
Field-Based Evaluation of Rice Genotypes for Enhanced Growth, Yield Attributes, Yield and Grain Yield Efficiency Index in Irrigated Lowlands of the Indo-Gangetic Plains

 , , ,
, , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Characteristics
2.2. Experimental Design and Treatments
2.3. Sampling and Measurements
2.4. Calculation of Related Indicators
2.5. Data Analysis
3. Results
3.1. Growth
3.1.1. Number of Tillers
3.1.2. Chlorophyll
3.1.3. Dry Matter Accumulation
3.1.4. Leaf Area Index
3.2. Yield-Attributing Characteristics
3.2.1. Number of Panicles
3.2.2. Filled Grains
3.2.3. 1000-Grain Weight
3.3. Grain Yield and Harvest Index
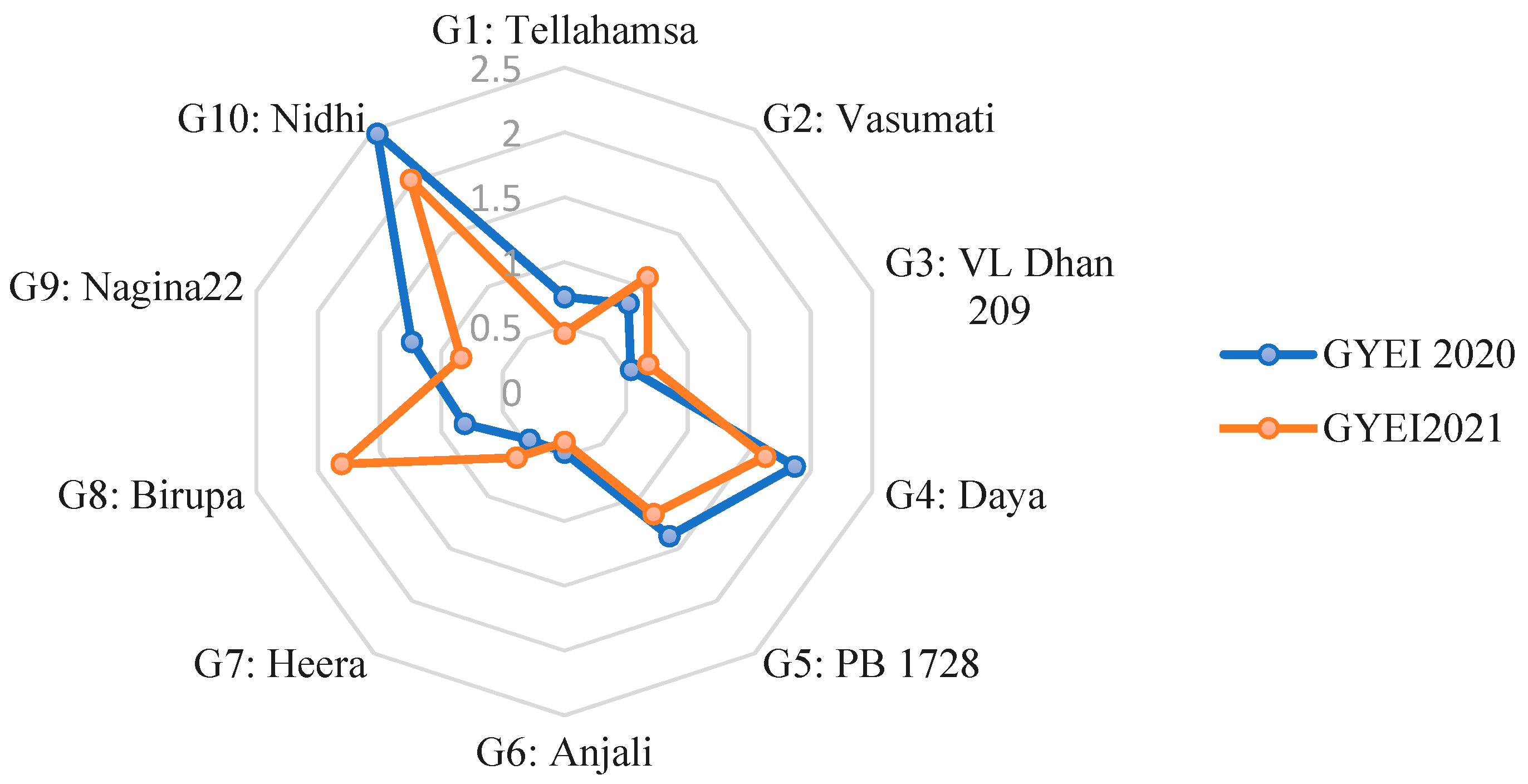
3.4. Grain Yield Efficiency Index
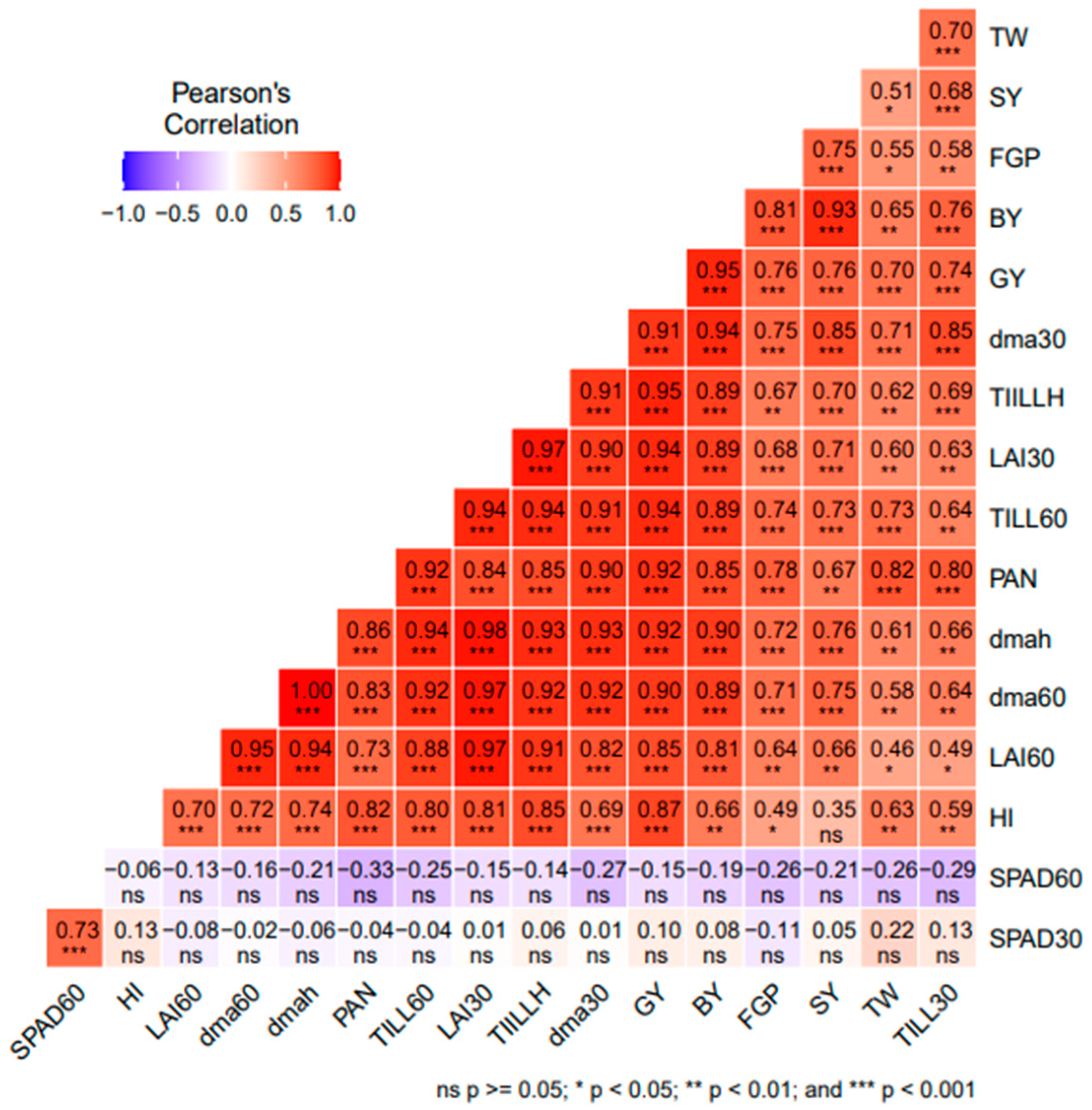
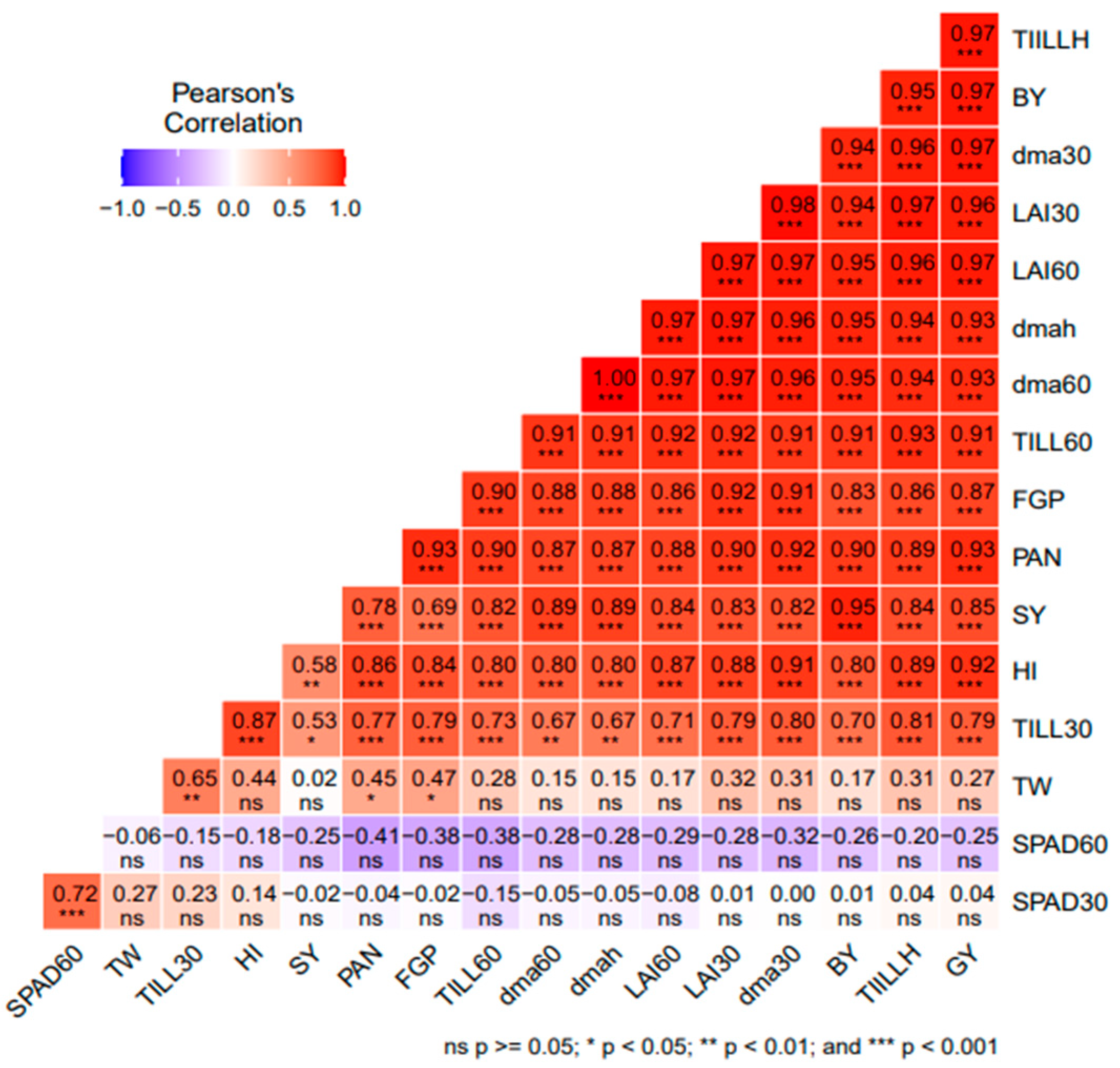
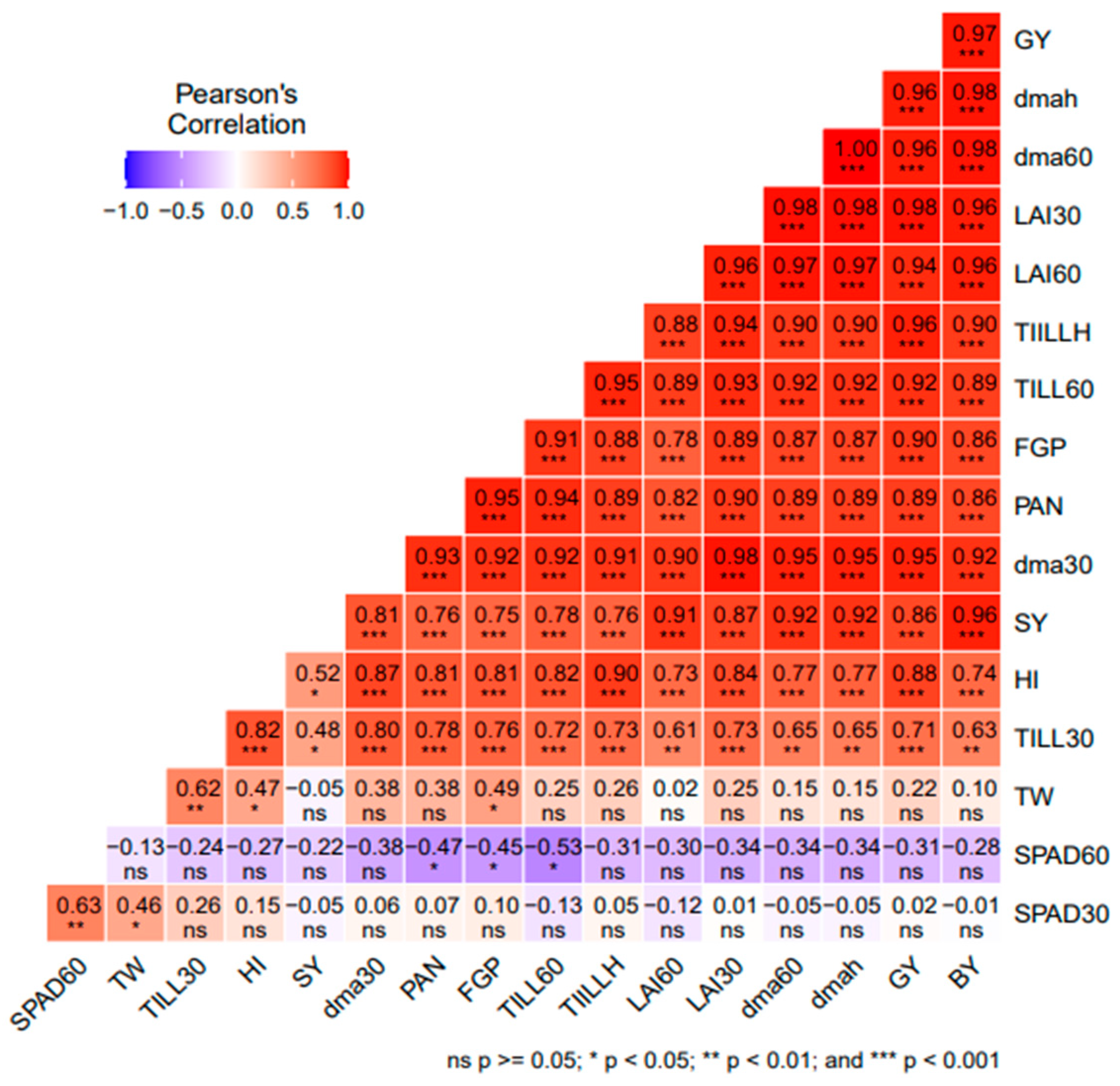
3.5. Correlation Analysis among Agro-Morphological Traits or Parameters of Rice
4. Discussion
4.1. Effect of Nitrogen
4.2. Rice Genotype
4.3. Interaction of Years, Nitrogen and Genotypes
4.4. Relationship between Agro-Morphological Traits or Parameters of Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Available online: https://www.faidelhi.org/statistics/statistical-database (accessed on 21 February 2022).
- Available online: https://www.rbi.org.in/scripts/AnnualPublications.aspx?head=Handbook%20of%20Statistics%20on%20Indian%20Economy (accessed on 21 February 2022).
- Food Grain Production Estimated to Reach New High of 150 Million Tonnes in 2021–2022: Centre. The Indian Express, 22 September 2021.
- Hakeem, K.R.; Chandna, R.; Altaf, A.; Muhammad, I. Physiological and molecular analysis of applied nitrogen in rice genotypes. Rice Sci. 2012, 19, 213–222. [Google Scholar] [CrossRef]
- Fu, Z.; Zhang, K.; Zhang, J.; Zhang, Y.; Cao, Q.; Tian, Y.; Zhu, Y.; Cao, W.; Liu, X. Optimizing nitrogen application and sowing date can improve environmental sustainability and economic benefit in wheat-rice rotation. Agric. Syst. 2023, 204, 103536. [Google Scholar] [CrossRef]
- Kumari, P.; Devi, L.L.; Kumar, A.; Pandey, A.; Sinha, S.K.; Singh, A.P. Differential response of rice genotypes to nitrogen availability is associated with the altered nitrogen metabolism and ionomic balance. Environ. Exp. Bot. 2022, 198, 104847. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Almaraz, M.; Aneja, V.; Austin, A.T.; Bai, E.; Cassman, K.G.; Compton, J.E.; Davidson, E.A.; Erisman, J.W.; Galloway, J.N.; et al. A World of Cobenefits: Solving the Global Nitrogen Challenge. Earth’s Future 2019, 7, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Chen, A.; Houlton, B.Z.; Zeng, Z.; Wei, S.; Zhao, C.; Lu, H.; Liao, Y.; Zheng, Z.; Luan, S.; et al. Spatial Variation of Reactive Nitrogen Emissions From China’s Croplands Codetermined by Regional Urbanization and Its Feedback to Global Climate Change. Geophys. Res. Lett. 2020, 47, e2019GL086551. [Google Scholar] [CrossRef]
- Cowan, N.; Bhatia, A.; Drewer, J.; Jain, N.; Singh, R.; Tomer, R.; Kumar, V.; Kumar, O.; Prasanna, R.; Ramakrishnan, B.; et al. Experimental comparison of continuous and intermittent flooding of rice in relation to methane, nitrous oxide and ammonia emissions and the implications for nitrogen use efficiency and yield. Agric. Ecosyst. Environ. 2021, 319, 107571. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, G.; Guo, S.; Li, X.; Yuan, J.; Hu, A. Improving Nitrogen Use Efficiency in Rice for Sustainable Agriculture: Strategies and Future Perspectives. Life 2022, 12, 1653. [Google Scholar] [CrossRef]
- Móring, A.; Hooda, S.; Raghuram, N.; Adhya, T.K.; Ahmad, A.; Bandyopadhyay, S.K.; Barsby, T.; Beig, G.; Bentley, A.R.; Bhatia, A.; et al. Nitrogen Challenges and Opportunities for Agricultural and Environmental Science in India. Front. Sustain. Food Syst. 2021, 5, 505347. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A Review of Methods to Improve Nitrogen Use Efficiency in Agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- de Vries, W. Impacts of nitrogen emissions on ecosystems and human health: A mini review. Curr. Opin. Environ. Sci. Health 2021, 21, 100249. [Google Scholar] [CrossRef]
- Rubaiyath, A.N.M.; Rahman, B.; Zhang, J. Trends in rice research: 2030 and beyond. Food Energy Secur. 2022, 12, e390. [Google Scholar] [CrossRef]
- Lianga, S.; Ren, G.; Liu, J.; Zhao, X.; Zhou, M.; McNeil, D.; Ye, G. Genotype-by-environment interaction is important for grain yield in irrigated lowland rice. Field Crops Res. 2015, 180, 90–99. [Google Scholar] [CrossRef]
- CGIAR-Consultative Group on International Agricultural Research; CGIAR Secretariat. CGIAR Annual Report; CGIAR: Washington, DC, USA, 1998. [Google Scholar]
- Panda, D.; Barik, J. Flooding Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Sci. 2021, 28, 43–57. [Google Scholar] [CrossRef]
- Singh, B.; Mishra, S.; Bisht, D.S.; Joshi, R. Growing Rice with Less Water: Improving Productivity by Decreasing Water Demand. In Rice Improvement; Ali, J., Wani, S.H., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Wang, W.; Huang, L.; Zhu, G.; Zhang, H.; Wang, Z.; Adnan, M.; Fahad, S. Screening of rice cultivars for nitrogen use efficiency and yield stability under varying nitrogen levels. J. Plant Growth Regul. 2022, 41, 1808–1819. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth–metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef]
- Fagaria, N.K. itrogen use efficiency in crop plants. In Nitrogen Management in Crop Production; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2014; pp. 297–314. [Google Scholar] [CrossRef]
- Pathak, H.; Bhatia, A.; Jain, N. Greenhouse Gas Emission from Indian Agriculture: Trends, Mitigation and Policy Needs; Indian Agricultural Research Institute: New Delhi, India, 2014; 39p. [Google Scholar]
- Eagle, A.J.; Bird, J.A.; Horwath, W.R.; Linquist, B.A.; Brouder, S.M.; Hill, J.E.; Kessel, C.V. Rice yield and nitrogen utilization efficiency under alternative straw management practices. Agron. J. 2000, 92, 1096–1103. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Voleti, S.R.; Subrahmanyam, D.; Surekha, K.; Rao Indian, P.R. Breeding rice for nitrogen use efficiency. Indian J. Genet. 2019, 79 (Suppl. 1), 208–215. [Google Scholar] [CrossRef]
- Singh, B. Agronomic Benefits of Neem Coated Urea—A Review; International Fertilizer Association Review Papers; International Fertilizer Association: Paris, France, 2016. [Google Scholar]
- Fageria, N.K.; Baligar, V.C. Methodology for evaluation of lowland rice genotypes for nitrogen use efficiency. J. Plant Nutr. 2003, 26, 1315–1333. [Google Scholar] [CrossRef]
- de Mendiburu, F. agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 3 February 2023).
- Hu, J.; Sun, S.Y.; Wang, X.L. Regulation of shoot branching by strigolactones and brassinosteroids: Conserved and specific functions of Arabidopsis BES1 and rice BZR1. Mol. Plant 2020, 13, 808–810. [Google Scholar] [CrossRef]
- Huang, M.; Chen, J.; Cao, F.; Jiang, L.; Zou, Y.; Deng, G. Improving physiological N-use efficiency by increasing harvest index in rice: A case in super-hybrid cultivar Guiliangyou 2. Arch. Agron. Soil Sci. 2016, 62, 725–743. [Google Scholar] [CrossRef]
- Xu, J.; Zha, M.; Li, Y.; Ding, Y.; Chen, L.; Ding, C.; Wang, S. The interaction between nitrogen availability and auxin, cytokinin, and strigolactone in the control of shoot branching in rice (Oryza sativa L.). Plant Cell Rep. 2015, 34, 1647–1662. [Google Scholar] [CrossRef] [PubMed]
- Zha, M.; Imran, M.; Wang, Y.; Xu, J.; Ding, Y.; Wang, S.H. Transcriptome analysis revealed the interaction among strigolactones, auxin, and cytokinin in controlling the shoot branching of rice. Plant Cell Rep. 2019, 38, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.; Feng, X.; Guangli, T.; Hu, X.; Shen, Q.; Guo, S. Optimizing nitrogen supply increases rice yield and nitrogen use efficiency by regulating yield formation factors. Field Crops Res. 2013, 150, 99–107. [Google Scholar] [CrossRef]
- Haque, M.A.; Haque, M.M. Growth, yield and nitrogen use efficiency of new rice variety under variable nitrogen rates. Am. J. Plant Sci. 2016, 7, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Shivay, Y.S.; Prasad, R.; Pal, M. Effect of nitrogen levels and coated urea on growth, yields and nitrogen use efficiency in aromatic rice. J. Plant Nutr. 2016, 39, 875–882. [Google Scholar] [CrossRef]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef]
- Zhou, W.; Lv, T.F.; Yang, Z.P.; Wang, T.; Fu, Y.; Chen, Y.; Ren, W.J. Morphophysiological mechanism of rice yield increase in response to optimized nitrogen management. Sci. Rep. 2017, 7, 17226. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Verma, A.; Ansari, M.W.; Shukla, A. Physiological response of rice (Oryza sativa L.) genotypes to elevated nitrogen applied under field conditions. Plant Signal. Behav. 2014, 9, e29015. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Biswas, D.K.; Xu, H.; Li, L.F.; Li, Y.H. Effects of nitrogen deficiency on photosynthetic traits of maize hybrids released in different years. Ann. Bot. 2005, 96, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Reddy, V. Nitrogen deficiency effects on plant growth, leaf photosynthesis, and hyperspectral reflectance properties of sorghum. Eur. J. Agron. 2005, 22, 391–403. [Google Scholar] [CrossRef]
- Sabri, R.S.; Rafii, M.Y.; Ismail, M.R.; Yusuff, O.; Chukwu, S.C.; Hasan, N.A. Assessment of agro-morphologic performance, genetic parameters and clustering pattern of newly developed blast resistant rice lines tested in four environments. Agronomy 2020, 10, 1098. [Google Scholar] [CrossRef]
- Ravikiran, K.T.; Krishnamurthy, S.L.; Warraich, A.S.; Sharma, P.C. Diversity and haplotypes of rice genotypes for seedling stage salinity tolerance analyzed through morpho-physiological and SSR markers. Field Crops Res. 2018, 220, 10–18. [Google Scholar] [CrossRef]
- Zhong, X.; Peng, S.; Sheehy, J.E.; Visperas, R.M.; Liu, H. Relationship between tillering and leaf area index: Quantifying critical leaf area index for tillering in rice. J. Agric. Sci. 2002, 138, 269–279. [Google Scholar] [CrossRef]
- Li, X.; Qian, Q.; Fu, Z.; Wang, Y.; Xiong, G.; Zeng, D.; Wang, X.; Liu, X.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef]
- Zou, J.; Chen, Z.; Zhang, S.; Zhang, W.; Jiang, G.; Zhao, X.; Zhaim, W.; Pan, X.; Zhu, L. Characterizations and fine mapping of a mutant gene for high tillering and dwarf in rice (Oryza sativa L.). Planta 2005, 222, 604–612. [Google Scholar] [CrossRef]
- Moldenhauer, K.A.K.; Gibbons, J.H. Rice morphology and development. In Rice: Origin, History, Technology, and Production; Smith, C.W., Dilday, R.H., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003; pp. 103–127. [Google Scholar]
- Yoshida, S.; Hayakawa, Y. Effects of mineral nutrition on tillering of rice. Soil Sci. Plant Nutr. 1970, 16, 186–191. [Google Scholar] [CrossRef]
- Ying, J.F.; Peng, S.B.; He, Q.R.; Yang, H.; Yang, C.; Visperas, R.M. Comparison of high yield rice in tropical and subtropical environments. I. Determinants of grain and dry matter yields. Field Crops Res. 1998, 57, 71–84. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.; Kong, X.S.; Hou, D.P.; Gu, J.F.; Liu, L.J.; Wang, Z.Q.; Yang, J.C. Progressive integrative crop managements increase grain yield, nitrogen use efficiency and irrigation water productivity in rice. Field Crops Res. 2018, 215, 1–11. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, T.T.; Liu, L.J.; Wang, Z.Q.; Yang, J.C.; Zhang, J.H. Performance in grain yield and physiological traits of rice in the Yangtze River basin of China during the last 60 yr. J. Integr. Agric. 2013, 12, 57–66. [Google Scholar] [CrossRef]
- Yoshida, S. Fundamentals of Rice Crop Science; International Rice Research Institute (IRRI): Los Baños, Philippines, 1981; p. 269. [Google Scholar]
- Counce, P.A.; Wells, B.R. Rice plant population density effect on early-season nitrogen requirement. J. Prod. Agric. 1990, 3, 390–393. [Google Scholar] [CrossRef]
- Jiang, L.G.; Cao, W.X.; Gan, X.Q.; Xu, J.Y.; Dong, D.F.; Chen, N.P.; Lu, F.Y.; Qin, H.D. Nitrogen uptake and utilization under different nitrogen management and influence on grain yield and quality in rice. Agri. Sci. China 2004, 37, 490–496. [Google Scholar]
- Fageria, N.K.; Santos, A.B.D.; Coelho, A.M. Growth, yield and yield components of lowland rice as Influenced by ammonium sulfate and urea fertilization. J. Plant Nutr. 2011, 34, 371–386. [Google Scholar] [CrossRef]
- Bocianowski, J.; Tratwal, A.; Nowosad, K. Genotype by environment interaction for main winter triticale varieties characteristics at two levels of technology using additive main effects and multiplicative interaction model. Euphytica 2021, 217, 26. [Google Scholar] [CrossRef]
- Chairi, F.; Aparicio, N.; Serret, M.D.; Araus, J.L. Breeding effects on the genotype× environment interaction for yield of durum wheat grown after the Green Revolution: The case of Spain. Crop J. 2020, 8, 623–634. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al Mamun, M. AMMI and GGE biplot analysis for yield performance and stability assessment of selected Bambara groundnut (Vigna subterranea L. Verdc.) genotypes under the multi-environmental trials (METs). Sci. Rep. 2021, 11, 22791. [Google Scholar] [CrossRef]
- Ferreira Coelho, I.; Peixoto, M.A.; Marcal, T.d.S.; Bernardeli, A.; Silva Alves, R.; de Lima, R.O.; Reis, E.F.d.; Bhering, L.L. Accounting for spatial trends in multi-environment diallel analysis in maize breeding. PLoS ONE 2021, 16, e0258473. [Google Scholar] [CrossRef]
- Li, C.-x.; Yun, S.; Zhang, L.-l. Effects of long-term organic fertilization on soil microbiologic characteristics, yield and sustainable production of winter wheat. J. Integr. Agric. 2018, 17, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, M.; Alharbi, K.; El-Denary, M.E.; Abd El-Megeed, T.; Naeem, E.-S.; Monir, S.; Al-Shaye, N.A.; Ammar, M.H.; Attia, K.; Dora, S.A.; et al. Detection of Superior Rice Genotypes and Yield Stability under Different Nitrogen Levels Using AMMI Model and Stability Statistics. Plants 2022, 11, 2775. [Google Scholar] [CrossRef]
- Huang, M.; Shan, S.; Cao, J.; Fang, S.; Tian, A.; Liu, Y.; Zou, Y. Primary-tiller panicle number is critical to achieving high grain yields in machine-transplanted hybrid rice. Sci. Rep. 2020, 10, 2811. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Sun, F.; Yuan, S.; Peng, S.; Wang, F. Different mechanisms underlying the yield advantage of ordinary hybrid and super hybrid rice over inbred rice under low and moderate N input conditions. Field Crops Res. 2018, 216, 150–157. [Google Scholar] [CrossRef]
- Wu, L.; Yuan, S.; Huang, L.; Sun, F.; Zhu, G.; Li, G.; Fahad, S.; Peng, S.; Wang, F. Physiological mechanisms underlying the high-grain yield and high-nitrogen use efficiency of elite rice cultivars under a low rate of nitrogen application in China. Trends Plant Sci. 2016, 7, 1024. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, J. Crop management techniques to enhance harvest index in rice. J. Exp. Bot. 2010, 61, 3177–3189. [Google Scholar] [CrossRef] [Green Version]
- Walkley, A.J.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chronic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Subbiah, B.V.; Asija, G.L. A rapid procedure for the estimation of available nitrogen in soils. Curr. Sci. 1956, 25, 259–260. [Google Scholar]
- Olsen, R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; Circular 939; United States Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Hanway, J.J.; Heidel, H. Soil Analysis Methods as Used in Iowa State College Soil Testing Laboratory; Bulletin 57; Iowa State College of Agriculture: Ames, IA, USA, 1952; 131p. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Crop Stages for N Fertilization | Amount of N Fertilizer Applied through Neem-Oil-Coated Urea (46.6% N) | SSP * (16% P2O5) | MOP ** (60% K2O) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Basal | Tillering | Panicle Initiation | Basal (Kg ha−1) | Tillering (Kg ha−1) | Panicle Initiation (Kg ha−1) | Total N (Kg ha−1) | Basal (Kg P2O5 ha−1) | Basal (Kg K2O ha−1) | |
| N0 | - | - | - | - | - | - | - | 60 | 40 |
| N60 | 10 days after transplanting (50% of N60) | 25% of N60 | 25% of N60 | 30 | 15 | 15 | 60 | 60 | 40 |
| N120 | 10 days after transplanting (half of N120) | 25% of N120 | 25% of N120 | 60 | 30 | 30 | 120 | 60 | 40 |
| Nitrogen/Genotype | SPAD Values at 30 DAT | Tillers m−2 at 30 DAT | ||
|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | |
| N0 | 25.3 b | 24.4 b | 89.0 c | 88.0 c |
| N60 | 31.3 a | 30.4 a | 112 b | 111 b |
| N120 | 33.5 a | 32.5 a | 129 a | 128 a |
| ‘Tella Hamsa’ | 31.1 bc | 30.2 bc | 103 ef | 102 ef |
| ‘Vasumati’ | 33.2 ab | 32.2 ab | 109 cde | 111 bcd |
| ‘VL Dhan 209’ | 27.6 de | 26.7 de | 107 de | 107 de |
| ‘Daya’ | 34.9 a | 34.0 a | 118 ab | 116 abc |
| ‘PB 1728’ | 33.3 ab | 32.5 ab | 116 abc | 113 bcd |
| ‘Anjali’ | 29.2 cd | 28.1 cd | 99 f | 99 f |
| ‘Heera’ | 31.1 bc | 30.3 bc | 102 ef | 103 e |
| ‘Birupa’ | 23.8 f | 22.6 f | 111 bcd | 118 ab |
| Nagina22 | 30.9 bc | 30.1 bc | 113 bcd | 109 cde |
| ‘Nidhi’ | 25.4 ef | 24.1 ef | 122 a | 122 a |
| N | 11.8 | 11.79 | 2.25 | 2.38 |
| V | 7.0 | 6.97 | 3.05 | 2.87 |
| N × V | ns | ns | ns | ns |
| Nitrogen × Genotype | ‘Tella Hamsa’ | ‘Vasumati’ | ‘VL Dhan 209’ | ‘Daya’ | ‘PB 1728’ | ‘Anjali’ | ‘Heera’ | ‘Birupa’ | ‘Nagina 22’ | ‘Nidhi’ | Mean | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SPAD value (60 DAT) | ||||||||||||
| 2020 | N0 | 38.7 mn | 42.3 ij | 33.6 pq | 40.9 kl | 43.9 efgh | 38.0 n | 42.6 hij | 24.6 s | 37.5 n | 34.8 op | 37.7 c |
| N60 | 44.9 de | 42.9 ghi | 38.8 mn | 46.5 ab | 45.1 cde | 41.4 jk | 44.0 efg | 30.3 r | 39.9 lm | 36.0 o | 41.0 b | |
| N120 | 46.3 bc | 47.1 ab | 46.1 bcd | 47.8 a | 44.9 de | 43.5 fghi | 44.7 ef | 32.7 q | 46.3 bc | 38.8 mn | 43.8 a | |
| Mean | 43.3 d | 44.1 bc | 39.5 f | 45.1 a | 44.6 ab | 40.9 e | 43.8 cd | 29.2 h | 41.2 e | 36.5 g | ||
| * N × G = 1.32/* G × N = 1.30 | ||||||||||||
| 2021 | N0 | 38.3 p | 41.4 l | 32.6 u | 40.0 n | 43.0 i | 37.0 q | 41.6 kl | 23.7 x | 36.5 r | 33.6 t | 36.8 c |
| N60 | 43.7 h | 41.8 jp | 38.4 p | 45.6 cd | 44.1 g | 40.0 m | 43.2 i | 29.4 w | 39.0 o | 35.0 s | 40.1 b | |
| N120 | 45.3 d | 46.1 b | 44.8 e | 46.7 a | 44.5 ef | 42.0 j | 44.1 fg | 31.1 v | 45.9 bc | 37.3 q | 42.8 a | |
| Mean | 42.4 d | 43.1 c | 38.6 g | 44.1 a | 43.9 b | 40.0 f | 43.0 c | 28.1 i | 40.4 e | 35.3 h | ||
| * N × V = 0.38/* V × N = 0.37 | ||||||||||||
| Tillers (60 DAT) | ||||||||||||
| 2020 | N0 | 292 op | 307 no | 280 pq | 395 k | 399 jk | 239 r | 268 q | 311 n | 368 m | 413 j | 327 c |
| N60 | 365 m | 385 kl | 378 lm | 538 e | 504 f | 297 nop | 361 m | 390 kl | 501 f | 564 d | 428 b | |
| N120 | 454 h | 474 g | 451 h | 620 b | 590 c | 375 lm | 433 i | 464 gh | 583 c | 647 a | 509 a | |
| Mean | 370 f | 388 e | 369 f | 517 b | 498 c | 304 h | 354 g | 388 e | 484 d | 541 a | ||
| * N × G = 16.94/* G × N = 16.21 | ||||||||||||
| 2021 | N0 | 268 t | 310 q | 280. st | 399 jk | 368 nop | 238 u | 292 rs | 393 kl | 305 qr | 411 j | 326 c |
| N60 | 361 p | 389 klm | 378 lmno | 505 f | 501 f | 295 qrs | 365 op | 536 e | 383 klmn | 562 d | 428 b | |
| N120 | 432 i | 464 gh | 450 h | 589 c | 582 c | 375 mnop | 453 h | 620 b | 473 g | 647 a | 508 a | |
| Mean | 354 g | 387 e | 369 f | 4976 c | 484 d | 303 h | 370 f | 516 b | 387 e | 540 a | ||
| Tillers at harvest | ||||||||||||
| 2020 | N0 | 269 pqr | 302 nop | 277 opq | 392 fghi | 375 ghij | 238 r | 251 qr | 320 lmn | 379 ghij | 409 fg | 321 c |
| N60 | 361 hijk | 375 ghij | 346 jkl | 509 cd | 489 de | 303 nop | 340 klm | 396 fgh | 461 e | 530 c | 411 b | |
| N120 | 408 fg | 391 fghi | 359 ijk | 595 ab | 569 b | 309 mno | 381 ghij | 416 f | 492 de | 621 a | 454 a | |
| Mean | 346 fg | 356 f | 328 gh | 499 b | 478 c | 284 i | 324 h | 377 e | 444 d | 520 a | ||
| * N × G = 34.9/* G × N = 34.5 | ||||||||||||
| 2021 | N0 | 250 qr | 319 lmn | 276 opq | 374 ghij | 378 ghij | 238 r | 268 pqr | 391 fghi | 301 nop | 408 fg | 320 c |
| N60 | 338 klm | 395 fgh | 346 jkl | 488 de | 460 e | 303 nop | 360 hijk | 508 cd | 374 ghij | 529 c | 410 b | |
| N120 | 380 ghij | 415 f | 358 ijk | 568 b | 492 de | 308 mno | 407 fg | 593 ab | 391 fghi | 619 a | 453 a | |
| Mean | 323 h | 376 e | 327 gh | 477 c | 443 d | 283 i | 345 fg | 497 b | 355 f | 519 a | ||
| * N × G = 34.6/* G × N = 34.2 | ||||||||||||
| Nitrogen × Genotype | ‘Tella Hamsa’ | ‘Vasumati’ | ‘VL Dhan 209’ | ‘Daya’ | ‘PB 1728’ | ‘Anjali’ | ‘Heera’ | ‘Birupa’ | ‘Nagina 22’ | ‘Nidhi’ | Mean | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAT | N0 | 88 no | 104 m | 103 m | 117 ijk | 115 jkl | 84 o | 92 n | 114 kl | 104 m | 119 ijk | 104 c |
| (2020) | N60 | 123 hi | 131 fg | 121 hij | 152 d | 160 c | 118 ijk | 108 lm | 134 ed | 149 d | 166 bc | 136 b |
| N120 | 132 fg | 140 e | 136 ef | 164 c | 172 ab | 126 gh | 121 hij | 149 d | 160 c | 178 a | 148 a | |
| Mean | 114 h | 125 f | 120 g | 145 c | 149 b | 109 i | 107 i | 132 e | 137 d | 154 a | ||
| * N × G = 7.1/* G × N = 7.8 | ||||||||||||
| 30 DAT | N0 | 84 n | 106 lm | 102 m | 116 ij | 113 jkl | 91 n | 87 n | 118 ij | 100 m | 113 jk | 103 c |
| (2021) | N60 | 117 ij | 148 d | 121 hi | 151 d | 133 ef | 107 klm | 122 hi | 165 bc | 130 fg | 159 c | 135 b |
| N120 | 125 gh | 159 c | 135 ef | 163 bc | 149 d | 120 hij | 131 fg | 177 a | 140 e | 170 ab | 147 a | |
| Mean | 109 g | 138 c | 120 e | 144 b | 131 d | 106 g | 114 f | 153 a | 123 e | 147 b | ||
| * N × G = 7.3/* G × N = 7.7 | ||||||||||||
| 60 DAT | N0 | 311 l | 456 hij | 331 l | 578 e | 579 e | 245 m | 239 m | 489 ghi | 447 ijk | 588 e | 426 c |
| (2020) | N60 | 473 ghi | 561 ef | 498 gh | 763 bc | 773 ab | 399 k | 396 k | 673 d | 689 d | 783 ab | 601 b |
| N120 | 491 ghi | 582 e | 523 fg | 789 ab | 800 ab | 414 jk | 408 jk | 697 d | 716 cd | 820 a | 624 a | |
| Mean | 425 d | 533 c | 451 d | 710 a | 717 a | 352 e | 348 e | 620 b | 617 b | 731 a | ||
| * N × G = ns/* G × N = ns | ||||||||||||
| 60 DAT | N0 | 245 v | 514 m | 330 t | 577 k | 488 o | 238 v | 310 u | 587 j | 387 s | 577 k | 425 c |
| (2021) | N60 | 397 r | 688 h | 498 n | 763 e | 672 i | 395 rs | 472 p | 782 c | 560 l | 772 d | 600 b |
| N120 | 413 q | 715 f | 522 m | 788 c | 697 g | 407 q | 491 no | 819 a | 582 jk | 799 b | 623 a | |
| Mean | 352 i | 639 d | 450 g | 709 c | 619 e | 347 i | 424 h | 730 a | 510 f | 716 b | ||
| * N × G = 8.5/* G × N = 8.2 | ||||||||||||
| Harvest | N0 | 523 l | 692 i | 552 kl | 862 f | 860 f | 444 m | 439 m | 788 gh | 699 i | 923 e | 678 c |
| (2020) | N60 | 709 i | 841 fg | 748 hi | 1145 b | 1160 b | 597 jk | 594 jk | 1009 d | 1033 cd | 1175 ab | 901 b |
| N120 | 737 hi | 873 ef | 784 gh | 1183 ab | 1100 ab | 620 j | 611 jk | 1046 cd | 1073 c | 1230 a | 936 a | |
| Mean | 656 f | 802 d | 695 e | 1063 b | 1073 b | 554 g | 548 g | 948 c | 935 c | 1109 a | ||
| * N × G = 60.1/* G × N = 60.4 | ||||||||||||
| Harvest | N0 | 444 v | 769 o | 551 t | 862 kl | 787 n | 438 v | 522 u | 922 j | 620 r | 858 l | 677 c |
| (2021) | N60 | 596 s | 1032 h | 747 p | 1144 e | 1008 i | 593 s | 708 q | 1174 c | 840 m | 1159 d | 900 b |
| N120 | 619 r | 1072 f | 783 n | 1182 c | 1045 g | 610 r | 736 p | 1228 a | 873 k | 1198 b | 935 a | |
| Mean | 553 i | 958 d | 694 g | 1062 c | 947 e | 547 i | 655 h | 1108 a | 778 f | 1072 b | ||
| * N × G = 12.1/* G × N = 24.5 | ||||||||||||
| Nitrogen × Genotype | ‘Tella Hamsa’ | ‘Vasumati’ | ‘VL Dhan 209’ | ‘Daya’ | ‘PB 1728’ | ‘Anjali’ | ‘Heera’ | ‘Birupa’ | ‘Nagina 22’ | ‘Nidhi’ | Mean | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LAI (30) | ||||||||||||
| 2020 | N0 | 0.44 r | 0.51 q | 0.32 s | 0.78 lmn | 0.72 no | 0.16 t | 0.21 t | 0.62 p | 0.67 op | 0.81 lm | 0.52 c |
| N60 | 1.12 j | 1.21 i | 1.00 k | 1.66 de | 1.50 f | 0.77 mn | 0.84 l | 1.36 h | 1.49 fg | 1.69 d | 1.26 b | |
| N120 | 1.54 f | 1.62 e | 1.42 g | 2.06 a | 1.91 b | 1.18 ij | 1.25 i | 1.76 c | 1.89 b | 2.11 a | 1.67 a | |
| Mean | 1.03 e | 1.11 d | 0.91 f | 1.5 a | 1.37 b | 0.7 h | 0.77 g | 1.25 c | 1.35 b | 1.54 a | ||
| * N × G = 0.07/* V × N = 0.11 | ||||||||||||
| 2021 | N0 | 0.15 r | 0.60 o | 0.31q | 0.66 no | 0.70 mn | 0.19 r | 0.42 p | 0.77 lm | 0.49 p | 0.80 l | 0.51 c |
| N60 | 0.76 lm | 1.34 h | 0.99 k | 1.47 fg | 1.48 f | 0.83 l | 1.11 j | 1.64 de | 1.20 i | 1.68 cd | 1.25 b | |
| N120 | 1.17 ij | 1.75 c | 1.40 gh | 1.87 b | 1.89 b | 1.24 i | 1.52 f | 2.05 a | 1.61 e | 2.09 a | 1.66 a | |
| Mean | 0.69 h | 1.23 c | 0.9 f | 1.33 b | 1.36 b | 0.75 g | 1.01 e | 1.49 a | 1.1 d | 1.52 a | ||
| * N × G = 0.07/* G × N = 0.12 | ||||||||||||
| LAI (60) | ||||||||||||
| 2020 | N0 | 3.07 r | 3.27 q | 2.71 s | 3.66 n | 3.57 no | 2.06 u | 2.42 t | 3.37 pq | 3.46 op | 3.86 m | 3.15 b |
| N60 | 4.66 j | 4.96 i | 4.36 k | 5.97 c | 5.67 e | 4.07 l | 4.17 l | 5.17 h | 5.47f | 6.20 a | 5.08 a | |
| N120 | 5.02 i | 5.32 g | 4.98 i | 6.11 b | 5.91 cd | 4.13 l | 4.58 j | 5.41 fg | 5.83 d | 6.36 a | 5.35 a | |
| Mean | 4.25 g | 4.52 f | 4.02 h | 5.25 b | 5.05 c | 3.42 j | 3.72 i | 4.65 e | 4.92 d | 5.47 a | ||
| * N × G = 0.13/* G × N = 0.34 | ||||||||||||
| 2021 | N0 | 2.05 u | 3.35 pq | 2.70 s | 3.45 op | 3.55 no | 2.40 t | 3.05 r | 3.65 n | 3.25 q | 3.85 m | 3.13 b |
| N60 | 4.05 l | 5.15 h | 4.35 k | 5.45 f | 5.65 e | 4.15 l | 4.65 j | 5.95 c | 4.95 i | 6.20 a | 5.07 a | |
| N120 | 4.10 l | 5.40 fg | 4.95 i | 5.80 d | 5.90 cd | 4.55 j | 5.00 i | 6.10 b | 5.30 g | 6.35 a | 5.33 a | |
| Mean | 3.4 j | 4.63 e | 4 h | 4.9 d | 5.03 c | 3.7 i | 4.23 g | 5.23 b | 4.5 f | 5.47 a | ||
| * N × G = 0.14/* G × N = 0.35 | ||||||||||||
| Panicles m−2 | ||||||||||||
| 2020 | N0 | 213 p | 293 n | 246 o | 363 fghi | 297 n | 228 op | 236 op | 366 fghi | 249 o | 385 ef | 288 c |
| N60 | 302 mn | 379 efgh | 334 jkl | 428 d | 384 efg | 315 lmn | 317 lmn | 436 d | 341 ijkl | 463.96 c | 370 b | |
| N120 | 326 klm | 395 e | 352 hijk | 478 bc | 491 ab | 335 jkl | 331 jkl | 492 ab | 357 ghij | 511 a | 407 a | |
| Mean | 281 f | 356 d | 311 e | 423 b | 391 c | 293 f | 294 f | 431 b | 316 e | 453 a | ||
| * N × G = 26.9/* G × N = 30.8 | ||||||||||||
| 2021 | N0 | 213 o | 294 m | 246 n | 363 fgh | 297 m | 228 no | 236 no | 366 fgh | 249 n | 385 ef | 287 c |
| N60 | 302 lm | 379 efg | 334 ijk | 428 d | 384 efg | 315 klm | 317 klm | 436 d | 341 hijk | 464 c | 370 b | |
| N120 | 325 jkl | 394 e | 351 hij | 477 bc | 491 ab | 335 ijk | 330 jk | 491 ab | 357 ghi | 510 a | 406 a | |
| Mean | 280 f | 356 d | 310 e | 423 b | 390 c | 292 f | 294 f | 431 b | 315 e | 453 a | ||
| * N × G = 26.9/* G × N = 30.8 | ||||||||||||
| Treatment | Filled Grains Panicle−1 | 1000-Grain Weight (g) | ||
|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | |
| N0 | 92 c | 91 c | 20.5 b | 19.5 b |
| N60 | 119 b | 118 b | 26.4 ab | 25.4 ab |
| N120 | 128 a | 127 a | 29.6 a | 28.6 a |
| ‘Tella Hamsa’ | 96 d | 95 d | 24.0 c | 23.1 cd |
| ‘Vasumati’ | 115 bc | 114 bc | 25.5 abc | 24.5 abcd |
| ‘VL Dhan209’ | 102 cd | 102 cd | 24.9 bc | 24.1 abcd |
| ‘Daya’ | 129 a | 128 a | 27.3 ab | 26.3 ab |
| ‘PB 1728’ | 121 ab | 120 ab | 24.6 bc | 23.9 abcd |
| ‘Anjali’ | 98 d | 97 d | 23.6 c | 22.5 d |
| ‘Heera’ | 101 d | 100 d | 25.5 abc | 24.7 abcd |
| ‘Birupa’ | 131 a | 130 a | 27.9 a | 26.8 a |
| ‘Nagina 22’ | 105 cd | 104 cd | 24.4 bc | 23.6 bcd |
| ‘Nidhi’ | 133 a | 132 a | 27.0 ab | 25.8 abc |
| Interaction | ns | ns | ns | ns |
| Treatment | Grain Yield of Rice (t ha−1) |
|---|---|
| Year | |
| 2020 | 3.86 a |
| 2021 | 3.77 a |
| Nitrogen | |
| N0 | 2.82 c |
| N60 | 4.15 b |
| N120 | 4.48 a |
| Genotype | |
| ‘Tella Hamsa’ | 2.85 e |
| ‘Vasumati’ | 3.86 d |
| ‘VL Dhan209’ | 2.94 e |
| ‘Daya’ | 5.06 b |
| ‘PB 1728’ | 4.40 c |
| ‘Anjali’ | 2.47 f |
| ‘Heera’ | 2.84 e |
| ‘Birupa’ | 4.40 c |
| ‘Nagina 22’ | 3.81 d |
| ‘Nidhi’ | 5.54 a |
| Year/Genotype | 2020 | 2021 | ||||||
|---|---|---|---|---|---|---|---|---|
| N0 | N60 | N120 | Mean | N0 | N60 | N120 | Mean | |
| ‘Tella Hamsa’ | 2.80 kl | 3.60 gh | 3.80 fg | 3.40 f | 1.93 o | 2.81 kl | 2.19 mno | 2.31 f |
| ‘Vasumati’ | 2.60 lm | 3.90 fg | 4.05 f | 3.52 ef | 3.10 jk | 4.35 f | 5.15 e | 4.20 c |
| ‘VL Dhan209’ | 2.40 mn | 3.20 ij | 3.15 ijk | 2.92 g | 2.15 no | 3.45 ghij | 3.28 hij | 2.96 e |
| ‘Daya’ | 3.60 gh | 5.30 c | 6.60 b | 5.17 b | 3.45 ghij | 5.33 de | 6.06 abc | 4.95 b |
| ‘PB 1728’ | 3.40 hi | 4.90 d | 5.30 c | 4.53 c | 3.14i jk | 4.53 f | 5.13 e | 4.27 c |
| ‘Anjali’ | 1.95 o | 2.90 jkl | 3.05 ijk | 2.63 f | 2.01 no | 2.62 lm | 2.28 mno | 2.30 h |
| ‘Heera’ | 2.20 no | 2.90 jkl | 3.00 jk | 2.70 gh | 2.05 no | 3.32 hij | 3.56 ghi | 2.98 e |
| ‘Birupa’ | 3.10 ijk | 3.75 fgh | 4.05 f | 3.63 e | 3.38 ghij | 5.62 cd | 6.48 a | 5.16 ab |
| ‘Nagina 22’ | 3.15 ijk | 4.45 e | 5.20 cd | 4.27 d | 2.42 lmn | 3.82 g | 3.79 g | 3.34 d |
| ‘Nidhi’ | 3.80 fg | 6.25 b | 7.35 a | 5.80 a | 3.68 gh | 5.91 bc | 6.24 ab | 5.28 a |
| Mean | 2.90 c | 4.12 b | 4.56 a | 2.73 c | 4.18 b | 4.41 a | ||
| * N:G:Y = 0.41/** N:G = 0.29/*** Y:G = 0.24/**** Y:N = 0.20 | ||||||||
| Nitrogen × Genotype | ‘Tella Hamsa’ | ‘Vasumati’ | ‘VL Dhan 209’ | ‘Daya’ | ‘PB 1728’ | ‘Anjali’ | ‘Heera’ | ‘Birupa’ | ‘Nagina 22’ | ‘Nidhi’ | Mean | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harvest index (%) | ||||||||||||
| 2020 | N0 | 35.5 jkl | 36.5 ijk | 28.5 p | 40.5 ef | 36.0 ijk | 30.5 op | 32.5 mno | 35.5 jkl | 39.0 fgh | 44.0 bcd | 35.8 c |
| N60 | 36.5 ijk | 37.5 hij | 30.5 op | 43.0 cd | 37.5 hij | 31.5 no | 32.5 mno | 36.5 ijk | 40.0 efg | 45.5 ab | 37.1 b | |
| N120 | 36.5i jk | 34.5 klm | 32.5 mno | 45.0 v | 38.0 ghi | 33.0 mn | 33.5 lmn | 37.5 hij | 42.0 de | 47.5 a | 38.0 a | |
| Mean | Mean | 36.2 d | 36.2 d | 30.5 f | 42.8 b | 37.2 d | 31.7 ef | 32.8 e | 36.5 d | 40.3 c | 45.7 a | |
| * N × G = 2.08/* G × N = 2.08 | ||||||||||||
| 2021 | N0 | 37.3 ij | 39.7 efghij | 37.0 j | 41.2 cdefghi | 40.1 efghij | 37.6 hij | 37.9 hij | 42.0 bcdefg | 37.9 hij | 43.2 abcde | 39.4 c |
| N60 | 38.1 ghij | 40.3 defghij | 38.9 fghij | 42.6 bcdef | 41.4 bcdefgh | 38.4 ghij | 39.1 fghij | 43.2 abcde | 39.2 fghij | 45.3 ab | 40.7 b | |
| N120 | 39.1 fghij | 40.9 cdefghij | 40.8 cdefghij | 44.2 abcd | 42.7 bcdef | 39.2 fghij | 40.3 defghij | 44.4 abc | 40.5 cdefghij | 46.8 a | 41.9 a | |
| Mean | 38.2 d | 40.0 cd | 38.9 d | 42.7 b | 41.4 bc | 38.4 d | 39.1 d | 43.2 ab | 39.2 cd | 45.1 a | ||
| * N × G = ns/* G × N = ns | ||||||||||||
| Genotype | 2020 | 2021 |
|---|---|---|
| ‘Tella Hamsa’ | 0.73 | 0.45 |
| ‘Vasumati’ | 0.84 | 1.09 |
| ‘VL Dhan 209’ | 0.54 | 0.68 |
| ‘Daya’ | 1.87 | 1.63 |
| ‘PB 1728’ | 1.38 | 1.17 |
| ‘Anjali’ | 0.47 | 0.39 |
| ‘Heera’ | 0.46 | 0.63 |
| ‘Birupa’ | 0.81 | 1.81 |
| ‘Nagina 22’ | 1.24 | 0.84 |
| ‘Nidhi’ | 2.46 | 2.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawdiya, S.; Kumar, D.; Shivay, Y.S.; Bhatia, A.; Mehrotra, S.; Chandra, M.S.; Kumawat, A.; Kumar, R.; Price, A.H.; Raghuram, N.; et al. Field-Based Evaluation of Rice Genotypes for Enhanced Growth, Yield Attributes, Yield and Grain Yield Efficiency Index in Irrigated Lowlands of the Indo-Gangetic Plains. Sustainability 2023, 15, 8793. https://doi.org/10.3390/su15118793
Gawdiya S, Kumar D, Shivay YS, Bhatia A, Mehrotra S, Chandra MS, Kumawat A, Kumar R, Price AH, Raghuram N, et al. Field-Based Evaluation of Rice Genotypes for Enhanced Growth, Yield Attributes, Yield and Grain Yield Efficiency Index in Irrigated Lowlands of the Indo-Gangetic Plains. Sustainability. 2023; 15(11):8793. https://doi.org/10.3390/su15118793
Chicago/Turabian StyleGawdiya, Sandeep, Dinesh Kumar, Yashbir S. Shivay, Arti Bhatia, Shweta Mehrotra, Mandapelli Sharath Chandra, Anita Kumawat, Rajesh Kumar, Adam H. Price, Nandula Raghuram, and et al. 2023. "Field-Based Evaluation of Rice Genotypes for Enhanced Growth, Yield Attributes, Yield and Grain Yield Efficiency Index in Irrigated Lowlands of the Indo-Gangetic Plains" Sustainability 15, no. 11: 8793. https://doi.org/10.3390/su15118793