting chemicals. Environ. Res. 2021, 204, 112063. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Hasbum, A.; Morales, D.; Thompson, L.M.; Crews, D.; Gore, A.C. Endocrine-disrupting chemicals alter the neuromolecular phenotype in F2 generation adult male rats. Physiol. Behav. 2019, 211, 112674. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Thompson, L.M.; Bell, M.; Mennigen, J.A. Transgenerational effects of polychlorinated biphenyls: 2. Hypothalamic gene expression in rats †. Biol. Reprod. 2021, 105, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Mennigen, J.A.; Thompson, L.M.; Bell, M.; Tellez Santos, M.; Gore, A.C. Transgenerational effects of polychlorinated biphenyls: 1. Development and physiology across 3 generations of rats. Environ. Health 2018, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, R.M.; Juenger, T.E.; Gore, A.C. The effects of prenatal PCBs on adult female paced mating reproductive behaviors in rats. Horm. Behav. 2007, 51, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, S.M.; Cunningham, S.L.; Patisaul, H.B.; Woller, M.J.; Gore, A.C. Endocrine disruption of brain sexual differentiation by developmental PCB exposure. Endocrinology 2011, 152, 581–594. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, E.F.; Belanger, E.E.; Gomez, M.I.; Cayo, M.; McCaffrey, R.J.; Seegal, R.F.; Jansing, R.L.; Hwang, S.A. Polychlorinated biphenyl exposure and neuropsychological status among older residents of upper Hudson River communities. Environ. Health Perspect. 2008, 116, 209–215. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, M.Y.; Vakulenko, M. Characterization of 50-kHz ultrasonic vocalizations in male and female rats. Physiol. Behav. 2003, 80, 81–88. [Google Scholar] [CrossRef]
- Simola, N.; Ma, S.T.; Schallert, T. Influence of acute caffeine on 50-kHz ultrasonic vocalizations in male adult rats and relevance to caffeine-mediated psychopharmacological effects. Int. J. Neuropsychopharmacol. 2010, 13, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liang, S.; Burgdorf, J.; Wess, J.; Yeomans, J. Ultrasonic vocalizations induced by sex and amphetamine in M2, M4, M5 muscarinic and D2 dopamine receptor knockout mice. PLoS ONE 2008, 3, e1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kassambara, R. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 13 November 2021).
- Lenth, R. Emmeans: Estimated Marginal Means, AKA least-Square Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 13 November 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S.K. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the ACM Symposium on User Interface Software and Technology (UIST 2021), Virtual Event, 10–14 October 2021; ACS Press: New York, NY, USA, 2021; pp. 754–768. [Google Scholar]
- Kay, M.; Elkin, L.; Higgins, J.J.; Wobbrock, J.O. ARTool: Aligned Rank Transform for Nonparametric Factorial ANOVAs. R package version 0.11.1. Available online: https://github.com/mjskay/ARTool (accessed on 13 November 2021). [CrossRef]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only Anova Procedures. In Proceedings of the ACM Conference on Human Factors in Computing Systems (CHI 2011), Vancouver, BC, USA, 7–12 May 2011; ACS Press: New York, NY, USA, 2021; pp. 143–146. [Google Scholar]
- Streifer, M.; Gore, A.C. Epigenetics, estrogenic endocrine-disrupting chemicals (EDCs), and the brain. Adv. Pharmacol. 2021, 92, 73–99. [Google Scholar] [PubMed]
- Walker, D.M.; Goetz, B.M.; Gore, A.C. Dynamic postnatal developmental and sex-specific neuroendocrine effects of prenatal polychlorinated biphenyls in rats. Mol. Endocrinol. 2014, 28, 99–115. [Google Scholar] [CrossRef]
- Kilic, N.; Sandal, S.; Colakoglu, N.; Kutlu, S.; Sevran, A.; Yilmaz, B. Endocrine disruptive effects of polychlorinated biphenyls on the thyroid gland in female rats. Tohoku J. Exp. Med. 2005, 206, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, A.J.; Cooke, G.M. Suppression of aromatase activity in vitro by PCBs 28 and 105 and Aroclor 1221. Toxicol. Lett. 2004, 152, 91–100. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, S.M.; Gore, A.C. Estrogenic environmental endocrine-disrupting chemical effects on reproductive neuroendocrine function and dysfunction across the life cycle. Rev. Endocr. Metab. Disord. 2007, 8, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Colciago, A.; Casati, L.; Mornati, O.; Vergoni, A.V.; Santagostino, A.; Celotti, F.; Negri-Cesi, P. Chronic treatment with polychlorinated biphenyls (PCB) during pregnancy and lactation in the rat. Part 2: Effects on reproductive parameters, on sex behavior, on memory retention and on hypothalamic expression of aromatase and 5alpha-reductases in the offspring. Toxicol. Appl. Pharmacol. 2009, 239, 46–54. [Google Scholar]
- Faass, O.; Schlumpf, M.; Reolon, S.; Henseler, M.; Maerkel, K.; Durrrer, S.; Lichtensteiger, W. Female sexual behavior, estrous cycle and gene expression in sexually dimorphic brain regions after pre- and postnatal exposure to endocrine active UV filters. Neurotoxicology 2009, 30, 249–260. [Google Scholar] [CrossRef]
- Henley, C.L.; Nunez, A.A.; Clemens, L.G. Estrogen treatment during development alters adult partner preference and reproductive behavior in female laboratory rats. Horm. Behav. 2009, 55, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.A.; Clemens, L.G.; Nunez, A.A. Exposure to PCB 77 affects partner preference but not sexual behavior in the female rat. Physiol. Behav. 2008, 95, 471–475. [Google Scholar] [CrossRef]
- Hull, E.M.; Meisel, R.L.; Sachs, B.D. Male Sexual Behavior. In Hormones, Brain and Behavior; Elsevier: Amsterdam, The Netherlands, 2002; pp. 139–214. [Google Scholar]
- Portfors, C.V. Types and functions of ultrasonic vocalizations in laboratory rats and mice. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 28–34. [Google Scholar] [PubMed]
- Barfield, R.J.; Thomas, D.A. The role of ultrasonic vocalizations in the regulation of reproduction in rats. Ann. N. Y. Acad. Sci. 1986, 474, 33–43. [Google Scholar] [CrossRef] [PubMed]
- White, N.R.; Barfield, R.J. Playback of female rat ultrasonic vocalizations during sexual behavior. Physiol. Behav. 1989, 45, 229–233. [Google Scholar] [CrossRef]
- Ahrens, A.M.; Ma, S.T.; Maier, E.Y.; Duvauchelle, C.L.; Schallert, T. Repeated intravenous amphetamine exposure: Rapid and persistent sensitization of 50-kHz ultrasonic trill calls in rats. Behav. Brain Res. 2009, 197, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Wöhr, M.; Houx, B.; Schwarting, R.K.W.; Spruijt, B. Effects of experience and context on 50-kHz vocalizations in rats. Physiol. Behav. 2008, 93, 766–776. [Google Scholar] [CrossRef]
- Kisko, T.M.; Schwarting, R.K.W.; Wöhr, M. Sex differences in the acoustic featuRes. of social play-induced 50-kHz ultrasonic vocalizations: A detailed spectrographic analysis in wild-type Sprague–Dawley and Cacna1c haploinsufficient rats. Dev. Psychobiol. 2021, 63, 262–276. [Google Scholar] [CrossRef]
- McGinnis, M.Y.; Kahn, D.F. Inhibition of male sexual behavior by intracranial implants of the protein synthesis inhibitor anisomycin into the medial preoptic area of the rat. Horm. Behav. 1997, 31, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Bogacki-Rychlik, W.; Rolf, M.; Bialy, M. Anticipatory 50-kHz Precontact Ultrasonic Vocalizations and Sexual Motivation: Characteristic Pattern of Ultrasound Subtypes in an Individual Analyzed Profile. Front. Behav. Neurosci. 2021, 15, 722456. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, S.M. Biological Functions of Rat Ultrasonic Vocalizations, Arousal Mechanisms, and Call Initiation. Brain Sci. 2021, 11, 605. [Google Scholar] [CrossRef] [PubMed]
- Matochik, J.A.; White, N.R.; Barfield, R.J. Variations in scent marking and ultrasonic vocalizations by Long-Evans rats across the estrous cycle. Physiol. Behav. 1992, 51, 783–786. [Google Scholar] [CrossRef]
- Sobolewski, M.; Abston, K.; Conrad, K.; Marvin, E.; Harvey, K.; Susiarjo, M.; Cory-Slechta, D.A. Lineage- and Sex-Dependent Behavioral and Biochemical Transgenerational Consequences of Developmental Exposure to Lead, Prenatal Stress, and Combined Lead and Prenatal Stress in Mice. Environ. Health Perspect. 2020, 128, 27001. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Females | Males | |||
|---|---|---|---|---|
| EB | A1221 | EB | A1221 | |
| Somatic (F1, F2, F3) | ||||
| Body weight | - | - | - |  (F2) (F2) |
| Adrenal weight (normalized) | - | - |  (trend, F1 and F3) (trend, F1 and F3) | - |
| Uterine weight (normalized) | - | A1221 > EB (F2) | - | - |
| Hormones (F1, F2)—Estradiol (females), Testosterone (males)—n.s. | ||||
| Mating Behaviors (F1, F2) | ||||
| Mount frequency |  (F2) (F2) | - | - | - |
| Lordosis latency | - |  (F1) (F1) | - | - |
| Partner Preference Behavior (F3) | ||||
| Total time active |  |  (trend) (trend) | - | - |
| Time rearing |  |  |  (trend) (trend) | - |
| # Rearing bouts |  (trend) (trend) | - | - | - |
| Time at Plexiglas |  |  (trend) (trend) |  | - |
| # Plexiglas bouts |  | - | - | - |
| Ultrasonic Vocalizations (F3) | ||||
| (combined for the sexes) | EB | A1221 | ||
| # Total calls |  |  | ||
| # Non-FM calls |  | |||
 Decreased compared to DMSO (or UNT for Partner Preference).
Decreased compared to DMSO (or UNT for Partner Preference).  Increased compared to DMSO (or UNT for Partner Preference). FM: Frequency modulated. n.s. and -: No significant effects. #: Number.
Increased compared to DMSO (or UNT for Partner Preference). FM: Frequency modulated. n.s. and -: No significant effects. #: Number.| F1 MALES | DMSO (n = 14) | EB (n = 11) | A1221 (n = 12) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | p-Values | |
| Body Weight (g) | 475.2 | (±10.9) | 462.9 | (±12.0) | 461.7 | (±11.6) | n.s |
| Norm Adrenal Weight (mg) | 0.105 | (±2.2 × 10−3) | 0.113 | (±2.6 × 10−3) | 0.107 | (±2.8 × 10−3) | p = 0.064 |
| Norm Testes Weight (mg) | 8.9 | (±0.24) | 9.0 | (±0.07) | 9.3 | (±0.25) | n.s. |
| Serum Testosterone (ng/mL) | 1.4 | (±0.2) | 1.2 | (±0.1) | 1.1 | (±0.2) | n.s. |
| F2 MALES | DMSO (n = 26) | EB (n = 22) | A1221 (n = 23) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Body Weight (g) | 449.3 | (±5.5) | 461.5 | (±8.2) | 482.2 | (±11.4) | p = 0.011 |
| Norm Adrenal Weight (mg) | 0.108 | (±1.8 × 10−3) | 0.111 | (±2.3 × 10−3) | 0.109 | (±2.7 × 10−3) | n.s. |
| Norm Testes Weight (mg) | 9.3 | (±0.009) | 9.2 | (±0.16) | 9.0 | (±0.18) | n.s. |
| Serum Testosterone (ng/mL) | 1.1 | (±0.1) | 1.2 | (±0.2) | 0.8 | (±0.1) | n.s. |
| F3 MALES | DMSO (n = 10) | EB (n = 9) | A1221 (n = 9) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Body Weight (g) | 472.4 | (±15.2) | 464.0 | (±12.9) | 491.3 | (±7.0) | n.s. |
| Norm Adrenal Weight (mg) | 0.108 | (±4.7 × 10−3) | 0.114 | (±3.5 × 10−3) | 0.100 | (±2.7 × 10−3) | p = 0.076 |
| Norm Testes Weight (mg) | 9.1 | (±0.27) | 9.4 | (±0.23) | 8.8 | (±0.17) | n.s. |
| F1 FEMALES | DMSO (n = 14) | EB (n = 11) | A1221 (n = 12) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Body Weight (g) | 286.2 | (±6.3) | 290.6 | (±8.7) | 286.0 | (±5.4) | n.s. |
| Norm Adrenal Weight (mg) | 0.219 | (±9.5 × 10−3) | 0.205 | (±4.3 × 10−3) | 0.208 | (±5.7 × 10−3) | n.s. |
| Norm Ovarian Weight (mg) | 0.540 | (±2.3 × 10−2) | 0.528 | (±1.9 × 10−2) | 0.538 | (±1.7 × 10−2) | n.s. |
| Norm Uterine Weight (mg) | 1.47 | (±0.1) | 1.99 | (±0.4) | 1.71 | (±0.2) | n.s. |
| Serum Estradiol (pg/mL) | 22.3 | (±2.9) | 21.8 | (±5.4) | 19.0 | (±2.3) | n.s. |
| F2 FEMALES | DMSO (n = 26) | EB (n = 22) | A1221 (n = 23) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Body Weight (g) | 283.0 | (±3.4) | 291.4 | (±4.6) | 289.6 | (±3.7) | n.s. |
| Norm Adrenal Weight (mg) | 0.208 | (±3.9 × 10−3) | 0.197 | (±4.9 × 10−3) | 0.203 | (±4.0 × 10−3) | n.s. |
| Norm Ovarian Weight (mg) | 0.540 | (±1.5 × 10−2) | 0.555 | (±1.7 × 10−2) | 0.547 | (±1.3 × 10−2) | n.s. |
| Norm Uterine Weight (mg) | 1.63 | (±0.1) | 1.45 | (±0.1) | 1.97 | (±0.2) * | p = 0.04 |
| Serum Estradiol (pg/mL) | 15.4 | (±1.6) | 14.4 | (±1.3) | 22.2 | (±3.4) | n.s. |
| F3 FEMALES | DMSO (n = 11) | EB (n = 7) | A1221 (n = 9) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Body Weight (g) | 285.8 | (±7.1) | 273.3 | (±8.4) | 287.4 | (±8.3) | n.s. |
| Norm Adrenal Weight (mg) | 0.202 | (±6.1 × 10−3) | 0.208 | (±8.3 × 10−3) | 0.207 | (±7.4 × 10−3) | n.s. |
| Norm Ovarian Weight (mg) | 0.437 | (±4.4 × 10−2) | 0.447 | (±1.8 × 10−2) | 0.475 | (±3.1 × 10−2) | n.s. |
| Norm Uterine Weight (mg) | 2.97 | (±0.5) | 2.33 | (±0.3) | 4.49 | (±1.2) | n.s. |
| MALES | UNT (n = 5) | DMSO (n = 10) | EB (n = 6) | A1221 (n = 6) | p-Values | (Sex-Combined) | ||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | Treatment | Sex | |
| Number of total calls | 101.2 | (±23.6) | 124.5 | (±28.3) | 33.5 | (±12.3) | 68.8 | (±7.8) | p < 0.001 | p < 0.001 |
| Number of non-FM calls | 48.2 | (±10.5) | 62.8 | (±13.9) | 22.8 | (±7.5) | 36.5 | (±10.1) | p = 0.024 | p < 0.001 |
| Number of FM calls | 53.0 | (±14.5) | 47.2 | (±11.5) | 17.4 | (±6.2) | 49.5 | (±9.4) | n.s. | p < 0.001 |
| Percentage of FM calls | 50.3 | (±4.8) | 47.3 | (±3.8) | 46.2 | (±7.1) | 59.7 | (±4.8) | n.s. | n.s. |
| Average call duration (ms) | 1.44 | (±0.22) | 1.41 | (±0.13) | 1.13 | (±0.13) | 1.61 | (±0.19) | n.s. | p = 0.032 |
| FEMALES | UNT (n = 5) | DMSO (n = 9) | EB (n = 6) | A1221 (n = 8) | ||||||
| Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | |||
| Number of total calls | 10.0 | (±2.1) | 21.4 | (±10.4) | 13.8 | (±6.7) | 7.3 | (±4.5) | ||
| Number of non-FM calls | 5.8 | (±1.5) | 10.7 | (±4.0) | 8.8 | (±4.1) | 9.6 | (±5.0) | ||
| Number of FM calls | 4.2 | (±0.7) | 26.5 | (±12.3) | 7.8 | (±3.6) | 4.3 | (±2.8) | ||
| Percentage of FM calls | 45.0 | (±5.7) | 40.5 | (±10.0) | 55.4 | (±12.4) | 32.0 | (±11.8) | ||
| Average call duration (ms) | 0.80 | (±0.10) | 1.06 | (±0.22) | 1.19 | (±0.20) | 1.28 | (±0.26) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kermath, B.A.; Thompson, L.M.; Jefferson, J.R.; Ward, M.H.B.; Gore, A.C. Transgenerational Effects of Prenatal Endocrine Disruption on Reproductive and Sociosexual Behaviors in Sprague Dawley Male and Female Rats. Toxics 2022, 10, 47. https://doi.org/10.3390/toxics10020047
Kermath BA, Thompson LM, Jefferson JR, Ward MHB, Gore AC. Transgenerational Effects of Prenatal Endocrine Disruption on Reproductive and Sociosexual Behaviors in Sprague Dawley Male and Female Rats. Toxics. 2022; 10(2):47. https://doi.org/10.3390/toxics10020047
Chicago/Turabian StyleKermath, Bailey A., Lindsay M. Thompson, Justin R. Jefferson, Mary H. B. Ward, and Andrea C. Gore. 2022. "Transgenerational Effects of Prenatal Endocrine Disruption on Reproductive and Sociosexual Behaviors in Sprague Dawley Male and Female Rats" Toxics 10, no. 2: 47. https://doi.org/10.3390/toxics10020047