


Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators

Department of Evolutionary Biology, Institute for Biological Research “Siniša Stanković”—National Institute of the Republic of Serbia, University of Belgrade, 11000 Belgrade, Serbia
*
Author to whom correspondence should be addressed.
Symmetry 2023, 15(6), 1149; https://doi.org/10.3390/sym15061149
Submission received: 20 April 2023
/
Revised: 12 May 2023
/
Accepted: 23 May 2023
/
Published: 25 May 2023
(This article belongs to the Special Issue Fluctuating Asymmetry in Evolutionary Biology)
Abstract
:To gain insight into the evolution of flower traits in the generalized food-deceptive plant Iris pumila, we assessed the color, size, shape, and fluctuating asymmetry (FA) of three functionally distinct floral organs—outer perianths (‘falls’), inner perianths (‘standards’), and style branches—and estimated pollinator-mediated selection on these traits. We evaluated the perianth color as the achromatic brightness of the fall, measured the flower stem height, and analyzed the floral organ size, shape, and FA using geometric morphometrics. Pollinated flowers had significantly higher brightness, longer flower stems, and larger floral organs compared to non-pollinated flowers. The shape and FA of the floral organs did not differ, except for the fall FA, where higher values were found for falls of pollinated flowers. Pollinator-mediated selection was confirmed for flower stem height and for subtle changes in the shape of the fall and style branch—organs that form the pollination tunnel. This study provides evidence that, although all analyzed flower traits play significant roles in pollinator attraction, flower stem height and pollination tunnel shape evolved under the pollinator-mediated selection, whereas achromatic brightness, size, and symmetry of floral organs did not directly affect pollination success.
1. Introduction
The main function of the flower is sexual reproduction, a phenomenon that involves two distinct phases: pollination, in which pollen grains are transferred from the anther to the stigma of the pistil, and fertilization, the fusion of male and female gametes [1,2]. Since the pollination process in entomophilous plants exclusively depends on insect pollinators, plants and insects have evolved a complex relationship mediated by the flower [3]. To establish a stable reciprocal relationship, plants provide insects with nectar and pollen as rewards for their pollination services [1,4]. The particular morphological traits of a flower, termed ‘honest signals’, are direct indicators of the quality and quantity of the reward, so the pollinators can learn to choose among different flowers that announce the presence and quality of pollen or nectar [5,6,7]. Therefore, it is not surprising that the remarkable diversity of floral traits, i.e., flower size, shape, color, and odor, is often attributed to pollinator behavior and preference for a particular flower phenotype [8,9,10].
In contrast to this mutualistic interaction, a generalized food-deceptive pollination strategy is a well-known phenomenon related to the utilization of a general set of floral signals that advertise a food reward without actually offering a reward to the pollinators [11,12]. Generalized food-deceptive species are characterized by large and attractive floral displays, a high degree of variability in flower traits, and early spring flowering [11,13]. Large flowers and early spring flowering allow the exploitation of the innate food foraging behavior of newly emerged, naïve pollinators, while high variability in flower traits prevents pollinators from learning visual signals associated with non-rewarding flowers [11,14,15]. This pollination strategy is widespread in orchids, but also occurs in other taxa, including some Iris species [16,17].
A larger floral display, attained through an increase in flower size or the number of open flowers within inflorescence, generally ensures a higher pollinator visitation rate, and consequently enhances pollen transfer efficiency and seed set [5,18,19]. In addition, large flower size, as an honest signal, often correlates with other flower traits that increase pollinator visitation rate, such as symmetry [20,21]. It has been suggested that pollinators tend to visit symmetrical flowers more frequently than asymmetrical ones, due to an innate or learned preference, as larger and more symmetrical flowers tend to offer greater reward [5,22,23]. However, the idea that pollinators act as selective agents for flower symmetry needs to be better understood, as previous studies have not clearly confirmed this hypothesis [21,24,25]. One of the most common methods contributing to this question is to quantify floral asymmetry using a fluctuating asymmetry—small, random deviations from a perfect symmetry that are considered to be the result of developmental instability or/and phenotypic plasticity [26,27,28,29].
The large Iris flowers may have evolved under strong selection pressure imposed by pollinators, either because larger flowers indicate greater reward or because pollinators can detect them from a greater distance [30]. Another distinguishing feature of the genus Iris, including the food-deceptive species Iris pumila, is the complex actinomorphic flower [31,32]. The Iris floral ground plan (and more generally in Iridaceae) includes four trimerous whorls made up of functionally distinct floral organs [33,34,35]. The two outer whorls are formed of morphologically distinct petaloid tepals (fall and standard, respectively), followed by a whorl of three stamens and a tricarpellar gynoecium, with petaloid style branches in the center of the flower (Figure 1). Falls, the external whorl tepals, are curved downward and their distal halves serve as a landing site for pollinating insects. On the adaxial surface, I. pumila falls have a ‘beard’, made of short fine pollen-imitating hairs as well as floral guides, which attract and direct the pollinators towards the pollination tunnel. The inner whorl tepals, called standards, stand upright and serve as long-distance visual signals to potential pollinators. The stamens consist of a filament and an anther, in which large pollen grains are produced. They are hidden under the ventral surface of the petaloid style branches. Each style branch has a receptive stigma at its distal end. The basal parts of the falls, standards, and stamens are fused together to form an elongated hypanthial tube. The proximal part of the fall with the overarching style branch and stamen form a pollination tunnel (Figure 1). Consequently, each I. pumila flower has three separate labiate pollination units (tunnels), which are perceived by pollinators as a single bilaterally symmetrical flower [34,35].
Although a generalized food-deceptive pollination system has been known for a long time, studies on plants other than orchids are scarce. We propose I. pumila as a good model system for studying the evolution of deceptive pollination due to consistent pollinator and pollen limitation, as the basis for pollinator-mediated selection [36], the marked variability in floral traits [37], and its self-incompatibility [12]. Based on the I. pumila flower morphology, with the pollen reward hidden under the upper surface of the pollination tunnel, it can be assumed that this species takes advantage of the naivety of pollinators, mostly generalist hymenopterans, searching for non-existent nectar [17,38]. Several hypotheses have been proposed to explain the existence of the deceptive pollination strategy, but the most available evidence suggests that food deception evolves because it strongly promotes cross-pollination and favors pollen flow over longer distances by causing pollinators to leave non-rewarding patches [9,39]. This could be the possible explanation in the case of I. pumila, as this long-lived plant forms large clones with multiple flowers, which may lead to increased geitonogamy and, consequently, reduce female reproductive success by saturating the stigma with self-pollen [40].
The objective of this study was to identify floral traits that play a functional role in pollinator attraction and, consequently, reproductive success in generalized deceptive pollination systems, and to determine whether they have evolved under the influence of this plant–pollinator interaction. To address these questions, we investigated the phenotypic expression of a suite of flower traits associated with attractiveness to pollinators: flower stem height and floral organs’ color, size, shape, and FA, in naturally pollinated and non-pollinated flowers of I. pumila plants, grown in an experimental garden. If variation in these flower traits prevents pollinators from discriminating between rewarding and non-rewarding flowers, it would be expected that pollinators might prefer certain flower traits without acting as the selection factor on them.
2. Materials and Methods
2.1. Study Species
Iris pumila L. is a long-lived rhizomatous clonal monocot native to the lowlands of central and southeast Europe. The species flowers in early spring and the flowering phase lasts about a few weeks. I. pumila is an enthomophilous, self-incompatible plant [38] that employs a generalized food-deceptive pollination strategy to attract its potential insect pollinators, mostly Bombus, Apis, and small solitary bees (e.g., Halictus and Andrena species) [17,41]. Flowers are large in size and polymorphic in color, with flower colors ranging from dark violet and blue to yellow and white [42].
The flowers of I. pumila are actinomorphic, with hierarchically structured symmetries. While the flower as a whole exhibits threefold rotational symmetry since all floral organs (falls, standards, stamens, and style branches) are arranged around a central axis with their three replicas separated by an angle of 120°, each individual floral organ, when considered as a separate part, is bilaterally symmetric [29].
2.2. Experimental Design and Sample Collection
The experiment was conducted on I. pumila clonal plants originating from Deliblato Sands and grown in clay pots for more than twenty years under common-garden conditions in an experimental garden of the Institute for Biological Research ‘Siniša Stanković’ in Belgrade, Serbia [29,35]. During the blooming phase in spring 2017, flowers were collected daily. Each newly opened flower was examined for the presence of pollen grains on the stigma surface. Flowers with deposited conspecific pollen were regarded as ‘pollinated’, while those ones without pollen on their stigmas as ‘non-pollinated’. At the same time, flower stem height was measured from the ground to the top of the standard (to the nearest 0.5 cm) and expressed in millimeters (mm). Immediately afterward, each flower was harvested, placed separately in a plastic bottle, and transferred to the laboratory. A total of 384 flowers were sampled: 193 pollinated and 191 non-pollinated ones. In the laboratory, the falls positioned beneath the style branches with pollinated stigmas, as well as the arbitrarily chosen falls (one per flower) from non-pollinated flowers, were cut at their bases and scanned using an optical scanner (CanoScan 5600F; Canon Inc., Tokyo, Japan; 600 dpi resolution). Digital images of fresh falls were used for analyses of achromatic brightness, which is defined as the mean RGB value and can vary between zero (black, no light) and one (white, full light intensity) [43,44]. For morphometric analyses, the complete sample of flowers, including the detached falls, was conserved in 70% ethanol at room temperature until dissection. To separate individual floral organs, each flower was cut at the border between the floral organ bases and the end of the floral tube. The organs of each floral whorl were separated, spread on a glass plate coated with 50% glycerol, and scanned (for details, see Tucić et al. [29]).
2.3. Morphometric Analyses
To quantify the variation in floral organ shapes, each organ was described by the relative positions of a set of morphological landmarks [45,46]. The two-dimensional coordinates of landmarks were digitized using a custom-written Iris flower plug-in and ImageJ software [47]. All images were digitized twice by the same person (K.H.).
As detailed in Tucić et al. [29], a set of 18 landmarks was used for the fall (7 pairs and 4 median landmarks) and the style branch (8 pairs and 2 median landmarks), as well as a set of 19 landmarks for the standard (8 pairs and 3 median landmarks).
Centroid size (CS), defined as the square root of the sum of squared distances of all the landmarks from their centroid, was used as a measure of size for each floral organ [46]. To make inferences about the mechanical fit of the pollination tunnel with pollinators, the ratio between fall and style branch CS was calculated.
Procrustes superimposition, which removes the variation in size, position, and orientation according to a least-squares criterion, was used to extract shape information from the landmark configurations of floral organs [46]. To eliminate variations in size, position, and orientation, each landmark configuration was scaled to centroid size 1, translated to the same center of gravity, and rotated so that the sum of squared distances between corresponding landmarks is minimal. To take into account the bilateral symmetry of each separated floral organ, the method for object symmetry was applied. This method uses the landmark configurations and their reflected and relabeled copies to obtain the symmetric and asymmetric components of shape variation. A symmetric component was calculated by averaging the original, reflected and relabeled copies, while the asymmetric component was obtained from their differences [27,28]. Procrustes superimposition and subsequent morphometric analyses were carried out with the MorphoJ software package [48].
To analyze individual shape variation in all three floral organs, principal component analysis (PCA) was carried out on the covariance matrix of the symmetric component of variation. To display characteristics of asymmetry shape variation, PCA was computed using an asymmetric component.
Discriminant function analysis (DFA) was applied to quantify differences in the mean shapes between pollinated and non-pollinated flowers. The statistical significance of pairwise differences in the mean shapes was assessed with a permutation test using Procrustes distances for the null hypothesis of equal group means (1000 permutations per test).
Procrustes ANOVA was performed in order to examine the effects of several factors on the overall variation in the shape of the analyzed floral organs: the effect of inter-individual variation, the effect of side (presence of directional asymmetry), and the effect of interaction between individual and side (presence of fluctuating asymmetry).
To explore the similarity of patterns for fluctuating asymmetry of pollinated and non-pollinated floral organs, covariance matrices for the effect of individuals by side (fluctuating asymmetry) obtained in Procrustes ANOVA were compared. This test evaluates the matrix correlation against the null hypothesis that there is no relationship of any kind between the matrices. To test the association statistically, a matrix permutation test was used. Since covariance matrices were used, the diagonal elements were included in the analysis and landmarks were permuted 10,000 times [27]. The covariance matrices of individual variation were also compared to evaluate the similarity of individual variation patterns between pollinated and non-pollinated flowers.
The amount of fluctuating asymmetry for the floral organs’ shape of pollinated and non-pollinated flowers was estimated using the FA10a index proposed by Palmer and Strobeck [26]. This index describes the magnitude of fluctuating asymmetry of shape for a structure after measurement error has been partitioned out and is calculated as:
where MSsj is the interaction MS, MSm is the error MS from a side’s x individuals Procrustes ANOVA, and M is the number of replicate measurements. Comparisons of FA10a indexes between pollinated and non-pollinated floral organs were performed using the F-test.
FA10a = 0.798 √2(MSsj − MSm)/M,
2.4. Selection Analyses
In this study, one-way ANOVA was used to test the differences in sizes of floral organs, flower stem height, fall-style branch CS ratio, and achromatic brightness between pollinated and non-pollinated flowers.
The selection of floral traits was estimated following the approach of Lande and Arnold [49]. The multivariate selection gradients were estimated by multiple regression of fitness on color, size, and shape variables. PC scores of the first three axes of the symmetric and asymmetric components of floral organ shape variation were used as shape variables. To assess the magnitude and direction of pollinator-mediated selection, plant pollination status was used as a proxy of fitness, with two categorical outcomes (value 1 for plants with pollinated flowers and value 0 for non-pollinated). The linear selection gradient (β) was estimated from the standardized partial regression coefficients and corresponded to the strength and direction of the directional selection acting on each trait. To obtain stabilizing or disruptive selection gradients, the non-linear selection gradient (γ) was estimated as doubled non-linear partial regression coefficients in a quadratic regression [50]. All selection analyses were performed in the Statistica 8 software package, using General Regression Models [51].
3. Results
3.1. The Variation in Floral Organs’ Color and Size in Pollinated and Non-Pollinated Flowers
To examine whether there was a significant difference in the mean values of floral traits between pollinated and non-pollinated flowers, one-way ANOVA was applied. Regarding flower color properties, pollinated flowers exhibited higher achromatic brightness compared to non-pollinated ones, implying that pollinators might prefer brighter flowers (Table 1). Pollinated flowers of I. pumila had significantly higher flower stem as well as CS of all three floral organs compared to non-pollinated flowers (Table 1). In addition, pollinated flowers were found to have a significantly higher ratio of fall and style branch CS, suggesting that a smaller style branch accompanied by a larger fall may be an efficient way to improve mechanical matching between pollinator and flower.
The phenotypic variation in flower color and size was estimated using the coefficient of variation (CV %). The highest phenotypic variation was observed for the color trait, and twice as low for floral organ size (Table 1). Both pollinated and non-pollinated flowers expressed similar levels of variation in all analyzed traits.
Considering flower traits of distinct color morphs separately, no significant differences were observed in flower stem height or in the floral organs’ size—the average stem height and size of falls, style branches, and standards were similar between violet, blue, and yellow color morphs A (Table S1). As expected, different color morphs expressed significantly different achromatic brightness (Table S2).
3.2. The Pattern of Floral Organs’ Shape Variation
Principal component analysis, PCA, was used to identify and display patterns of individual variation and fluctuating asymmetry for the fall, standard, and style branch. The first three PC axes described most of the individual variation in overall shape variance for all analyzed floral organs (fall: 87%; style branch: 87.7%; standard: 87.4%).
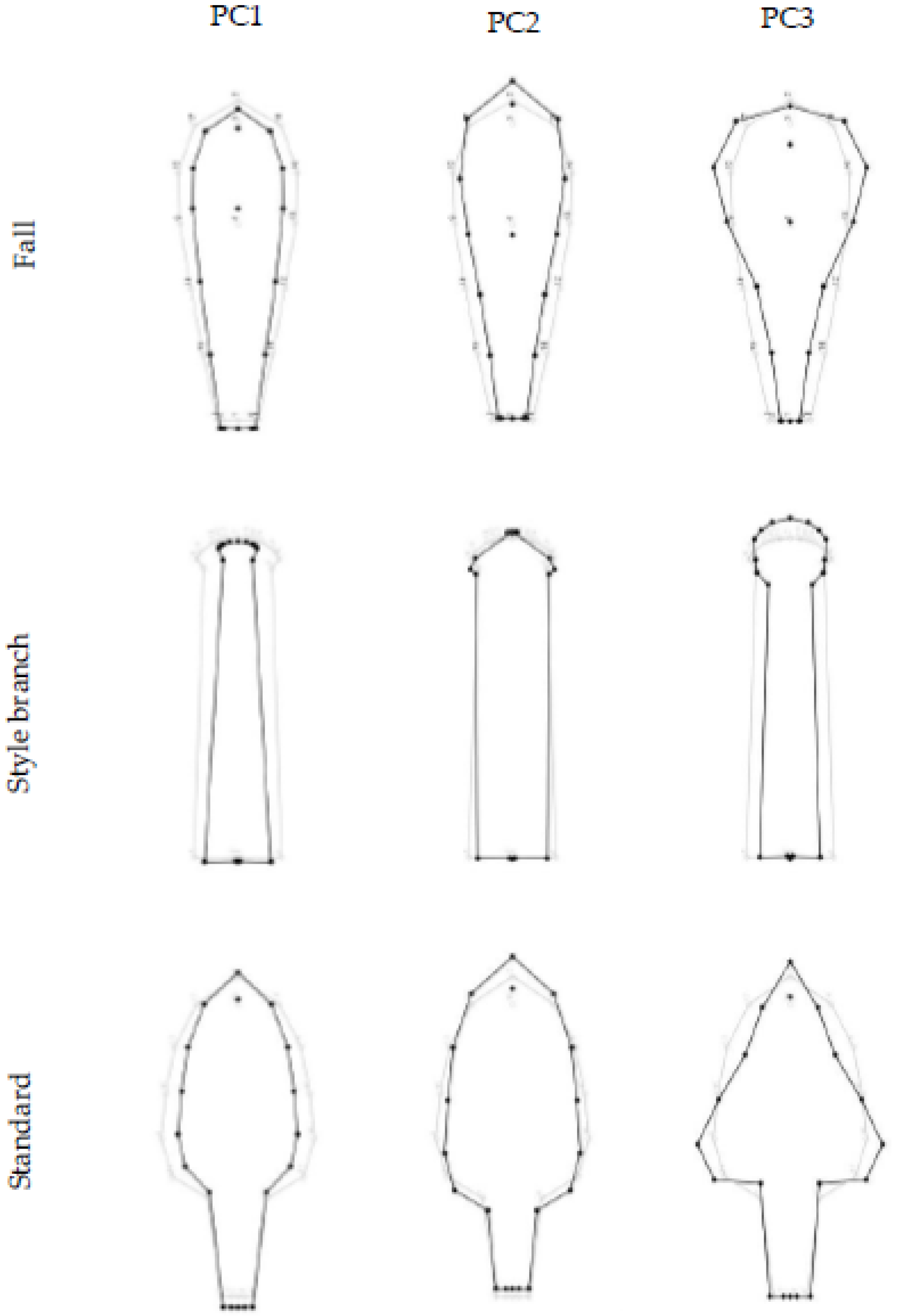
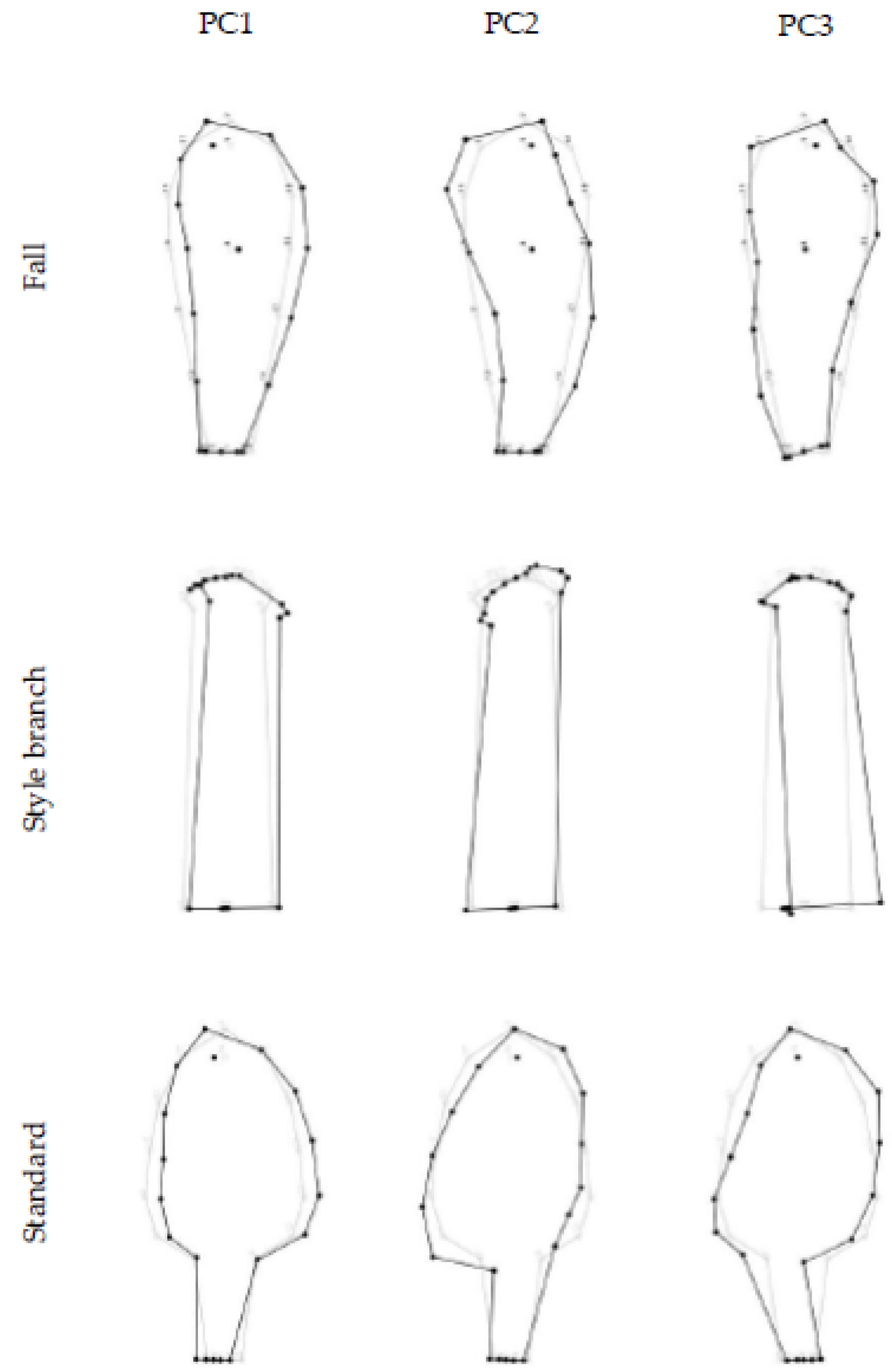
The PC1, PC2, and PC3 for the fall individual shape variation described 42.8%, 33.2%, and 11% of the total phenotypic variation, respectively (Figure 2). PC1 and PC2 shape changes were related to the narrowing and expansion of the entire structure around the longitudinal axis, and additionally, for PC2, to the lengthening and shortening of the fall blade. PC3 corresponded to synchronous changes of the fall blade and haft in opposite directions.
The highest percentage of total individual variation in the shape of the style branch was described by the first three PC axes (63.8%, 16.3%, and 7.6%, respectively) (Figure 2). PC1 referred to the narrowing and expansion of the entire style branch in the transverse direction. PC2 and PC3 described changes in the width and length of the upper part, followed by changes in the width of the basal part of the style branch.
Regarding the pattern of the standard symmetric component of shape variation, PC1, PC2, and PC3 axes described 46.4%, 35.2%, and 5.8% of the total variation, respectively (Figure 2). PC1 was related to the narrowing and the extension of the standard in the transverse direction. PC2 described the lengthening of the blade and the simultaneous shortening of the haft and vice versa, while PC3 was primarily associated with changes in the shape of the standard blade.
The first three PC axes described most of the variation in the fluctuating asymmetry of all three floral organs (fall: 78.8%; style branch: 82.5%; standard: 69.4%) (Figure 3). For the asymmetric component of fall shape variation, PC1, PC2, and PC3 described 55.8%, 14.6%, and 8.4% of the total shape variation, respectively (Figure 3). PC1 described a slight leftward twist of the entire fall relative to the longitudinal axis. PC2 reflected a downward curvature of the right side of the fall blade and an associated upward change in the left side of the blade. PC3 described a more pronounced twist of the blade to the left and haft to the right relative to the longitudinal axis.
The first three PCs of the asymmetric component of the style branch shape variation described 44.9%, 29.6%, and 8% of the total shape variation (Figure 3). PC1 described the bending of the lip area relative to the longitudinal axis of the style branch. PC2 reflected the elevation of the right and a downward movement of the left side of the lip area. PC3 was related to the shift of the style branch base, relative to the longitudinal axis.
PCA of the asymmetric component of standard shape variation revealed that the PC1, PC2, and PC3 accounted for 42.3%, 14.5%, and 12.6% of the total variance, respectively (Figure 3). PC1 characterized the bending of the standard to the left side. PC2 represents an opposite movement of the standard relative to the longitudinal axis. PC3 was associated with the torsion of the blade to the left and the haft to the right side relative to the longitudinal axis.
The difference between the shapes of pollinated and non-pollinated floral organs was evaluated separately for the symmetric and asymmetric components. The discriminant function analysis showed no significant difference in the shape of all three floral organs for both symmetric and asymmetric components of shape variation, as all p values for Procrustes distances after permutation tests were statistically insignificant for all floral organs analyzed (Table S3, Figure S1).
3.3. Analyses of the Asymmetry of Pollinated and Non-Pollinated Floral Organs
The results of Procrustes ANOVA showed that all causal effects had a highly significant impact on shape variation for each of all three organs analyzed, in both pollinated and non-pollinated flowers (Table 2). The term for variation among individuals was significant, indicating the presence of inter-individual variation in the shape of a particular flower organ. The side effect was significant, proving the existence of directional asymmetry in all three floral organs. A significant effect of interaction between individual and side indicated the presence of floral organs’ fluctuating asymmetry in both pollinated and non-pollinated flowers.
Matrix correlations were calculated for the Procrustes ANOVA covariance matrices for the effects of individuals between floral organs of pollinated and non-pollinated flowers. Patterns of individual variation for all three organs analyzed showed a high degree of similarity between pollinated and non-pollinated flowers (fall: 0.992; style branch: 0.992; standard: 0.968; all p < 0.0001). Likewise, high matrix correlations were obtained when covariance matrices of fluctuating asymmetry of floral organs in pollinated and non-pollinated flowers were compared (fall: 0.975; style branch: 0.972; standard: 0.948; all p < 0.0001), indicating a similar pattern of FA.
To quantify the level of floral organs’ fluctuating asymmetry of pollinated and non-pollinated I. pumila flowers, the FA10a index was calculated. The level of fluctuating asymmetry was higher for pollinated falls than for non-pollinated ones (4.08 × 10−3 vs. 3.74 × 10−3). The F-test revealed that for the F value calculated from Procrustes ANOVA (F(1993,1857) = 1.09), the difference between the FA level of pollinated and non-pollinated falls is statistically significant (p = 0.03, at α = 0.05). On the contrary, FA10a indexes for style branches and standards of pollinated flowers exhibited lower values than those obtained for non-pollinated ones (2.57 × 10−3 vs. 2.81 × 10−3, 1.71 × 10−3 vs. 2.15 × 10−3, respectively). Nevertheless, the differences between FA levels of style branches and standards in pollinated and non-pollinated flowers were not found to be statistically significant.
3.4. Phenotypic Selection on Flower Traits
To determine the role of pollinators in flower trait evolution, linear and quadratic selection gradients were estimated (Table 3). It appeared that flower color and floral organ size were not influenced by pollinator-mediated selections, both linear and non-linear. In contrast, it was discovered that flower stem height was significantly affected by positive directional selection (Table 3), implying a positive effect of flower height on the frequency of insect visits. Regarding the floral organs’ shape, positive directional selection was found for the fall symmetric component of shape variation described by PC3, and disruptive selection was found for the style branch symmetric component of shape variation, described by PC2 (Table 3, Figure 2).
Selection analyses for fluctuating asymmetry of floral organs were performed separately, and the obtained linear and non-linear selection gradients were found to be statistically insignificant; therefore, the data are not presented.
4. Discussion
The objective of this study was to gain insight into the evolution of the generalized food-deceptive pollination system in I. pumila by identifying floral traits that increase pollinator visitation and, consequently, the reproductive success of the species.
Deceptive species are expected to exhibit a high degree of variation in floral traits related to conspicuousness and pollinator attraction [37]. The variability of flower traits has evolved to slow pollinators’ associative learning of visual cues that enable them to discriminate non-rewarding flowers from rewarding ones [11,52]. Of all the analyzed I. pumila flower traits, the flower color displayed the greatest phenotypic variation, up to three times greater compared to that found in the floral organ size. This result is supported by the impressive variety of flower color phenotypes this species displays—among nine color phenotypes identified, the three most common ones are purple, blue, and yellow, with estimated frequencies of 0.50, 0.40, and 0.10, respectively [42]. Investigating whether pollinators act as the selection agents for the flower color in irises, Souto-Vilarós et al. [17] did not discover an association between pollinators and female reproductive success of color morphs in two Iris species: polymorphic I. pumila and dimorphic I. lutescens. The only exception was pollinator-mediated stabilizing selection for anthocyanin concentration observed for the I. pumila blue color morph. In the present study, the flower color was characterized in an alternative way—as achromatic brightness, determined as RGB mean score. Higher brightness implies that a particular flower reflects more light, increasing its visibility over long distances and attractiveness to pollinators, so selection for higher flower brightness could occur [44]. Pollinated I. pumila flowers generally exhibited higher brightness values than non-pollinated flowers. Comparing the brightness of pollinated and non-pollinated flowers for each color morph separately, a significantly higher brightness value was found only for the blue morph. It is important to emphasize that this study was conducted in an experimental garden located in urban area. Although the pollinator community probably differs from that in the nature, the presence of potential I. pumila pollinators, such as Bombus, Apis, and small solitary bees, was confirmed [41], and it is likely that this pollinator’s diversity may have led to this result. In addition to brightness, different aspects of the floral visual signals, such as green contrast, represent important cues for distant approach and are considered to provide good proxies for flower conspicuousness to pollinators [53,54]. Therefore, this would be an important direction for further investigation.
The flower stem height represents an important functional trait, as it affects plant performance, i.e., its survival and reproduction [55]. The significant phenotypic correlation of the flower stem height with ovule and seed number may be attributed to the intrinsic developmental relationships [17]. In this study, pollinated flowers exhibited significantly longer flower stems as well as larger sizes of all three floral organs compared to their non-pollinated counterparts. The observed variation in flower stem height was twofold greater than the variation found in floral organ size. These results may suggest that pollinators are generally more attracted to plants with larger and taller flowers compared to the surrounding less sizable ones.
Whether variation in the shape and FA of floral organs is related to attracting pollinators in food-deceptive pollination is still unclear [21,22,23,25]. In I. pumila, the shape and symmetry of the individual floral organs do not seem to play an important role in attracting pollinators, since no difference between the shapes of pollinated and non-pollinated floral organs was observed for both the symmetric and asymmetric components of shape variation. In the previous studies on I. pumila, FA has been used as a tool to evaluate developmental instability in response to environmental and pollution stresses [56,57]. The underlying hypothesis was that a more stressful environment would be reflected by a higher floral organ shape FA [26,28,56,57]. In addition, symmetric structures are thought to be a pollinator’s innate preference [23]. To corroborate this hypothesis, a higher FA of floral organs would be expected in the non-pollinated flowers than in the pollinated ones. For all analyzed I. pumila floral organs, the contribution of FA to the total floral organ shape variation was highly significant and had similar patterns in both pollinated and non-pollinated flowers. The only difference in floral organs’ FA between pollinated and non-pollinated flowers was observed for the fall, being significantly larger in pollinated than in non-pollinated ones. This result contradicts the hypothesis that FA of floral organs could be a potential indicator of fitness in I. pumila. Given that the high variation in floral traits is the key mechanism in the maintenance of plant–pollinator interaction in deceptive pollination systems [12,37], a higher FA detected in the falls of pollinated flowers could be understood in light of this view.
For pollinator-mediated selection to operate on floral traits, both trait variability and pollen limitation must exist [36]. Previously, pollinator limitation was identified in I. pumila, and it appears to be common through the Iris genus [17,30,58,59]. Here, even though the required conditions for pollinator-mediated selection existed, in addition to the significant differences in flower traits between pollinated and non-pollinated flowers, pollinators were not recognized to be strong selection agents. More specifically, pollinators preferred I. pumila flowers with higher brightness, longer flower stems, and larger floral organs, but positive directional pollinator-mediated selection was detected exclusively for the flower stem height. This implies that for successful insect attraction, enhanced long-distance flower visibility, afforded by a longer stem, is of crucial importance. In contrast, the size and shape of the floral organs were not found to be under pollinator-mediated selection [60,61]. Comparable results were reported for other Iris species. Except for the extreme size of the I. atropurpurea flower that was probably selected by pollinators, in other Iris species, pollinator behavior had no effect on the flower size and flower stem height, despite the high correlation between these traits and fitness [17,30,59]. Similarly, no pollinator-mediated selection was observed for brightness. Studies on other deceptive plant species showed that when analyzed over consecutive seasons, pollinator-mediated selection on plant height, size, and color of floral organs is weak and variable, suggesting that changes over time in the strength and direction of pollinator-mediated selection may maintain high phenotypic variation in floral traits in food-deceptive orchids [62,63].
Whether the variation in the floral organs’ shape and FA is subject to selection mediated by pollinators is still an open question. Numerous studies investigating the insect’s ability to consistently discriminate between the flowers based on flower shape and symmetry have not led to a clear conclusion [21,23,25,52,64,65]. To ensure pollen transfer and successful reproduction, I. pumila, like all early-flowering food-deceptive plants, exploits the naivety of recently emerged pollinators [13,17]. The flowers of I. pumila are pollinated by a quite heterogeneous group of large, medium, and small-sized generalist hymenopterans [41]. In addition to their strong innate sensory biases [66,67,68], these insects are also known for their ability to learn and discriminate visual cues based on experience [8,69]. Furthermore, they have an innate preference for higher and larger objects, as well as for particular colors, shapes, and symmetries [21,23,62,69]. Though it is expected that food-deceptive plants take advantage of pollinator preferences for particular colors and shapes, we have revealed that, in the case of I. pumila, pollinators play a minor role in the evolution of floral organ shape. Considering the symmetric component of shape variation, pollinator-mediated selection was discovered for subtle shape changes in fall and style branch—floral organs that form the pollination tunnel. Positive directional selection was found for changes in the shape of the fall, and disruptive selection for the shape of the style branch. These results suggest that even delicate changes in the shape of floral organs related to the pollinator visitation efficiency or to the pollination tunnel assembly are critical. For instance, expansion of the upper part of the fall, which serves as a landing platform for insects, or narrowing of the base of the fall and style branch, which causes a narrower pollination tunnel, affect the flower–pollinator relationship and successful pollen transfer. Furthermore, the larger ratio of fall and style branch sizes found in pollinated flowers points toward a preferred smaller style branch relative to the corresponding fall. All of the above findings indicate that selection on floral organ shape could be due to a better mechanical fitting of the pollination tunnel with the pollinator body rather than a deliberate choice of pollinator [70]. Accordingly, a study by Morinaga and Sakai [71] aimed to examine the functional differentiation between the outer and inner perianths in Iris gracilipes. By experimental manipulation of the inner and outer perianths’ length, the authors showed that different parts of the perianth have different significance for pollinator attraction and fitness. They concluded that the length of outer perianths (i.e., falls) may have evolved as an adaptive trait because flowers with reduced outer perianths gained less pollination success than control flowers. In contrast, flowers with reduced inner perianths (i.e., standards) had similar pollination success to control flowers; hence, the adaptive length of the inner perianths may be shorter than in the contemporary flowers [71]. Considering that the role of the standard is to advertise the flower over a long distance and increase its visibility, their result, although counterintuitive, is consistent with ours, since we found that standard shape had no direct influence on pollination success in I. pumila.
The fitness of organisms may also be related to the bilateral symmetry of a morphological trait, and usually, this symmetry is expressed as fluctuating asymmetry [26,28]. In our study, no pollinator-mediated selection on the asymmetric component of shape variation was observed in any of the three floral organs, confirming that fluctuating asymmetry of floral organ shape is neither universal nor a strong visual cue for pollinators to discriminate between rewarding and non-rewarding flowers. It is important to emphasize that in this study, the shape and FA were analyzed at the level of the floral organs, not at the level of the whole flower. Given the complex structure of the Iris flower, there is a possibility that an analysis of shape and symmetry at the level of the whole flower would suggest a different result.
5. Conclusions
The pollinators of I. pumila showed higher affinity for the flowers with taller flower stem and greater brightness and size of floral organs. The shape and FA of floral organs did not affect pollinator visitation rate, with the exception of the fall FA, where a higher level of FA was found for the pollinated flowers.
Pollinator-mediated selection was confirmed for flower stem height and for subtle changes in the shape of the fall and style branch—organs that form the pollination tunnel. Conversely, FA of floral organs was not identified as an important visual cue for pollinators to discriminate between rewarding and non-rewarding flowers in I. pumila.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/sym15061149/s1, Table S1: The mean (), standard error (SE), and phenotypic coefficient of variation (CV%) for achromatic brightness of perianth and size traits in three I. pumila color morphs; Table S2: Achromatic brightness of perianth in three I. pumila color morphs. The mean (), standard error (SE), and phenotypic coefficient of variation (CV%) were calculated for each trait; Table S3: Discriminant function analysis of the difference in the mean shape of all three floral organs for both symmetric and asymmetric components of shape variation among pollinated and non-pollinated I. pumila flowers; Figure S1: The mean shape diagrams of pollinated and non-pollinated flower organs: fall, style branch, and standard.
Author Contributions
Conceptualization, S.M.J. and A.V.; methodology, S.B. and A.V.; validation, S.B., S.M.J., K.H. and A.V.; formal analysis, K.H. and A.V.; investigation, S.B., K.H., S.M.J. and A.V.; writing—original draft preparation, A.V.; writing—review and editing, S.B., K.H., S.M.J. and A.V.; visualization, A.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia, grant number 451-03-47/2023-01/ 200007.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available because the authors will use them in future research.
Acknowledgments
We thank the four anonymous reviewers for their helpful comments on a previous version of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harder, L.D.; Barrett, S.C.H. Pollen Dispersal and Mating Patterns in Animal-Pollinated Plants. In Floral Biology, 1st ed.; Lloyd, D.G., Barrett, S.C.H., Eds.; Springer: Boston, MA, USA, 1996; pp. 140–190. [Google Scholar]
- Harder, L.D.; Barrett, S.C.H. Pollen and ovule fates and reproductive performance by flowering plants. In Ecology and Evolution of Flowers, 1st ed.; Harder, L.D., Routley, M.B., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2006; pp. 61–80. [Google Scholar]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Simpson, B.B.; Neff, J.L. Floral rewards: Alternatives to pollen and nectar. Ann. Missouri Bot. Gard. 1981, 68, 301–322. [Google Scholar] [CrossRef]
- Blarer, A.; Keasar, T.; Shmida, A. Possible mechanisms for the formation of flower size preferences by foraging bumblebees. Ethology 2002, 108, 341–351. [Google Scholar] [CrossRef]
- Knauer, A.C.; Schiestl, F.P. Bees use honest floral signals as indicators of reward when visiting flowers. Ecol. Lett. 2015, 18, 135–143. [Google Scholar] [CrossRef]
- Ito, K.; Suzuki, M.F.; Mochizuki, K. Evolution of honest reward signal in flowers. Proc. R. Soc. B 2021, 288, 20202848. [Google Scholar] [CrossRef]
- Spaethe, J.; Tautz, J.; Chittka, L. Visual constraints in foraging bumblebees: Flower size and color affect search time and flight behavior. Proc. Natl. Acad. Sci. USA 2001, 98, 3898–3903. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Johnson, S.D. Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 2013, 28, 307–315. [Google Scholar] [CrossRef]
- Gómez, J.M.; Torices, R.; Lorite, J.; Klingenberg, C.P.; Perfectti, F. The role of pollinators in the evolution of corolla shape variation, disparity and integration in a highly diversified plant family with a conserved floral bauplan. Ann. Bot. 2016, 117, 889–904. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Jürgens, A. Deceptive behavior in plants. II. Food deception by plants: From generalized systems to specialized floral mimicry. In Plant-Environment Interaction, 1st ed.; Baluška, F., Ed.; Springer: Berlin, Germany, 2009; pp. 223–246. [Google Scholar]
- Johnson, S.D.; Schiestl, F.P. Floral Mimicry, 1st ed.; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Pellissier, L.; Vittoz, P.; Internicola, A.I.; Gigord, L.D.B. Generalized food-deceptive orchid species flower earlier and occur at lower altitudes than rewarding ones. J. Plant Ecol. 2010, 3, 243–250. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235. [Google Scholar] [CrossRef]
- Juillet, N.; Scopece, G. Does floral trait variability enhance reproductive success in deceptive orchids? Perspect. Plant Ecol. Evol. Syst. 2010, 12, 317–322. [Google Scholar] [CrossRef]
- Renner, S.S. Rewardless flowers in the angiosperms and the role of insect cognition in their evolution. In Plant-Pollinator Interactions: From Specialization to Generalization, 1st ed.; Waser, N.M., Olletron, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2006; pp. 123–144. [Google Scholar]
- Souto-Vilarós, D.; Vuleta, A.; Jovanović, S.M.; Budečević, S.; Wang, H.; Sapir, Y.; Imbert, E. Are pollinators the agents of selection on flower colour and size in irises? Oikos 2018, 127, 834–846. [Google Scholar] [CrossRef]
- Engel, E.C.; Irwin, R.E. Linking pollinator visitation rate and pollen receipt. Am. J. Bot. 2003, 90, 1612–1618. [Google Scholar] [CrossRef]
- Makino, T.T.; Sakai, S. Experience changes pollinator responses to floral display size: From size-based to reward-based foraging. Funct. Ecol. 2007, 21, 854–863. [Google Scholar] [CrossRef]
- Fenster, C.B.; Cheely, G.; Dudash, M.R.; Reynolds, R.J. Nectar reward and advertisement in hummingbird-pollinated Silene virginica (Caryophyllaceae). Am. J. Bot. 2006, 93, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Frey, F.M.; Bukoski, M. Floral symmetry is associated with flower size and pollen production but not insect visitation rates in Geranium robertianum (Geraniaceae). Plant Species Biol. 2014, 29, 272–280. [Google Scholar] [CrossRef]
- Møller, A.P.; Eriksson, M. Patterns of fluctuating asymmetry in flowers: Implications for sexual selection in plants. J. Evol. Biol. 1994, 7, 97–113. [Google Scholar] [CrossRef]
- Møller, A.P. Bumblebee preference for symmetrical flowers. Proc. Natl. Acad. Sci. USA 1995, 92, 2288–2292. [Google Scholar] [CrossRef]
- Midgley, J.J.; Johnson, S.D. Some pollinators do not prefer symmetrically marked or shaped daisy (Asteraceae) flowers. Evol. Ecol. 1998, 12, 123–126. [Google Scholar] [CrossRef]
- Frey, F.M.; Davis, R.; Delph, L.F. Manipulation of floral symmetry does not affect seed production in Impatiens pallida. Int. J. Plant Sci. 2005, 166, 659–662. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry analyses revisited. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 279–319. [Google Scholar]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [PubMed]
- Klingenberg, C.P. Analyzing fluctuating asymmetry with geometric morphometrics: Concepts, methods, and applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Tucić, B.; Budečević, S.; Manitašević Jovanović, S.; Vuleta, A.; Klingenberg, C.P. Phenotypic plasticity in response to environmental heterogeneity contributes to fluctuating asymmetry in plants: First empirical evidence. J. Evol. Biol. 2018, 31, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Lavi, R.; Sapir, Y. Are pollinators the agents of selection for the extreme large size and dark color in Oncocyclus irises? New Phytol. 2015, 205, 369–377. [Google Scholar] [CrossRef]
- Mathew, B. The Iris, 1st ed.; Timber Press: Portland, OR, USA, 1981. [Google Scholar]
- Guo, J.; Wilson, C.A. Organization, available space and organ morphology within floral buds of Iris (Iridaceae). Flora 2018, 249, 67–76. [Google Scholar] [CrossRef]
- Remizowa, M.V.; Rudall, P.J.; Choob, V.V.; Sokoloff, D.D. Racemose inflorescences of monocots: Structural and morphogenetic interaction at the flower/inflorescence level. Ann. Bot. 2013, 112, 1553–1566. [Google Scholar] [CrossRef] [PubMed]
- Radović, S.; Urošević, A.; Hočevar, K.; Vuleta, A.; Manitašević Jovanović, S.; Tucić, B. Geometric morphometrics of functionally distinct floral organs in Iris pumila: Analyzing patterns of symmetric and asymmetric shape variations. Arch. Biol. Sci. 2017, 69, 223–231. [Google Scholar] [CrossRef]
- Budečević, S.; Manitašević Jovanović, S.; Vuleta, A.; Tucić, B.; Klingenberg, C.P. Directional asymmetry and direction-giving factors: Lessons from flowers with complex symmetry. Evol. Dev. 2022, 24, 92–108. [Google Scholar] [CrossRef]
- Ashman, T.L.; Knight, T.M.; Steets, J.A.; Amarasekare, P.; Burd, M.; Campbell, D.R.; Dudash, M.R.; Johnston, M.O.; Mazer, S.J.; Mitchell, R.J. Pollen limitation of plant reproduction: Ecological and evolutionary causes and consequences. Ecology 2004, 85, 2408–2421. [Google Scholar] [CrossRef]
- Ackerman, J.D.; Cuevas, A.A.; Hof, D. Are deception-pollinated species more variable than those offering a reward? Plant Syst. Evol. 2011, 293, 91–99. [Google Scholar] [CrossRef]
- Tarasjev, A. Impact of genet size and flowering stage on fruit set in Iris pumila L. clones in wild. Acta Oecologica 2005, 27, 93–98. [Google Scholar] [CrossRef]
- Walsh, R.P.; Michaels, H.J. When it pays to cheat: Examining how generalized food deception increases male and female fitness in a terrestrial orchid. PLoS ONE 2017, 12, e0171286. [Google Scholar] [CrossRef] [PubMed]
- Vallejo-Marín, M.; Dorken, M.E.; Barrett, S.C.H. The ecological and evolutionary consequences of clonality for plant mating. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 193–213. [Google Scholar] [CrossRef]
- Vuleta, A.; Hočevar, K.; Manitašević Jovanović, S.; Raičević, J.; Plećaš, M. Assessment of Iris pumila L. pollinator and florivore diversity in a common garden: A pan-trap experiment. In Proceedings of the 4th International Conference on Plant Biology (23rd SPPS Meeting), Belgrade, Serbia, 6–8 October 2022. [Google Scholar]
- Tucić, B.; Milojković, S.; Tarasjev, A.; Vijčić, S. The influence of climatic factors on clonal diversity in a population of Iris pumila. Oikos 1989, 56, 115–120. [Google Scholar] [CrossRef]
- Lambert, P.; Carron, T. Symbolic fusion of luminance-hue-chroma features for region segmentation. Pattern Recognit. 1999, 32, 1857–1872. [Google Scholar] [CrossRef]
- Sletvold, N.; Trunschke, J.; Smit, M.; Verbeek, J.; Ågren, J. Strong pollinator-mediated selection for increased flower brightness and contrast in a deceptive orchid. Evolution 2016, 70, 716–724. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis: With Applications in R, 1st ed.; John Wiley & Sons: Chichester, UK, 2016. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Lande, R.; Arnold, S.J. The measurement of selection on correlated characters. Evolution 1983, 37, 1210–1226. [Google Scholar] [CrossRef]
- Stinchcombe, J.R.; Agrawal, A.F.; Hohenlohe, P.A.; Arnold, S.J.; Blows, M.W. Estimating nonlinear selection gradients using quadratic regression coefficients: Double or nothing? Evolution 2008, 62, 2435–2440. [Google Scholar] [CrossRef]
- Weiß, C.H. Statsoft, Inc., Tulsa, OK: STATISTICA, version 8. Asta-Adv. Stat. Anal. 2007, 91, 339–341. [Google Scholar] [CrossRef]
- Kagawa, K.; Takimoto, G. Inaccurate color discrimination by pollinators promotes evolution of discrete color polymorphism in food-deceptive flowers. Am. Nat. 2016, 187, 194–204. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Dyer, A.G.; Kevan, P.G.; Lunau, K. Functional significance of the optical properties of flowers for visual signalling. Ann Bot. 2019, 123, 263–276. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Spaethe, J. Caution with colour calculations: Spectral purity is a poor descriptor of flower colour visibility. Ann Bot. 2022, 130, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Barišić Klisarić, N.; Miljković, D.; Avramov, S.; Živković, U.; Tarasjev, A. Radial and bilateral fluctuating asymmetry of Iris pumila flowers as indicators of environmental stress. Symmetry 2019, 11, 818. [Google Scholar] [CrossRef]
- Tucić, B.; Miljković, D. Fluctuating asymmetry of floral organ traits in natural populations of Iris pumila from contrasting light habitats. Plant Species Biol. 2010, 25, 173–184. [Google Scholar] [CrossRef]
- Segal, B.; Sapir, Y.; Carmel, Y. Fragmentation and pollination crisis in the self-incompatible Iris bismarckiana (Iridaceae), with implications for conservation. Isr. J. Ecol. Evol. 2006, 52, 111–122. [Google Scholar] [CrossRef]
- Pellegrino, G. Pollinator limitation on reproductive success in Iris tuberosa. AoB Plants 2015, 7, plu089. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.; Flick, A.J.; Bauer, A.A. Phenotypic selection on flower color and floral display size by three bee species. Front. Plant Sci. 2021, 11, 587528. [Google Scholar] [CrossRef]
- Caruso, C.M.; Eisen, K.E.; Martin, R.A.; Sletvold, N. A meta-analysis of the agents of selection on floral traits. Evolution 2019, 73, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Trunschke, J.; Lunau, K.; Pyke, G.H.; Ren, Z.-X.; Wang, H. Flower Color Evolution and the Evidenceof Pollinator-Mediated Selection. Front. Plant Sci. 2021, 12, 617851. [Google Scholar] [CrossRef] [PubMed]
- Scopece, G.; Juillet, N.; Lexer, C.; Cozzolino, S. Fluctuating selection across years and phenotypic variation in food-deceptive orchids. PeerJ 2017, 5, e3704. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Burczyk, J. Selection on floral traits through male fertility in a natural plant population. Evol. Ecol. 2008, 22, 39–54. [Google Scholar] [CrossRef]
- Lehrer, M.; Horridge, G.A.; Zhang, S.W.; Gadagkar, R. Shape vision in bees: Innate preference for flower-like patterns. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 1995, 347, 123–137. [Google Scholar]
- Lunau, K.; Wacht, S.; Chittka, L. Colour choices of naive bumble bees and their implications for colour perception. J. Comp. Physiol. A 1996, 178, 477–489. [Google Scholar] [CrossRef]
- Gumbert, A. Color choices by bumble bees (Bombus terrestris): Innate preferences and generalization after learning. Behav. Ecol. Sociobiol. 2000, 48, 36–43. [Google Scholar] [CrossRef]
- Raine, N.E.; Ings, T.C.; Dornhaus, A.; Saleh, N.; Chittka, L. Adaptation, genetic drift, pleiotropy, and history in the evolution of bee foraging behavior. Adv. Study Behav. 2006, 36, 305–354. [Google Scholar]
- Lehrer, M. Shape perception in the honeybee: Symmetry as a global framework. Int. J. Plant Sci. 1999, 160, S51–S65. [Google Scholar] [CrossRef]
- La Rosa, R.J.; Conner, J.K. Floral function: Effects of traits on pollinators, male and female pollination success, and female fitness across three species of milkweeds (Asclepias). Am. J. Bot. 2017, 104, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Morinaga, S.I.; Sakai, S. Functional differentiation in pollination processes between the outer and inner perianths in Iris gracilipes (Iridaceae). Botany 2006, 84, 164–171. [Google Scholar] [CrossRef]
Figure 1.
Ground plan of an Iris flower, illustrated by the example of I. pumila. (A) Lateral view of pollination tunnel. Floral organs are indicated with arrows. (B) Image of a bumblebee visiting I. pumila flower in an experimental garden.
Figure 1.
Ground plan of an Iris flower, illustrated by the example of I. pumila. (A) Lateral view of pollination tunnel. Floral organs are indicated with arrows. (B) Image of a bumblebee visiting I. pumila flower in an experimental garden.

Figure 2.
PCA for individual variation in I. pumila floral organs: fall, style branch, and standard. The diagrams represent shape changes from the mean shape (gray lines) to a configuration with an arbitrary PC score of +0.1 unit of Procrustes distance (black line). The sign for PCs is arbitrary and the opposite shape change is equivalent.
Figure 2.
PCA for individual variation in I. pumila floral organs: fall, style branch, and standard. The diagrams represent shape changes from the mean shape (gray lines) to a configuration with an arbitrary PC score of +0.1 unit of Procrustes distance (black line). The sign for PCs is arbitrary and the opposite shape change is equivalent.

Figure 3.
PCA for the fluctuating asymmetry in I. pumila floral organs: fall, style branch, and standard. The diagrams represent shape changes from the mean shape (gray lines) to a configuration with an arbitrary PC score of +0.1 unit of Procrustes distance (black line). The sign for PCs is arbitrary and the opposite shape change is equivalent.
Figure 3.
PCA for the fluctuating asymmetry in I. pumila floral organs: fall, style branch, and standard. The diagrams represent shape changes from the mean shape (gray lines) to a configuration with an arbitrary PC score of +0.1 unit of Procrustes distance (black line). The sign for PCs is arbitrary and the opposite shape change is equivalent.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The mean (), standard error (SE), and phenotypic coefficient of variation (CV%) for color (achromatic brightness) and size (flower stem height in mm; centroid size of fall, style branch, and standard) flower traits expressed in pollinated and non-pollinated I. pumila flowers. One-way ANOVA was used to compare the mean values of flower traits between pollinated and non-pollinated flowers. For all ANOVAs, df = 1.
Table 1.
The mean (), standard error (SE), and phenotypic coefficient of variation (CV%) for color (achromatic brightness) and size (flower stem height in mm; centroid size of fall, style branch, and standard) flower traits expressed in pollinated and non-pollinated I. pumila flowers. One-way ANOVA was used to compare the mean values of flower traits between pollinated and non-pollinated flowers. For all ANOVAs, df = 1.
| Pollinated Flower | Non-Pollinated Flower | ANOVA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trait | n | SE | CV % | n | SE | CV % | F | p | ||
| Achromatic brightness | 193 | 0.266 | 0.005 | 26.19 | 191 | 0.251 | 0.005 | 24.85 | 5.143 | 0.024 |
| Flower stem height | 193 | 97.42 | 1.49 | 21.29 | 191 | 87.54 | 1.26 | 19.89 | 25.58 | <0.0001 |
| CS fall | 193 | 338.7 | 3.0 | 12.20 | 191 | 321.5 | 2.8 | 12.18 | 17.06 | <0.0001 |
| CS style branch | 193 | 198.2 | 1.4 | 9.62 | 191 | 192.2 | 1.4 | 10.09 | 9.19 | 0.003 |
| CS standard | 193 | 335.1 | 3.1 | 13.00 | 191 | 315.3 | 3.0 | 13.12 | 20.60 | <0.0001 |
| Fall-style branch CS ratio | 193 | 1.713 | 0.013 | 10.39 | 191 | 1.676 | 0.011 | 9.45 | 4.43 | 0.036 |
Table 2.
Procrustes ANOVA of floral organs’ shape variation in pollinated and non-pollinated I. pumila flowers. The effects of individual (inter-individual variation), side (directional asymmetry), interaction between individual and side (fluctuating asymmetry), and measurement errors are shown. SS—Procrustes sums of squares, MS—Procrustes mean squares, df—degree of freedom, F—Goodall’s F statistic, p—level of statistical significance.
Table 2.
Procrustes ANOVA of floral organs’ shape variation in pollinated and non-pollinated I. pumila flowers. The effects of individual (inter-individual variation), side (directional asymmetry), interaction between individual and side (fluctuating asymmetry), and measurement errors are shown. SS—Procrustes sums of squares, MS—Procrustes mean squares, df—degree of freedom, F—Goodall’s F statistic, p—level of statistical significance.
| Pollinated Floral Organs | Non-Pollinated Floral Organs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Effect | SS | MS | df | F | p | SS | MS | df | F | p |
| Fall | ||||||||||
| Individual | 0.73483 | 0.00024 | 3056 | 7.51 | <0.0001 | 0.68965 | 0.00023 | 2992 | 8.34 | <0.0001 |
| Side | 0.05519 | 0.00345 | 16 | 107.7 | <0.0001 | 0.04922 | 0.00308 | 16 | 111.3 | <0.0001 |
| Ind * Side | 0.09787 | 0.00003 | 3056 | 5.39 | <0.0001 | 0.08273 | 0.00003 | 2992 | 4.90 | <0.0001 |
| Error 1 | 0.03593 | 0.00001 | 6048 | 0.03338 | 0.00001 | 5920 | ||||
| Style branch | ||||||||||
| Individual | 0.47477 | 0.00016 | 3056 | 6.64 | <0.0001 | 0.43848 | 0.00014 | 3040 | 5.25 | <0.0001 |
| Side | 0.00449 | 0.00028 | 16 | 11.99 | <0.0001 | 0.00461 | 0.00029 | 16 | 10.49 | <0.0001 |
| Ind * Side | 0.07150 | 0.00002 | 3056 | 1.80 | <0.0001 | 0.08352 | 0.00003 | 3040 | 1.82 | <0.0001 |
| Error 1 | 0.08004 | 0.00001 | 6144 | 0.09210 | 0.00002 | 6112 | ||||
| Standard | ||||||||||
| Individual | 1.13133 | 0.00035 | 3264 | 7.68 | <0.0001 | 1.26642 | 0.00039 | 3247 | 8.59 | <0.0001 |
| Side | 0.07219 | 0.00425 | 17 | 94.11 | <0.0001 | 0.06520 | 0.00384 | 17 | 84.46 | <0.0001 |
| Ind * Side | 0.14727 | 0.00005 | 3264 | 1.11 | 0.0002 | 0.14746 | 0.00005 | 3247 | 1.19 | <0.0001 |
| Error 1 | 0.26593 | 0.00004 | 6562 | 0.24915 | 0.00004 | 6528 | ||||
Table 3.
Standardized linear (β) and non-linear (γ) selection gradients (±SE) for floral traits related to pollinator attraction, in pollinated and non-pollinated I. pumila flowers.
Table 3.
Standardized linear (β) and non-linear (γ) selection gradients (±SE) for floral traits related to pollinator attraction, in pollinated and non-pollinated I. pumila flowers.
| Trait | β ± SE | p | γ ± SE | p |
|---|---|---|---|---|
| Achromatic brightness | 0.074 ± 0.050 | 0.143 | −0.052 ± 0.246 | 0.915 |
| Flower stem height | 0.176 ± 0.069 | 0.011 | 0.094 ± 0.318 | 0.883 |
| CS fall | −0.392 ± 0.686 | 0.568 | 3.273 ± 5.116 | 0.749 |
| CS style branch | 0.276 ± 0.532 | 0.604 | −2.271 ± 3.642 | 0.755 |
| CS standard | 0.142 ± 0.132 | 0.282 | −1.885 ± 1.106 | 0.395 |
| Fall-style branch CS ratio | 0.278 ± 0.532 | 0.602 | 1.053 ± 3.849 | 0.891 |
| Fall shape | ||||
| PC1 | −0.014 ± 0.052 | 0.791 | 0.022 ± 0.052 | 0.830 |
| PC2 | 0.012 ± 0.052 | 0.821 | 0.089 ± 0.055 | 0.418 |
| PC3 | 0.151 ± 0.052 | 0.004 | 0.051 ± 0.052 | 0.627 |
| Style branch shape | ||||
| PC1 | 0.048 ± 0.051 | 0.349 | −0.026 ± 0.053 | 0.807 |
| PC2 | −0.065 ± 0.051 | 0.203 | 0.208 ± 0.052 | 0.047 |
| PC3 | 0.090 ± 0.051 | 0.080 | 0.199 ± 0.052 | 0.055 |
| Standard shape | ||||
| PC1 | −0.055 ± 0.051 | 0.285 | −0.105 ± 0.052 | 0.311 |
| PC2 | 0.021 ± 0.051 | 0.680 | −0.016 ± 0.052 | 0.878 |
| PC3 | 0.038 ± 0.051 | 0.456 | −0.034 ± 0.052 | 0.744 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Budečević, S.; Hočevar, K.; Manitašević Jovanović, S.; Vuleta, A. Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators. Symmetry 2023, 15, 1149. https://doi.org/10.3390/sym15061149
AMA Style
Budečević S, Hočevar K, Manitašević Jovanović S, Vuleta A. Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators. Symmetry. 2023; 15(6):1149. https://doi.org/10.3390/sym15061149
Chicago/Turabian StyleBudečević, Sanja, Katarina Hočevar, Sanja Manitašević Jovanović, and Ana Vuleta. 2023. "Phenotypic Selection on Flower Traits in Food-Deceptive Plant Iris pumila L.: The Role of Pollinators" Symmetry 15, no. 6: 1149. https://doi.org/10.3390/sym15061149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.