
Quantifying the Unvoiced Carbon Pools of the Nilgiri Hill Region in the Western Ghats Global Biodiversity Hotspot—First Report
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
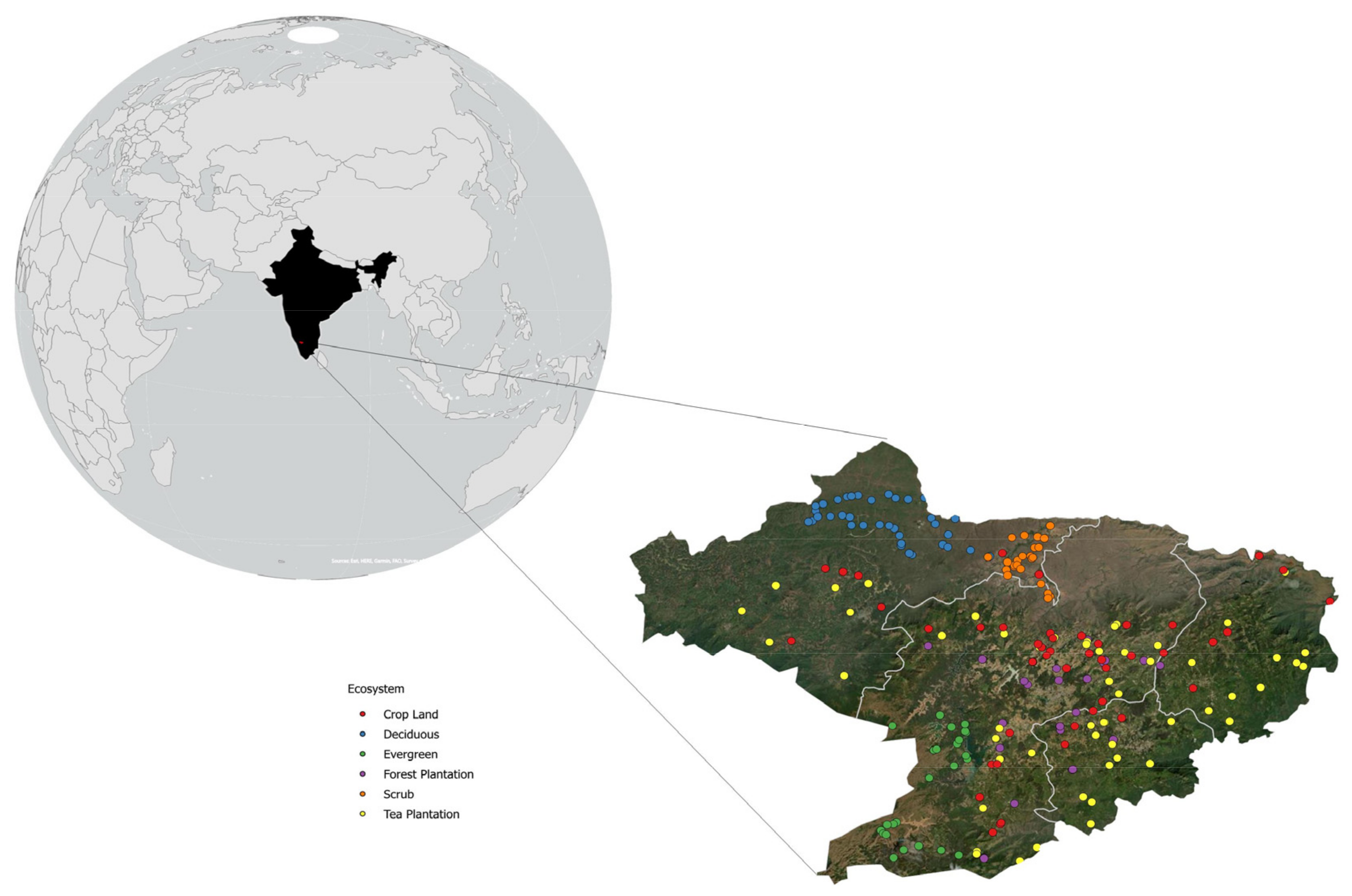
2.2. Study Area
2.3. Geomorphology and Soils
2.4. Soil Sampling and Analysis
2.5. Water-Soluble Organic Carbon (WSOC)
2.6. Water-Soluble Carbohydrates (WSCs)
2.7. Dehydrogenase
2.8. Microbial Biomass Carbon (MBC)
2.9. Microbial Biomass Nitrogen (MBN)
2.10. Aggregate-Size Organic Carbon (ASOC)
2.11. Land-Degradation Index (LDI)
2.12. Statistical Analyses
3. Results
3.1. Total Organic Carbon (TOC) (%) Distribution among Different Ecosystems in NHR
3.2. WSOC (mg kg−1) Distribution among Different Ecosystems in NHR
3.3. Hot Water-Soluble Carbohydrate (WSC) (mg kg−1) Distribution among Different Ecosystems in NHR
3.4. Dehydrogenase (µg TPF g−1 day−1) Distribution among Different Ecosystems in NHR
3.5. Microbial Biomass Carbon (MBC) (mg kg−1) Distribution among Different Ecosystems in NHR
3.6. MBN (mg kg−1) Distribution among Different Ecosystems in NHR
3.7. Aggregate-Size Organic Carbon (ASOC) (g kg−1) Distribution among Different Ecosystems in NHR
3.7.1. ASOC (2 mm) (g kg−1) Distribution among Different Ecosystems in NHR
3.7.2. ASOC (0.25 mm) (g kg−1) Distribution among Different Ecosystems in NHR
3.7.3. ASOC (0.053 mm) (g kg−1) Distribution among Different Ecosystems in NHR
3.7.4. ASOC (<0.053 mm) (g kg−1) Distribution among Different Ecosystems in NHR
3.8. Relationship between TOC and Carbon Pools
3.9. Land-Degradation Indices (LDIs) of Different Land Uses in NHR
4. Discussion
4.1. Effect of Land-Use Change on Water-Soluble Carbon (WSOC)
4.2. Effect of Land-Use Change on Water-Soluble Carbohydrates (WSC)
4.3. Effect of Land-Use Change on Microbial Biomass Carbon (MBC)
4.4. Effect of Land Use on Microbial Biomass Nitrogen (MBN)
4.5. Effect of Land-Use Change on Dehydrogenase
4.6. Effect of Land-Use Change on Aggregate-Size Organic Carbon (ASOC)
4.7. Land-Degradation Index (LDI) in NHR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scharlemann, J.P.W.; Tanner, E.V.J.; Hiederer, R.; Kapos, V. Global Soil Carbon: Understanding and Managing the Largest Terrestrial Carbon Pool. Carbon Manag. 2014, 5, 81–91. [Google Scholar] [CrossRef]
- Lal, R. Sequestration of Atmospheric CO2 in Global Carbon Pools. Energy Environ. Sci. 2008, 1, 86–100. [Google Scholar] [CrossRef]
- Lal, R. Soil Health and Carbon Management. Food Energy Secur. 2016, 5, 212–222. [Google Scholar] [CrossRef]
- Villarino, S.H.; Studdert, G.A.; Laterra, P. How Does Soil Organic Carbon Mediate Trade-Offs between Ecosystem Services and Agricultural Production? Ecol. Indic. 2019, 103, 280–288. [Google Scholar] [CrossRef]
- Laik, R.; Kumara, B.H.; Pramanick, B.; Singh, S.K.; Nidhi; Alhomrani, M.; Gaber, A.; Hossain, A. Labile soil organic matter pools are influenced by 45 years of applied farmyard manure and mineral nitrogen in the wheat—Pearl millet cropping system in the sub-tropical condition. Agronomy 2021, 11, 2190. [Google Scholar] [CrossRef]
- Pramanick, B.; Kumar, M.; Naik, B.M.; Kumar, M.; Singh, S.K.; Maitra, S.; Naik, B.S.S.S.; Rajput, V.D.; Minkina, T. Long-term conservation tillage and precision nutrient management in maize–wheat cropping system: Effect on soil properties, crop production, and economics. Agronomy 2022, 12, 2766. [Google Scholar] [CrossRef]
- Victoria, R.; Banwart, S.; Black, H.; Ingram, J.; Joosten, H.; Milne, E.; Noellemeyer, E.; Baskin, Y. The Benefits of Soil Carbon. Foresight Chapter UNEP Yearb. 2012, 2012, 19–33. [Google Scholar]
- Paustian, K.; Collins, H.P.; Paul, E.A. Management Controls on Soil Carbon. In Soil Organic Matter in Temperate Agroecosystems; CRC Press: Boca Raton, FL, USA, 2019; pp. 15–49. [Google Scholar] [CrossRef]
- Hati, K.M.; Biswas, A.K.; Somasundaram, J.; Mohanty, M.; Singh, R.K.; Sinha, N.K.; Chaudhary, R.S. Soil Organic Carbon Dynamics and Carbon Sequestration under Conservation Tillage in Tropical Vertisols. In Carbon Management in Tropical and Sub-Tropical Terrestrial Systems; Springer: Singapore City, Singapore, 2019; pp. 201–212. [Google Scholar] [CrossRef]
- Zhang, L.; Zhuang, Q.; He, Y.; Liu, Y.; Yu, D.; Zhao, Q.; Shi, X.; Xing, S.; Wang, G. Toward Optimal Soil Organic Carbon Sequestration with Effects of Agricultural Management Practices and Climate Change in Tai-Lake Paddy Soils of China. Geoderma 2016, 275, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Jagadesh, M.; Selvi, D.; Thiyageshwari, S.; Srinivasarao, C.; Kalaiselvi, T.; Lourdusamy, K.; Kumaraperumal, R.; Allan, V. Soil Carbon Dynamics Under Different Ecosystems of Ooty Region in the Western Ghats Biodiversity Hotspot of India. J. Soil Sci. Plant Nutr. 2023, 23, 1374–1385. [Google Scholar] [CrossRef]
- Wolters, V. Invertebrate Control of Soil Organic Matter Stability. Biol. Fertil. Soils 2000, 31, 1–19. [Google Scholar] [CrossRef]
- Six, J.; Callewaert, P.; Lenders, S.; de Gryze, S.; Morris, S.J.; Gregorich, E.G.; Paul, E.A.; Paustian, K. Measuring and Understanding Carbon Storage in Afforested Soils by Physical Fractionation. Soil Sci. Soc. Am. J. 2002, 66, 1981–1987. [Google Scholar] [CrossRef] [Green Version]
- Blair, G.J.; Lefroy, R.D.B.; Lisle, L. Soil Carbon Fractions Based on Their Degree of Oxidation, and the Development of a Carbon Management Index for Agricultural Systems. Aust. J. Agric. Res. 1995, 46, 1459–1466. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of Microbial Biomass Carbon and Nitrogen in Soil. Adv. Nitrogen Cycl. 1988, 16, 368–386. [Google Scholar]
- Cambardella, C.A.; Elliott, E.T. Particulate Soil Organic-matter Changes across a Grassland Cultivation Sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Trumbore, S.E. Potential Responses of Soil Organic Carbon to Global Environmental Change. Proc. Natl. Acad. Sci. USA 1997, 94, 8284–8291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, B.; Majumuder, B.; Adhya, T.K.; Bandyopadhyay, P.K.; Gangopadhyay, A.; Sarkar, D.; Kundu, M.C.; Choudhury, S.G.; Hazra, G.C.; Kundu, S.; et al. Potential of Double-Cropped Rice Ecology to Conserve Organic Carbon under Subtropical Climate. Glob. Change Biol. 2008, 14, 2139–2151. [Google Scholar] [CrossRef]
- Chan, K.Y.; Bowman, A.; Oates, A. Oxidizible organic carbon fractions and soil quality changes in an oxic paleustalf under different pasture leys. Soil Sci. 2001, 166, 61–67. [Google Scholar] [CrossRef]
- Nath, A.J.; Brahma, B.; Sileshi, G.W.; Das, A.K. Impact of Land Use Changes on the Storage of Soil Organic Carbon in Active and Recalcitrant Pools in a Humid Tropical Region of India. Sci. Total Environ. 2018, 624, 908–917. [Google Scholar] [CrossRef]
- Luo, Y.; White, L.W.; Canadell, J.G.; DeLucia, E.H.; Ellsworth, D.S.; Finzi, A.; Lichter, J.; Schlesinger, W.H. Sustainability of Terrestrial Carbon Sequestration: A Case Study in Duke Forest with Inversion Approach. Glob. Biogeochem. Cycles 2003, 17, 1021. [Google Scholar] [CrossRef] [Green Version]
- Paul, E.A.; Collins, H.P.; Leavitt, S.W. Dynamics of Resistant Soil Carbon of Midwestern Agricultural Soils Measured by Naturally Occurring 14C Abundance. Geoderma 2001, 104, 239–256. [Google Scholar] [CrossRef]
- Maskell, L.C.; Crowe, A.; Dunbar, M.J.; Emmett, B.; Henrys, P.; Keith, A.M.; Norton, L.R.; Scholefield, P.; Clark, D.B.; Simpson, I.C. Exploring the Ecological Constraints to Multiple Ecosystem Service Delivery and Biodiversity. J. Appl. Ecol. 2013, 50, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil Carbon Debt of 12,000 Years of Human Land Use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldewijk, K.K.; Beusen, A.; Van Drecht, G.; De Vos, M. The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Glob. Ecol. Biogeogr. 2011, 20, 73–86. [Google Scholar] [CrossRef]
- Kooch, Y.; Tavakoli, M.; Akbarinia, M. Tree Species Could Have Substantial Consequences on Topsoil Fauna: A Feedback of Land Degradation/Restoration. Eur. J. For. Res. 2018, 137, 793–805. [Google Scholar] [CrossRef]
- Olsson, L.; Barbosa, H.; Bhadwal, S.; Cowie, A.; Delusca, K.; Flores-Renteria, D.; Hermans, K.; Jobbagy, E.; Kurz, W.; Li, D. Land Degradation: IPCC Special Report on Climate Change, Desertification, Land 5 Degradation, Sustainable Land Management, Food Security, and 6 Greenhouse Gas Fluxes in Terrestrial Ecosystems. In IPCC Special Report on Climate Change, Desertification, Land 5 Degradation, Sustainable Land Management, Food Security, and 6 Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC), UNEP: Nairobi, Kenya, 2019; p. 1. [Google Scholar]
- FAO. Recarbonization of Global Soils: A Dynamic Response to Offset Global Emissions; FAO: Rome, Italy, 2019. [Google Scholar]
- Wei, X.; Shao, M.; Gale, W.; Li, L. Global Pattern of Soil Carbon Losses Due to the Conversion of Forests to Agricultural Land. Sci. Rep. 2015, 4, 4062. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Sharma, L.K.; Sharma, K.C. Assessment of Land Use Change and Its Effect on Soil Carbon Stock Using Multitemporal Satellite Data in Semiarid Region of Rajasthan, India. Ecol. Process. 2019, 8, 1–17. [Google Scholar] [CrossRef]
- Vasantha, K.S.; Bhagavanulu, D.V.S. Effect of Deforestation on Landslides in Nilgiris District—A Case Study. J. Indian Soc. Remote Sens. 2008, 36, 105–108. [Google Scholar] [CrossRef]
- Sasmitha, R.; Iqshanullah, A.M.; Arunachalam, R. Ecosystem Changes in Shola Forest-Grassland Mosaic of the Nilgiri Biosphere Reserve (NBR). Environ. Issues Sustain. Dev. IntechOpen 2021. [Google Scholar] [CrossRef]
- Sasmitha, R.; Arunachalam, R.; Iqshanullah, M. Documentation of Soil Related Environmental Issues and It’s Contributing Factors: A Study among the Hilly Tribes of the Nilgiri District. Madras Agric. J. 2019, 106, 1. [Google Scholar] [CrossRef]
- Krishnan, S. Landscape, Labor, and Label: The Second World War, Pastoralist Amelioration, and Pastoral Conservation in the Nilgiris, South India (1929–1945). Int. Labor Work. Cl. Hist. 2015, 87, 92–110. [Google Scholar] [CrossRef]
- Baldi, G.; Paruelo, J.M. Land-Use and Land Cover Dynamics in South American Temperate Grasslands. Ecol. Soc. 2008, 13, 6. [Google Scholar] [CrossRef]
- Thirumalai, P.; Anand, P.H.; Murugesan, J. Changing Land Use Pattern in Nilgiris Hill Environment Using Geospatial Technology. Int. J. Recent Sci. Res. 2015, 6, 3679–3683. [Google Scholar]
- Vega, E.; Baldi, G.; Jobbágy, E.G.; Paruelo, J. Land Use Change Patterns in the Río de La Plata Grasslands: The Influence of Phytogeographic and Political Boundaries. Agric. Ecosyst. Environ. 2009, 134, 287–292. [Google Scholar] [CrossRef]
- Nalina, P.; Meenambal, T.; Sridhar, R.S. Land use land cover dynamics of nilgiris district, india inferred from satellite imageries. Am. J. Appl. Sci. 2014, 11, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Gunnell, Y.; Louchet, A. The Influence of Rock Hardness and Divergent Weatherin g on the Interpretation of Apatite Fission-Track Denudation Rates. Evid. Charnockites South India Sri Lanka. Z. Für Geomorphol. 2000, 44, 33–57. [Google Scholar] [CrossRef]
- Caner, L.; Bourgeon, G.; Bourgeon Andisols, G.; Bourgeon, G.; Gunnell, Y.; Radhakrishna, B. Andisols of the Nilgiri Highlands: New Insight into Their Classification, Age and Genesis; Geological Society of India: Bangalore, India, 2001. [Google Scholar]
- McGill, W.B.; Cannon, K.R.; Robertson, J.A.; Cook, F.D. Dynamics of soil microbial biomass and water-soluble organic c in breton l after 50 years of cropping to two rotations. Can. J. Soil Sci. 1986, 66, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Brink, R.H.; Dubach, P.; Lynch, D.L. Measurement of carbohydrates in soil hydrolyzates with anthrone. Soil Sci. 1960, 89, 157–166. [Google Scholar] [CrossRef]
- Thalmann, A. Zur Methodik Der Bestimmung Der DehydrogenaseaktivitAt Im Boden Mittels Triphenytetrazoliumchlorid (TTC). Landwirtsch 1968, 21, 249–258. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Barrow, C.J. Land Degradation: Development and Breakdown of Terrestrial Environments; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Tollefson, M. Introduction: Plot (), Qplot (), and Ggplot (), Plus Some. In Visualizing Data in R 4; Springer: Berkeley, CA, USA, 2021; pp. 3–7. [Google Scholar]
- Scaglia, B.; Adani, F. Biodegradability of Soil Water Soluble Organic Carbon Extracted from Seven Different Soils. J. Environ. Sci. 2009, 21, 641–646. [Google Scholar] [CrossRef]
- Jha, P.; Garg, N.; Lakaria, B.L.; Biswas, A.K.; Rao, A.S. Soil and Residue Carbon Mineralization as Affected by Soil Aggregate Size. Soil Tillage Res. 2012, 121, 57–62. [Google Scholar] [CrossRef]
- Ma, X.Z.; Chen, L.J.; Chen, Z.H.; Wu, Z.J.; Zhang, L.L.; Zhang, Y.L. Soil Glycosidase Activities and Water Soluble Organic Carbon under Different Land Use Types. Rev. De La Cienc. Del Suelo Y Nutr. Veg. 2010, 10, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Moharana, P.C.; Naitam, R.K.; Verma, T.P.; Meena, R.L.; Kumar, S.; Tailor, B.L.; Singh, R.S.; Singh, S.K.; Samal, S.K. Effect of Long-Term Cropping Systems on Soil Organic Carbon Pools and Soil Quality in Western Plain of Hot Arid India. Arch. Agron. Soil Sci. 2017, 63, 1661–1675. [Google Scholar] [CrossRef]
- Chantigny, M.H. Dissolved and Water-Extractable Organic Matter in Soils: A Review on the Influence of Land Use and Management Practices. Geoderma 2003, 113, 357–380. [Google Scholar] [CrossRef]
- Christ, M.J.; David, M.B. Dynamics of Extractable Organic Carbon in Spodosol Forest Floors. Soil Biol. Biochem. 1996, 28, 1171–1179. [Google Scholar] [CrossRef]
- Sheng, H.; Zhou, P.; Zhang, Y.; Kuzyakov, Y.; Zhou, Q.; Ge, T.; Wang, C. Loss of Labile Organic Carbon from Subsoil Due to Land-Use Changes Insubtropical China. Soil Biol. Biochem. 2015, 88, 148–157. [Google Scholar] [CrossRef]
- Deng, L.; Shangguan, Z. Afforestation Drives Soil Carbon and Nitrogen Changes in China. Land Degrad. Dev. 2017, 28, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Zsolnay, A.; Görlitz, H. Water Extractable Organic Matter in Arable Soils: Effects of Drought and Long-Term Fertilization. Soil Biol. Biochem. 1994, 26, 1257–1261. [Google Scholar] [CrossRef]
- Jensen, L.S.; Mueller, T.; Magid, J.; Nielsen, N.E. Temporal Variation of C and N Mineralization, Microbial Biomass and Extractable Organic Pools in Soil after Oilseed Rape Straw Incorporation in the Field. Soil Biol. Biochem. 1997, 29, 1043–1055. [Google Scholar] [CrossRef]
- Martin-Olmedo, P.; Rees, R.M. Short-Term N Availability in Response to Dissolved-Organic-Carbon from Poultry Manure, Alone or in Combination with Cellulose. Biol Fertil Soils 1999, 29, 386–393. [Google Scholar] [CrossRef]
- Jha, P.; De, A.; Lakaria, B.L.; Biswas, A.K.; Singh, M.; Reddy, K.S.; Rao, A.S. Soil Carbon Pools, Mineralization and Fluxes Associated with Land Use Change in Vertisols of Central India. Natl. Acad. Sci. Lett. 2012, 35, 475–483. [Google Scholar] [CrossRef]
- Ghani, A.; Dexter, M.; Sarathchandra, U.; Perrott, K.W.; Singleton, P. Assessment of Extractable Hot-Water Carbon as an Indicator of Soil Quality on Soils under Long-Term Pastoral, Cropping, Market Gardening and Native Vegetation. In Proceedings of the Australian and New Zealand Second Joint Soils Conference, Lincoln, New Zealand, 14–21 August 2000; pp. 119–120. [Google Scholar]
- Haynes, R.J.; Swift, R.S. Stability of Soil Aggregates in Relation to Organic Constituents and Soil Water Content. J. Soil Sci. 1990, 41, 73–83. [Google Scholar] [CrossRef]
- Puget, P.; Angers, D.A.; Chenu, C. Nature of Carbohydrates Associated with Water-Stable Aggregates of Two Cultivated Soils. Soil Biol. Biochem. 1998, 31, 55–63. [Google Scholar] [CrossRef]
- Takele, L.; Chimdi, A.; Abebaw, A. Dynamics of Soil Fertility as Influenced by Different Land Use Systems and Soil Depth in West Showa Zone, Gindeberet District, Ethiopia. Agric. For. Fish. 2014, 3, 489–494. [Google Scholar]
- Six, J.; Paustian, K.; Elliott, E.T.; Combrink, C. Soil Structure and Organic Matter, I. Distribution of Aggregate-size Classes and Aggregate-associated Carbon. Soil Sci. Soc. Am. J. 2000, 64, 681–689. [Google Scholar] [CrossRef]
- Haynes, R.J.; Francis, G.S. Changes in Microbial Biomass C, Soil Carbohydrate Composition and Aggregate Stability Induced by Growth of Selected Crop and Forage Species under Field Conditions. J. Soil Sci. 1993, 44, 665–675. [Google Scholar] [CrossRef]
- Hernández, T.; Garcia, C.; Reinhardt, I. Short-Term Effect of Wildfire on the Chemical, Biochemical and Microbiological Properties of Mediterranean Pine Forest Soils. Biol. Fertil. Soils 1997, 25, 109–116. [Google Scholar] [CrossRef]
- Arunachalam, K.; Arunachalam, A.; Melkania, N.P. Influence of Soil Properties on Microbial Populations, Activity and Biomass in Humid Subtropical Mountainous Ecosystems of India. Biol. Fertil. Soils 1999, 30, 217–223. [Google Scholar] [CrossRef]
- Dos Santos, U.J.; de Medeiros, E.V.; Duda, G.P.; Marques, M.C.; de Souza, E.S.; Brossard, M.; Hammecker, C. Land Use Changes the Soil Carbon Stocks, Microbial Biomass and Fatty Acid Methyl Ester (FAME) in Brazilian Semiarid Area. Arch. Agron. Soil Sci. 2019, 65, 755–769. [Google Scholar] [CrossRef]
- Mganga, K.Z.; Kuzyakov, Y. Glucose Decomposition and Its Incorporation into Soil Microbial Biomass Depending on Land Use in Mt. Kilimanjaro Ecosystems. Eur. J. Soil Biol. 2014, 62, 74–82. [Google Scholar] [CrossRef]
- Van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L.; de Ruiter, P.C.; Lair, G.J. Effects of Land Use on Soil Microbial Biomass, Activity and Community Structure at Different Soil Depths in the Danube Floodplain. Eur. J. Soil Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Soleimani, A.; Hosseini, S.M.; Bavani, A.R.M.; Jafari, M.; Francaviglia, R. Influence of Land Use and Land Cover Change on Soil Organic Carbon and Microbial Activity in the Forests of Northern Iran. Catena 2019, 177, 227–237. [Google Scholar] [CrossRef]
- Feng, W.; Zou, X.; Schaefer, D. Above-and Belowground Carbon Inputs Affect Seasonal Variations of Soil Microbial Biomass in a Subtropical Monsoon Forest of Southwest China. Soil Biol. Biochem. 2009, 41, 978–983. [Google Scholar] [CrossRef]
- Jin, H.; Sun, O.J.; Liu, J. Changes in Soil Microbial Biomass and Community Structure with Addition of Contrasting Types of Plant Litter in a Semiarid Grassland Ecosystem. J. Plant Ecol. 2010, 3, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-H.; Chiu, C.-Y.; Tian, G. Seasonal Dynamics of Soil Microbial Biomass in Coastal Sand Dune Forest. Pedobiologia 2005, 49, 645–653. [Google Scholar] [CrossRef]
- Chen, C.; Liu, W.; Jiang, X.; Wu, J. Effects of Rubber-Based Agroforestry Systems on Soil Aggregation and Associated Soil Organic Carbon: Implications for Land Use. Geoderma 2017, 299, 13–24. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S. Response of Labile Soil Organic Matter to Changes in Forest Vegetation in Subtropical Regions. Appl. Soil Ecol. 2011, 47, 210–216. [Google Scholar] [CrossRef]
- Padalia, K.; Bargali, S.S.; Bargali, K.; Khulbe, K. Microbial Biomass Carbon and Nitrogen in Relation to Cropping Systems in Central Himalaya, India. Curr. Sci. 2018, 115, 1741–1750. [Google Scholar] [CrossRef]
- Lepcha, N.T.; Devi, N.B. Effect of Land Use, Season, and Soil Depth on Soil Microbial Biomass Carbon of Eastern Himalayas. Ecol. Process. 2020, 9, 1–14. [Google Scholar] [CrossRef]
- Fall, D.; Diouf, D.; Zoubeirou, A.M.; Bakhoum, N.; Faye, A.; Sall, S.N. Effect of Distance and Depth on Microbial Biomass and Mineral Nitrogen Content under Acacia Senegal (L.) Willd. Trees. J. Environ. Manag. 2012, 95, S260–S264. [Google Scholar] [CrossRef]
- Anderson, T.-H.; Domsch, K.H. Ratios of Microbial Biomass Carbon to Total Organic Carbon in Arable Soils. Soil Biol. Biochem. 1989, 21, 471–479. [Google Scholar] [CrossRef]
- Priha, O. Microbial Activities in Soils Under Scots Pine, Norway Spruce and Silver Birch; Hakapaino Oy: Helsinki, Finland, 1999; ISBN 9514016785. [Google Scholar]
- Vásquez-Murrieta, M.S.; Govaerts, B.; Dendooven, L. Microbial Biomass C Measurements in Soil of the Central Highlands of Mexico. Appl. Soil Ecol. 2007, 35, 432–440. [Google Scholar] [CrossRef]
- Diaz-Ravina, M.; Carballas, T.; Acea, M.J. Microbial Biomass and Metabolic Activity in Four Acid Soils. Soil Biol. Biochem. 1988, 20, 817–823. [Google Scholar] [CrossRef]
- McCulley, R.L.; Burke, I.C. Microbial Community Composition across the Great Plains: Landscape versus Regional Variability. Soil Sci. Soc. Am. J. 2004, 68, 106–115. [Google Scholar] [CrossRef]
- Schimel, D.S.; Braswell, B.H.; Holland, E.A.; McKeown, R.; Ojima, D.S.; Painter, T.H.; Parton, W.J.; Townsend, A.R. Climatic, Edaphic, and Biotic Controls over Storage and Turnover of Carbon in Soils. Glob. Biogeochem. Cycles 1994, 8, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, P.; Kong, C.H. Urease, Invertase, Dehydrogenase and Polyphenoloxidase Activities in Paddy Soil Influenced by Allelopathic Rice Variety. Eur. J. Soil Biol. 2009, 45, 436–441. [Google Scholar] [CrossRef]
- Nannipieri, P.; Greco, S.; Ceccanti, B. Ecological Significance of the Biological Activity in Soil. In Soil Biochemistry, 1st ed.; Routledge: Oxfordshire, UK, 1990. [Google Scholar]
- Zhang, Z.; Liew, C.W.; Handy, D.E.; Zhang, Y.; Leopold, J.A.; Hu, J.; Guo, L.; Kulkarni, R.N.; Loscalzo, J.; Stanton, R.C. High Glucose Inhibits Glucose-6-phosphate Dehydrogenase, Leading to Increased Oxidative Stress and Β-cell Apoptosis. FASEB J. 2010, 24, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Salazar, S.; Sánchez, L.E.; Alvarez, J.; Valverde, A.; Galindo, P.; Igual, J.M.; Peix, A.; Santa-Regina, I. Correlation among Soil Enzyme Activities under Different Forest System Management Practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- Bing-Cheng, Y.; Dong-Xia, Y.U.E. Soil Microbial and Enzymatic Activities across a Chronosequence of Chinese Pine Plantation Development on the Loess Plateau of China. Pedosphere 2012, 22, 1–12. [Google Scholar]
- Chodak, M.; Niklińska, M. Effect of Texture and Tree Species on Microbial Properties of Mine Soils. Appl. Soil Ecol. 2010, 46, 268–275. [Google Scholar] [CrossRef]
- Moeskops, B.; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; de Neve, S. Soil Microbial Communities and Activities under Intensive Organic and Conventional Vegetable Farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Romero, E.; Fernández-Bayo, J.; Díaz, J.M.C.; Nogales, R. Enzyme Activities and Diuron Persistence in Soil Amended with Vermicompost Derived from Spent Grape Marc and Treated with Urea. Appl. Soil Ecol. 2010, 44, 198–204. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, J.; Zhang, J.; Qin, S. Soil Microbial Biomass and Activity Response to Repeated Drying–Rewetting Cycles along a Soil Fertility Gradient Modified by Long-Term Fertilization Management Practices. Geoderma 2010, 160, 218–224. [Google Scholar] [CrossRef]
- Zhang, N.; Xing-Dong, H.E.; Yu-Bao, G.A.O.; Yong-Hong, L.I.; Hai-Tao, W.; Di, M.A.; Zhang, R.; Yang, S. Pedogenic Carbonate and Soil Dehydrogenase Activity in Response to Soil Organic Matter in Artemisia Ordosica Community. Pedosphere 2010, 20, 229–235. [Google Scholar] [CrossRef]
- Chu, F.; Lidstrom, M.E. XoxF Acts as the Predominant Methanol Dehydrogenase in the Type I Methanotroph Methylomicrobium Buryatense. J. Bacteriol. 2016, 198, 1317–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macci, C.; Doni, S.; Peruzzi, E.; Masciandaro, G.; Mennone, C.; Ceccanti, B. Almond Tree and Organic Fertilization for Soil Quality Improvement in Southern Italy. J. Environ. Manag. 2012, 95, S215–S222. [Google Scholar] [CrossRef]
- Xie, W.; Zhou, J.; Wang, H.; Chen, X.; Lu, Z.; Yu, J.; Chen, X. Short-Term Effects of Copper, Cadmium and Cypermethrin on Dehydrogenase Activity and Microbial Functional Diversity in Soils after Long-Term Mineral or Organic Fertilization. Agric Ecosyst. Environ. 2009, 129, 450–456. [Google Scholar] [CrossRef]
- Tejada, M.; García-Martínez, A.M.; Gómez, I.; Parrado, J. Application of MCPA Herbicide on Soils Amended with Biostimulants: Short-Time Effects on Soil Biological Properties. Chemosphere 2010, 80, 1088–1094. [Google Scholar] [CrossRef]
- Ghorbani, M.; Amirahmadi, E.; Konvalina, P.; Moudrý, J.; Kopecký, M.; Hoang, T.N. Carbon Pool Dynamic and Soil Microbial Respiration Affected by Land Use Alteration: A Case Study in Humid Subtropical Area. Land 2023, 12, 459. [Google Scholar] [CrossRef]
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and Rewetting Effects on C and N Mineralization and Microbial Activity in Surface and Subsurface California Grassland Soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Gajda, A.M. Effect of Different Tillage Systems on Some Microbiological Properties of Soils under Winter Wheat. Int. Agrophys. 2008, 22, 201–208. [Google Scholar]
- Benbi, D.K.; Boparai, A.K.; Brar, K. Decomposition of Particulate Organic Matter Is More Sensitive to Temperature than the Mineral Associated Organic Matter. Soil Biol. Biochem. 2014, 70, 183–192. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Gajda, A.M.; Doran, J.W.; Wienhold, B.J.; Kettler, T.A.; Lal, R. Estimation of Particulate and Total Organic Matter by Weight Loss-on-Ignition. In Assessment Methods for Soil Carbon; CRC Press: Boca Raton, FL, USA, 2001; pp. 349–359. [Google Scholar]
- Garcia-Pausas, J.; Casals, P.; Camarero, L.; Huguet, C.; Sebastia, M.-T.; Thompson, R.; Romanya, J. Soil Organic Carbon Storage in Mountain Grasslands of the Pyrenees: Effects of Climate and Topography. Biogeochemistry 2007, 82, 279–289. [Google Scholar] [CrossRef]
- Zhu, G.; Shangguan, Z.; Hu, X.; Deng, L. Effects of Land Use Changes on Soil Organic Carbon, Nitrogen and Their Losses in a Typical Watershed of the Loess Plateau, China. Ecol. Indic. 2021, 133, 108443. [Google Scholar] [CrossRef]
- Elliott, E.T.; Palm, C.A.; Reuss, D.E.; Monz, C.A. Organic matter contained in soil aggregates from a tropical chronosequence: Correction for sand and light fraction. Agric. Ecosyst. Environ. 1991, 34, 443–451. [Google Scholar] [CrossRef]
- Bongiovanni, M.D.; Lobartini, J.C. Particulate Organic Matter, Carbohydrate, Humic Acid Contents in Soil Macro-and Microaggregates as Affected by Cultivation. Geoderma 2006, 136, 660–665. [Google Scholar] [CrossRef]
- Udom, B.E.; Ogunwole, J.O. Soil Organic Carbon, Nitrogen, and Phosphorus Distribution in Stable Aggregates of an Ultisol under Contrasting Land Use and Management History. J. Plant Nutr. Soil Sci. 2015, 178, 460–467. [Google Scholar] [CrossRef]
- Briedis, C.; de Moraes Sá, J.C.; Caires, E.F.; de Fátima Navarro, J.; Inagaki, T.M.; Boer, A.; Neto, C.Q.; de Oliveira Ferreira, A.; Canalli, L.B.; Dos Santos, J.B. Soil Organic Matter Pools and Carbon-Protection Mechanisms in Aggregate Classes Influenced by Surface Liming in a No-till System. Geoderma 2012, 170, 80–88. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; He, H.; Xie, H.; Yan, Y.; Zhu, P.; Ren, J.; Wang, L. Carbon and Nitrogen Pools in Different Aggregates of a Chinese Mollisol as Influenced by Long-Term Fertilization. J. Soils Sediments 2010, 10, 1018–1026. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic Matter and Water-Stable Aggregates in Soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Liao, H.; Long, J.; Li, J. Conversion of Cropland to Chinese Prickly Ash Orchard Affects Soil Organic Carbon Dynamics in a Karst Region of Southwest China. Nutr. Cycl. Agroecosyst. 2016, 104, 15–23. [Google Scholar] [CrossRef]
- Delelegn, Y.T.; Purahong, W.; Blazevic, A.; Yitaferu, B.; Wubet, T.; Göransson, H.; Godbold, D.L. Changes in Land Use Alter Soil Quality and Aggregate Stability in the Highlands of Northern Ethiopia. Sci. Rep. 2017, 7, 1–12. [Google Scholar]
- Wang, J.; Liu, L.; Qiu, X.; Wei, Y.; Li, Y.; Shi, Z. Contents of Soil Organic Carbon and Nitrogen in Water-Stable Aggregates in Abandoned Agricultural Lands in an Arid Ecosystem of Northwest China. J. Arid Land 2016, 8, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Tisdall, J.M. Possible Role of Soil Microorganisms in Aggregation in Soils. Plant Soil 1994, 159, 115–121. [Google Scholar] [CrossRef]
- Cao, R.; Chen, L.; Hou, X.; Lu, X.; Li, H. Nitrogen addition reduced carbon mineralization of aggregates in forest soils but enhanced in paddy soils in South China. Ecol. Process. 2021, 10, 45. [Google Scholar] [CrossRef]
- Dorji, T.; Field, D.J.; Odeh, I.O.A. Soil Aggregate Stability and Aggregate-associated Organic Carbon under Different Land Use or Land Cover Types. Soil Use Manag 2020, 36, 308–319. [Google Scholar] [CrossRef]
- Debasish-Saha; Kukal, S.S.; Bawa, S.S. Soil organic carbon stock and fractions in relation to land use and soil depth in the degraded shiwaliks hills of lower himalayas. Land Degrad. Dev. 2014, 25, 407–416. [Google Scholar] [CrossRef]
- Li, X.-G.; Li, F.-M.; Zed, R.; Zhan, Z.-Y. Soil Physical Properties and Their Relations to Organic Carbon Pools as Affected by Land Use in an Alpine Pastureland. Geoderma 2007, 139, 98–105. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustian, K. Soil Macroaggregate Turnover and Microaggregate Formation: A Mechanism for C Sequestration under No-Tillage Agriculture. Soil Biol. Biochem. 2000, 32, 2099–2103. [Google Scholar] [CrossRef]
- Chidozie, E.I.; Ifeanyi, I.F.; Johnbosco, O.M.; Onyekachi, I.A.; Anthony, C.C.; Obinna, O.M.; Glory, M.O. Assessment of Hydraulic Conductivity and Soil Quality of Similar Lithology under Contrasting Landuse and Land Cover in Humid Tropical Nigeria. Soil Environ. 2019, 38, 75–80. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecosystem | Soil Depth | ||
|---|---|---|---|
| 0–15 cm | 15–30 cm | 30–45 cm | |
| Evergreen forest | 102.53 a (63.22–156.72) | 80.80 a (51.97–136.30) | 72.85 a (35.62–103.40) |
| Deciduous forest | 71.55 b (53.56–110.90) | 57.57 b (41.33–94.45) | 41.68 b (29.22–64.12) |
| Scrubland | 56.59 d (35.31–90.56) | 43.53 c (22.33–83.55) | 29.38 c (10.36–68.87) |
| Forest plantation | 64.40 c (39.22–96.49) | 49.34 c (24.44–76.84) | 32.85 c (11.65–59.86) |
| Tea plantation | 31.90 e (13.00–54.89) | 25.98 d (11.55–50.58) | 13.65 d (5.95–38.72) |
| Cropland | 25.61 e (18.94–47.03) | 20.97 d (13.66–35.13) | 8.80 d (1.82–24.70) |
| Mean | 58.76 (37.21–92.77) | 46.37 (27.55–79.48) | 33.20 (15.77–59.95) |
| Land-Degradation Index | |||||
|---|---|---|---|---|---|
| S. No | Ecosystem | 0–15 cm | 15–30 cm | 30–45 cm | Mean |
| 1 | Evergreen | 0.00 | 0.00 | 0.00 | 0.00 |
| 2 | Deciduous | −30.21 | −28.75 | −42.78 | −33.91 |
| 3 | Scrub | −44.81 | −46.12 | −59.67 | −50.20 |
| 4 | Forest Plantation | −37.19 | −38.94 | −54.91 | −43.68 |
| 5 | Tea Plantation | −68.89 | −67.85 | −81.27 | −72.67 |
| 6 | Crop Land | −75.02 | −74.04 | −87.93 | −79.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jagadesh, M.; Srinivasarao, C.; Selvi, D.; Thiyageshwari, S.; Kalaiselvi, T.; Kumari, A.; Singh, S.K.; Lourdusamy, K.; Kumaraperumal, R.; Allan, V.; et al. Quantifying the Unvoiced Carbon Pools of the Nilgiri Hill Region in the Western Ghats Global Biodiversity Hotspot—First Report. Sustainability 2023, 15, 5520. https://doi.org/10.3390/su15065520
Jagadesh M, Srinivasarao C, Selvi D, Thiyageshwari S, Kalaiselvi T, Kumari A, Singh SK, Lourdusamy K, Kumaraperumal R, Allan V, et al. Quantifying the Unvoiced Carbon Pools of the Nilgiri Hill Region in the Western Ghats Global Biodiversity Hotspot—First Report. Sustainability. 2023; 15(6):5520. https://doi.org/10.3390/su15065520
Chicago/Turabian StyleJagadesh, M., Cherukumalli Srinivasarao, Duraisamy Selvi, Subramanium Thiyageshwari, Thangavel Kalaiselvi, Aradhna Kumari, Santhosh Kumar Singh, Keisar Lourdusamy, Ramalingam Kumaraperumal, Victor Allan, and et al. 2023. "Quantifying the Unvoiced Carbon Pools of the Nilgiri Hill Region in the Western Ghats Global Biodiversity Hotspot—First Report" Sustainability 15, no. 6: 5520. https://doi.org/10.3390/su15065520