
Asymmetric Pattern of Correlations of Leucine Aminopeptidase Activity between Left or Right Frontal Cortex versus Diverse Left or Right Regions of Rat Brains
 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
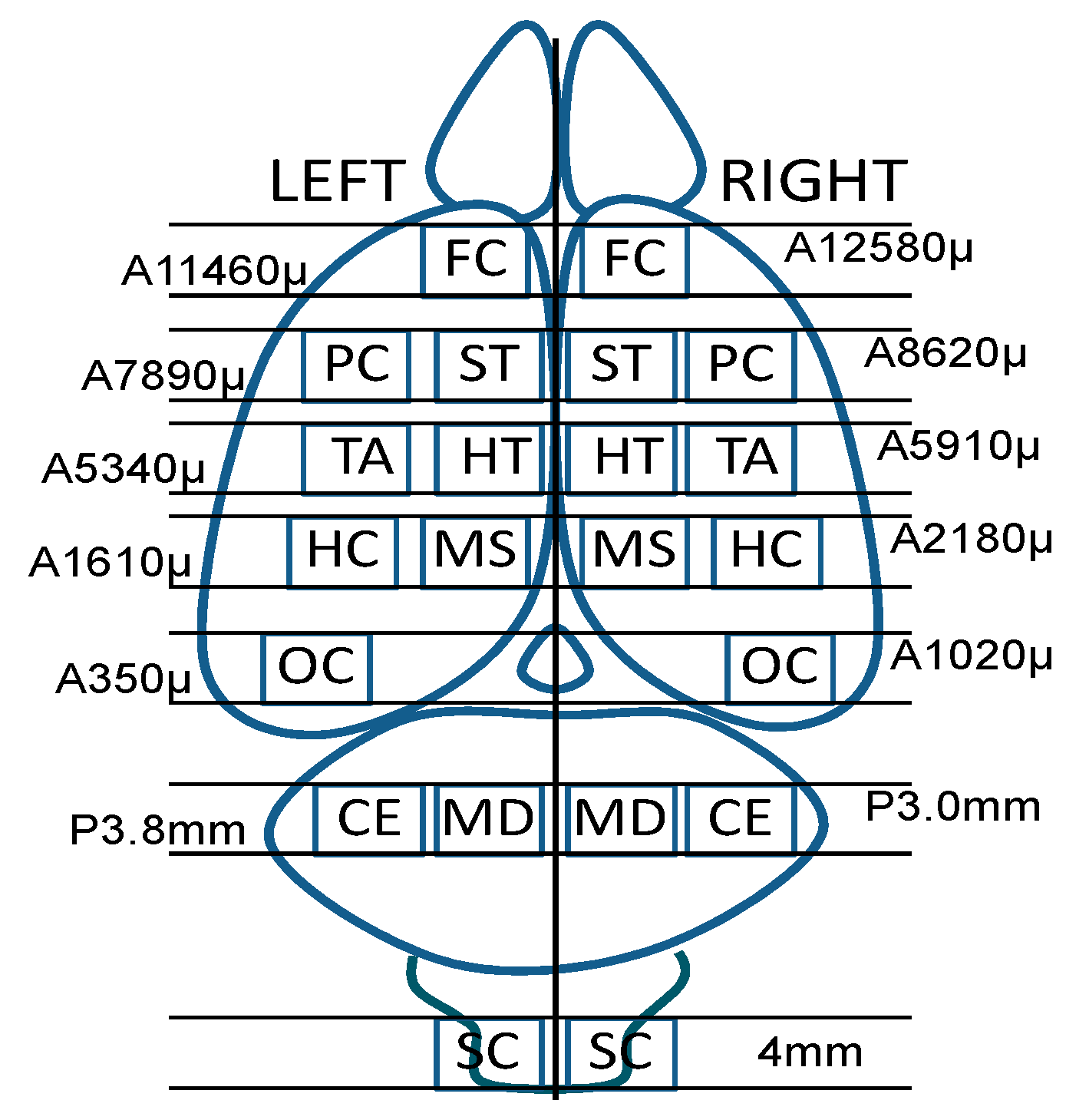
2.1. Animal Design, Collection and Sample Preparation
2.2. Determination of LeuAP Activity
2.3. Determination of LDH Activity
2.4. Statistical Analysis


3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ramírez-Sánchez, M.; Prieto, I.; Segarra, A.B.; Banegas, I.; Martínez-Cañamero, M.; Domínguez-Vías, G.; de Gasparo, M. Brain Asymmetry: Towards an Asymmetrical Neurovisceral Integration. Symmetry 2021, 13, 2409. [Google Scholar] [CrossRef]
- Corballis, M.C. Bilaterally Symmetrical: To Be or Not to Be? Symmetry 2020, 12, 326. [Google Scholar] [CrossRef] [Green Version]
- Corballis, M.C. How Asymmetries Evolved: Hearts, Brains, and Molecules. Symmetry 2021, 13, 914. [Google Scholar] [CrossRef]
- Homberg, J.R.; Adan, R.A.H.; Alenina, N.; Asiminas, A.; Bader, M.; Beckers, T.; Begg, D.P.; Blokland, A.; Burger, M.E.; van Dijk, G.; et al. The continued need for animals to advance brain research. Neuron 2021, 109, 2374–2379. [Google Scholar] [CrossRef]
- Alba, F.; Ramirez, M.; Cantalejo, E.S.; Iribar, C. Aminopeptidase activity is asymmetrically distributed in selected zones of rat brain. Life Sci. 1988, 43, 935–939. [Google Scholar] [CrossRef]
- Alba, F.; Iríbar, C.; Ramírez, M.; Arenas, C. Un método fluorimétrico para la determinación de aminopeptidasas cerebrales [A fluorimetric method for the determination of brain aminopeptidases]. Arch. Neurobiol. 1989, 52, 169–173. [Google Scholar]
- Hui, M.; Hui, K.S. A new type of neuron-specific aminopeptidase NAP-2 in rat brain synaptosomes. Neurochem. Int. 2008, 53, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.; Prieto, I.; Banegas, I.; Segarra, A.B.; Alba, F. Neuropeptidases. Methods Mol. Biol. 2011, 789, 287–294. [Google Scholar]
- Konig, J.F.R.; Klippel, R.A. The Rat Brain; Krieger: Huntington, NY, USA, 1967. [Google Scholar]
- Pellegrino, L.J.; Cushman, A.J. A Stereotaxic Atlas of the Rat Brain; Appleton-Century-Crofts: Norwalk, CT, USA, 1967. [Google Scholar]
- Greenberg, L.J. Fluorometric measurement of alkaline phosphatase and aminopeptidase activities in the order of 10−14 mole. Biochem. Biophys. Res. Commun. 1962, 9, 430–435. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Bernt, E. Lactate dehydrogenase. UV-assay with pyruvate and NADH. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; pp. 574–578. [Google Scholar]
- Barrett, A.J.; Rawlings, N.D.; Woessner, J.F. (Eds.) The Handbook of Proteolytic Enzymes; Academic Press: London, UK, 1998. [Google Scholar]
- Fricker, L.D.; Margolis, E.B.; Gomes, I.; Devi, L.A. Five Decades of Research on Opioid Peptides: Current Knowledge and Unanswered Questions. Mol. Pharmacol. 2020, 98, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Glick, S.D.; Cox, R.D. Striatal asymmetry and morphine reinforcement. Brain Res. 1980, 197, 253–255. [Google Scholar] [CrossRef]
- Hung, C.R.; Hong, J.S.; Bondy, S.C. Lack of asymmetrical distribution of receptor binding sites and of neurally active peptides within rat brain. Neuroscience 1982, 7, 2295–2298. [Google Scholar] [CrossRef]
- Deco, G.; Kringelbach, M.L.; Arnatkeviciute, A.; Oldham, S.; Sabaroedin, K.; Rogasch, N.C.; Aquino, K.M.; Fornito, A. Dynamical consequences of regional heterogeneity in the brain’s transcriptional landscape. Sci. Adv. 2021, 7, eabf4752. [Google Scholar] [CrossRef]
- Perez-Nieves, N.; Leung, V.C.H.; Dragotti, P.L.; Goodman, D.F.M. Neural heterogeneity promotes robust learning. Nat. Commun. 2021, 12, 5791. [Google Scholar] [CrossRef]
- Gazzaniga, M.S.; Sperry, R.W. Language after section of the cerebral commissures. Brain 1967, 90, 131–148. [Google Scholar] [CrossRef]
- Sperry, R. Some effects of disconnecting the cerebral hemispheres. Nobel Lecture, 8 December 1981. Biosci. Rep. 1982, 2, 265–276. [Google Scholar] [CrossRef]
- Knijnenburg, A.C.S.; Steinbusch, C.V.M.; Janssen-Potten, Y.J.M.; Defesche, A.; Vermeulen, R.J. Neuro-imaging characteristics of sensory impairment in cerebral palsy; a systematic review. Front. Rehab. Sci. 2023, 4, 1084746. [Google Scholar] [CrossRef]
- Fabri, M.; Polonara, G. Functional topography of the corpus callosum as revealed by fMRI and behavioural studies of control subjects and patients with callosal resection. Neuropsychologia 2023, 183, 108533. [Google Scholar] [CrossRef]
- Cotovio, G.; Rodrigues da Silva, D.; Real Lage, E.; Seybert, C.; Oliveira-Maia, A.J. Hemispheric asymmetry of motor cortex excitability in mood disorders—Evidence from a systematic review and meta-analysis. Clin. Neurophysiol. 2022, 137, 25–37. [Google Scholar] [CrossRef]
- Lindell, A.K. Lateral thinkers are not so laterally minded: Hemispheric asymmetry, interaction, and creativity. Laterality 2011, 16, 479–498. [Google Scholar] [CrossRef] [PubMed]
- Prieto, I.; Segarra, A.B.; Martinez-Canamero, M.; De Gasparo, M.; Zorad, S.; Ramirez-Sanchez, M. Bidirectional asymmetry in the neurovisceral communication for the cardiovascular control: New insights. Endocr. Regul. 2017, 51, 157–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
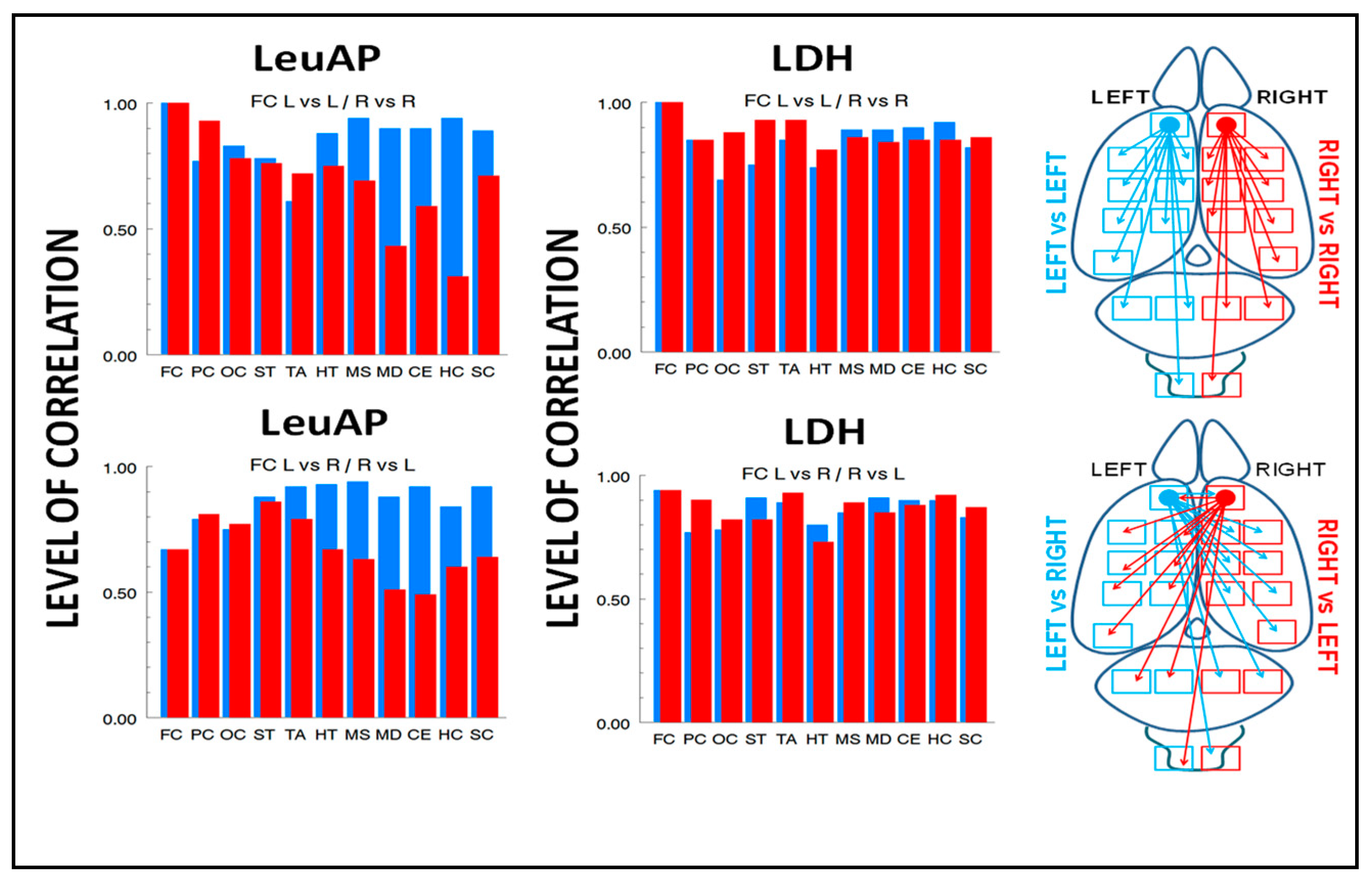
| LeuAP (n = 21) | |||||
|---|---|---|---|---|---|
| A | B | C | D | E | F |
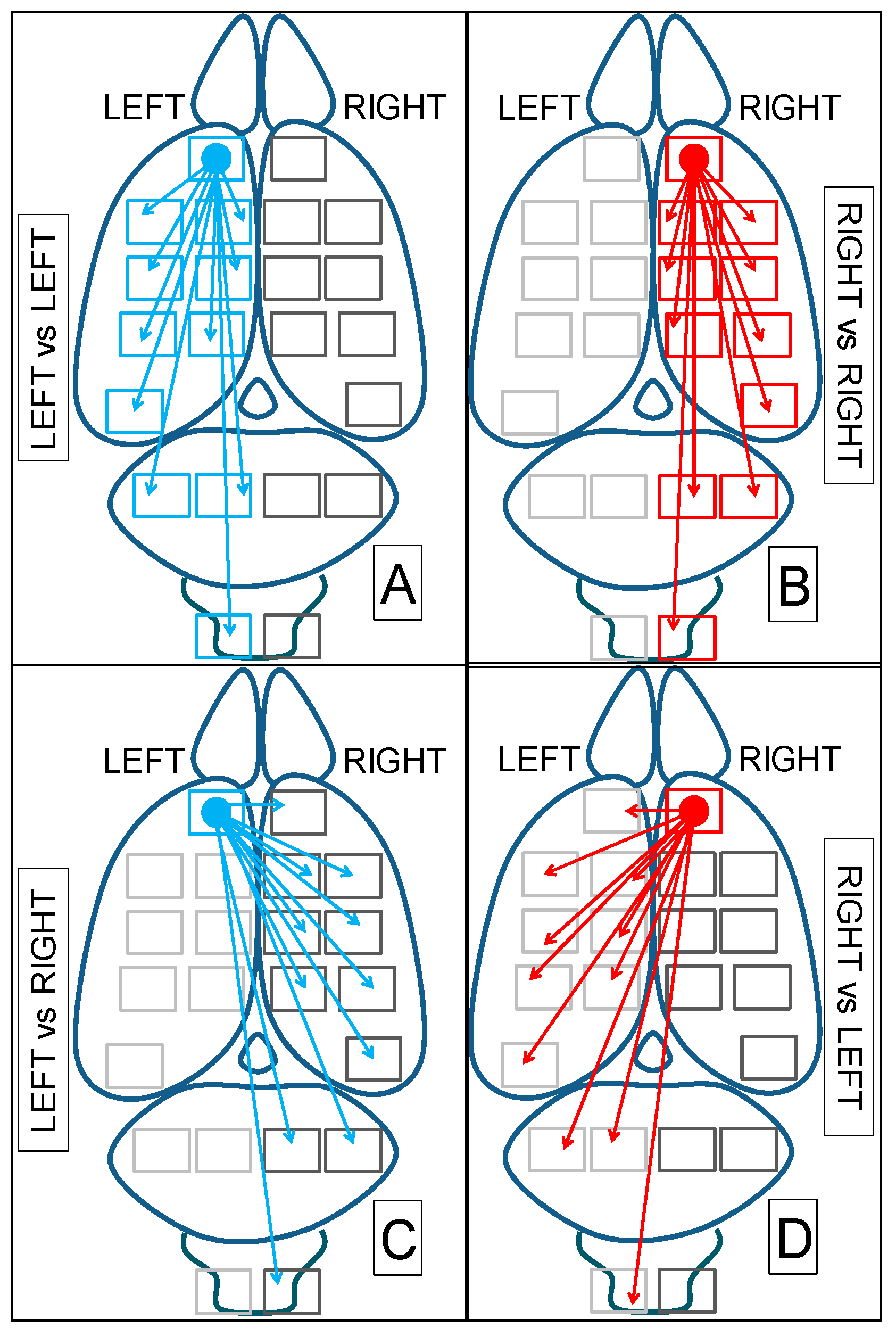
| L vs. L | R vs. R | L vs. R | R vs. L | ||
| LFC | RFC | % | LFC | RFC | % |
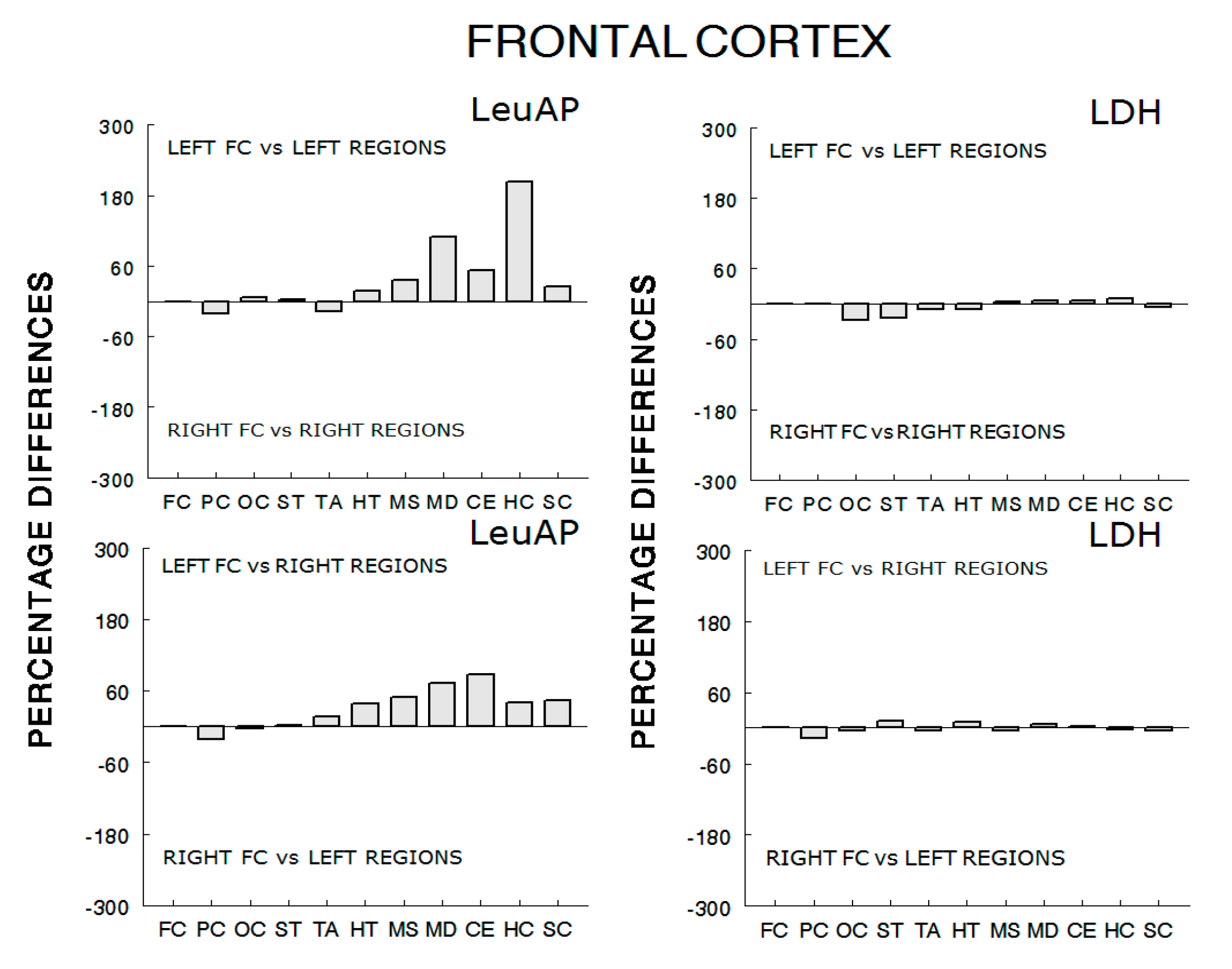
| LFC 1 | RFC 1 | 0 | RFC 0.67 | LFC 0.67 | 0 |
| LPC 0.77 | RPC 0.93 | −20.7 | RPC 0.79 | LPC 0.81 | −20.8 |
| LOC 0.83 | ROC 0.78 | +6.4 | ROC 0.75 | LOC 0.77 | −2.6 |
| LST 0.78 | RST 0.76 | +2.6 | RST 0.88 | LST 0.86 | +2.3 |
| LTA 0.61 | RTA 0.72 | −18 | RTA 0.92 | LTA 0.79 | +16.4 |
| LHT 0.88 | RHT 0.75 | +17.3 | RHT 0.93 | LHT 0.67 | +38.8 |
| LMS 0.94 | RMS 0.69 | +36.2 | RMS 0.94 | LMS 0.63 | +49.2 |
| LMD 0.90 | RMD 0.43 | +109.3 | RMD 0.88 | LMD 0.51 | +72.5 |
| LCE 0.90 | RCE 0.59 | +52.5 | RCE 0.92 | LCE 0.49 | +87.7 |
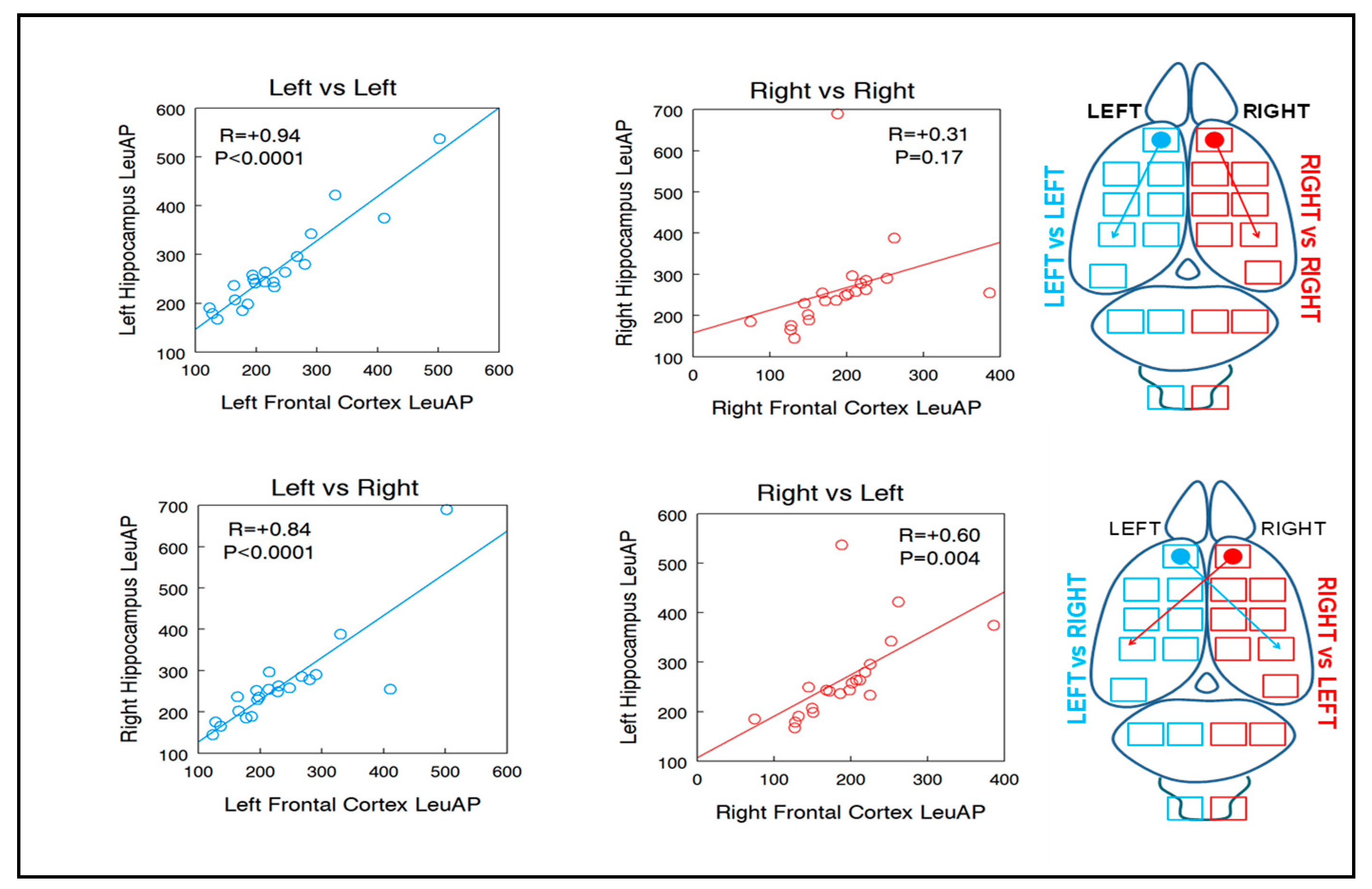
| LHC 0.94 | RHC 0.31 | +203.2 | RHC 0.84 | LHC 0.60 | +40 |
| LSC 0.89 | RSC 0.71 | +25.3 | RSC 0.92 | LSC 0.64 | +43.7 |
| 63.9% | 222.5% | 40.2% | 75.5% | ||
| LDH (n = 18) | |||||
|---|---|---|---|---|---|
| A | B | C | D | E | F |
| L vs. L | R vs. R | L vs. R | R vs. L | ||
| LFC | RFC | % | LFC | RFC | % |
| LFC 1 | RFC 1 | 0 | RFC 0.94 | LFC 0.94 | 0 |
| LPC 0.85 | RPC 0.85 | 0 | RPC 0.77 | LPC 0.90 | −16.8 |
| LOC 0.69 | ROC 0.88 | −27.5 | ROC 0.78 | LOC 0.82 | −5.1 |
| LST 0.75 | RST 0.93 | −24 | RST 0.91 | LST 0.82 | +10.9 |
| LTA 0.85 | RTA 0.93 | −9.4 | RTA 0.89 | LTA 0.93 | −4.4 |
| LHT 0.74 | RHT 0.81 | −9.4 | RHT 0.80 | LHT 0.73 | +9.5 |
| LMS 0.89 | RMS 0.86 | +3.4 | RMS 0.85 | LMS 0.89 | −4.7 |
| LMD 0.89 | RMD 0.84 | +5.9 | RMD 0.91 | LMD 0.85 | +7 |
| LCE 0.90 | RCE 0.85 | +5.8 | RCE 0.90 | LCE 0.88 | +2.2 |
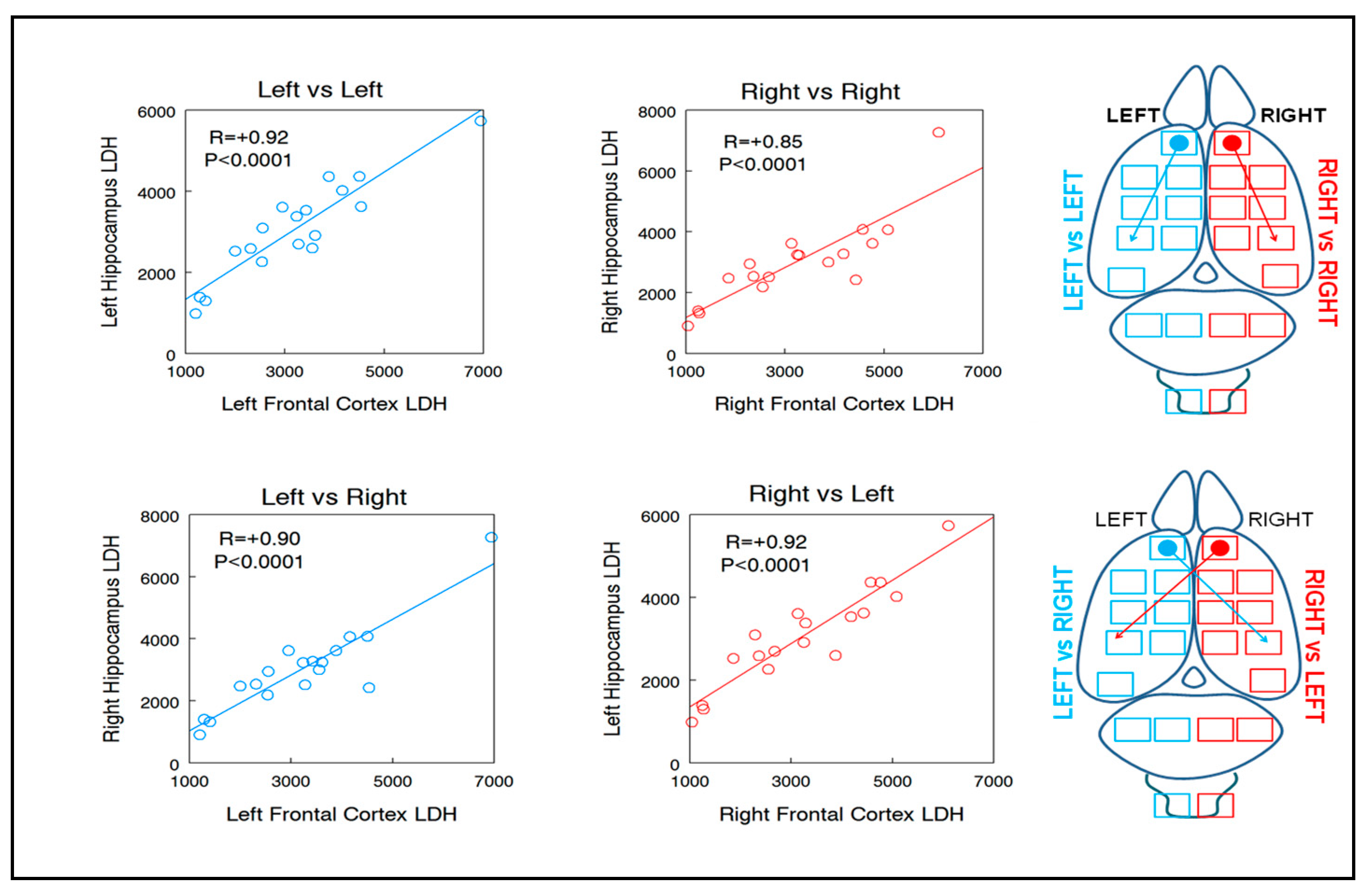
| LHC 0.92 | RHC 0.85 | +8.2 | RHC 0.90 | LHC 0.92 | −2.2 |
| LSC 0.82 | RSC 0.86 | −4.8 | RSC 0.83 | LSC 0.87 | −4.8 |
| 44.9% | 23.4% | 22% | 28.7% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Sánchez, M.; Prieto, I.; Segarra, A.B.; Banegas, I.; Martínez-Cañamero, M.; Domínguez-Vías, G.; Durán, R.; Vives, F.; Alba, F. Asymmetric Pattern of Correlations of Leucine Aminopeptidase Activity between Left or Right Frontal Cortex versus Diverse Left or Right Regions of Rat Brains. Symmetry 2023, 15, 1320. https://doi.org/10.3390/sym15071320
Ramírez-Sánchez M, Prieto I, Segarra AB, Banegas I, Martínez-Cañamero M, Domínguez-Vías G, Durán R, Vives F, Alba F. Asymmetric Pattern of Correlations of Leucine Aminopeptidase Activity between Left or Right Frontal Cortex versus Diverse Left or Right Regions of Rat Brains. Symmetry. 2023; 15(7):1320. https://doi.org/10.3390/sym15071320
Chicago/Turabian StyleRamírez-Sánchez, Manuel, Isabel Prieto, Ana Belén Segarra, Inmaculada Banegas, Magdalena Martínez-Cañamero, Germán Domínguez-Vías, Raquel Durán, Francisco Vives, and Francisco Alba. 2023. "Asymmetric Pattern of Correlations of Leucine Aminopeptidase Activity between Left or Right Frontal Cortex versus Diverse Left or Right Regions of Rat Brains" Symmetry 15, no. 7: 1320. https://doi.org/10.3390/sym15071320