
Stronger Hurricanes and Climate Change in the Caribbean Sea: Threats to the Sustainability of Endangered Coral Species

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Temporal Variation in Storm Frequency and Intensity across the Northeastern Caribbean
2.3. Status of Acropora palmata Populations
2.4. Status of Acropora cervicornis, Orbicella spp., and Dendrogyra cylindrus Populations
3. Results
3.1. Hurricane Intensification across the Northeastern Caribbean
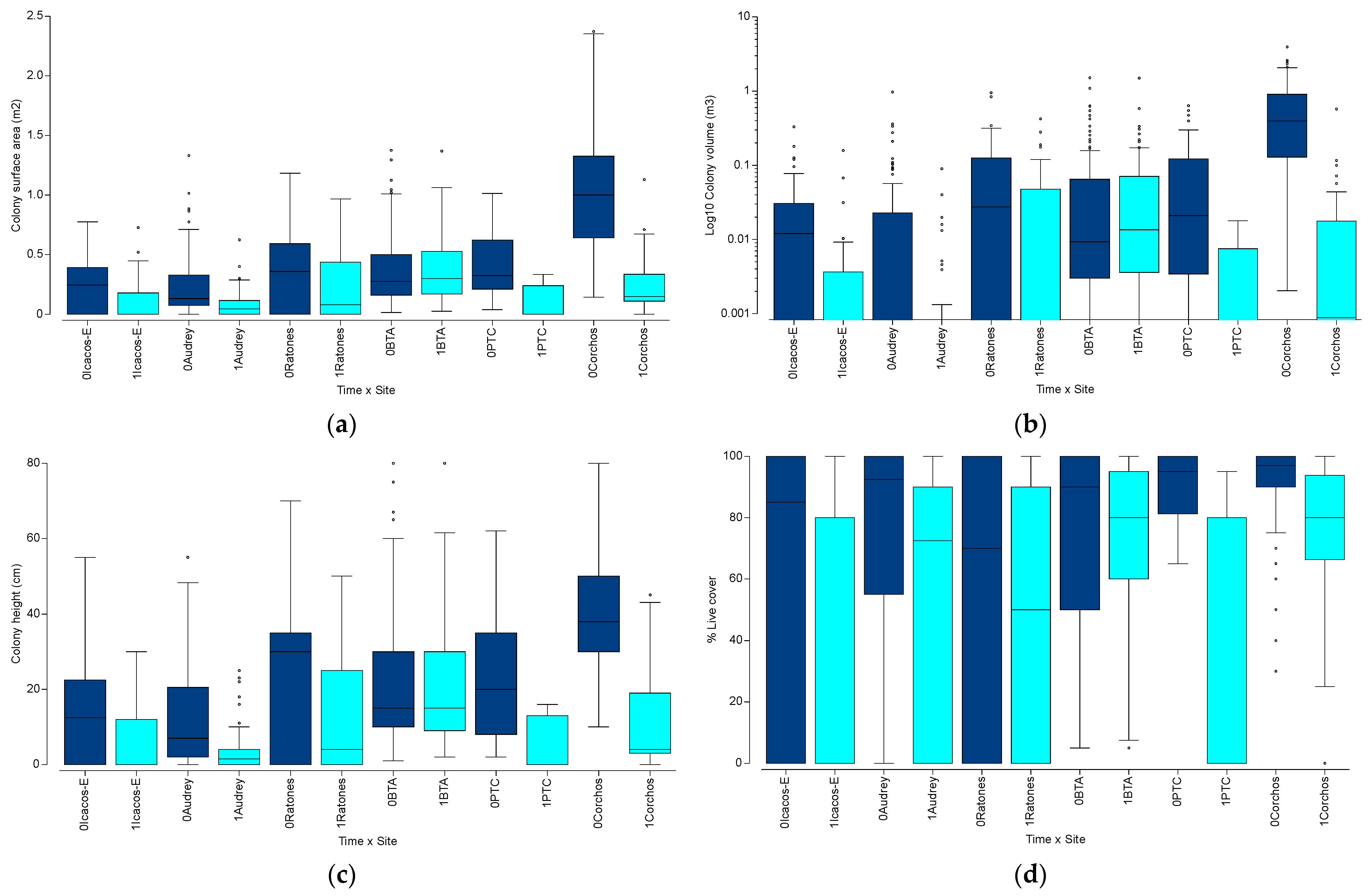
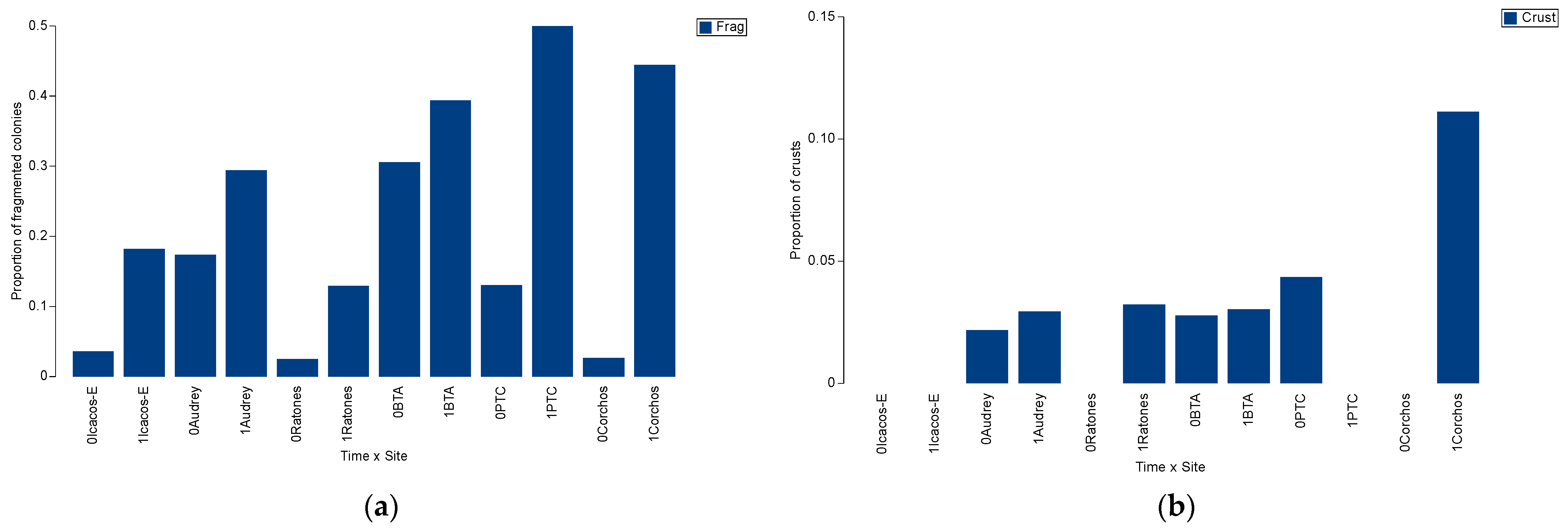
3.2. Spatio-Temporal Variation in Acropora palmata
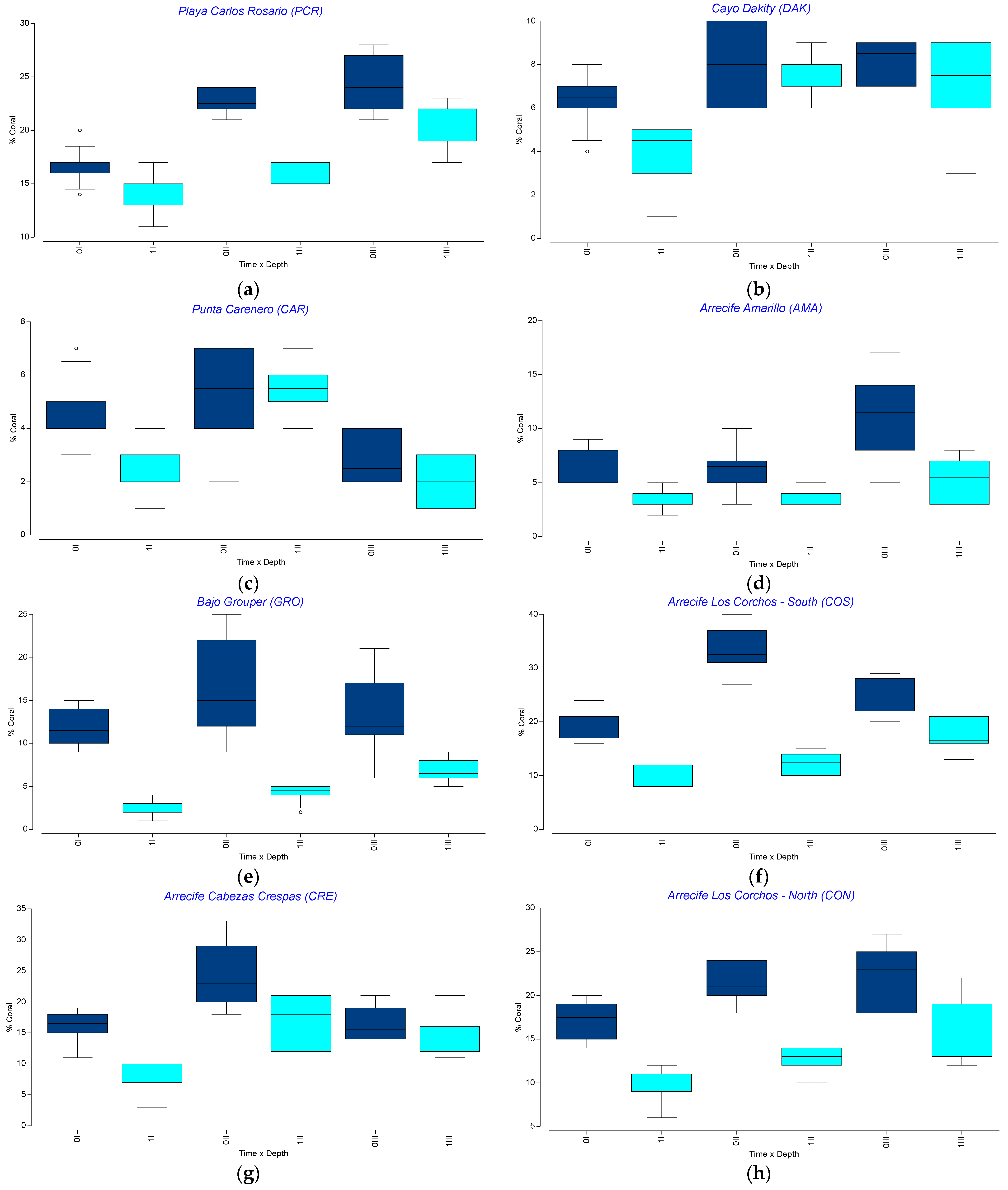
3.3. Spatio-Temporal Variation in Endangered Corals Percent Live Cover
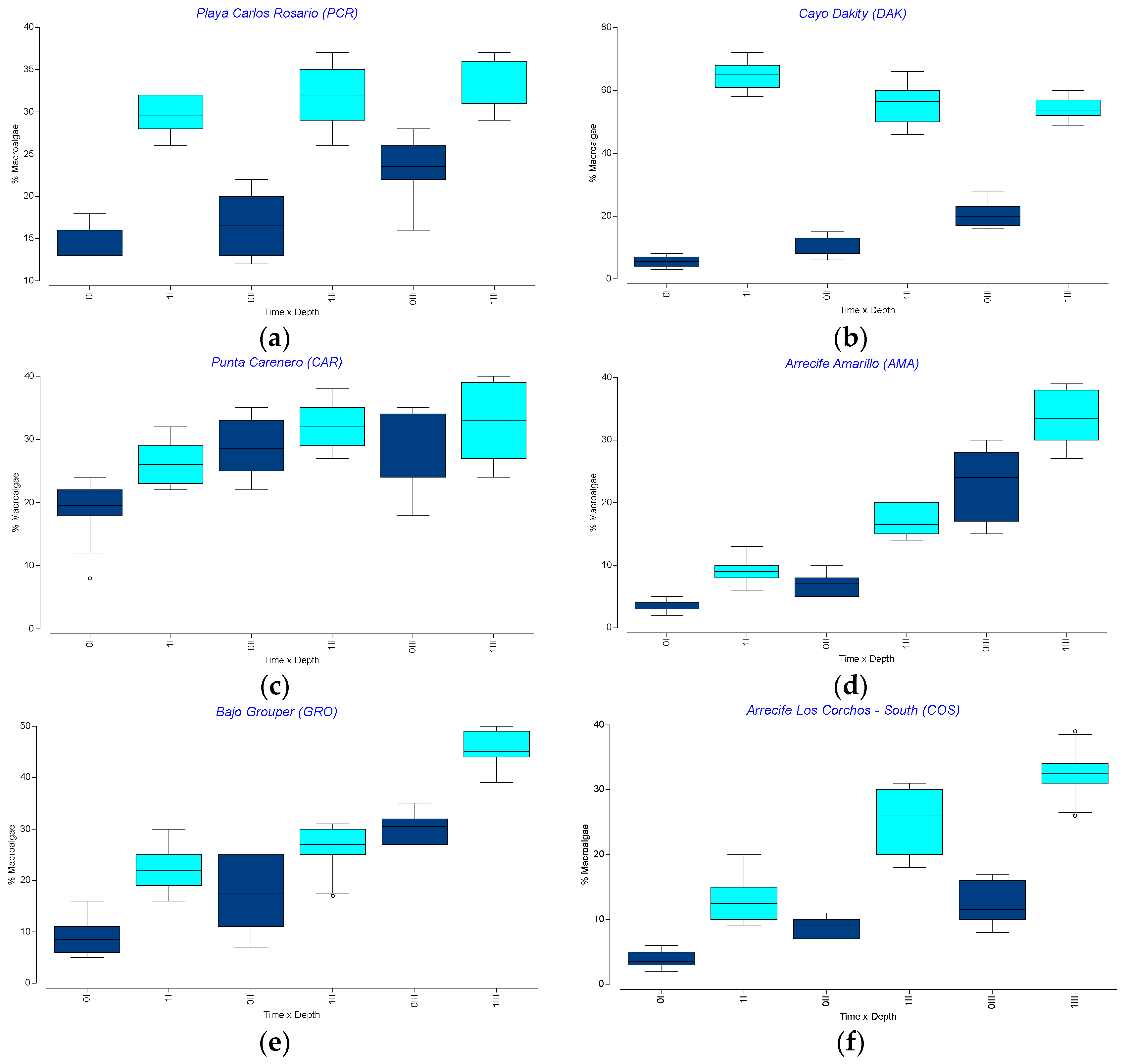
3.4. Spatio-Temporal Variation in the Percent Cover of Other Benthic Components
3.5. Spatio-Temporal Variation in Benthic Community Structure
3.6. Spatio-Temporal Variation in Biodiversity
4. Discussion
4.1. Hurricane Impacts to Endangered Corals
4.2. Can Coral Reefs Recover Rapidly from Major Hurricane Impacts?
4.3. A Future of Stronger Hurricanes and Increasing Damage to Caribbean Coral Reefs?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A









































References
- Haas, A.F.; Fairoz, M.F.; Kelly, L.W.; Nelson, C.E.; Dinsdale, E.A.; Edwards, R.A.; Giles, S.; Hatay, M.; Hisakawa, N.; Knowles, B.; et al. Global microbialization of coral reefs. Nat. Microbiol. 2016, 1, 16042. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Poloczanska, E.S.; Skirving, W.; Dove, S. Coral reef ecosystems under climate change and ocean acidification. Front. Mar. Sci. 2017, 4, 158. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Guillén Bolaños, T.; Bindi, M.; Brown, S.; Camilloni, I.A.; Diedhiou, A.; Djalante, R.; Ebi, K.; et al. The human imperative of stabilizing global climate change at 1.5 C. Science 2019, 365, eaaw6974. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.; Kleypas, J.; Van De Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Griffin, A.; Park, A.; Porter, J.W.; Heron, S.F.; Eakin, C.M.; Berry, B.; Kemp, D.W.; Kemp, K.M.; Lipp, E.K.; et al. Twenty-year record of white pox disease in the Florida Keys: Importance of environmental risk factors as drivers of coral health. Dis. Aquat. Orgs. 2023, 154, 15–31. [Google Scholar] [CrossRef]
- Hayes, N.K.; Walton, C.J.; Gilliam, D.S. Tissue loss disease outbreak significantly alters the Southeast Florida stony coral assemblage. Front. Mar. Sci. 2022, 26, 9. [Google Scholar] [CrossRef]
- Toth, L.T.; Courtney, T.A.; Colella, M.A.; Ruzicka, R.R. Stony coral tissue loss disease accelerated shifts in coral composition and declines in reef accretion potential in the Florida Keys. Front. Mar. Sci. 2023, 10, 1276400. [Google Scholar] [CrossRef]
- Nelson, H.R.; Altieri, A.H. Oxygen: The universal currency on coral reefs. Coral Reefs 2019, 38, 177–198. [Google Scholar] [CrossRef]
- Hughes, D.J.; Alderdice, R.; Cooney, C.; Kühl, M.; Pernice, M.; Voolstra, C.R.; Suggett, D.J. Coral reef survival under accelerating ocean deoxygenation. Nat. Clim. Change 2020, 10, 296–307. [Google Scholar] [CrossRef]
- Mann, M.E.; Emanuel, K.A. Atlantic hurricane trends linked to climate change. Eos Trans. Am. Geophys. Union 2006, 87, 233–241. [Google Scholar] [CrossRef]
- Knutson, T.R.; Sirutis, J.J.; Garner, S.T.; Held, I.M.; Tuleya, R.E. Simulation of the recent multidecadal increase of Atlantic hurricane activity using an 18-km-grid regional model. Bull. Am. Meteor. Soc. 2007, 88, 1549–1565. [Google Scholar] [CrossRef]
- Saunders, M.A.; Lea, A.S. Large contribution of sea surface warming to recent increase in Atlantic hurricane activity. Nature 2008, 451, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Grinsted, A.; Moore, J.C.; Jevrejeva, S. Projected Atlantic hurricane surge threat from rising temperatures. Proc. Nat. Acad. Sci. USA 2013, 110, 5369–5373. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.K.; Willig, M.R.; Hernández-Delgado, E.A. Resistance, resilience, and vulnerability of social-ecological systems to hurricanes in Puerto Rico. Ecosphere 2020, 11, e03159. [Google Scholar] [CrossRef]
- Smith, T.B.; Brandt, M.E.; Calnan, J.M.; Nemeth, R.S.; Blondeau, J.; Kadison, E.; Taylor, M.; Rothenberger, P. Convergent mortality responses of Caribbean coral species to seawater warming. Ecosphere 2013, 4, 1–40. [Google Scholar] [CrossRef]
- De Bakker, D.M.; Van Duyl, F.C.; Bak, R.P.; Nugues, M.M.; Nieuwland, G.; Meesters, E.H. 40 Years of benthic community change on the Caribbean reefs of Curaçao and Bonaire: The rise of slimy cyanobacterial mats. Coral Reefs 2017, 36, 355–367. [Google Scholar] [CrossRef]
- Perry, C.T.; Murphy, G.N.; Kench, P.S.; Smithers, S.G.; Edinger, E.N.; Steneck, R.S.; Mumby, P.J. Caribbean-wide decline in carbonate production threatens coral reef growth. Nature Comm. 2013, 4, 1402. [Google Scholar] [CrossRef]
- Jackson, J.; Donovan, M.; Cramer, K.; Lam, V. Status and Trends of Caribbean Coral Reefs: 1970–2012; Global Coral Reef Monitoring Network; International Union for the Conservation of Nature (IUCN): Gland, Switzerland, 2014; Volume 1–304. [Google Scholar]
- González-Figueroa, M.C.; Hernández-Delgado, E.A. Variación espacial en los patrones de recuperación natural de los arrecifes de coral someros urbanos en Puerto Rico. Persp. Asuntos Amb. 2021, 9, 90–111. [Google Scholar]
- Hernández-Delgado, E.A.; Ortiz-Flores, M.F. The long and winding road of coral reef recovery in the Anthropocene: A case study from Puerto Rico. Diversity 2022, 14, 804. [Google Scholar] [CrossRef]
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Hurricanes and Caribbean coral reefs: Impacts, recovery patterns, and role in long-term decline. Ecology 2005, 86, 174–184. [Google Scholar] [CrossRef]
- Roth, F.; Saalmann, F.; Thomson, T.; Coker, D.J.; Villalobos, R.; Jones, B.H.; Wild, C.; Carvalho, S. Coral reef degradation affects the potential for reef recovery after disturbance. Mar. Environ. Res. 2018, 142, 48–58. [Google Scholar] [CrossRef]
- Speare, K.E.; Adam, T.C.; Winslow, E.M.; Lenihan, H.S.; Burkepile, D.E. Size-dependent mortality of corals during marine heatwave erodes recovery capacity of a coral reef. Glob. Change Biol. 2022, 28, 1342–1358. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.S.; Miller, J. Permanent ‘phase shifts’ or reversible declines in coral cover? Lack of recovery of two coral reefs in St. John, US Virgin Islands. Mar. Ecol. Progr. Ser. 2006, 306, 103–114. [Google Scholar] [CrossRef]
- Miller, J.; Waara, R.; Muller, E.; Rogers, C. Coral bleaching and disease combine to cause extensive mortality on reefs in US Virgin Islands. Coral Reefs 2006, 25, 418. [Google Scholar] [CrossRef]
- Rogers, C. Coral bleaching and disease should not be underestimated as causes of Caribbean coral reef decline. Proc. R. Soc. B Biol. Sci. 2009, 276, 197–198. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Delgado, E.A.; González-Ramos, C.M.; Alejandro-Camis, P.J. Large-scale coral recruitment patterns on Mona Island, Puerto Rico: Evidence of a transitional community trajectory after massive coral bleaching and mortality. Rev. Biol. Trop. 2014, 62, 283–298. [Google Scholar]
- Edmunds, P.J.; Elahi, R. The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecol. Monogr. 2007, 77, 3–18. [Google Scholar] [CrossRef]
- Miller, J.; Muller, E.; Rogers, C.; Waara, R.; Atkinson, A.; Whelan, K.R.; Patterson, M.; Witcher, B. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs 2009, 28, 925–937. [Google Scholar] [CrossRef]
- Hernández-Pacheco, R.; Hernández-Delgado, E.A.; Sabat, A.M. Demographics of bleaching in a major Caribbean reef-building coral: Montastraea annularis. Ecosphere 2011, 2, art9. [Google Scholar] [CrossRef]
- Hughes, T.P. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Anthony, K.R.; Maynard, J.A.; Díaz-Pulido, G.; Mumby, P.J.; Marshall, P.A.; Cao, L.; Hoegh-Guldberg, O. Ocean acidification and warming will lower coral reef resilience. Global Change Biol. 2011, 17, 1798–1808. [Google Scholar] [CrossRef]
- Mumby, P.J. Embracing a world of subtlety and nuance on coral reefs. Coral Reefs 2017, 36, 1003–1011. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implications for conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef]
- Graham, N.A.; Cinner, J.E.; Norström, A.V.; Nyström, M. Coral reefs as novel ecosystems: Embracing new futures. Curr. Opinion Environ. Sust. 2014, 7, 9–14. [Google Scholar] [CrossRef]
- Perry, C.T.; Alvarez-Filip, L. Changing geo-ecological functions of coral reefs in the Anthropocene. Funct. Ecol. 2019, 33, 976–988. [Google Scholar] [CrossRef]
- Williams, G.J.; Graham, N.A. Rethinking coral reef functional futures. Funct. Ecol. 2019, 33, 942–947. [Google Scholar] [CrossRef]
- Norström, A.V.; Nyström, M.; Jouffray, J.B.; Folke, C.; Graham, N.A.; Moberg, F.; Olsson, P.; Williams, G.J. Guiding coral reef futures in the Anthropocene. Front. Ecol. Environ. 2016, 14, 490–498. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Pratchett, M.S.; Morrison, T.H.; Gurney, G.G.; Hughes, T.P.; Álvarez-Romero, J.G.; Day, J.C.; Grantham, R.; Grech, A.; Hoey, A.S.; et al. Coral reef conservation in the Anthropocene: Confronting spatial mismatches and prioritizing functions. Biol. Conserv. 2019, 236, 604–615. [Google Scholar] [CrossRef]
- Mcleod, E.; Anthony, K.R.; Mumby, P.J.; Maynard, J.; Beeden, R.; Graham, N.A.; Heron, S.F.; Hoegh-Guldberg, O.; Jupiter, S.; MacGowan, P.; et al. The future of resilience-based management in coral reef ecosystems. J. Environ. Mgmt. 2019, 233, 291–301. [Google Scholar] [CrossRef]
- Williams, S.M.; García-Sais, J.R. A potential new threat on the coral reefs of Puerto Rico: The recent emergence of Ramicrusta spp. Mar. Ecol. 2020, 41, e12592. [Google Scholar] [CrossRef]
- Souter, D.W.; Linden, O. The health and future of coral reef systems. Ocean Coast. Mgmt. 2000, 43, 657–688. [Google Scholar] [CrossRef]
- Hernández-Delgado, E.A. The emerging threats of climate change on tropical coastal ecosystem services, public health, local economies and livelihood sustainability of small islands: Cumulative impacts and synergies. Mar. Poll. Bull. 2015, 101, 5–28. [Google Scholar] [CrossRef] [PubMed]
- National Marine Fisheries Service. Final Endangered Species Act 4 (d) Regulations for Threatened Elkhorn and Staghorn Corals; National Oceanic and Atmospheric Administration, Southeast Regional Center: St. Petersburg, FL, USA, 2008; pp. 1–24.
- Hernández-Delgado, E.A. Effects of Anthropogenic Stress Gradients in the Structure of Coral Reef Epibenthic and Fish Communities. Ph.D. Thesis, Department Biology, University of Puerto Rico, San Juan, Puerto Rico, 2000; pp. 1–330. [Google Scholar]
- Hernández-Delgado, E.A. Historia natural, caracterización, distribución y estado actual de los arrecifes de coral Puerto Rico. In Biodiversidad de Puerto Rico: Vertebrados Terrestres y Ecosistemas. Serie Historia Natural; Joglar, R.L., Ed.; Editorial Instituto de Cultura Puertorriqueña: San Juan, Puerto Rico, 2005; pp. 281–356. [Google Scholar]
- García, J.R.; Morelock, J.; Castro, R.; Goenaga, C.; Hernández, E. Puerto Rican reefs: Research synthesis, present threats and management perspectives. In Latin American Coral Reefs; Cortés, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 111–130. [Google Scholar]
- García-Sais, J.R.; Appeldoorn, R.; Batista, T.; Bauer, L.; Bruckner, A.; Caldow, C.; Carruba, L.M.; Corredor, J.; Díaz, E.; Lyliestrom, C.; et al. The state of coral reef ecosystems in Puerto Rico. In The State of Coral Reef Ecosystems of the United States and Pacific Freely Associated States; Waddell, E.E., Clarke, A.M., Eds.; National Oceanic and Atmospheric Administration Technical Memorandum National Ocean Service, National Centers for Coastal Ocean Science 73. National Oceanic and Atmospheric Administration/National Centers for Coastal Ocean Science; Center for Coastal Monitoring and Assessment’s Biogeography Team: Silver Spring, MD, USA, 2008; pp. 75–116. [Google Scholar]
- Ballantine, D.L.; Appeldoorn, R.S.; Yoshioka, P.; Weil, E.; Armstrong, R.; García, J.R.; Otero, E.; Pagán, F.; Sherman, C.; Hernández-Delgado, E.A.; et al. Biology and ecology of Puerto Rican coral reefs. In Coral Reefs of the World, Vol. I. Coral Reefs of the USA; Riegl, B.M., Dodge, R.E., Eds.; Springer-Science + Business Media B.V.: Berlin/Heidelberg, Germany, 2008; pp. 375–406. [Google Scholar]
- Ramos-Scharrón, C.E.; Arima, E. Hurricane María’s precipitation signature in Puerto Rico: A conceivable presage of rains to come. Sci. Rep. 2019, 9, 15612. [Google Scholar] [CrossRef] [PubMed]
- Cangialosi, J.P.; Latto, A.S.; Berg, R. National Hurricane Center Tropical Cyclone Report—Hurricane Irma (AL112017); National Hurricane Center: Miami, FL, USA, 2021; pp. 1–111. [Google Scholar]
- Pasch, R.J.; Penny, A.B.; Berg, R. National Hurricane Center Tropical Cyclone Report—Hurricane María (AL152017); National Hurricane Center: Miami, FL, USA, 2023; pp. 1–48. [Google Scholar]
- NOAA. Status of Puerto Rico’s Coral Reefs in the aftermath of Hurricanes Irma and María; Assessment Report Submitted by NOAA to the FEMA Natural and Cultural Resources Recovery Support Function; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2018; pp. 1–37.
- Toledo-Hernández, C.; Ruiz-Diaz, C.P.; Hernández-Delgado, E.A.; Suleimán-Ramos, S.E. Devastation of 15-year old community-based coral farming and reef-restoration sites in Puerto Rico by major hurricanes Irma and María. Caribb. Nat. 2018, 53, 1–6. [Google Scholar]
- Wachnicka, A.; Armitage, A.R.; Zink, I.; Browder, J.; Fourqurean, J.W. Major 2017 hurricanes and their cumulative impacts on coastal waters of the USA and the Caribbean. Estuaries Coasts 2020, 43, 941–942. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Williams, D.E.; Miller, M.W.; Kramer, K.L. Demographic Monitoring Protocols for Threatened Caribbean Acropora spp. Corals; National Oceanic and Atmospheric Administration Technical Memorandum National Marine Fisheries Service-Southeast Fisheries Science Center-543; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2006; pp. 1–91. [Google Scholar]
- Anderson, M.J. Permutational Multivariate Analysis of Variance; Department of Statistics, University of Auckland: Auckland, New Zealand, 2005; Volume 26, pp. 32–46. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Massey University, Auckland and PRIMER-e Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate Dispersion as a Measure of Beta Diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Rogers, C.S.; Suchanek, T.H.; Pecora, F.A. Effects of hurricanes David and Frederic (1979) on shallow Acropora palmata reef communities: St. Croix, US Virgin Islands. Bull. Mar. Sci. 1982, 32, 532–548. [Google Scholar]
- Rogers, C.S.; McLain, L.N.; Tobias, C.R. Effects of Hurricane Hugo (1989) on a coral reef in St. John, USVI. Mar. Ecol. Progr. Ser. 1991, 78, 189–199. [Google Scholar] [CrossRef]
- Rogers, C.S. Hurricanes and coral reefs: The intermediate disturbance hypothesis revisited. Coral Reefs 1993, 12, 127–137. [Google Scholar] [CrossRef]
- Kjerfve, B.; Magill, K.E.; Porter, J.W.; Woodley, J.D. Hindcasting of hurricane characteristics and observed storm damage on a fringing reef, Jamaica, West Indies. J. Mar. Res. 1986, 44, 119–148. [Google Scholar] [CrossRef]
- Woodley, J.D.; Chornesky, E.A.; Clifford, P.A.; Jackson, J.B.; Kaufman, L.S.; Knowlton, N.; Lang, J.C.; Pearson, M.P.; Porter, J.W.; Rooney, M.C.; et al. Hurricane Allen’s impact on Jamaican coral reefs. Science 1981, 214, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Woodley, J.D. The incidence of hurricanes on the north coast of Jamaica since 1870: Are the classic reef descriptions atypical? Hydrobiologia 1992, 247, 133–138. [Google Scholar] [CrossRef]
- Liddell, W.D.; Ohlhorst, S.L. Patterns of reef community structure, North Jamaica. Bull. Mar. Sci. 1987, 40, 311–329. [Google Scholar]
- Aronson, R.B.; Precht, W.F. Evolutionary paleoecology of Caribbean coral reefs. In Evolutionary Paleoecology: The Ecological Context of Macroevolutionary Change; Columbia University Press: New York, NY, USA, 2001; pp. 171–234. [Google Scholar]
- Mumby, P.J.; Hastings, A.; Edwards, H.J. Thresholds and the resilience of Caribbean coral reefs. Nature 2007, 450, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Mumby, P.J.; Foster, N.L.; Fahy, E.A. Patch dynamics of coral reef macroalgae under chronic and acute disturbance. Coral Reefs 2005, 24, 681–692. [Google Scholar] [CrossRef]
- Gleason, A.C.; Lirman, D.; Williams, D.; Gracias, N.R.; Gintert, B.E.; Madjidi, H.; Reid, R.P.; Boynton, C.G.; Negahdaripour, S.; Miller, M.; et al. Documenting hurricane impacts on coral reefs using two-dimensional video-mosaic technology. Mar. Ecol. 2007, 28, 254–258. [Google Scholar] [CrossRef]
- Bythell, J.C.; Hillis-Starr, Z.M.; Rogers, C.S. Local variability but landscape stability in coral reef communities following repeated hurricane impacts. Mar. Ecol. Progr. Ser. 2000, 204, 93–100. [Google Scholar] [CrossRef]
- Highsmith, R.C.; Riggs, A.C.; D’Antonio, C.M. Survival of hurricane-generated coral fragments and a disturbance model of reef calcification/growth rates. Oecologia 1980, 46, 322–329. [Google Scholar] [CrossRef]
- Edmunds, P.J. The demography of hurricane effects on two coral populations differing in dynamics. Ecosphere 2019, 10, e02836. [Google Scholar] [CrossRef]
- Mumby, P.J. Bleaching and hurricane disturbances to populations of coral recruits in Belize. Mar. Ecol. Progr. Ser. 1999, 190, 27–35. [Google Scholar] [CrossRef]
- Stoddart, D.R. Catastrophic storm effects on the British Honduras reefs and cays. Nature 1962, 196, 512–515. [Google Scholar] [CrossRef]
- Stoddart, D.R. Effects of Hurricane Hattie on the British Honduras reefs and cays, 30–31 October 1961. Atoll Res. Bull. 1963, 95, 1–142. [Google Scholar] [CrossRef]
- Lirman, D.; Fong, P. Sequential storms cause zone-specific damage on a reef in the northern Florida reef tract: Evidence from Hurricane Andrew and the 1993 Storm of the Century. Fla. Sci. 1996, 59, 50–64. [Google Scholar]
- Stoddart, D.R. Post-hurricane changes on the British Honduras reefs and cays: Re-survey of 1965. Atoll Res. Bull. 1969, 131, 50–64. [Google Scholar] [CrossRef]
- Stoddart, D.R. Coral reefs and islands and catastrophic storms. In Applied Coastal Geomorphology; Palgrave Macmillan: London, UK, 1971; pp. 155–197. [Google Scholar]
- Williams, A.H. The effects of Hurricane Allen on back reef populations of Discovery Bay, Jamaica. J. Exp. Mar. Biol. Ecol. 1984, 75, 233–243. [Google Scholar] [CrossRef]
- Knowlton, N.; Lang, J.C.; Rooney, C.M.; Clifford, P. Evidence for delayed mortality in hurricane-damaged Jamaican staghorn corals. Nature 1981, 294, 251–252. [Google Scholar] [CrossRef]
- Knowlton, N.; Lang, J.C.; Keller, B.D. Case study of natural population collapse: Post-hurricane predation on Jamaican staghorn corals. Smith. Contrib. Mar. Sci. 1990, 31, 1–125. [Google Scholar] [CrossRef]
- Kaufman, L.S. Effects of Hurricane Allen on reef fish assemblages near Discovery Bay, Jamaica. Coral Reefs 1983, 2, 43–47. [Google Scholar] [CrossRef]
- Álvarez-Filip, L.; Millet-Encalada, M.; Reyes-Bonilla, H. Impact of Hurricanes Emily and Wilma on the coral community of Cozumel Island, Mexico. Bull. Mar. Sci. 2009, 84, 295–306. [Google Scholar]
- Bernal-Sotelo, K.; Acosta, A.; Cortés, J. Decadal change in the population of Dendrogyra cylindrus (Scleractinia: Meandrinidae) in old providence and St. Catalina Islands, Colombian Caribbean. Front. Mar. Sci. 2019, 5, 513. [Google Scholar] [CrossRef]
- Foster, N.L.; Baums, I.B.; Sanchez, J.A.; Paris, C.B.; Chollett, I.; Agudelo, C.L.; Vermeij, M.J.; Mumby, P.J. Hurricane-driven patterns of clonality in an ecosystem engineer: The Caribbean coral Montastraea annularis. PLoS ONE 2013, 8, e53283. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Filip, L.; Dulvy, N.K.; Gill, J.A.; Côté, I.M.; Watkinson, A.R. Flattening of Caribbean coral reefs: Region-wide declines in architectural complexity. Proc. R. Soc. B Biol. Sci. 2009, 276, 3019–3025. [Google Scholar] [CrossRef] [PubMed]
- Gochfeld, D.J.; Olson, J.B.; Chaves-Fonnegra, A.; Smith, T.B.; Ennis, R.S.; Brandt, M.E. Impacts of hurricanes Irma and Maria on coral reef sponge communities in St. Thomas, US Virgin Islands. Estuaries Coasts 2020, 43, 1235–1247. [Google Scholar] [CrossRef]
- Doropoulos, C.; Roff, G.; Zupan, M.; Nestor, V.; Isechal, A.L.; Mumby, P.J. Reef-scale failure of coral settlement following typhoon disturbance and macroalgal bloom in Palau, Western Pacific. Coral Reefs 2014, 33, 613–623. [Google Scholar] [CrossRef]
- Gouezo, M.; Golbuu, Y.; Van Woesik, R.; Rehm, L.; Koshiba, S.; Doropoulos, C. Impact of two sequential super typhoons on coral reef communities in Palau. Mar. Ecol. Progr. Ser. 2015, 540, 73–85. [Google Scholar] [CrossRef]
- Anticamara, J.A.; Go, K.T.B. Impacts of super-typhoon Yolanda on Philippine reefs and communities. Reg. Environ. Change 2017, 17, 703–713. [Google Scholar] [CrossRef]
- Yang, H.; Yu, K.; Zhao, M.; Shi, Q.; Tao, S.; Yan, H.; Chen, T.; Liu, G. Impact on the coral reefs at Yongle Atoll, Xisha Islands, South China Sea from a strong typhoon direct sweep: Wutip, September 2013. J. Asian Earth Sci. 2015, 114, 457–466. [Google Scholar] [CrossRef]
- Ogg, J.G.; Koslow, J.A. The impact of typhoon Pamela (1976) on Guam’s coral reefs and beaches. Pac. Sci. 1978, 32, 105–118. [Google Scholar]
- Harmelin-Vivien, M.L.; Laboute, P. Catastrophic impact of hurricanes on atoll outer reef slopes in the Tuamotu (French Polynesia). Coral Reefs 1986, 5, 55–62. [Google Scholar] [CrossRef]
- White, K.N.; Ohara, T.; Fujii, T.; Kawamura, I.; Mizuyama, M.; Montenegro, J.; Shikiba, H.; Naruse, T.; McClelland, T.; Denis, V.; et al. Typhoon damage on a shallow mesophotic reef in Okinawa, Japan. PeerJ 2013, 1, e151. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, N.; Chabanet, P.; Le Pape, O. Cyclone effects on coral reef habitats in New Caledonia (South Pacific). Coral Reefs 2010, 29, 445–453. [Google Scholar] [CrossRef]
- White, K.N.; Weinstein, D.K.; Ohara, T.; Denis, V.; Montenegro, J.; Reimer, J.D. Shifting communities after typhoon damage on an upper mesophotic reef in Okinawa, Japan. PeerJ 2017, 5, e3573. [Google Scholar] [CrossRef] [PubMed]
- Vila-Concejo, A.; Duce, S.; Nagao, M.; Nakashima, Y.; Ito, M.; Fujita, K.; Kan, H. Typhoon waves on coral reefs. Coast. Dyn. 2017, 263, 697–701. [Google Scholar]
- Hongo, C.; Kawamata, H.; Goto, K. Catastrophic impact of typhoon waves on coral communities in the Ryukyu Islands under global warming. J. Geophys. Res. Biogeosci. 2012, 117, G02029. [Google Scholar] [CrossRef]
- Hernández, W. Quantifying the Effects of Hurricanes Irma and Maria on Coastal Water Quality, Habitats, and Resources in Puerto Rico Using Moderate and High Resolution Satellite Sensors; Technical Report; National Oceanic and Atmospheric Administration Restoration Center and Coral Reef Conservation Program: Silver Spring, MD, USA, 2018; pp. 1–40.
- Edmunds, P.J.; Tsounis, G.; Boulon, R.; Bramanti, L. Acute effects of back-to-back hurricanes on the underwater light regime of a coral reef. Mar. Biol. 2019, 166, 20. [Google Scholar] [CrossRef]
- Hernández-Delgado, E.A.; Mercado-Molina, A.E.; Alejandro-Camis, P.J.; Candelas-Sánchez, F.; Fonseca-Miranda, J.S.; González-Ramos, C.M.; Guzmán-Rodríguez, R.; Mège, P.; Montañez-Acuña, A.A.; Olivo-Maldonado, I.; et al. Community-based coral reef rehabilitation in a changing climate: Lessons learned from hurricanes, extreme rainfall, and changing land use impacts. Open J. Ecol. 2014, 4, 918–944. [Google Scholar] [CrossRef]
- Ramos-Scharrón, C.E.; Amador, J.M.; Hernández-Delgado, E.A. An interdisciplinary erosion mitigation approach for coral reef protection—A case study from the eastern Caribbean. In Marine Ecosystems; Cruzado, A., Ed.; Intech Publications: Rijeka, Croatia, 2012; pp. 127–160. [Google Scholar]
- Ramos-Scharrón, C.; Torres-Pulliza, D.; Hernández-Delgado, E.A. 2015. Watershed- and island-scale land cover changes in Puerto Rico (1930s-2004) and their potential effects on coral reef ecosystems. Sci. Total Environ. 2015, 506–507, 241–251. [Google Scholar] [CrossRef]
- Otaño-Cruz, A.; Montañez-Acuña, A.A.; Torres-López, V.I.; Hernández-Figueroa, E.M.; Hernández-Delgado, E.A. Effects of changing weather, oceanographic conditions, and land uses on spatio-temporal variation of sedimentation dynamics along near-shore coral reefs. Front. Mar. Sci. 2017, 4, 249. [Google Scholar] [CrossRef]
- Otaño-Cruz, A.; Montañez-Acuña, A.A.; Benson, E.; Cuevas, E.P.; Ortiz, J.; Hernández-Delgado, E.A. Response of near-shore coral reefs benthic communities to changes of sedimentation dynamics and environmental conditions. Front. Mar. Sci. 2019, 6, 551. [Google Scholar] [CrossRef]
- Lugo, A.E. Effects and outcomes of Caribbean hurricanes in a climate change scenario. Sci. Total Environ. 2000, 262, 243–251. [Google Scholar] [CrossRef]
- Heron, S.; Morgan, J.; Eakin, M.A.; Skirving, W. Hurricanes and their effects on coral reefs. In Status of Caribbean Coral Reefs After Bleaching and Hurricanes in 2005; Wilkinson, C., Souter, D., Eds.; Global Coral Reef Monitoring Network, and Reef and Rainforest Research Centre: Townsville, AU, USA, 2008; pp. 31–36. [Google Scholar]
- Hernández-Delgado, E.A.; Toledo-Hernández, C.; Ruíz-Díaz, C.P.; Gómez-Andújar, N.; Medina-Muñiz, J.L.; Canals-Silander, M.F.; Suleimán-Ramos, S.E. Hurricane impacts and the resilience of the invasive sea vine, Halophila stipulacea: A case study from Puerto Rico. Estuaries Coasts 2020, 43, 1263–1283. [Google Scholar] [CrossRef]
- Wilson, S.S.; Furman, B.T.; Hall, M.O.; Fourqurean, J.W. Assessment of Hurricane Irma impacts on South Florida seagrass communities using long-term monitoring programs. Estuaries Coasts 2020, 43, 1119–1132. [Google Scholar] [CrossRef]
- Roff, G.; Chollett, I.; Doropoulos, C.; Golbuu, Y.; Steneck, R.S.; Isechal, A.L.; van Woesik, R.; Mumby, P.J. Exposure-driven macroalgal phase shift following catastrophic disturbance on coral reefs. Coral Reefs 2015, 34, 715–725. [Google Scholar] [CrossRef]
- Donahue, S.; Acosta, A.; Akins, L.; Ault, J.; Bohnsack, J.; Boyer, J.; Callahan, M.; Causey, B.; Cox, C.; Delaney, J.; et al. The state of coral reef ecosystems of the Florida Keys. In The State of Coral Reef Ecosystems of the United States and Pacific Freely Associated States; NOAA Technical Mem. NOS NCCOS 73; NOAA/NCCOS Center for Coastal Monitoring and Assessment’s Biogeography Team: Silver Spring, MD, USA, 2008; pp. 161–187. [Google Scholar]
- Lugo-Fernández, A.; Gravois, M. Understanding impacts of tropical storms and hurricanes on submerged bank reefs and coral communities in the northwestern Gulf of Mexico. Cont. Shelf Res. 2010, 30, 1226–1240. [Google Scholar] [CrossRef]
- Zimmer, B.; Duncan, L.; Deis, D.R.; Robbart, M.L.; Aronson, R.B.; Deslarzes, K.J.; Precht, W.F.; Sinclair, J.; Hickerson, E.L.; Schmah, G.P.; et al. Post-hurricane assessment of reefs and banks in the NW Gulf of Mexico. In Proceedings of the 11th International Coral Reef Symposium, Fort Lauderdale, FL, USA, 7–11 July 2008; Session 23. pp. 1–5. [Google Scholar]
- Fong, P.; Lirman, D. Hurricanes cause population expansion of the branching coral Acropora palmata (Scleractinia): Wound healing and growth patterns of asexual recruits. Mar. Ecol. 1995, 16, 317–335. [Google Scholar] [CrossRef]
- Lirman, D.; Fong, P. Patterns of damage to the branching coral Acropora palmata following Hurricane Andrew: Damage and survivorship of hurricane-generated asexual recruits. J. Coast. Res. 1997, 1, 67–72. [Google Scholar]
- Lirman, D. Fragmentation in the branching coral Acropora palmata (Lamarck): Growth, survivorship, and reproduction of colonies and fragments. J. Exp. Mar. Biol. Ecol. 2000, 251, 41–57. [Google Scholar] [CrossRef]
- Rasser, M.; Riegl, B. Holocene coral reef rubble and its binding agents. Coral Reefs 2002, 21, 57–72. [Google Scholar] [CrossRef]
- Shannon, A.M.; Power, H.E.; Webster, J.M.; Vila-Concejo, A. Evolution of coral rubble deposits on a reef platform as detected by remote sensing. Remote Sens. 2013, 5, 1–18. [Google Scholar] [CrossRef]
- Cameron, C.M.; Pausch, R.E.; Miller, M.W. Coral recruitment dynamics and substrate mobility in a rubble-dominated back reef habitat. Bull. Mar. Sci. 2016, 92, 123–136. [Google Scholar] [CrossRef]
- Kenyon, T.M.; Doropoulos, C.; Dove, S.; Webb, G.E.; Newman, S.P.; Sim, C.W.; Arzan, M.; Mumby, P.J. The effects of rubble mobilisation on coral fragment survival, partial mortality and growth. J. Exp. Mar. Biol. Ecol. 2020, 533, 151467. [Google Scholar] [CrossRef]
- Kenyon, T.M.; Doropoulos, C.; Wolfe, K.; Webb, G.E.; Dove, S.; Harris, D.; Mumby, P.J. Coral rubble dynamics in the Anthropocene and implications for reef recovery. Limnol. Oceanogr. 2023, 68, 110–147. [Google Scholar] [CrossRef]
- Vroom, P.; Walters, L.; Beach, K.; Coyer, J.; Smith, J.; Abgrall, M.J.; Byron, D.; DeAngelis, K.; Konar, B.; Liss, J.; et al. Hurricane-induced propagation and rapid regrowth of the weedy brown alga Dictyota in the Florida Keys. Fla. Sci. 2005, 1, 161–174. [Google Scholar]
- Wolfe, K.; Kenyon, T.M.; Mumby, P.J. The biology and ecology of coral rubble and implications for the future of coral reefs. Coral Reefs 2021, 40, 1769–1806. [Google Scholar] [CrossRef]
- Madden, I.A.; Mariwala, A.; Lindhart, M.; Narayan, S.; Arkema, K.K.; Beck, M.W.; Baker, J.W.; Suckale, J. Quantifying the fragility of coral reefs to hurricane impacts: A case study of the Florida Keys and Puerto Rico. Environ. Res. Lett. 2023, 18, 024034. [Google Scholar] [CrossRef]
- Weil, E.; Hernández-Delgado, E.A.; González, M.; Williams, S.; Suleimán-Ramos, S.E.; Figuerola, M.; Metz-Estrella, T. Spread of the new coral disease “SCTLD” into the Caribbean: Implications for Puerto Rico. Reef Enc. 2019, 34, 38–43. [Google Scholar]
- Brandt, M.E.; Ennis, R.S.; Meiling, S.S.; Townsend, J.; Cobleigh, K.; Glahn, A.; Quetel, J.; Brandtneris, V.; Henderson, L.M.; Smith, T.B. The emergence and initial impact of stony coral tissue loss disease (SCTLD) in the United States Virgin Islands. Front. Mar. Sci. 2021, 8, 715329. [Google Scholar] [CrossRef]
- Williams, S.M.; García-Sais, J.; Sabater-Clavell, J. Prevalence of stony coral tissue loss disease at el seco, a mesophotic reef system off Vieques Island, Puerto Rico. Front. Mar. Sci. 2021, 8, 668669. [Google Scholar] [CrossRef]
- Hallam, S.; Marsh, R.; Josey, S.A.; Hyder, P.; Moat, B.; Hirschi, J.J. Ocean precursors to the extreme Atlantic 2017 hurricane season. Nat. Comm. 2019, 10, 896. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, K. Assessing the present and future probability of Hurricane Harvey’s rainfall. Proc. Nat. Acad. Sci. USA 2017, 114, 12681–12684. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Cheng, L.; Jacobs, P.; Zhang, Y.; Fasullo, J. Hurricane Harvey links to ocean heat content and climate change adaptation. Earth’s Future 2018, 6, 730–744. [Google Scholar] [CrossRef]
- Guzmán, O.; Jiang, H. Heavier inner-core rainfall of major hurricanes in the North Atlantic Basin than in other global basins. J. Clim. 2021, 34, 5707–5721. [Google Scholar] [CrossRef]
- Emanuel, K. Atlantic tropical cyclones downscaled from climate reanalyses show increasing activity over past 150 years. Nat. Comm. 2021, 12, 7027. [Google Scholar] [CrossRef]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled impact of anthropogenic warming on the frequency of intense Atlantic hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, L.; Hodges, K.I.; Esch, M.; Keenlyside, N.; Kornblueh, L.; Luo, J.J.; Yamagata, T. How may tropical cyclones change in a warmer climate? Tellus A 2007, 59, 539–561. [Google Scholar] [CrossRef]
- Hill, K.A.; Lackmann, G.M. The impact of future climate change on TC intensity and structure: A downscaling approach. J. Clim. 2011, 24, 4644–4661. [Google Scholar] [CrossRef]
- Staid, A.; Guikema, S.D.; Nateghi, R.; Quiring, S.M.; Gao, M.Z. Simulation of tropical cyclone impacts to the US power system under climate change scenarios. Clim. Change 2014, 127, 535–546. [Google Scholar] [CrossRef]
- Walsh, K.J.; McBride, J.L.; Klotzbach, P.J.; Balachandran, S.; Camargo, S.J.; Holland, G.; Knutson, T.R.; Kossin, J.P.; Lee, T.C.; Sobel, A.; et al. Tropical cyclones and climate change. Wiley Int. Rev. Clim. Change 2016, 7, 65–89. [Google Scholar] [CrossRef]
- Knutson, T.R.; Chung, M.V.; Vecchi, G.; Sun, J.; Hsieh, T.L.; Smith, A.J. Climate change is probably increasing the intensity of tropical cyclones. In Critical Issues in Climate Change Science, Science Brief Review; No. 4570334; Tyndall Centre, University of East Anglia: Norwich, UK, 2021. [Google Scholar] [CrossRef]
- Landsea, C.W.; Vecchi, G.A.; Bengtsson, L.; Knutson, T.R. Impact of duration thresholds on Atlantic tropical cyclone counts. J. Clim. 2010, 23, 2508–2519. [Google Scholar] [CrossRef]
- Holland, G.; Bruyère, C.L. Recent intense hurricane response to global climate change. Clim. Dyn. 2014, 42, 617–627. [Google Scholar] [CrossRef]
- Tsuboki, K.; Yoshioka, M.K.; Shinoda, T.; Kato, M.; Kanada, S.; Kitoh, A. Future increase of supertyphoon intensity associated with climate change. Geophys. Res. Lett. 2015, 42, 646–652. [Google Scholar] [CrossRef]
- Liu, M.; Vecchi, G.A.; Smith, J.A.; Knutson, T.R. Causes of large projected increases in hurricane precipitation rates with global warming. NPJ Clim. Atmos. Sci. 2019, 2, 38. [Google Scholar] [CrossRef]
- Huprikar, A.; Stansfield, A.; Reed, K. A storyline analysis of Hurricane Irma’s precipitation under various levels of climate warming. Environ. Res. Lett. 2023, 19, 014004. [Google Scholar] [CrossRef]
- Ranson, M.; Kousky, C.; Ruth, M.; Jantarasami, L.; Crimmins, A.; Tarquinio, L. Tropical and extratropical cyclone damages under climate change. Clim. Change 2014, 127, 227–241. [Google Scholar] [CrossRef]
- Gettelman, A.; Bresch, D.N.; Chen, C.C.; Truesdale, J.E.; Bacmeister, J.T. Projections of future tropical cyclone damage with a high-resolution global climate model. Clim. Change 2018, 146, 575–585. [Google Scholar] [CrossRef]
- Bjarnadottir, S.; Li, Y.; Stewart, M.G. A probabilistic-based framework for impact and adaptation assessment of climate change on hurricane damage risks and costs. Struct. Saf. 2011, 33, 173–185. [Google Scholar] [CrossRef]
- Knutson, T.R.; Tuleya, R.E. Impact of CO 2-induced warming on simulated hurricane intensity and precipitation: Sensitivity to the choice of climate model and convective parameterization. J. Clim. 2004, 17, 3477–3495. [Google Scholar] [CrossRef]
- Balaguru, K.; Foltz, G.R.; Leung, L.R. Increasing magnitude of hurricane rapid intensification in the central and eastern tropical Atlantic. Geophys. Res. Lett. 2018, 45, 4238–4247. [Google Scholar] [CrossRef]
- Balaguru, K.; Foltz, G.R.; Leung, L.R.; Xu, W.; Kim, D.; López, H.; West, R. Increasing hurricane intensification rate near the US Atlantic coast. Geophys. Res. Lett. 2022, 49, e2022GL099793. [Google Scholar] [CrossRef]
- Bhatia, K.T.; Vecchi, G.A.; Knutson, T.R.; Murakami, H.; Kossin, J.; Dixon, K.W.; Whitlock, C.E. Recent increases in tropical cyclone intensification rates. Nat. Comm. 2019, 10, 635. [Google Scholar] [CrossRef]
- Bhatia, K.; Baker, A.; Yang, W.; Vecchi, G.; Knutson, T.; Murakami, H.; Kossin, J.; Hodges, K.; Dixon, K.; Bronselaer, B.; et al. A potential explanation for the global increase in tropical cyclone rapid intensification. Nat. Comm. 2022, 13, 6626. [Google Scholar] [CrossRef] [PubMed]
- Haarsma, R.J.; Hazeleger, W.; Severijns, C.; De Vries, H.; Sterl, A.; Bintanja, R.; Van Oldenborgh, G.J.; Van Den Brink, H.W. More hurricanes to hit western Europe due to global warming. Geophys. Res. Lett. 2013, 40, 1783–1788. [Google Scholar] [CrossRef]
- Studholme, J.; Fedorov, A.V.; Gulev, S.K.; Emanuel, K.; Hodges, K. Poleward expansion of tropical cyclone latitudes in warming climates. Nat. Geosci. 2022, 15, 14–28. [Google Scholar] [CrossRef]
- Mei, W.; Primeau, F.; McWilliams, J.C.; Pasquero, C. Sea surface height evidence for long-term warming effects of tropical cyclones on the ocean. Proc. Nat. Acad. Sci. USA 2013, 110, 15207–15210. [Google Scholar] [CrossRef]
- Kossin, J.P. A global slowdown of tropical-cyclone translation speed. Nature 2018, 558, 104–107. [Google Scholar] [CrossRef]
- Gründemann, G.J.; van de Giesen, N.; Brunner, L.; van der Ent, R. Rarest rainfall events will see the greatest relative increase in magnitude under future climate change. Commun. Earth Environ. 2022, 3, 235. [Google Scholar] [CrossRef]
- Reed, K.A.; Wehner, M.F.; Zarzycki, C.M. Attribution of 2020 hurricane season extreme rainfall to human-induced climate change. Nat. Commun. 2022, 13, 1905. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Smith, J.A.; Villarini, G.; Marchok, T.P.; Baeck, M.L. Modeling extreme rainfall, winds, and surge from Hurricane Isabel (2003). Weather Forecast. 2010, 25, 1342–1361. [Google Scholar] [CrossRef]
- Lin, N.; Goni, G.J.; Knaff, J.A.; Forbes, C.; Ali, M.M. Ocean heat content for tropical cyclone intensity forecasting and its impact on storm surge. Nat. Hazards 2013, 66, 1481–1500. [Google Scholar] [CrossRef]
- Lin, N.; Kopp, R.E.; Horton, B.P.; Donnelly, J.P. Hurricane Sandy’s flood frequency increasing from year 1800 to 2100. Proc. Nat. Acad. Sci. USA 2016, 113, 12071–12075. [Google Scholar] [CrossRef]
- Lin, N.; Lane, P.; Emanuel, K.A.; Sullivan, R.M.; Donnelly, J.P. Heightened hurricane surge risk in northwest Florida revealed from climatological-hydrodynamic modeling and paleorecord reconstruction. J. Geophys. Res. Atmos. 2014, 119, 8606–8623. [Google Scholar] [CrossRef]
- Collier, C.; Ruzicka, R.; Banks, K.; Barbieri, L.; Beal, J.; Bingham, D.; Bohnsack, J.A.; Brooke, S.; Craig, N.; Fisher, L.E.; et al. The state of coral reef ecosystems of southeast Florida. In The State of Coral Reef Ecosystems of the United States and Pacific Freely Associated States; NOAA Technical Mem. NOS NCCOS 73; NOAA/NCCOS Center for Coastal Monitoring and Assessment’s Biogeography Team: Silver Spring, MD, USA, 2008; pp. 131–159. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
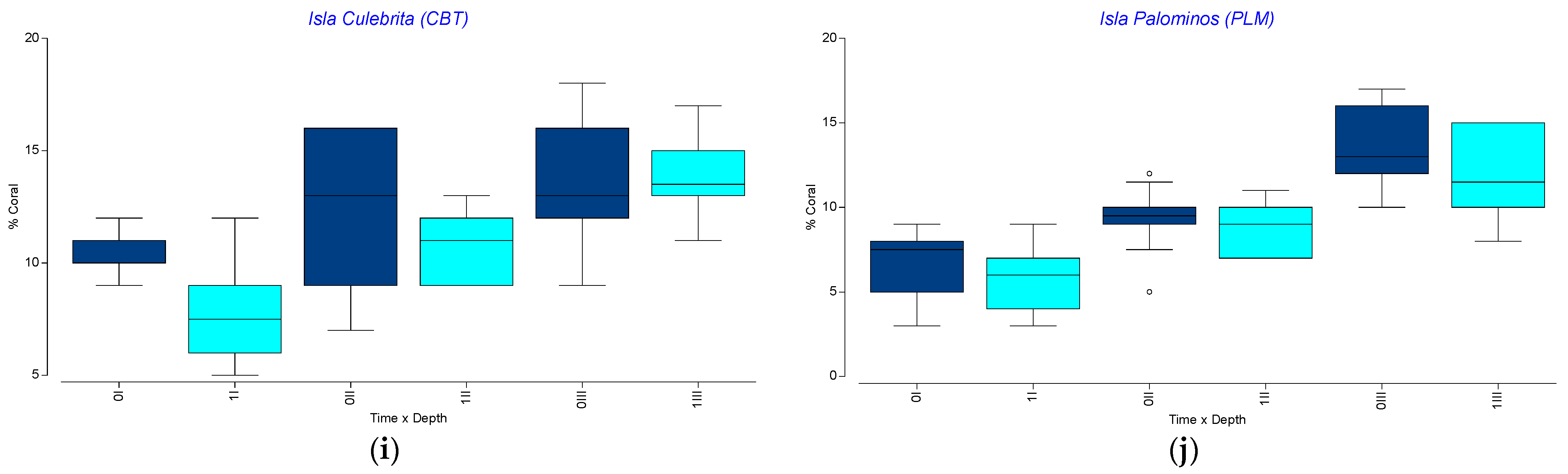{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Acronym | Latitude | Longitude |
|---|---|---|---|
| Cayo Icacos | ICA | 18.379958° | −65.585074° |
| Audrey Rock | AUD | 18.379338° | −65.583138° |
| Cayo Ratones | RAT | 18.380443° | −65.581537° |
| Palominos Island | PLM | 18.353366° | −65.570911° |
| Bahía Tamarindo | BTA | 18.314467° | −65.317659° |
| Punta Tamarindo Chico | PTC | 18.309368° | −65.317045° |
| Playa Carlos Rosario | PCR | 18.327083° | −65.330703° |
| Cayo Dákity | DAK | 18.284020° | −65.278355° |
| Punta Carenero | CAR | 18.293450° | −65.274178° |
| Bajo Amarillo | AMA | 18.279045° | −65.273172° |
| Bajo Grouper | GRO | 18.285689° | −65.270431° |
| Los Corchos-South | COS | 18.283938° | −65.245307° |
| Cabezas Crespás | CRE | 18.291187° | −65.233909° |
| Los Corchos-North | CON | 18.303623° | −65.230626° |
| Culebrita Island | CBT | 18.315294° | −65.233359° |
| Source 1 | Rho | p |
|---|---|---|
| ATM [CO2] | 0.105 | 0.0187 |
| Annual increment [CO2] | −0.042 | 0.7771 |
| Temperature anomaly L-O | 0.053 | 0.1460 |
| NAO Index | −0.027 | 0.6674 |
| Ocean Heat Content | 0.110 | 0.0181 |
| Thermosteric component SLC | 0.104 | 0.0234 |
| Source 1 | df | Surface Area | Colony Volume | Colony Height | % Live Cover |
|---|---|---|---|---|---|
| Time | 1 | 4.09 | 3.43 | 8.15 | 7.26 |
| 0.1063 | 0.1291 | 0.0307 | 0.0369 | ||
| Region | 1 | 4.09 | 2.69 | 9.98 | 11.05 |
| 0.1525 | 0.177 | 0.0081 | 0.0044 | ||
| Site (Region) | 4 | 11 | 11.84 | 6.75 | 7 |
| <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||
| Time × Region | 1 | 0.6 | 0.74 | 0.21 | 0.004 |
| 0.557 | 0.5149 | 0.8493 | 0.9711 | ||
| Time × Site (region) | 4 | 16.62 | 17.89 | 9.06 | 6.92 |
| <0.0001 | <0.0001 | <0.0001 | 0.0002 | ||
| Residual | 544 |
| Source 1 | df | Proportion Fragmented Colonies | Proportion Recruits (Crusts) |
|---|---|---|---|
| Time | 1 | 10.5 | 0.64 |
| 0.0396 | 0.5049 | ||
| Region | 1 | 5.63 | 2.42 |
| 0.178 | 0.2303 | ||
| Site (region) | 4 | 3.09 | 0.72 |
| 0.0158 | 0.5698 | ||
| Time × Region | 1 | 1.73 | 0.05 |
| 0.287 | 0.8341 | ||
| Time × Site (region) | 4 | 2.76 | 2.29 |
| 0.0243 | 0.0604 | ||
| Residual | 544 |
| Source 1 | df | MS | Pseudo_F | P (Perm) | P (Perm) |
|---|---|---|---|---|---|
| Time | 1 | 8830.5 | 114.91 | <0.0001 | 9935 |
| Site | 9 | 6225.2 | 81.01 | <0.0001 | 9919 |
| Depth | 2 | 2960.2 | 38.52 | <0.0001 | 9955 |
| Time × Site | 9 | 741.84 | 9.65 | <0.0001 | 9891 |
| Time × Depth | 2 | 794.55 | 10.34 | <0.0001 | 9919 |
| Site × Depth | 18 | 299.50 | 3.90 | <0.0001 | 9850 |
| Time × Site × Depth | 18 | 133.11 | 1.73 | 0.0019 | 9844 |
| Residual | 300 | 76.849 |
| Source 1 | df | Acropora cervicornis | Orbicella annularis | Orbicella faveolata | Orbicella franksi | Dendrogyra cylindrus |
|---|---|---|---|---|---|---|
| Time | 1 | 142.84 <0.0001 | 1.39 0.2406 | 0.42 0.9072 | 0.88 0.3486 | 6.71 0.0063 |
Site Depth Time × Site Time × Depth Site × Depth Time × Site × Depth Residual | 9 2 9 2 18 18 300 | 77.83 <0.0001 14.69 <0.0001 12.67 <0.0001 8.43 0.0002 6.92 <0.0001 3.65 <0.0001 | 50.87 <0.0001 35.92 <0.0001 0.87 0.5522 0.16 0.8932 3.55 <0.0001 0.36 0.9967 | 31.82 <0.0001 30.68 <0.0001 0.55 0.8495 0.03 0.9908 2.83 <0.0001 0.28 0.9994 | 4.55 0.0002 13.90 <0.0001 0.84 0.5848 0.01 0.9952 0.78 0.7171 0.36 0.9930 | 1.10 0.3535 2.74 0.0651 0.66 0.7522 1.70 0.1852 0.93 0.5502 0.84 0.6616 |
| Source 1 | df | Macroalgae | Algal Turf | Ramicrusta textilis | Crustose Coralline Algae (Cca) | Cyanobacteria | Sand, Pavement, Rubble (SPR) | Sponge |
|---|---|---|---|---|---|---|---|---|
| Time | 1 | 1102.3 <0.0001 | 120.55 <0.0001 | 92.78 <0.0001 | 70.90 <0.0001 | 162.73 <0.0001 | 51.21 <0.0001 | 3.73 0.0512 |
Site Depth Time × Site Time × Depth Site × Depth Time × Site × Depth Residual | 9 2 9 2 18 18 300 | 45.75 <0.0001 396.33 <0.0001 25.50 <0.0001 57.27 <0.0001 9.97 <0.0001 3.75 <0.0001 | 52.71 <0.0001 111.14 <0.0001 9.53 <0.0001 1.83 0.1544 20.53 <0.0001 6.78 <0.0001 | 37.15 <0.0001 33.01 <0.0001 10.27 <0.0001 4.94 0.0014 15.20 <0.0001 4.15 <0.0001 | 9.55 <0.0001 109.90 <0.0001 4.24 <0.0001 2.74 0.0602 4.32 <0.0001 2.11 0.0026 | 5.90 <0.0001 64.85 <0.0001 4.22 0.0002 5.89 0.0020 4.40 <0.0001 1.65 0.0385 | 28.42 <0.0001 44.81 <0.0001 18.71 <0.0001 15.47 <0.0001 9.20 <0.0001 2.76 <0.0001 | 0.80 0.6119 11.52 <0.0001 0.62 0.7869 0.94 0.3966 0.82 0.6710 0.74 0.7677 |
| Source 1 | df | MS | Pseudo_F | P (Perm) | P (Perm) |
|---|---|---|---|---|---|
| Time | 1 | 21019 | 128.78 | <0.0001 | 9950 |
| Site | 9 | 5966.5 | 36.56 | <0.0001 | 9850 |
| Depth | 2 | 11826 | 72.46 | <0.0001 | 9938 |
| Time × Site | 9 | 1161.2 | 7.11 | <0.0001 | 9871 |
| Time × Depth | 2 | 1682.7 | 10.37 | <0.0001 | 9943 |
| Site × Depth | 18 | 1246.9 | 7.64 | <0.0001 | 9824 |
| Time × Site × Depth | 18 | 319.78 | 1.96 | <0.0001 | 9834 |
| Residual | 300 | 163.22 |
| Source 1 | df | Species Richness (S) | Diversity Index (H’c) | Evenness (J’c) |
|---|---|---|---|---|
| Time | 1 | 88.83 | 66.23 | 5.57 |
| <0.0001 | <0.0001 | 0.028 | ||
| Site | 9 | 51.39 | 42.41 | 11.86 |
| <0.0001 | <0.0001 | <0.0001 | ||
| Depth | 2 | 30.2 | 19.78 | 6.02 |
| <0.0001 | <0.0001 | 0.0058 | ||
| Time × Site | 9 | 3.3 | 4.01 | 7.91 |
| 0.0006 | <0.0001 | <0.0001 | ||
| Time × Depth | 2 | 5.86 | 11.28 | 4.53 |
| 0.0026 | <0.0001 | 0.0195 | ||
| Site × Depth | 18 | 4.58 | 4.58 | 5.34 |
| <0.0001 | <0.0001 | <0.0001 | ||
| Time × Site × Depth | 18 | 1.04 | 1.69 | 2.35 |
| 0.4107 | 0.0378 | 0.0055 | ||
| Residual | 300 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Delgado, E.A.; Alejandro-Camis, P.; Cabrera-Beauchamp, G.; Fonseca-Miranda, J.S.; Gómez-Andújar, N.X.; Gómez, P.; Guzmán-Rodríguez, R.; Olivo-Maldonado, I.; Suleimán-Ramos, S.E. Stronger Hurricanes and Climate Change in the Caribbean Sea: Threats to the Sustainability of Endangered Coral Species. Sustainability 2024, 16, 1506. https://doi.org/10.3390/su16041506
Hernández-Delgado EA, Alejandro-Camis P, Cabrera-Beauchamp G, Fonseca-Miranda JS, Gómez-Andújar NX, Gómez P, Guzmán-Rodríguez R, Olivo-Maldonado I, Suleimán-Ramos SE. Stronger Hurricanes and Climate Change in the Caribbean Sea: Threats to the Sustainability of Endangered Coral Species. Sustainability. 2024; 16(4):1506. https://doi.org/10.3390/su16041506
Chicago/Turabian StyleHernández-Delgado, Edwin A., Pedro Alejandro-Camis, Gerardo Cabrera-Beauchamp, Jaime S. Fonseca-Miranda, Nicolás X. Gómez-Andújar, Pedro Gómez, Roger Guzmán-Rodríguez, Iván Olivo-Maldonado, and Samuel E. Suleimán-Ramos. 2024. "Stronger Hurricanes and Climate Change in the Caribbean Sea: Threats to the Sustainability of Endangered Coral Species" Sustainability 16, no. 4: 1506. https://doi.org/10.3390/su16041506