
Microbial Enzyme Systems in the Production of Second Generation Bioethanol


1
Department of Microbiology, Panjab University, Chandigarh 160014, India
2
Department of Biotechnology, D.A.V. College, Chandigarh 160011, India
*
Author to whom correspondence should be addressed.
Sustainability 2023, 15(4), 3590; https://doi.org/10.3390/su15043590
Submission received: 18 December 2022
/
Revised: 31 January 2023
/
Accepted: 13 February 2023
/
Published: 15 February 2023
(This article belongs to the Special Issue Sustainable Development of Enzyme System for Industrial Applications)
Abstract
:The primary contributor to global warming has been the careless usage of fossil fuels. Urbanization’s threat to the depletion of these resources has made it necessary to find alternatives due to the rising demand. Four different forms of biofuels are now available and constitute a possible replacement for fossil fuels. The first generation of biofuels is generated from the edible portion of biomass, the second generation is made from the non-edible portion of biomass, the third generation is made from algal biomass, and the fourth generation is made using molecular biology to improve the algal strain. Second-generation biofuels are extremely important because they are derived from non-edible biomass, such as agricultural and agro-industrial wastes rich in cellulose, hemicellulose, pectin, and starch impregnated with lignin, and are hydrolyzed after delignification by physio-chemical or biological pretreatments using ligninases. The enzymes involved in the hydrolysis of feedstocks for the production of second-generation bioethanol, a highly acceptable biofuel, are discussed in this article. Furthermore, the article discusses various fermentation technologies as well as significant developments in second-generation biofuel production by combining various microbial enzyme systems.
1. Introduction
Society’s advancement has raised the standard of living and made jobs easier, but it has also resulted in environmental issues as a result of excessive use of automobiles, machines, and other items, which has contributed to the depletion of fossil fuel sources. Urban areas house 52.5% of the world’s population, with that figure expected to rise to 70% by 2050 [1]. This urbanization is causing excessive use of fossil fuels in the transportation sector. Cities contribute significantly to CO2 emissions and the indiscriminate use of fossil fuels has put their reserves at risk [2]. Annual global carbon dioxide emissions are increasing and are expected to reach 38,836.98 MT (Metric Ton) in 2025. China’s expected CO2 emissions in 2025 are 11,521.21 MT, the United States’ is 11,521.21 MT, India’s is 3158.37 MT, Canada’s is 699.65 MT, Brazil’s is 605.61 MT, Argentina’s is 207.44 MT, Germany’s is 738.77 MT, Turkey’s is 374.40 MT, Iran’s is 822.46 MT, Saudi Arabia’s is 714.08 MT, South Africa’s is 338.17 MT and Japan’s is 1091.78 MT [3]. This has prompted researchers all over the world to focus on environmentally friendly alternatives to fossil fuels. Biofuels are one type of such fuel that emits fewer GHGs over their entire life cycle [4]. A biofuel is any fuel that is made from plant biomass and can generate energy for use in a variety of ways [5]. For the production of biofuels and energy, biomass that primarily consists of starchy crops, such as cereals, and root tubers, sugary crops, such as sugar cane and beet, agricultural residues, such as grasses, straws, bagasse, and brans, forestry crops, wood processing residues, dedicated energy crops, and biodegradable municipal solid waste, can be used [6]. Solid, liquid, or gaseous biofuels are all possible. Wood, and refuse-derived fuel (RDF), are some examples of solid biofuels. Biodiesel, biomethanol, bioethanol, biobutanol, etc are examples of liquid biofuels, while biohydrogen and biomethane are examples of gaseous biofuels. Given the increasing global biofuel consumption trend, major research attention has been directed toward feasible and low-cost biofuel resources, as reported by research publications in the last 20 years around the world, particularly in Asia, Europe, and the United States [7].
According to the proposed sustainable development scenario, biofuels must meet 9% of total transportation fuel demand by 2030, up from 3% in 2018. Between 2010 and 2021, the use of modern bioenergy increased by about 7% per year on average, and it is on the rise. More efforts are required to accelerate modern bioenergy deployment to meet the Net Zero Scenario. Biofuel production is not increasing at a rate sufficient to meet this demand, and it grew 6% year on year in 2019, with an average of 3% growth expected over the next five years, leaving total production short of 10% by 2030 to meet the pace required for sustainable development [8]. Food crops account for the majority of biofuels produced. For better sustainability, advanced biofuel production using non-food feedstocks must improve and gain a significant share of total biofuel production. Scaling up the production of these biofuels to a commercial level will require a great deal of effort and innovative research. Bioethanol and biomass-to-liquid synthetic fuels are among the most important advanced biofuels because they can be produced using low-cost, abundantly available feedstocks such as agricultural and agro-industrial residues [9,10].
Traditional biofuels are made from edible feedstocks such as sugar cane juice, molasses, sugar beet juice, molasses, cereals such as corn, rice, barley, wheat, sorghum, and oils such as soybean oil and palm oil. These are known as first-generation biofuels. Advanced biofuels are made from non-edible parts of biomass and are classified into three types based on the type of substrate used in the production process: second, third, and fourth-generation biofuels. Waste biomass resources such as agricultural, agro-industrial, municipal solid waste, and forest residue are used in second-generation biofuels. Third-generation biofuels are primarily made from algal biomass, which can be used to produce a wide range of biofuels and other value-added products [11]. The fourth generation of biofuels is a newer type that uses synthetic biology tools to create electro fuels and photobiological solar fuels by converting solar energy directly into fuels [12,13]. Recent concerns about the production of first-generation biofuels caused by the conflict between food and fuel have prompted experts to investigate alternative biofuel production routes [14]. According to numerous reports, the cost of food ingredients has risen due to the production of first-generation bioethanol [15]. The primary reason for preferring second-generation biofuels over first-generation biofuels is the use of waste and inedible agricultural biomass as a substrate for fuel generation. Because of its abundance and underutilization in comparison to other natural resources, lignocellulosic, agro-industrial, and biodegradable municipal solid waste biomass is a promising feedstock for the production of biofuels.
Due to the extensive food versus fuel debate associated with first-generation biofuels, the emphasis has shifted to the production of second-generation biofuels because the feedstock is easily accessible and has a less significant impact on the food web, water resources, and ecosystem [16,17,18]. The current methods for producing second-generation bioethanol are neither cost-effective nor eco-friendly [19]. As a result, the entire manufacturing process must be improved to be environmentally friendly and to make the cost of the fuel produced competitive with other fuels already on the market [20,21]. Biologically mediated lignocellulosic biomass conversion into biofuels appears to be more promising. The primary goal of this article is to review the environmentally friendly approaches used in biofuel production, with a focus on the enzymes used in the production of second-generation bioethanol, a highly acceptable liquid biofuel.
2. Composition of Agricultural and Agro-Industrial Waste Biomass, the Feedstocks for Second Generation Bioethanol
The majority of plant waste biomass, also known as lignocellulosics, consists primarily of carbohydrates in the form of cellulose, hemicellulose, and phenolic polymers such as lignin. Starch, pectin, proteins, acids, salts, and minerals are also present in varying amounts in some agro-industrial and biodegradable municipal solid waste biomass residues [18]. The structural composition of some common lignocellulosic biomass residues with the potential to be used as feedstocks in the production of second-generation ethanol, also known as cellulosic ethanol, is shown in Table 1 and discussed along with the structural architecture of pectin and starch, commonly found in agro-industrial wastes such as brans, spent grains and kitchen waste residues, etc.
2.1. Cellulose (C6H10O5)n
One of the major constituents of plant cell walls which is abundantly available on earth is cellulose which exists as a fibrous structure. It is an unbranched long-chain polymer consisting of several repeated units of cellobiose which are linked to each other by β-1,4-glycosidic bonds [29]. These long chains of cellulose are linked together by Van der Waals and hydrogen bonds packing the cellulose into microfibrils which further bundle together to build cellulose fibers. The straightness of the chain is determined by the hydrogen bonds within these microfibrils. The crystalline and amorphous structures within the cellulose are introduced by interchain hydrogen bonding which imparts order or disorder to the cellulose structure [18].
2.2. Hemicellulose (C5H8O4)n
Hemicellulose is the second notable and prevalent polymer in plant waste. Being chemically heterogeneous sets it apart from cellulose. These pentoses (xylose, rhamnose, and arabinose), hexoses (glucose, mannose, and galactose), and uronic acids (4-o-methyl-glucuronic, D-glucuronic acids) are branching, heterogeneous polymers [29]. In various materials, hemicelluloses have varying proportions. For instance, conifers and hardwoods have widely different proportions and types of xylans and mannan. In conifers, galactoglucomananas (5–8%), arabinoglucouronoxilanes (7–15%), and glucomannan (10–15%) are the primary components, whereas glucomannans (2–5%) and glycoronoxilanes (15–35%) predominate in hardwoods. The primary hemicellulosic components of grass and cereal cell walls are arabinoxylans [18]. The general structure of hemicellulose is depicted in Figure 1.
2.3. Lignin
Lignin is the third polymer that is widely distributed in nature (Figure 2). This polymer, which is found in plant cell walls, gives the cell wall of the plant a strong defense against any microbial invasion. The major three forms of phenyl propane units found in lignin are guaiacyl propanol also known as coniferyl alcohol, syringyl alcohols also known as sinapyl alcohol, and p-hydroxyphenyl propanol also known as coumaryl alcohol. Lignin is primarily viewed as the glue that binds the various parts of lignocellulosic biomass together, making it water-insoluble. It is extremely challenging to hydrolyze biomass using enzymatic or microbiological processes because of how tightly lignin is bound to the cellulose structure [18,30].
2.4. Pectin
Only a limited amount of pectin can be found in plant cell walls. Pectins are heteropolysaccharides comprised of 1,4-linked units of α-D-galactosyluronic acid residues. Rhamnogalacturonan-I, homogalacturonan, and substituted galacturonans are the three main pectins that have been identified from plant cell walls [31]. The general structure of pectin is shown in Figure 3.
2.5. Starch
Glucose units in starch are connected by glycosidic linkages. Figure 4 shows the architecture of the two types of polymeric units that make it up: amylose and amylopectin. By α-1,4 glycosidic linkages, amylose is made up of linearly linked glucose units. Amylopectin is made up of linear glucose chains with an α-1,4 linkage that is joined to the side chains by α-1,6 linkage [32,33].
The preceding section summarised the various carbohydrates in the form of cellulose, hemicellulose, phenolic polymer, and other ingredients in lesser amounts such as pectin and starch that comprise the overall skeleton of agricultural and agro-industrial waste biomass used in the production of second-generation biofuels. Following the hydrolysis of polysaccharides into simpler sugar molecules to be converted into bioethanol, the phenolic polymers are disintegrated first for conversion into various value-added products. Many developed countries are investing heavily in microbial fermentation and product regeneration from lignocellulosic feedstock, which necessitates complete exploitation of the lignocellulosic biomass. Knowing the lignocellulosic biomass composition allows industries, researchers, and biorefineries to invest in and exploit microorganisms and their enzyme systems for the development of second-generation bioethanol.
3. Conversion of Agricultural and Agro-Industrial Residues into Bioethanol
The development of a biorefinery for the production of numerous value-added products, including second-generation biofuels from plant biomass waste, has been the subject of extensive research. Effective cellulose utilization is crucial for making use of lignocellulosic biomass because it produces sugars that can be fermented further. But because lignin acts as a significant barrier to the existing carbohydrates, pretreatment is an extremely important step in the processing of biomass to disrupt lignin and hemicellulose for efficient hydrolysis of cellulose in the following stage. Pretreatment, enzymatic hydrolysis, and fermentation are the three primary processes in the bioconversion of lignocellulosic biomass into bioethanol [33,34,35].
3.1. Pretreatment
The fundamental obstacle to the generation of biofuels seems to be the resistant and crystalline structure of plant biomass [36]. Enzyme interaction with cellulose is necessary for enzymatic hydrolysis, however, cellulose’s crystalline structure makes enzymatic attacks difficult. The lignin and hemicellulose matrices are another obstacle because they operate as physical barriers that reduce the accessibility of activated cellulose to enzymes. Additionally, lignin reduces the effectiveness of enzymes by binding cellulase [37]. Therefore, a pretreatment technique is needed to soften the crystalline structure of plant biomass before enzymatic hydrolysis [38]. The pretreatment process alters the structure and composition of the biomass and increases the surface area of the cellulose, making it more porous and more accessible for enzymatic hydrolysis [39,40,41].
3.1.1. Goal of Pretreatment
Different pretreatment techniques have been proposed and put into practice for the maximum release of fermentable sugars from lignocellulosic biomass to improve the enzymatic hydrolysis of biomass and fermentation yields [42,43].
Pretreatment must have the following qualities to be effective: (i) it must be economical and environmentally friendly, (ii) the most lignin can be eliminated, (iii) minimum production of phenols, furans, and furfurals, which prevents fermentation, (iv) recovering lignin to create other products with extra value, (v) minimal energy required, (vi) pretreatment chemicals must be recovered for future use, (vii) minimum costs of operation and minimum labor needs.
3.1.2. Factors Affecting the Choice of Pretreatment
3.1.3. Types of Pretreatments
Different types of pretreatment technologies have been studied so far and basically, four types of strategies have been categorized including (i) mechanical or physical involving mechanical milling and exposure to high temperature using steam (ii) chemical involving the use of acids, bases, oxidizing agents or ionic liquids alone or in combination with steam and are energy intensive (iii) physicochemical involving acid or ammonia explosion which are also energy intensive, (iv) biological involving the microorganisms or microbial enzyme systems for disrupting lignin and hemicellulose. Table 2 summarises different pretreatment methodologies and their effects on biomass. Because this manuscript is about enzyme systems for the production of second-generation biofuels, biological pretreatment is more relevant here and will be discussed in detail hereafter.
Biological Pretreatment
Biological pretreatment uses less energy and is less harmful to the environment than chemical and physical procedures. Natural diversity includes a variety of ligninolytic and hemicellulolytic microorganisms that can be used for the pretreatment of biomass [38]. Because they destroy lignin and hemicellulose with only a small amount of cellulose, a variety of white, brown, and soft rot fungi have been employed for biological pretreatment [57]. White-rot fungi degrade lignin due to the presence of lignin-degrading enzymes including peroxidases and laccases. With the aid of mediators, laccase can directly target the nonphenolic and phenolic subunits of lignocellulosic biomass, causing structural changes [58].
Some of the white-rot fungal species that have been investigated for the biological pretreatment of biomass include Pycnoporus cinnarbarinus. Phanerochaete chrysosporium, Cyathus stercolerus, Ceriporia lacerata, Ceriporiopsis subvermispora, Pleurotus ostreaus. Other basidiomycetes used for biological pretreatment include Fomes fomentarius, Ganoderma resinaceum, Lepista nuda, Irpex lacteus, Trametes versicolor, and Pycnoporus sanguineus [59,60,61,62,63,64]. The biological pretreatment of the biomass can be accomplished in three different ways, which include the use of enzymes, a consortium of microorganisms, or fungi that can degrade lignin [22]. Ma and Ruan [65] explored simultaneous delignification and hydrolysis of corn stover by co-culturing Coprinus comatus and Trichoderma reesei. A range of white-rot fungi was investigated in a study to discover the optimum biological pretreatment for corn stover, and Cyathus stercoreus NRRL-6573 produced the highest carbohydrate conversion [62]. Although biological pretreatment has advantages, it is not favored on an industrial scale because it is too sluggish [66]. Therefore, for biological pretreatment to be applied at the industrial level, it is necessary to discover more fungi that can delignify biomass but at faster rates. Rastogi et al. [67] observed that Pyrenophora phaeocomes S-1 cultivation on rice straw led to 63 and 51% lignin and hemicellulose breakdown, respectively. Further extraction of these components using a mild alkali revealed that the overall losses for lignin and hemicellulose were 78 and 60%, respectively. An increase in hydrolytic efficiency was seen in a study by Yan et al. [68] by using the Cupriavidus basilensis B-8 strain of bacteria in conjunction with diluted acid pretreatment. By forming pores in the biomass and removing the lignin droplets created by the acid treatment, the bacteria increased the surface area available for enzymatic action.
3.2. Hydrolysis to Release Free Sugars for Fermentation into Ethanol
Pretreatment is followed by hydrolysis of the pretreated substrate to saccharify it leading to the release of monomeric sugars. Hydrolysis can be performed by acid or enzymatic treatments.
3.2.1. Acid Hydrolysis
For a remarkably long time, diverse substrates have been hydrolyzed using acid. The two most frequently used acids are H2SO4 and HCl, which can be utilized in both diluted and concentrated forms and at varied concentrations. Dilute acid hydrolysis involves two processes. The first step in the process is the saccharification of carbohydrates, and if the reaction persists, sugars will then be converted to furfurals. Because cellulose breaks down more slowly than hemicellulose, a two-stage process is necessary to prevent the formation of furfurals from the sugars released from hemicellulose. The first stage of the process recovers the sugars from the hemicellulose under mild conditions, and stage two recovers the sugars from the cellulose under harsher conditions. The effective enzyme from Penicillium consortium and acid hydrolysis of poplar were also compared by Liang et al. [69], who concluded that the sugar yield from enzymatic hydrolysis is superior.
3.2.2. Enzymatic Hydrolysis
Since it does not result in the production of inhibitors, enzymatic hydrolysis of the pretreated substrate is preferred to acid hydrolysis. Furthermore, the enzymes contain no secondary reactions and work in a highly precise manner. By pretreating the substrate, cellulose and hemicellulose’s crystalline structure is broken down, allowing the enzymes to attack them and liberate sugars (Figure 5). Cellulases and hemicellulases are needed to break down cellulose and hemicellulose, which are the two main carbohydrates found in the cell wall structure [70]. The pretreated substrate must also include starch and pectin for amylases and pectinases, the corresponding enzymes, to fully saccharify the substrate.
Enzymatic hydrolysis has several benefits, such as high specificity, a higher sugar yield, milder reaction conditions, and a reduced formation of undesirable products [71]. Additionally, enzymatic saccharification offers a more cost-effective, environmentally friendly method for releasing sugars from lignocellulosic biomass.
Microbial Enzymes Involved in the Hydrolysis of Feedstocks for the Production of Second-Generation Bioethanol
Rice straw, wheat straw, corn stover, corn cobs, barley straw, sugarcane bagasse, rice husk, switchgrass, cotton stalks, and poplar biomass, among others, contain 30–48% cellulose and 15–30% hemicellulosic carbohydrates [72]. Cellulases and hemicellulases are thus essential for the efficient saccharification of these residues and the production of free sugars from them. Other agro-industrial residues, such as wheat bran, fruit peels, vegetable waste, rice bran, maize bran, and apple pomace, contain starch and pectin in addition to cellulose and hemicellulose [28,73,74]. As a result, amylases and pectinases are required for the hydrolysis of these biomass residues. Enzyme systems containing cocktails of various hydrolytic enzymes are required for complete and simultaneous hydrolysis of all carbohydrates in various feedstocks for the production of second-generation bioethanol. The following sections discuss the individual enzymes of various systems, along with their modes of action, required for the efficient hydrolysis of various polysaccharides in feedstocks for the generation of second-generation bioethanol.
Cellulases
The majority of the time, lignocellulosic biomass requires a combination of numerous enzymes, the most crucial of which are cellulases. Cellulases are classified structurally as glycosyl hydrolases, which hydrolyze cellulose’s β-1,4-D-glucan connections to create cellobiose and glucose [75]. To completely dissolve the cellulose framework, three enzymes must act together as depicted in Figure 6 and the role of various enzymes is as follows:
Endoglucanase or Endo-β-1,4-glucanase (EC 3.2.1.4): It makes short-chain oligomers containing non-reducing and reducing tails by randomly cutting the amorphous area of cellulose.
Cellobiohydrolase or Exo-β-1,4-glucanase (EC 3.2.1.91): Endoglucanase’s catalytic activity produces non-reducing endings that are hydrolyzed to produce cellobiose, a repetitive unit containing two glucose molecules.
Cellobiase or β-glucosidase (BG) (EC 3.2.1.21): To generate monomeric glucose units, it hydrolyzes cellobiose units.
Cellulose is the primary growth medium needed by the microbes that make cellulases, while they can also use other carbohydrates. Cellulase-producing microorganisms include fungi such as Aspergillus flavus, Aspergillus fumigatus, Aspergillus niger, Aspergillus oryzae, Fusarium oxysporum, Trichoderma viride [60,61,62,63,64,65].
Hemicellulases
The second most abundant polymer in nature is hemicellulose which comprises xylan, mannan, arabinan, and galactan. It is soluble in aqueous alkali but not in water or any chelating agent [76]. The enzyme market for hemicellulases is expanding quickly because these enzymes are used in a variety of industrial processes. The second-most prevalent carbohydrate in lignocellulosic is called xylan, which is a hetero-polysaccharide made up of 1,4-β-D-xylose monomers with different substituents [77]. Figure 7 shows the mode of action of xylanase for the breakdown of xylan [18]. When xylan is hydrolyzed by xylanase, oligosaccharides are produced, which are then hydrolyzed by 1,4-β-xylosidase to produce xylose [78]. For complete hydrolysis of xylans, other enzymes such as ferulic and p-coumaric esterases, xylan esterases, α-4-O-methyl glucoronosidases, and α-1-arabinofuranosidases work in concert [57].
In addition to xylanase, mannans, and heteromannanas are additional polysaccharides that are found in the hemicellulose of plant cell walls. D-mannose, a six-carbon sugar, makes up the majority of mannan, but because plant mannans have a complex and heterogeneous structure, it takes a combination of endo-1,4-β-mannanases, exo-mannosidases, and other enzymes to completely break them down [79]. These enzymes can also remove the side chain sugars that are present at various locations on mannans. The following enzymes are involved in the hydrolysis of different hemicellulosic structures. Xylan degradation is carried out by three different types of xylanases [80].
Endo-β-1,4-xylanase (EC 3.2.1.8): By hydrolyzing glycosidic linkages to release linear and branching oligosaccharides, it randomly splits the xylan chain.
Exo-β-1,4-xylanase or β -1,4-xylan xylohydrolase: It eliminates monomeric xylose units from the xylan polymer’s non-reducing terminus.
β-1,4-xylosidase or Xylobiase. (EC 3.2.1.37): This enzyme hydrolyzes disaccharides such as xylobiose and the higher xylooligosaccharides that have a lower specific affinity.
The following enzymes, whose modes of action are also shown in Figure 8, are considered to be involved in the hydrolysis of mannan and galactomannans by Moreira and Filho [81].
Endo-β-1,4-mannanase (EC 3.2.1.78): It generates new chain endpoints by randomly cleaving the mannan’s β-1,4-linkage internal links.
Exo-β-mannosidase (EC 3.2.1.25): It releases mannose sugar moieties by cleaving β-1,4-linked mannosides from the non-reducing ends of mannan and mannooligosaccharides.
β-glucosidase (EC 3.2.1.21): This enzyme hydrolyzes the 1,4-β-D-glucopyranose found at the non-reducing ends of the oligosaccharides produced from glucomannan and galactoglucomannan.
α-galactosidase (EC 3.2.1.22): It is a debranching enzyme that breaks down the α-1,6-linked D-galactopyranosyl side chains of galactomannan and galactoglucomannan.
Acetyl mannan esterase: It is a debranching enzyme that causes galactoglucomannan to release its acetyl groups.
Agaricus [82], Aspergillus [83,84], Fusarium [84,85], and Trichoderma [86,87,88,89] are fungi that have been discovered to break down hemicellulose. Hemicellulases are produced mostly by gram-positive bacteria, such as Bacillus species [90,91] and Clostridia species [92,93]. Among the actinomycetes, some species of Streptomycetes group [94].
Pectinases
Pectinases are the enzymes that hydrolyze pectic polysaccharides into monomers such as galacturonic acids. Pectin is a major component of plant cell walls, so to completely break down the lignocellulosic biomass, pectinases are required to completely hydrolyze the pectic materials. This lowers the viscosity of the medium and creates an ideal environment for the other enzymes to act on different polysaccharides. The following are the primary enzymes [95] involved in the hydrolysis of pectic substances:
Protopectinases: To liberate soluble form polymerized pectin, they dissolve protopectin. These are divided into two types: type A, which acts with protopectin at the polygalacturonic acid chain area, and type B, which acts with the polysaccharide chains tying the polygalacturonic acid chain to the components of the cell wall.
Pectin Methyl Esterases (PME) (EC 3.1.1.11): Pectin methyl esterases de-esterify the methyl group of pectin, releasing pectic acid and methanol in the process. Before pectate lyases and polygalacturonases, which require non-esterified substrates, it catalyzes de-esterification.
Pectin Acetyl Esterases (PAE): To liberate pectic acid and acetate, it catalyzes the hydrolysis of the acetyl esters found in pectin.
Polymethylgalacturonases (PMG): The pectin backbone’s α-1,4-glycosidic linkages are broken down, resulting in the formation of 6-methyl-D-galacturonate. It has both endo and exo modes of action. Exo-PMG catalyzes a reaction at the non-reducing end of the substrate while endo-PMG randomly cleaves the substrate.
Polygalacturonases (PG): To create D-galacturonate, it cleaves the polygalacturonic acid’s α-1,4-glycosidic linkages. It can act in both endo and exo modes, just as PMG. Exo-PG (EC 3.2.1.67) catalyzes the reaction at the non-reducing end of the substrate while endo-PG (EC 3.2.1.15) randomly cleaves the substrate.
Pectate Lyases (PGL): To release α-4,5-D-galacturonate from the glycosidic bonds in polygalacturonic acid, it performs a trans-elimination reaction. Exo-PGL (EC 4.2.2.9) cleaves the substrate at the nonreducing end, whereas endo-PGL (EC 4.2.2.2) operates on the substrate at random.
Pectin Lyases (PL): It performs trans elimination of glycosidic connections to randomly break the esterified pectin and create unsaturated methyloligogalacturonates.
Numerous bacteria and fungi that cause plant disease produce pectinolytic enzymes to aid in host invasion. Additionally, they aid in the recycling of carbon ponds in nature by decomposing dead plant materials. Numerous organisms have been shown to generate pectinolytic enzymes, including Aspergillus [96], Fusarium [97], Penicillium [98], Trichoderma [99], Bacillus, Erwinia, and actinomycetes such as Streptomycetes [100].
Amylases
The three main categories of amylases, also known as glycosyl hydrolases (GH), according to the International Union of Biochemistry and Molecular Biology (UIBMB), are endo-amylases, exo-amylases, and debranching enzymes. Figure 9 shows how all of these enzymes work to break down starch. The various types of starch-degrading enzymes are as follows [101].
Endoamylases or α-amylase (EC 3.2.1.1): It cleaves the α-1,4-bonds present in the inner regions of amylose and amylopectin to break into oligosaccharides and dextrins, decreasing the solution’s viscosity.
Exoamylase or β-amylase (EC 3.2.1.2): Only the α-1,4-bonds at the non-reducing ends are broken, releasing limit dextrins and β-maltose.
ƴ-amylase or Amyloglucosidase or Glucoamylase (EC 3.2.1.3): It functions as a debranching enzyme by cleaving the final α-1,4 links at the non-reducing end of amylase and amylopectin, which releases glucose.
Many fungi, bacteria, and actinomycetes have been found to produce amylases. Several species of the genera Aspergillus and Penicillium are effective fungal amylase producers. Aspergillus flavus, Aspergillus fumigatus, and Aspergillus niger are among the fungi that produce amylases [102,103,104]. Bacillus species are the most common types of the many bacteria that produce amylases. Rhodothermus, Corynebacterium, Geobacillus, Lactobacillus, and Pseudomonas are some more species. Streptomyces and Thermonospora have been discovered to make amylase among the actinomycetes [105].
Enzyme technology is typically regarded as the most environmentally friendly method of saccharification in any biorefinery. Using additional enzymes to allow for more extensive exploitation of plant biomass could result in processing that uses less energy and chemicals while recovering more fermentable sugar. Table 3 depicts the use of multiple hydrolytic enzymes produced by fungi and bacteria to aid in the process of polysaccharide bioconversions in various biomass residues for the production of second-generation bioethanol.
To completely hydrolyze biomass residues and produce second-generation bioethanol, a large number of enzymes are required. These enzymes are required for hydrolyzing a specific linkage at a specific phase in any biorefinery. Furthermore, the method of action provides any biorefinery with critical information for overcoming any flaws in the hydrolysis of any type of sugar or linkage between them. A microorganism with the ability to release a variety of enzymes involved in the hydrolysis of lignocellulosic biomass could be a candidate for use in a biorefinery producing second-generation biofuel.
4. Production of Microbial Enzymes for Use in the Generation of Second-Generation Bioethanol
Two different fermentation procedures can be used to produce enzymes at the industrial level while taking production costs and using natural substrates into account. There are two types of fermentation: liquid-state fermentation and solid-state fermentation.
4.1. Solid-State Fermentation (SSF)
For the growth of microorganisms, this type of fermentation often uses a moist solid substrate. SSF is a fermentation procedure that uses either a natural or inert solid substrate in the absence of freely flowing water [112,113]. A key component of SSF is the choice of solid material, which must be insoluble and serve as both a physical support and a source of nutrition for the bacteria. This imitates their natural environment and promotes the synthesis of enzymes and other useful metabolites for industry [114,115]. Due to the utilization of lignocellulosic as a medium or substrate for the development of microorganisms to create cellulases, hemicellulases, pectinases, and amylases, this fermentation is cost-effective. SSF cultures were discovered to produce more enzymes as compared to liquid cultures. SSF might be viewed as a superior method for the industrial synthesis of enzymes while taking into account production costs and employing natural substrates. Higher fermentation productivities, higher product stability, higher product concentration, decreased chances of contamination due to lower water activity need, and development of microorganisms specialized for water-insoluble substrates are all benefits of SSF [116]. Other benefits include the use of straightforward instrumentation, compactness of the fermenter due to a smaller volume of water, lack of foam formation, higher fermentation capacity, decreased catabolic repression, cost-effectiveness, and a reduced need for solvents in the product recovery process [117,118].
Various researchers have used different lignocellulosic, agro-industrial, and biodegradable municipal solid waste feedstocks to produce various hydrolytic enzyme systems important in the industries working in the field of second-generation biofuels using bacterial and fungal cultures. In an attempt to investigate the potential of Aspergillus niger CECT2088 on brewer’s spent grain for the production of cellulases and xylanases, Leite et al. [108] used brewer’s spent grain. Kaur et al. [119] used a natural variant of Aspergillus niger P-19 to produce a cellulase-hemicellulase consortium on rice straw for efficient and low-cost saccharification, whereas Chugh et al. [9] produced multiple carbohydrases including cellulases, hemicellulases, pectinases, and amylases through solid-state fermentation of de-oiled rice bran. Recently, our group developed an enzyme cocktail comprised of 19 hydrolytic enzymes for the generation of bioethanol from various lignocellulosic and agro-industrial waste biomass residues in solid, surface, and submerged state fermentation using a standardized kitchen waste-based medium [120]. More studies on optimizing various physical and cultural factors, as well as enzyme characterization, have been published in the literature to achieve the highest enzyme productivity and activity. Table 4 compiles some examples of solid-state fermentations for the production of various hydrolytic enzymes important in second-generation biofuels with significant breakthroughs.
4.2. Liquid State Fermentation (Submerged and Surface)
Under stationary or shaking circumstances, liquid-state fermentation involves the development of microorganisms in a liquid medium that contains the necessary nutrients. This type of fermentation is appealing for the development of microbes and the creation of products with added value due to several factors, including (a) homogenous distribution of nutrients for the proliferation of microorganisms; (b) simplicity of monitoring of variables such as moisture, temperature, pH, agitation, oxygen, and nutrient levels; (c) powerful technology that has already been best adapted with automatic grade and equipment availability. Cellulolytic enzymes, ligninolytic enzymes, and other beneficial metabolites can all be produced through liquid-state fermentation [135].
Liquid-state fermentation is divided into submerged and surface culture fermentation depending primarily on whether the incubation is being carried out in stationary or rocking circumstances. In surface culture, fermentation microorganisms develop on the shallow nutritional media’s surface, consume the nutrients necessary for their growth, and simultaneously release products into the medium. Since fungi are filamentous in nature and agitation might break their mycelia, segregating biomass from the liquid medium, this mode of fermentation does not call for agitation in the case of fungi [136].
However, surface culture fermentation has a lower bio-reaction rate and longer fermentation periods as compared to submerged fermentation, which involves robust aeration and agitation [137]. Submerged fermentation is preferred over surface culture fermentation as a result of this drawback. Through submerged cultivation, many strains of bacteria, yeast, fungus, and algae have been employed for fermentation. These methods of fermentation can use either synthetically manufactured or lignocellulosic biomass-produced fermentation media.
The fungal hyphae are not desiccated as a result of the continual immersion in a liquid medium during liquid-state fermentation, which is also the most effective, easiest to sterilize, and most cost-effective approach for producing bioagents in large quantities [138]. Except for high-density cultures, microorganisms are exposed to a fixed temperature throughout their life cycle. Additionally, oxygen availability to biomass can be regulated at a specific level of medium saturation. When compared to solid substrates, submerged culture has various benefits, including easier control of fermentation parameters such as pH and temperature, improved contamination control, and a lower labor and space demand. The nature and amplitude of forces in a bioreactor are studied using fermenters that offer the organism a low-shear environment. Surface culture fermentation is preferred to submerged fermentation for several reasons, including equipment expense, energy usage, aeration breakdown, improved productivity, and yield [139].
Elegbede et al. [140] synthesized in-house xylanases in submerged fermentation conditions using corn cob as the substrate. Irfan et al. [141] used a peanut shell to produce cellulases in a submerged fermentation process. Recent studies investigated the much-needed potential of the submerged fermentation process in the production of various hydrolytic enzymes involved in the production of second-generation biofuels. Table 5 depicts a few examples of liquid-state fermentation producing hydrolytic enzymes on various substrates, as well as a significant breakthrough in their production.
Enzymatic saccharification of lignocellulosic feedstock is followed by fermentation of the hydrolysate by suitable fermentative microorganisms for ethanol production. The hydrolysate produced by enzymes after saccharification of pretreated lignocellulosic feedstock contains a mixture of hexoses and pentoses, including glucose, mannose, xylose, arabinose, galactose, and some oligosaccharides. Saccharomyces cerevisiae, Pachysolen tannophilus, Escherichia coli, Zymomonas mobilis, Candida brassicae, Candida shehatae, Bacillus macerans, Clostridium sp., etc. are used to ferment these monomeric sugars to produce ethanol [157,158]. However, for an effective ethanol production method, the fermentative microorganism should be able to use a wide range of substrates, including pentoses and hexoses, and have high ethanol productivities, tolerance for high ethanol concentrations and inhibitors present in the hydrolysate [159,160]. So far, four types of fermentation have been studied: (i) separate hydrolysis and fermentation (SHF), (ii) simultaneous saccharification and fermentation (SSF), (iii) simultaneous saccharification and co-fermentation (SSCF), and (iv) Consolidated bioprocessing (CBP), and the key features of each are summarised in Table 6.
Many reports from around the world suggested that the use of enzymes in the conversion of lignocellulosic and other waste biomass residues to second-generation bioethanol provides a much-needed boost to this sector. Roberto et al. [162] reported SSF using a vertical ball mill reactor with a high loading of rice straw. The study concluded that feeding the substrate gradually at an initial load of 16% with 4% additions after 10 and 24 h using an inoculum level of 3 g/L resulted in a high ethanol concentration of 52.3 g/L. As a result, the findings demonstrated that a suitable fed-batch feeding strategy of biomass aids in overcoming the limitations of SSF in batch mode. Zhu et al. [163] used SSCF to ferment ethylenediamine-treated corn stover with Saccharomyces cerevisiae and xylose utilizing yeast, yielding 59.8 g/L ethanol at 42 °C. In a study by da Silva et al. [165], pretreatment used alkaline hydrogen peroxide, which efficiently removed lignin and hemicellulose from carnauba waste, yielding 57.49% and 56.13%, respectively. Chen et al. [166] obtained 72.3% ethanol yield on total sugar by co-fermenting with S. cerevisiae IPE005 in corn stover hydrolysate. In another study, using a statistical approach significantly increased sugar yields and the tool was successful in designing simple conditions of pre-treatment and hydrolysis of deoiled rice bran for maximum saccharification of all carbohydrates present in the substrate [9].
Recently, our group investigated the potential of biodegradable solid waste, primarily kitchen waste, as a feedstock for the production of second-generation biofuel. A multiple hydrolytic enzyme cocktail was created using 19 concentrated enzyme components with an enzymatic yield of 150–250 IU/mL of CMCase. 30–40 IU/mL of FPase, 25–35 IU/mL of Avicelase, 30–40 IU/mL of β-glucosidase, 135–145 IU/mL of cellobiase, 160–175 IU/mL of salicinase, 800–900 IU/mL of xylanase, 50–70 IU/mL of xylosidase, 260–275 IU/mL of mannanase, 25–35 IU/mL of mannosidase, 25–35 IU/mL of pectin-lyase, 25–35 IU/mL of polygalacturonase, 12,500–15,000 U/mL of α-amylase, 50–75 IU/mL of pullulanase, 400–500 IU/mL of glucoamylase, 140–165 IU/mL of α-glucosidase, 2100–2300 U/mL of protease, 190–210 U/mL of lipase and 190–210 U/mL of alginate lyase [120]. This breakthrough has paved the way for biodegradable solid waste to be used as a substrate for enzymes in second-generation biofuels.
Different countries use different feedstocks for bioethanol production based on regional availability, local climate, and economic drivers. Sugars and starches are the primary feedstocks for commercial bioethanol production. The ethanol produced in the United States and Brazil accounts for 85% of all bioethanol produced globally [167]. The United States, the world’s largest bioethanol producer, primarily uses corn as a feedstock, which is also used in China and Slovakia whereas Brazil, the world’s second-largest bioethanol producer, primarily uses sugarcane juice and molasses as a feedstock which are also employed in India, Indonesia, Brazil, China, Thailand, and Colombia. Wheat is generally used in Denmark, Austria, Germany, Canada, Belgium, France, and Russia [75]. Because most of these feedstocks compete with human feed, lignocellulosic biomass, as well as agro-industrial and biodegradable municipal solid waste residues, which are abundant and the most untapped natural reservoir on the planet, are promising feedstocks for second-generation bioethanol generation.
5. Conclusions and Future Outlook
The world’s increasing energy requirements as a result of urbanization, excessive use of fossil fuels, and the issue of disposing of agricultural waste residues are all scenarios that make the use of biofuels made from waste biomass an essential solution that can solve all of these problems. Second-generation ethanol production is significantly more expensive than first-generation ethanol, which uses existing technology for converting biomass to bioethanol, and it is difficult to predict when its cost will approach that of corn/sugarcane ethanol. Cellulosic ethanol’s superior environmental benefits require drastic cost reductions at all levels. The cost of pretreatment, enzymes for hydrolysis, fermentation of all sugars, and distillation, all significantly increase the final cost of producing cellulosic ethanol. Many countries around the world have launched Ethanol Blending Programmes to reduce their reliance on crude oil imports, reduce carbon emissions, and increase farmer income. Because of the coordinated efforts of the Public Sector Oil Marketing Companies, the program’s target of 10% blending has been met much ahead of the November 2022 deadline in India. The Government of India announced its ‘National Policy on Biofuels’ in 2018, with an indicative target of 20% ethanol blending in gasoline by 2030. However, given the encouraging performance and various interventions implemented by the government since 2014, the target of 20% ethanol blending has been pushed back from 2030 to 2025–26. In this context cellulosic ethanol and enzyme systems especially cellulases and hemicellulases are emerging as the stronger contenders to increase the indigenous production of second-generation bioethanol. Globally, research at all levels is currently being conducted to reduce the overall cost of the process. Furthermore, government-level incentives for second-generation ethanol and mandated ethanol blending into gasoline in several countries may pave the way for future bioethanol production from waste biomass.
The scientific community has switched to biofuels that are made from a variety of biomass residues, including municipal and agricultural waste, as a result of the rising cost of fossil fuels, the global warming caused by the careless use of these fuels, and the unscientific disposal of agricultural and agro-industrial waste residues. The commercial manufacture of bioethanol, which is now the highest-volume industrial fermentation product, generally uses sweet and starchy substrates. However, specialists are careful about their utilization due to the utility of such starchy residues as human nourishment. Even yet, many nations have established limitations on their permissible usage. Scientists are working to use agricultural, agroindustrial, and municipal solid waste as second-generation bioethanol feedstocks as the biofuel industry develops as a result of the rise in ethanol demand. These feedstocks are used by a small number of companies that pretreat and hydrolyze materials using chemical processes, which results in increased costs and significant chemical loading that eventually enters our life and environment. Enzymatic hydrolysis is advised, even though it adds between 30 and 50% to the overall cost of producing ethanol from lignocellulosic wastes. Enzymes with higher substrate specificity, lower dose requirements, and improved cost-effectiveness are required. The process economy as a whole can gain from the creation of innovative enzymes that can hydrolyze a variety of substrates, high-titer production of such enzymes, further development using genetic and molecular methods, and lower costs associated with the enzyme production process. Technologies that reuse the enzyme that washed away during hydrolysis can help address the issue of enzyme cost. The development of effective and environmentally friendly process technology for converting lignocellulosic residues to bioethanol may be made possible by advancements in enzyme technology and commercialization. This technology may prove to be a panacea for pressing global issues such as the depletion of fossil fuels and the improper disposal of these priceless resources.
Author Contributions
S.K.S.: Conceptualization, writing—review and editing, supervision; A.S.: investigation, writing—original draft preparation, writing—review and editing; R.S.: investigation, writing—original draft preparation, writing—review and editing, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Bank. Urban Population (% of Total). Available online: http://data.worldbank.org/indicator/SP.URB.TOTL.IN.ZS. (accessed on 15 January 2019).
- Leahy, S. City Emits 60% More Carbon than Thought. National Geographic. 6 March 2018. Available online: https://www.nationalgeographic.com/news/2018/03/city-consumption-greenhouse-gases-carbon-c40-spd/ (accessed on 16 November 2022).
- Qiao, W.; Lu, H.; Zhou, G.; Azimi, M.; Yang, Q.; Tian, W. A hybrid algorithm for carbon dioxide emissions forecasting based on improved lion swarm optimizer. J. Clean. Prod. 2020, 244, 118612. [Google Scholar] [CrossRef]
- Hanaki, K.; Portugal-Pereira, J. The effect of biofuel production on greenhouse gas emission reductions. In Biofuels and Sustainability; Springer: Tokyo, Japan, 2018; pp. 53–71. [Google Scholar]
- Moukamnerd, C.; Kawahara, H.; Katakura, Y. Feasibility study of ethanol production from food wastes by consolidated continuous solid-state fermentation. J. Sustain. Bioenerg. Syst. 2013, 3, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Xue, J.; Yang, Y.; Lee, S.W. Thermal properties and combustion-related problems prediction of agricultural crop residues. Energies 2021, 14, 4619. [Google Scholar] [CrossRef]
- Subramaniam, Y.; Masron, T.A. The impact of economic globalization on biofuel in developing countries. Energy Convers. Manag. 2021, 1, 100064. [Google Scholar] [CrossRef]
- IEA. Bioenergy; IEA: Paris, France, 2022; Available online: https://www.iea.org/reports/bioenergy (accessed on 14 November 2022).
- Chugh, P.; Kaur, J.; Soni, R.; Sharma, A.; Soni, S.K. A low-cost process for efficient hydrolysis of deoiled rice bran and ethanol production using an inhouse produced multi-enzyme preparation from Aspergillus niger P-19. J. Mater. Cycles Waste Manag. 2022, 25, 359–375. [Google Scholar] [CrossRef]
- Kim, J.R.; Karthikeyan, K.G. Effects of severe pretreatment conditions and lignocellulose-derived furan byproducts on anaerobic digestion of dairy manure. Bioresour. Technol. 2021, 340, 125632. [Google Scholar] [CrossRef]
- Soni, S.K.; Dhull, N.P.; Soni, R.; Sharma, A. Microbiofuels: The Sustainable Energy Source for the Future. In Genomic, Proteomics, and Biotechnology; CRC Press: Boca Raton, FA, USA, 2022; pp. 357–380. [Google Scholar]
- Ziolkowska, J.R. Biofuels technologies: An overview of feedstocks, processes, and technologies. In Biofuels for a More Sustainable Future; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar]
- Xie, Y.; Khoo, K.S.; Chew, K.W.; Devadas, V.V.; Phang, S.J.; Lim, H.R.; Show, P.L. Advancement of renewable energy technologies via artificial and microalgae photosynthesis. Bioresour. Technol. 2022, 363, 127830. [Google Scholar] [CrossRef]
- Arun, N.; Dalai, A.K. Environmental and socioeconomic impact assessment of biofuels from lignocellulosic biomass. In Lignocellulosic Biomass to Liquid Biofuels; Academic Press: Cambridge, MA, USA, 2020; pp. 283–299. [Google Scholar]
- Ahmed, J.O. The effect of biofuel crops cultivation on food prices stability and food security-A Review. Eurasian J. Biosci. 2020, 14, 613–621. [Google Scholar]
- Lima, D.R.S.; de Oliveira Paranhos, A.G.; Adarme, O.F.H.; Baêta, B.E.L.; Gurgel, L.V.A.; dos Santos, A.S.; de Aquino, S.F. Integrated production of second-generation ethanol and biogas from sugarcane bagasse pretreated with ozone. Biomass Convers. Biorefin. 2022, 12, 809–825. [Google Scholar] [CrossRef]
- Singh, A.; Prajapati, P.; Vyas, S.; Gaur, V.K.; Sindhu, R.; Binod, P.; Varjani, S. A comprehensive review of feed-stocks as sustainable substrates for next-generation biofuels. Bioenergy Res. 2022, 1–18. Available online: https://link.springer.com/article/10.1007/s12155-022-10440-2 (accessed on 21 November 2022).
- Machineni, L. Lignocellulosic biofuel production: Review of alternatives. Biomass Convers. Bioref. 2020, 10, 779–791. [Google Scholar] [CrossRef]
- Babu, S.; Rathore, S.S.; Singh, R.; Kumar, S.; Singh, V.K.; Yadav, S.K.; Wani, O.A. Exploring agricultural waste biomass for energy, food and feed production and pollution mitigation: A review. Bioresour. Technol. 2022, 360, 127566. [Google Scholar] [CrossRef]
- Mina, D.; Hadi, S.; Jalal, A. The incorporated environmental policies and regulations into bioenergy supply chain management: A literature review. Sci. Total Environ. 2022, 820, 153202. [Google Scholar]
- Santos, F.; Eichler, P.; de Queiroz, J.H.; Gomes, F. Production of second-generation ethanol from sugarcane. In Sugarcane Biorefinery, Technology and Perspectives; Academic Press: Cambridge, MA, USA, 2020; pp. 195–228. [Google Scholar]
- Oliva-Taravilla, A.; Moreno, A.D.; Demuez, M.; Ibarra, D.; Tomás-Pejó, E.; González-Fernández, C. Unraveling the effects of laccase treatment on enzymatic hydrolysis of steam-exploded wheat straw. Bioresour. Technol. 2015, 175, 209–215. [Google Scholar] [CrossRef]
- Khan, M.F.S.; Akbar, M.; Xu, Z.; Wang, H. A review on the role of pretreatment technologies in the hydrolysis of lignocellulosic biomass of corn stover. Biomass Bioenergy 2021, 155, 106276. [Google Scholar] [CrossRef]
- Verma, N.; Kumar, V. Impact of process parameters and plant polysaccharide hydrolysates in cellulase production by Trichoderma reesei and Neurospora crassa under wheat bran based solid state fermentation. Biotechnol. Rep. 2020, 25, e00416. [Google Scholar] [CrossRef]
- Partida-Sedas, G.; Montes-García, N.; Carvajal-Zarrabal, O.; López-Zamora, L.; Gómez-Rodríguez, J.; Aguilar-Uscanga, M.G. Optimization of hydrolysis process to obtain fermentable sugars from sweet sorghum bagasse using a Box–Behnken design. Sugar Tech. 2017, 19, 317–325. [Google Scholar] [CrossRef]
- Díaz-González, A.; Perez Luna, M.Y.; Ramírez Morales, E.; Saldaña-Trinidad, S.; Rojas Blanco, L.; de la Cruz-Arreola, S.; Robles-Ocampo, J.B. Assessment of the Pretreatments and Bioconversion of Lignocellulosic Biomass Recovered from the Husk of the Cocoa Pod. Energies 2022, 15, 3544. [Google Scholar] [CrossRef]
- Kou, L.; Song, Y.; Zhang, X.; Tan, T. Comparison of four types of energy grasses as lignocellulosic feedstock for the production of bio-ethanol. Bioresour. Technol. 2017, 241, 424–429. [Google Scholar] [CrossRef]
- Chugh, P.; Soni, R.; Soni, S.K. Deoiled rice bran: A substrate for co-production of a consortium of hydrolytic enzymes by Aspergillus niger P-19. Waste Biomass. Valor. 2016, 7, 513–525. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, L.; Liu, D. Biomass recalcitrance Part I: The chemical compositions and physical structures affecting the enzymatic hydrolysis of lignocellulose. Biofuel Bioprod, Bioref. 2012, 6, 465–482. [Google Scholar] [CrossRef]
- Prasad, V.; Siddiqui, L.; Mishra, P.K.; Ekielski, A.; Talegaonkar, S. Recent advancements in lignin valorization and biomedical applications: A patent review. Recent Pat. Nanotechnol. 2022, 16, 107–127. [Google Scholar] [CrossRef]
- Picot-Allain, M.C.N.; Ramasawmy, B.; Emmambux, M.N. Extraction, characterisation, and application of pectin from tropical and sub-tropical fruits: A review. Food Rev. Int. 2022, 38, 282–312. [Google Scholar] [CrossRef]
- Gunaratne, A.; Corke, H. Starch, Analysis of Quality. Ref. Modul. Food Sci. 2016, 3, 202–212. [Google Scholar]
- Saini, J.K.; Kaur, A.; Mathur, A. Strategies to enhance enzymatic hydrolysis of lignocellulosic biomass for biorefinery applications: A review. Bioresour. Technol. 2022, 360, 127517. [Google Scholar] [CrossRef]
- Lay, C.H.; Dharmaraja, J.; Shobana, S.; Arvindnarayan, S.; Priya, R.K.; Saratlae, R.; Kumar, G. Lignocellulose biohydrogen towards net zero emission: A review on recent developments. Bioresour. Technol. 2022, 364, 128084. [Google Scholar] [CrossRef]
- Periyasamy, S.; Isabel, J.B.; Kavitha, S.; Karthik, V.; Mohamed, B.A.; Gizaw, D.G.; Aminabhavi, T.M. Recent Advances in Consolidated Bioprocessing for Conversion of Lignocellulosic Biomass into Bioethanol-A Review. Chem. Eng. J. 2022, 453, 139783. [Google Scholar] [CrossRef]
- Sharma, A.; Aggarwal, N.K. Pretreatment Strategies: Unlocking of Lignocellulosic Substrate. In Water Hyacinth: A Potential Lignocellulosic Biomass for Bioethanol; Springer: Berlin/Heidelberg, Germany, 2020; pp. 37–49. [Google Scholar]
- Meng, X.; Yoo, C.G.; Li, M.; Ragauskas, A.J. Physicochemical structural changes of cellulosic substrates during enzymatic saccharification. J. Appl. Biotechnl. Bioeng. 2016, 1, 87–94. [Google Scholar]
- Mahmood, H.; Moniruzzaman, M.; Iqbal, T.; Khan, M.J. Recent advances in the pretreatment of lignocellulosic biomass for biofuels and value-added products. Curr. Opin. Green Sustain. Chem. 2019, 20, 18–24. [Google Scholar] [CrossRef]
- Mosier, N.; Wyman, C.E.; Dale, B.E.; Elander, R.; Lee, Y.Y.; Holtzapple, M.T. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol. 2005, 96, 673–686. [Google Scholar] [CrossRef]
- García, V.; Päkkilä, J.; Ojamo, H.; Muurinen, E.; Keiski, R.L. Challenges in biobutanol production: How to improve the efficiency? Renew. Sustain. Energ. Rev. 2011, 15, 964–980. [Google Scholar] [CrossRef]
- Sun, S.; Sun, S.; Cao, X.; Sun, R. The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Bioresour. Technol. 2016, 199, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.V.T.; Vergaram, P.; Carbajo, J.M.; Villar, J.C.; dos Santos Rocha, J.M.; de Sousa, M.D.G.V. Bioconversion of pine stumps to ethanol: Pretreatment and simultaneous saccharification and fermentation. Holzforschung 2020, 74, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Masran, R.; Zanirun, Z.; Bahrin, E.K.; Ibrahim, M.F.; Lai Yee, P.; Abd-Aziz, S. Harnessing the potential of ligninolytic enzymes for lignocellulosic biomass pretreatment. Appl. Microbiol. Biotechnol. 2016, 100, 5231–5246. [Google Scholar] [CrossRef] [PubMed]
- Gandam, P.K.; Chinta, M.L.; Pabbathi, N.P.P.; Baadhe, R.R.; Sharma, M.; Thakur, V.K.; Gupta, V.K. Second-generation bioethanol production from corncob–A comprehensive review on pretreatment and bioconversion strategies, including techno-economic and lifecycle perspective. Ind. Crops Prod. 2022, 186, 115245. [Google Scholar]
- Yoo, C.G.; Meng, X.; Pu, Y.; Ragauskas, A.J. The critical role of lignin in lignocellulosic biomass conversion and recent pretreatment strategies: A comprehensive review. Bioresour. Technol. 2020, 301, 122784. [Google Scholar] [CrossRef]
- Periyasamy, S.; Karthik, V.; Senthil Kumar, P.; Isabel, J.B.; Temesgen, T.; Hunegnaw, B.M.; Vo, D.V.N. Chemical, physical and biological methods to convert lignocellulosic waste into value-added products. A review. Environ. Chem. Lett. 2022, 20, 1129–1152. [Google Scholar] [CrossRef]
- Aftab, M.N.; Iqbal, I.; Riaz, F.; Karadag, A.; Tabatabaei, M. Different Pretreatment Methods of Lignocellulosic Biomass for Use in Biofuel Production. In Biomass for Bioenergy-Recent Trends and Future Challenges; Intechopen: London, UK, 2019; pp. 15–92. [Google Scholar]
- Gu, B.J.; Dhumal, G.S.; Wolcott, M.P.; Ganjyal, G.M. Disruption of lignocellulosic biomass along the length of the screws with different screw elements in a twin-screw extruder. Bioresour. Technol. 2019, 275, 266–271. [Google Scholar] [CrossRef]
- Kumari, D.; Chahar, P.; Singh, R. Effect of ultrasonication on biogas and ethanol production from rice straw pretreated with petha waste water and dairy waste water. Int. J. Curr. Eng. Sci. Res. 2018, 5, 65–73. [Google Scholar]
- Puligundla, P.; Oh, S.E.; Mok, C. Microwave-assisted pretreatment technologies for the conversion of lignocellulosic biomass to sugars and ethanol: A review. Carbon Lett. 2016, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mikulski, D.; Kłosowski, G.; Menka, A.; Koim-Puchowska, B. Microwave-assisted pretreatment of maize distillery stillage with the use of dilute sulfuric acid in the production of cellulosic ethanol. Bioresour. Technol. 2019, 278, 318–328. [Google Scholar] [CrossRef]
- Han, S.Y.; Park, C.W.; Endo, T.; Febrianto, F.; Kim, N.H.; Lee, S.H. Extrusion process to enhance the pretreatment effect of ionic liquid for improving enzymatic hydrolysis of lignocellulosic biomass. In Wood Science and Technology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–15. [Google Scholar]
- Meng, X.; Bhagia, S.; Wang, Y.; Zhou, Y.; Pu, Y.; Dunlap, J.R.; Shuai, L.; Ragauskas, A.J.; Yoo, C.G. Effects of the advanced organosolv pretreatment strategies on structural properties of woody biomass. Ind. Crops Prod. 2020, 146, 112144. [Google Scholar] [CrossRef]
- Yuan, Z.; Li, G.; Wei, W.; Wang, J.; Fang, Z. A comparison of different preextraction methods followed by steam pretreatment of bamboo to improve the enzymatic digestibility and ethanol production. Energy 2020, 196, 117156. [Google Scholar] [CrossRef]
- Sarker, T.R.; Pattnaik, F.; Nanda, S.; Dalai, A.K.; Meda, V.; Naik, S. Hydrothermal pretreatment technologies for lignocellulosic biomass: A review of steam explosion and subcritical water hydrolysis. Chemosphere 2021, 284, 131372. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Shen, F.; Yang, G.; Deng, S.; Long, L.; He, J.; Luo, L. Liquid hot water extraction followed by mechanical extrusion as a chemical-free pretreatment approach for cellulosic ethanol production from rigid hardwood. Fuel 2019, 252, 589–597. [Google Scholar] [CrossRef]
- Sanchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Curran, L.M.L.K.; Sale, K.L.; Simmons, B.A. Review of advances in the development of laccases for the valorization of lignin to enable the production of lignocellulosic biofuels and bioproducts. Biotechnol. Adv. 2021, 54, 107809. [Google Scholar] [CrossRef]
- Shi, J.; Chinn, M.S.; Sharma-Shivappa, R.R. Microbial pretreatment of cotton stalks by solid state cultivation of Phanerochaete chrysosporium. Bioresour Technol. 2008, 99, 6556–6564. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Verma, P. An overview of key pretreatment processes employed for bioconversion of lignocellulosic biomass into biofuels and value added products. Biotech 2013, 5, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Rabemanolontsoa, H.; Saka, S. Various pretreatments of lignocellulosics. Bioresour Technol. 2016, 199, 83–91. [Google Scholar] [CrossRef]
- Saha, B.C.; Qureshi, N.; Kennedy, G.J.; Cotta, M.A. Biological pretreatment of corn stover with white-rot fungus for improved enzymatic hydrolysis. Int. Biodeter. Biodegrad. 2016, 109, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.K.; Sharma, S. Recent updates on different methods of pretreatment of lignocellulosic feedstocks: A review. Bioresour. Bioprocess. 2017, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Hafid, H.S.; Baharuddin, A.S.; Mokhtar, M.N.; Omar, F.N.; Mohammed, M.A.; Wakisaka, M. Enhanced laccase production for oil palm biomass delignification using biological pretreatment and its estimation at biorefinary scale. Biomass Bioenergy 2021, 144, 105904. [Google Scholar] [CrossRef]
- Ma, K.; Ruan, Z. Production of a lignocellulolytic enzyme system for simultaneous biodelignification and saccharification of corn stover employing co-culture of fungi. Bioresour Technol. 2015, 175, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Soni, R.; Kaur, J.; Soni, S.K. Unravelling the capability of Pyrenophora phaeocomes S-1 for the production of ligno-hemicellulolytic enzyme cocktail and simultaneous bio-delignification of rice straw for enhanced enzymatic saccharification. Bioresour. Technol. 2016, 222, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Wang, Z.; Zhang, K.; Si, M.; Liu, M.; Chai, L. Bacteria-enhanced dilute acid pretreatment of lignocellulosic biomass. Bioresour. Technol. 2017, 245, 419425. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Q.; Wang, W.; Lin, C.S.K.; Hu, Y.; Qi, W. Enhancement of an efficient enzyme cocktail from Penicillium consortium on biodegradation of pretreated poplar. Chem. Eng. J. 2023, 452, 139352. [Google Scholar] [CrossRef]
- Bhat, M.K. Cellulases and related enzymes in biotechnology. Biotechnol. Adv. 2000, 18, 355–383. [Google Scholar] [CrossRef] [PubMed]
- Sadaf, A.; Khare, S.K. Production of Sporotrichum thermophile xylanase by solid state fermentation utilizing deoiled Jatropha curcas seed cake and its application in xylooligosachharide synthesis. Bioresour. Technol. 2014, 153, 126–130. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, M.; Upadhyay, S.N.; Mishra, P.K.; Ramteke, P.W. Biofuels from Protein-Rich Lignocellulosic Biomass: New Approach. In Sustainable Approaches for Biofuels Production Technologies; Springer: Cham, Switherland, 2019; pp. 83–92. [Google Scholar]
- Janveja, C.; Rana, S.S.; Soni, S.K. Kitchen waste residues as potential renewable biomass resources for the production of multiple fungal carbohydrases and second generation bioethanol. J. Technol. Innov. Renew. Energy. 2013, 2, 186–200. [Google Scholar]
- Venkatanagaraju, E.; Bharathi, N.; Sindhuja, R.H.; Chowdhury, R.R.; Sreelekha, Y. Extraction and Purification of Pectin from Agro-Industrial Wastes. In Pectins-Extraction, Purification, Characterization and Applications; Intechopen: London, UK, 2019. [Google Scholar]
- Gupta, A.; Verma, J.P. Sustainable bio-ethanol production from agro-residues: A review. Renew. Sustain. Energ. Rev. 2015, 41, 550–567. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Z.; Li, X.; Liu, X.; Fan, J.; Clark, J.H.; Hu, C. The production of furfural directly from hemicellulose in lignocellulosic biomass: A review. Catal. Today 2019, 319, 14–24. [Google Scholar] [CrossRef]
- Peralta, A.G.; Venkatachalam, S.; Stone, S.C.; Pattathil, S. Xylan epitope profiling: An enhanced approach to study organ development-dependent changes in xylan structure, biosynthesis, and deposition in plant cell walls. Biotechnol. Biofuels 2017, 10, 245. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Li, X.; Zhao, J. Production and characteristics of a novel Xylose-and Alkali-tolerant GH 43 β-xylosidase from Penicillium oxalicum for promoting hemicellulose degradation. Sci. Rep. 2017, 7, 11600. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, S.; Kaur, J. Microbial mannases: An overview of production and applications. Crit. Rev. Biotechnol. 2007, 27, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Bastawde, K.B. Xylan structure, microbial xylanases, and their mode of action. World J. Microb. Biotechnol. 1992, 8, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.R.S.; Filho, E.X.F. An overview of mannan structure and mannan-degrading enzyme systems. Appl. Microbiol. Biotechnol. 2008, 79, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Kabel, M.A.; Jurak, E.; Mäkelä, M.R.; De Vries, R.P. Occurrence and function of enzymes for lignocellulose degradation in commercial Agaricus bisporus cultivation. Appl. Microbiol. Biotechnol. 2017, 101, 4363–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Soren, J.P.; Mondal, J.; Rakshit, S.; Halder, S.K.; Mondal, K.C. Contemporaneous synthesis of multiple carbohydrate debranching enzymes from newly isolated Aspergillus fumigatus SKF-2 under solid state fermentation: A unique enzyme mixture for proficient saccharification of plant bioresources. Ind. Crops Prod. 2020, 150, 112409. [Google Scholar] [CrossRef]
- Zehra, M.; Syed, M.N.; Sohail, M. Banana Peels: A Promising Substrate for the Coproduction of Pectinase and Xylanase from Aspergillus fumigatus MS16. Pol. J. Microbiol. 2020, 69, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olajuyigbe, F.M.; Fatokun, C.O.; Oni, O.I. Effective Substrate Loading for Saccharification of Corn Cob and Concurrent Production of Lignocellulolytic Enzymes by Fusarium oxysporum and Sporothrix carnis. Curr. Biotechnol. 2019, 8, 109–115. [Google Scholar] [CrossRef]
- Ezeilo, U.R.; Lee, C.T.; Huyop, F.; Zakaria, I.I.; Wahab, R.A. Raw oil palm frond leaves as cost-effective substrate for cellulase and xylanase productions by Trichoderma asperellum UC1 under solid-state fermentation. J. Environ. Manag. 2019, 243, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Ezeilo, U.R.; Wahab, R.A.; Mahat, N.A. Optimization studies on cellulase and xylanase production by Rhizopus oryzae UC2 using raw oil palm frond leaves as substrate under solid state fermentation. Renew. Energy 2019, 156, 1301–1312. [Google Scholar] [CrossRef]
- Cekmecelioglu, D.; Demirci, A. Production of Cellulase and Xylanase Enzymes Using Distillers Dried Grains with Solubles (DDGS) by Trichoderma reesei at Shake-Flask Scale and the Validation in the Benchtop Scale Bioreactor. Waste Biomass Valor. 2020, 11, 6575–6584. [Google Scholar] [CrossRef]
- Yan, S.; Xu, Y.; Yu, X.W. Rational engineering of xylanase hyper-producing system in Trichoderma reesei for efficient biomass degradation. Biotechnol. Biofuels. 2021, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Blibech, M.; Farhat-Khemakhem, A.; Kriaa, M.; Aslouj, R.; Boukhris, I.; Alghamdi, O.A.; Chouayekh, H. Optimization of β-mannanase production by Bacillus subtilis US191 using economical agricultural substrates. Biotechnol. Prog. 2020, 36, 2989. [Google Scholar] [CrossRef]
- Yadav, A.; Ali, A.A.M.; Ingawale, M.; Raychaudhuri, S.; Gantayet, L.; Pandit, A. Enhanced co-production of pectinase, cellulase and xylanase enzymes from Bacillus subtilis ABDR01 upon ultrasonic irradiation. Proc. Biochem. 2020, 92, 197–203. [Google Scholar] [CrossRef]
- Khan, M.I.M.; Zafar, M.; Anwar, Z.; Imran, M. Effect of expression of additional catalytic domain on characteristics of Xylanase Z of Clostridium thermocellum. Biologia. 2019, 74, 1395–1403. [Google Scholar] [CrossRef]
- Hamann, P.R.; Gomes, T.C.; de MB Silva, L.; Noronha, E.F. Influence of lignin-derived phenolic compounds on the Clostridium thermocellum endo-β-1, 4-xylanase XynA. Proc. Biochem. 2020, 92, 1–9. [Google Scholar] [CrossRef]
- Sinjaroonsak, S.; Chaiyaso, T.; Aran, H. Optimization of Cellulase and Xylanase Productions by Streptomyces thermocoprophilus Strain TC13W Using Oil Palm Empty Fruit Bunch and Tuna Condensate as Substrates. Appl. Biochem. Biotechnol. 2019, 189, 76–86. [Google Scholar] [CrossRef]
- Pedrolli, D.B.; Monteiro, A.C.; Gomes, E.; Carmona, E.C. Pectin and pectinases: Production, characterization and industrial application of microbial pectinolytic enzymes. Open Biotechnol. J. 2009, 3, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Begum, G.; Munjam, S. Carbon and Nitrogen Sources Effect on Pectinase Synthesis by Aspergillus niger Under Submerged Fermentation. Biosci. Biotechnol. Res. Asia 2021, 18, 185–195. [Google Scholar] [CrossRef]
- Kumar, Y.S.; Varakumar, S.; Reddy, O.V. Production and optimization of polygalacturonase from mango (Mangifera indica L.) peel using Fusarium moniliforme in solid state fermentation. World J. Microbiol. Biotechnol. 2010, 26, 1973–1980. [Google Scholar] [CrossRef]
- Amin, F.; Mohsin, A.; Bhatti, H.N.; Bilal, M. Production, thermodynamic characterization, and fruit juice quality improvement characteristics of an Exo-polygalacturonase from Penicillium janczewskii. Biochim Biophy. Acta Proteins Proteom. 2020, 1868, 140379. [Google Scholar] [CrossRef]
- Siamphan, C.; Arnthong, J.; Tharad, S.; Zhang, F.; Yang, J.; Laothanachareon, T. Production of D-galacturonic acid from pomelo peel using the crude enzyme from recombinant Trichoderma reesei expressing a heterologous exopolygalacturonase gene. J. Clean. Prod. 2022, 331, 129958. [Google Scholar] [CrossRef]
- Zeni, J.; Cence, K.; Grando, C.E.; Tiggermann, L.; Colet, R.; Lerin, L.A.; Valduga, E. Screening of pectinase-producing microorganisms with polygalacturonase activity. Appl. Biochem. Biotechnol. 2011, 163, 383–392. [Google Scholar] [CrossRef]
- Saranraj, P.; Stella, D. Fungal amylase—A review. Int. J. Microbiol. Res. 2013, 4, 203–211. [Google Scholar]
- Adejuwon, A.O.; Tsygankova, V.A.; Alonge, O. Effect of cultivation conditions on activity of α-amylase from a tropical strain Aspergillus flavusLink. J. Microbiol. Biotechnol. Food Sci. 2021, 7, 571–575. [Google Scholar] [CrossRef]
- Bano, S.; Iqbal, S.; Siddiqui, K.; Abbasi, K. Purification and characterization of [beta]-galactosidase from Aspergillus fumigatus PCSIR-2013. Pak. J. Pharm. Sci. 2021, 34, 1333–1341. [Google Scholar]
- Bellaouchi, R.; Abouloifa, H.; Rokni, Y.; Hasnaoui, A.; Ghabbour, N.; Hakkou, A.; Bechchari, A.; Asehraou, A. Characterization and optimization of extracellular enzymes production by Aspergillus niger strains isolated from date by-products. J. Genet. Eng. Biotechnol. 2021, 19, 50. [Google Scholar] [CrossRef]
- Gopinath, S.C.; Anbu, P.; Arshad, M.M.; Lakshmipriya, T.; Voon, C.H.; Hashim, U.; Chinni, S.V. Biotechnological process in microbial amylase production. BioMed Res. Int. 2017, 2017, 1272193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanram, S.; Amat, D.; Choudhary, J.; Arora, A.; Nain, L. Novel perspectives for evolving enzyme cocktails for lignocellulose hydrolysis in biorefineries. Sustain. Chem. Process. 2013, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sadh, P.K.; Chawla, P.; Bhandari, L.; Duhan, J.S. Bio-enrichment of functional properties of peanut oil cakes by solid state fermentation using Aspergillus oryzae. J. Food Meas. Character. 2017, 12, 622–633. [Google Scholar] [CrossRef]
- Leite, P.; Silva, C.; Salgado, J.M.; Belo, I. Simultaneous production of lignocellulolytic enzymes and extraction of antioxidant compounds by solid-state fermentation of agro-industrial wastes. Ind. Crops Prod. 2019, 137, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Marzo, C.; Díaz, A.B.; Caro, I.; Blandino, A. Valorization of agro-industrial wastes to produce hydrolytic enzymes by fungal solid-state fermentation. Waste Manag. Res. 2019, 37, 149–156. [Google Scholar] [CrossRef]
- Amande, T.; Adebayo-Tayo, B.; Ndubuisi-Nnaji, U.; Ado, B. Production and partial characterization of pectinases from mango peels by Aspergillus tamarii. J. Microbiol. Biotechnol. Food Sci. 2020, 9, 59–62. [Google Scholar]
- Melnichuk, N.; Braia, M.J.; Anselmi, P.A.; Meini, M.R.; Romanini, D. Valorization of two agroindustrial wastes to produce alpha-amylase enzyme from Aspergillus oryzae by solid-state fermentation. Waste Manag. 2020, 106, 155–161. [Google Scholar] [CrossRef]
- Iqbal, H.M.N.; Ahmed, I.; Zia, M.A.; Irfan, M. Purification and characterization of the kinetic parameters of cellulase produced from wheat straw by Trichoderma viride under SSF and its detergent compatibility. Adv. Biosci. Biotechnol. 2011, 2, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Farinas, C.S. Developments in solid-state fermentation for the production of biomass-degrading enzymes for the bioenergy sector. Renew. Sustain. Energ. Rev. 2015, 52, 179–188. [Google Scholar] [CrossRef]
- Nene, S.N.; Joshi, K.S. A comparative study of production of hydrophobin like proteins (HYD-LPs) in submerged liquid and solid state fermentation from white rot fungus Pleurotus ostreatus. Biocatal. Agric. Biotechnol. 2020, 23, 101440. [Google Scholar]
- Barrios-González, J. Secondary Metabolites Production: Physiological Advantages in Solid-State Fermentation. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 257–283. [Google Scholar]
- El-Bakry, M.; Abraham, J.; Cerda, A.; Barrena, R.; Ponsa, S.; Gea, T.; Sánchez, A. From wastes to high value added products: Novel aspects of SSF in the production of enzymes. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1999–2042. [Google Scholar] [CrossRef] [Green Version]
- Rudakiya, D.M. Strategies to improve solid-state fermentation technology. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 155–180. [Google Scholar]
- Kaur, J.; Chugh, P.; Soni, R.; Soni, S.K. A low-cost approach for the generation of enhanced sugars and ethanol from rice straw using in-house produced cellulase-hemicellulase consortium from A. niger P-19. Bioresour. Technol. Rep. 2020, 11, 100469. [Google Scholar] [CrossRef]
- Soni, S.K.; Sharma, A.; Soni, R. Fungal cocktail of multiple hydrolytic enzymes and method of production thereof. Indian Patent 202213059023, 2022. Available online: https://ipindiaservices.gov.in/PatentSearch/PatentSearch/ViewApplicationStatus (accessed on 21 November 2022).
- Kalogeris, E.; Christakopoulos, P.; Katapodis, P.; Alexiou, A.; Vlachou, S.; Kekos, D.; Macris, B.J. Production and characterization of cellulolytic enzymes from the thermophilic fungus Thermoascus aurantiacus under solid state cultivation of agricultural wastes. Process Biochem. 2003, 38, 1099–1104. [Google Scholar] [CrossRef]
- Negi, S.; Banerjee, R. Optimization of extraction and purification of glucoamylase produced by A. awamori in solid state fermentation. Biotechnol. Bioprocess. Eng. 2009, 14, 60–66. [Google Scholar] [CrossRef]
- Prasoulas, G.; Gentikis, A.; Konti, A.; Kalantzi, S.; Kekos, D.; Mamma, D. Bioethanol Production from Food Waste Applying the Multienzyme System Produced On-Site by Fusarium oxysporum F3 and Mixed Microbial Cultures. Fermentation 2020, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Grover, A.; Maninder, A.; Sarao, L.K. Production of fungal amylase and cellulase enzyme via solid state fermentation using Aspergillus oryzae and Trichoderma reesei. Int. J. Adv. Res. Technol. 2013, 2, 108–124. [Google Scholar]
- Rana, S.S.; Janveja, C.; Soni, S.K. Brewer’s spent grain as a valuable substrate for low cost production of fungal cellulases by statistical modeling in solid state fermentation and generation of cellulosic ethanol. Int. J. Food. Ferment. Technol. 2013, 3, 41–55. [Google Scholar] [CrossRef]
- Kaur, P.S.; Kaur, S.; Kaur, H.; Sharma, A.; Raj, P.; Panwar, S. Solid substrate fermentation using agro industrial waste: New approach for amylase production by Bacillus licheniformis. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 712–717. [Google Scholar]
- Mahalakshmi, N.; Jayalakshmi, S. Amylase, cellulase and xylanase production from a novel bacterial isolate Achromobacter xylosoxidans isolated from marine environment. Int. J. Adv. Res. Biol. Sci. 2016, 3, 230–233. [Google Scholar]
- Marín, M.; Artola, A.; Sánchez, A. Optimization of down-stream for cellulases produced under solid-state fermentation of coffee husk. Waste Biomass Valor. 2019, 10, 2761–2772. [Google Scholar] [CrossRef] [Green Version]
- Marín, M.; Sánchez, A.; Artola, A. Production and recovery of cellulases through solid-state fermentation of selected lignocellulosic wastes. J. Clean. Prod. 2019, 209, 937–946. [Google Scholar] [CrossRef]
- Salomão, G.S.; Agnezi, J.C.; Paulino, L.B.; Hencker, L.B.; de Lira, T.S.; Tardioli, P.W.; Pinotti, L.M. Production of cellulases by solid state fermentation using natural and pretreated sugarcane bagasse with different fungi. Biocatal. Agric. Biotechnol. 2019, 17, 1–6. [Google Scholar] [CrossRef]
- Teles, A.S.; Chávez, D.W.; Oliveira, R.A.; Bon, E.P.; Terzi, S.C.; Souza, E.F.; Tonon, R.V. Use of grape pomace for the production of hydrolytic enzymes by solid-state fermentation and recovery of its bioactive compounds. Food Res. Int. 2019, 120, 441–448. [Google Scholar] [CrossRef]
- Almanaa, T.N.; Vijayaraghavan, P.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Alyahya, S.A. Solid state fermentation of amylase production from Bacillus subtilis D19 using agro-residues. J. King Saud. Univ. Sci. 2020, 32, 1555–1561. [Google Scholar] [CrossRef]
- Jovanović, M.; Vučurović, D.; Bajić, B.; Dodić, S.; Vlajkov, V.; Jevtić-Mučibabić, R. Optimization of the simultaneous production of cellulase and xylanase by submerged and solid-state fermentation of wheat chaff. J. Serb. Chem Soc. 2020, 85, 177–189. [Google Scholar] [CrossRef]
- Rodrigues, I.D.; Barreto, J.T.; Moutinho, B.L.; Oliveira, M.M.; da Silva, R.S.; Fernandes, M.F.; Fernandes, R.P.M. Production of xylanases by Bacillus sp. TC-DT13 in solid state fermentation using bran wheat. Prep. Biochem. Biotechnol. 2020, 50, 91–97. [Google Scholar]
- Khade, S.M.; Srivastava, S.K.; Kumar, K.; Sharma, K.; Goyal, A.; Tripathi, A.D. Optimization of clinical uricase production by Bacillus cereus under submerged fermentation, its purification and structure characterization. Process. Biochem. 2018, 75, 49–58. [Google Scholar] [CrossRef]
- Letti, L.A.; Vítola, F.M.; de Melo Pereira, G.V.; Karp, S.G.; Medeiros, A.B.; da Costa, E.S.; Soccol, C.R. Solid-State Fermentation for the Production of Mushrooms. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 285–318. [Google Scholar]
- Rodrigues, A.C.; Fontão, A.I.; Coelho, A.; Leal, M.; da Silva, F.A.; Wan, Y.; Dourado, F.; Gama, M. Response surface statistical optimization of bacterial nanocellulose fermentation in static culture using a low-cost medium. New Biotechnol. 2019, 49, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, S.; Chen, G.; Wu, R.; Cao, X.; Zeng, W.; Liang, Z. Non-sterile submerged fermentation of fibrinolytic enzyme by marine Bacillus subtilis harboring antibacterial activity with starvation strategy. Front. Microbiol. 2019, 10, 1025. [Google Scholar] [CrossRef] [PubMed]
- Darouneh, E.; Alavi, A.; Vosoughi, M.; Arjmand, M.; Seifkordi, A.; Rajabi, R. Citric acid production: Surface culture versus submerged culture. Afr. J. Microbiol. Res. 2009, 3, 541–545. [Google Scholar]
- Elegbede, J.A.; Lateef, A. Valorization of Corn-Cob by Fungal Isolates for Production of Xylanase in Submerged and Solid State Fermentation Media and Potential Biotechnological Applications. Waste Biomass Valor. 2018, 9, 1273. [Google Scholar] [CrossRef]
- Irfan, M.; Bakhtawar, J.; Shakir, H.A.; Khan, M.; Ali, S. Utilization of peanut shells as substrate for cellulase production in submerged fermentation through Box-Behnken Design. Int. J. Biol. Chem. 2020, 12, 28–39. [Google Scholar] [CrossRef]
- Fawole, O.B.; Odunfa, S.A. Some factors affecting production of pectic enzymes by Aspergillus niger. Int. Biodeterior. Biodegrad. 2003, 52, 223–227. [Google Scholar] [CrossRef]
- Krough, K.B.R.; Morkeberg, A.; Jorgensen, H.; Frisvad, J.C.; Olsson, L. Screening genus Penicillium for producers of cellulolytic and xylanolytic enzymes. Appl. Biochem. Biotechnol. 2004, 114, 389–401. [Google Scholar] [CrossRef]
- Elisashvili, V.; Penninckx, M.; Kachlishvili, E.; Asatiani, M.; Kvesitadze, G. Use of Pleurotus dryinus for lignocellulolytic enzymes production in submerged fermentation of mandarin peels and tree leaves. Enzym. Microb. Technol. 2006, 38, 998–1004. [Google Scholar] [CrossRef]
- Vidyalakshmi, R.; Paranthaman, R.; Indhumathi, J. Amylase production on submerged fermentation by Bacillus spp. World J. Chem. 2009, 4, 89–91. [Google Scholar]
- de Castro, A.M.; Pedro, K.C.N.R.; da Cruz, J.C.; Ferreira, M.C.; Leite, S.G.F.; Pereira, N. Trichoderma harzianum IOC-4038: A promising strain for the production of a cellulolytic complex with significant β-glucosidase activity from sugarcane bagasse cellulignin. Appl. Biochem. Biotechnol. 2010, 162, 2111–2122. [Google Scholar] [CrossRef]
- de Almeida, M.N.; Guimarães, V.M.; Bischoff, K.M.; Falkoski, D.L.; Pereira, O.L.; Gonçalves, D.S. Cellulases and hemicellulases from endophytic Acremonium species and its application on sugarcane bagasse hydrolysis. Appl. Biochem. Biotechnol. 2011, 165, 594–610. [Google Scholar] [CrossRef]
- Nagar, S.; Gupta, V.K.; Kumar, D.; Kumar, L.; Kuhad, R.C. Production and optimization of cellulase-free, alkali-stable xylanase by Bacillus pumilus SV-85S in submerged fermentation. J. Ind. Microbiol. Biotechnol. 2010, 37, 71–83. [Google Scholar] [CrossRef]
- Kavuthodi, B.; Sebastian, D. Biotechnological valorization of pineapple stem for pectinase production by Bacillus subtilis BKDS1: Media formulation and statistical optimization for submerged fermentation. Biocatal. Agricl. Biotechnol. 2018, 16, 715–722. [Google Scholar] [CrossRef]
- Ismail, S.A.; Khattab, O.K.H.; Nour, S.A.; Awad, G.E.; Abo-Elnasr, A.A.; Hashem, A.M. A Thermodynamic Study of Partially-Purified Penicillium humicola β-mannanase Produced by Statistical Optimization. Jordan J. Biol. Sci. 2019, 12, 209–217. [Google Scholar]
- Sharma, D.; Mahajan, R. Development of Methodology for Concurrent Maximum Production of Alkaline Xylanase–Pectinase Enzymes in Short Submerged Fermentation Cycle. Waste Biomass Valor. 2019, 11, 6065–6072. [Google Scholar] [CrossRef]
- Arekemase, M.O.; Omotosho, I.O.; Agbabiaka, T.O.; Ajide-Bamigboye, N.T.; Lawal, A.K.; Ahmed, T. Optimization of bacteria pectinolytic enzyme production using banana peel as substrate under submerged fermentation. Sci. World J. 2020, 15, 56–63. [Google Scholar]
- Kote, N.V.; Manjula, A.C.; Vishwanatha, T.; Aravind, G.P. Production, Partial Purification and Characterisation of α-Amylase from Aspergillus niger using Aqueous Two Phase System (ATPS). Res. J. Biotechnol. 2020, 15, 5. [Google Scholar]
- Liu, J.; Yang, J.; Wang, R.; Liu, L.; Zhang, Y.; Bao, H.M. Comparative characterization of extracellular enzymes secreted by Phanerochaete chrysosporium during solid-state and submerged fermentation. Int. J. Biol. Macromol. 2020, 152, 288–294. [Google Scholar] [CrossRef]
- Stephen, A.C.; Adeniyi, O.A.; Hadiza, J. Effect of optimization conditions on submerged fermentation of corn bran for the production of xylanase enzyme. World J. Adv. Res. Rev. 2020, 5, 19–25. [Google Scholar] [CrossRef]
- Thite, V.S.; Nerurkar, A.S.; Baxi, N.N. Optimization of concurrent production of xylanolytic and pectinolytic enzymes by Bacillus safensis M35 and Bacillus altitudinis J208 using agro-industrial biomass through Response Surface Methodology. Sci. Rep. 2020, 10, 3824. [Google Scholar] [CrossRef] [Green Version]
- Girio, F.M.; Fonseca, C.; Carvalheiro, F.; Duarte, C.L.; Marques, S.; Bogel-qukasik, R. Hemicelluloses for fuel ethanol: A review. Bioresour. Technol. 2010, 101, 4775–4800. [Google Scholar] [CrossRef]
- Melendez, J.R.; Mátyás, B.; Hena, S.; Lowy, D.A.; El Salous, A. Perspectives in the production of bioethanol: A review of sustainable methods, technologies, and bioprocesses. Renew. Sustain. Energy Rev. 2022, 160, 112260. [Google Scholar] [CrossRef]
- Sarkar, N.; Aikat, K. Aspergillus fumigatus NITDGPKA3 provides for increased cellulase production. Int. J. Chem. Eng. 2014, 5, 959845. [Google Scholar]
- Tayeh, A.H.; Naim, N.; Carlos, D.; Ahmed, T.; Hassan, A. Potential of bioethanol production from olive mill solid wastes. Bioresour. Technol. 2014, 152, 24–30. [Google Scholar] [CrossRef]
- Sarbishei, S.; Goshadrou, A.; Hatamipour, M.S. Mild sodium hydroxide pretreatment of tobacco product waste to enable efficient bioethanol production by separate hydrolysis and fermentation. Biomass Convers. Biorefin. 2021, 11, 2963–2973. [Google Scholar] [CrossRef]
- Roberto, I.C.; Castro, R.C.; Silva, J.P.; Mussatto, S.I. Ethanol Production from High Solid Loading of Rice Straw by Simultaneous Saccharification and Fermentation in a Non-Conventional Reactor. Energies 2020, 13, 2090. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Zong, Q.J.; Li, W.C.; Chai, M.Z.; Xu, T.; Liu, H.; Fan, H.; Li, B.Z.; Yuan, Y.J. Temperature profiled simultaneous saccharification and co-fermentation of corn stover increases ethanol production at high solid loading. Energy Convers. Manag. 2020, 205, 112344. [Google Scholar] [CrossRef]
- Pandiyan, K.; Singh, A.; Singh, S.; Saxena, A.K.; Nain, L. Technological interventions for utilization of crop residues and weedy biomass for second generation bio-ethanol production. Renew. Energy 2019, 132, 723–741. [Google Scholar] [CrossRef]
- da Silva, F.L.; Dos Santos, D.A.; de Oliveira, C.A.; Magalhães, E.R.B.; Dos Santos, E.S. Evaluation of blend production of cellulases and xylanases Using pretreated and recycled carnauba straw. Appl. Biochem. Biotechnol. 2022, 194, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Fu, X. Industrial technologies for bioethanol production from lignocellulosic biomass. Renew. Sustain. Energy Rev. 2016, 57, 468–478. [Google Scholar] [CrossRef]
- Bhatt, S.M.; Bal, J.S. Bioprocessing perspective in biorefineries. In Sustainable Approaches for Biofuels Production Technologies; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–23. [Google Scholar]
Figure 1.
Generalized structure of hemicellulose (Xylan type) (Modified from [18]).
Figure 1.
Generalized structure of hemicellulose (Xylan type) (Modified from [18]).

Figure 2.
Generalized structure of lignin (Modified from [18]).
Figure 2.
Generalized structure of lignin (Modified from [18]).

Figure 3.
Generalized structure of pectin (Modified from [31]).
Figure 3.
Generalized structure of pectin (Modified from [31]).

Figure 4.
Generalized structure of starch.

Figure 5.
Schematic representation of pretreatment of lignocellulosic residue (Modified from [21]).
Figure 5.
Schematic representation of pretreatment of lignocellulosic residue (Modified from [21]).

Figure 6.
Mode of action of enzymes involved in the breakdown of cellulose (Modified from [21]).
Figure 6.
Mode of action of enzymes involved in the breakdown of cellulose (Modified from [21]).

Figure 7.
Mode of action of enzymes involved in the breakdown of xylan (Modified from [18]).
Figure 7.
Mode of action of enzymes involved in the breakdown of xylan (Modified from [18]).

Figure 8.
Schematic representation of O-acetylated galacto-glucomannan and enzymes involved in its degradation and the oligosaccharides released.
Figure 8.
Schematic representation of O-acetylated galacto-glucomannan and enzymes involved in its degradation and the oligosaccharides released.

Figure 9.
Generalized mode of action of amylases.


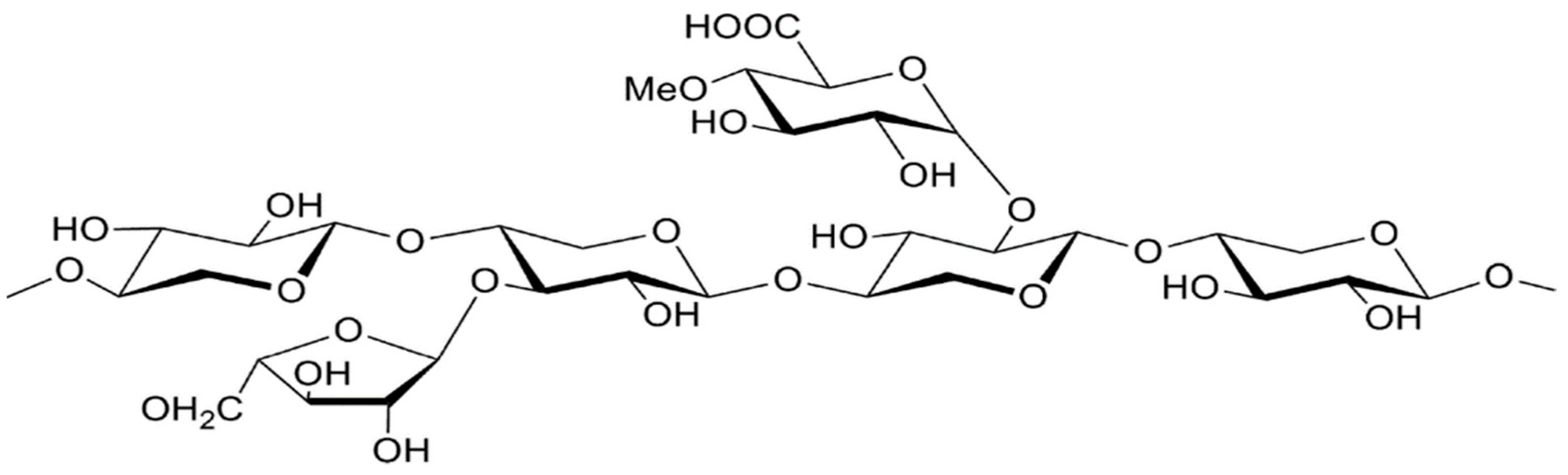{kind=link}
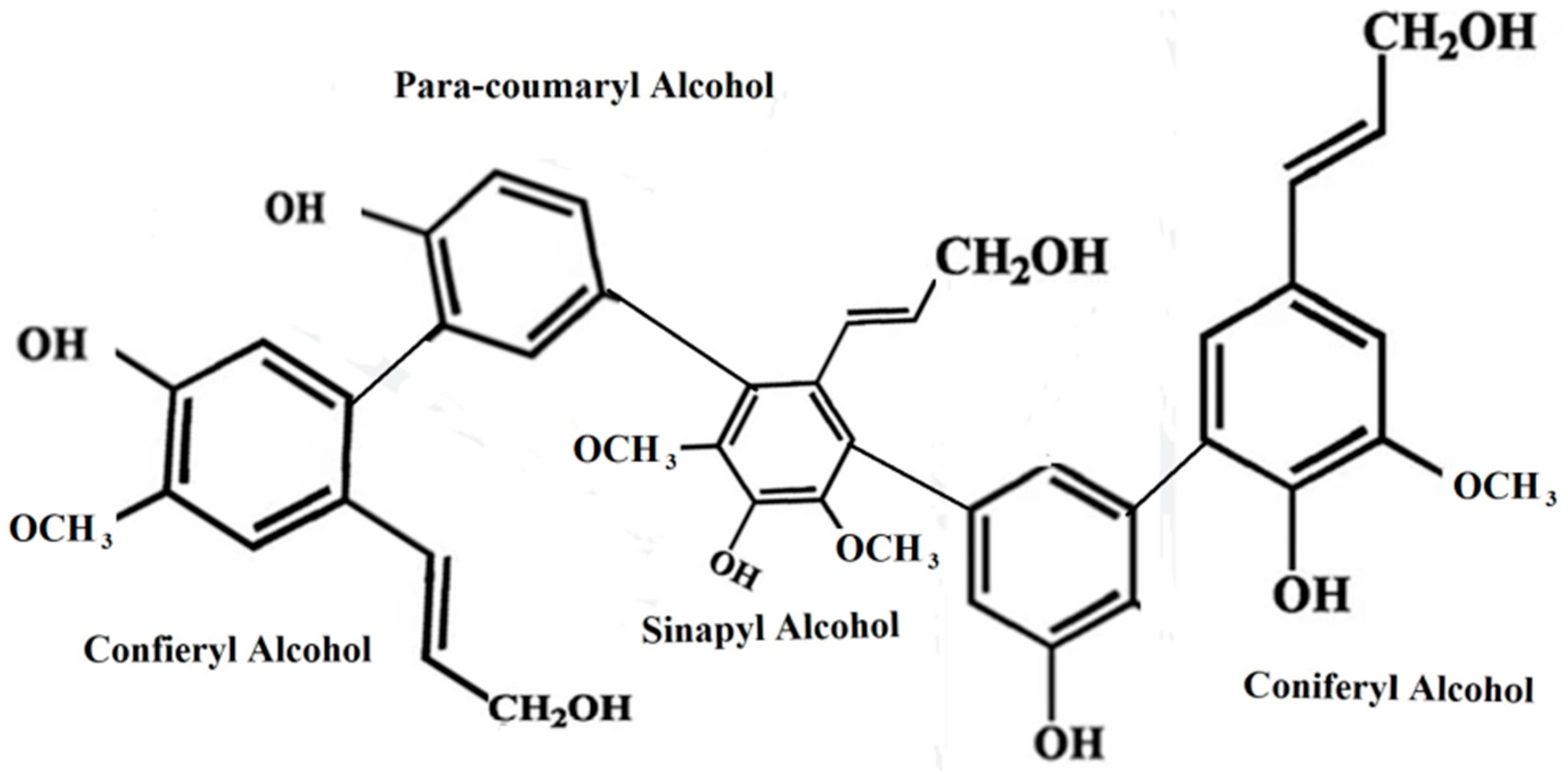{kind=link}
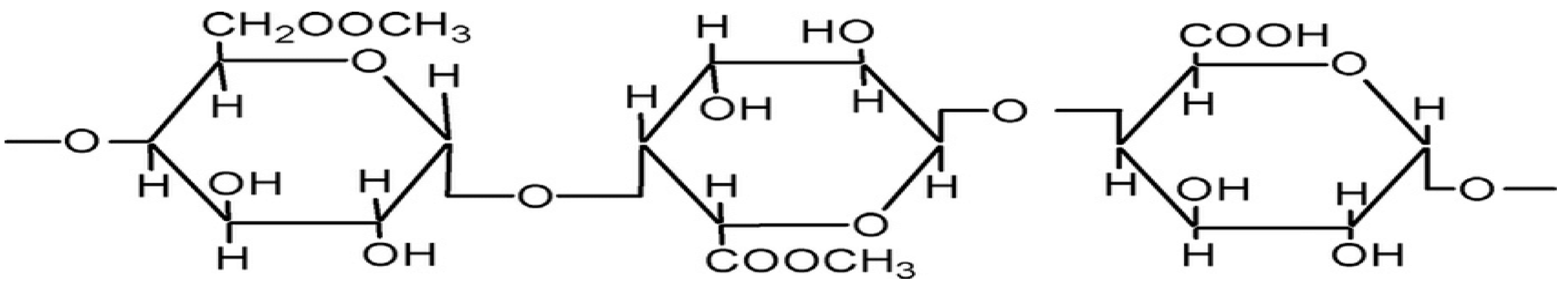{kind=link}
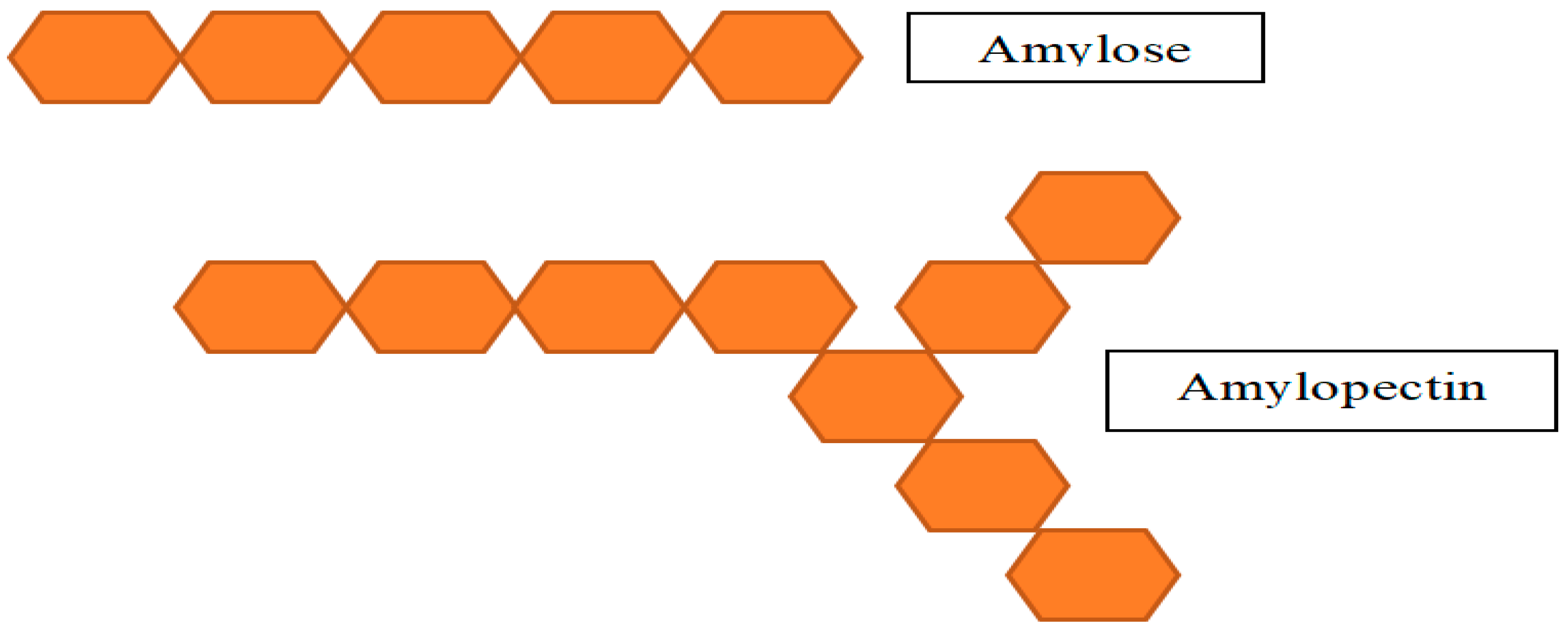{kind=link}
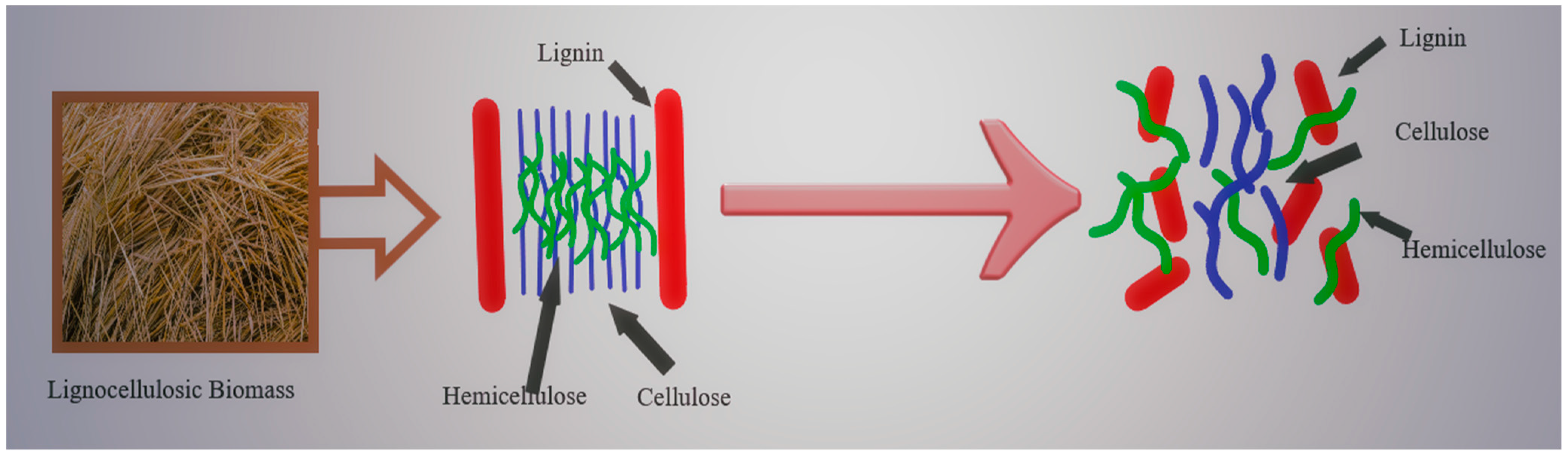{kind=link}
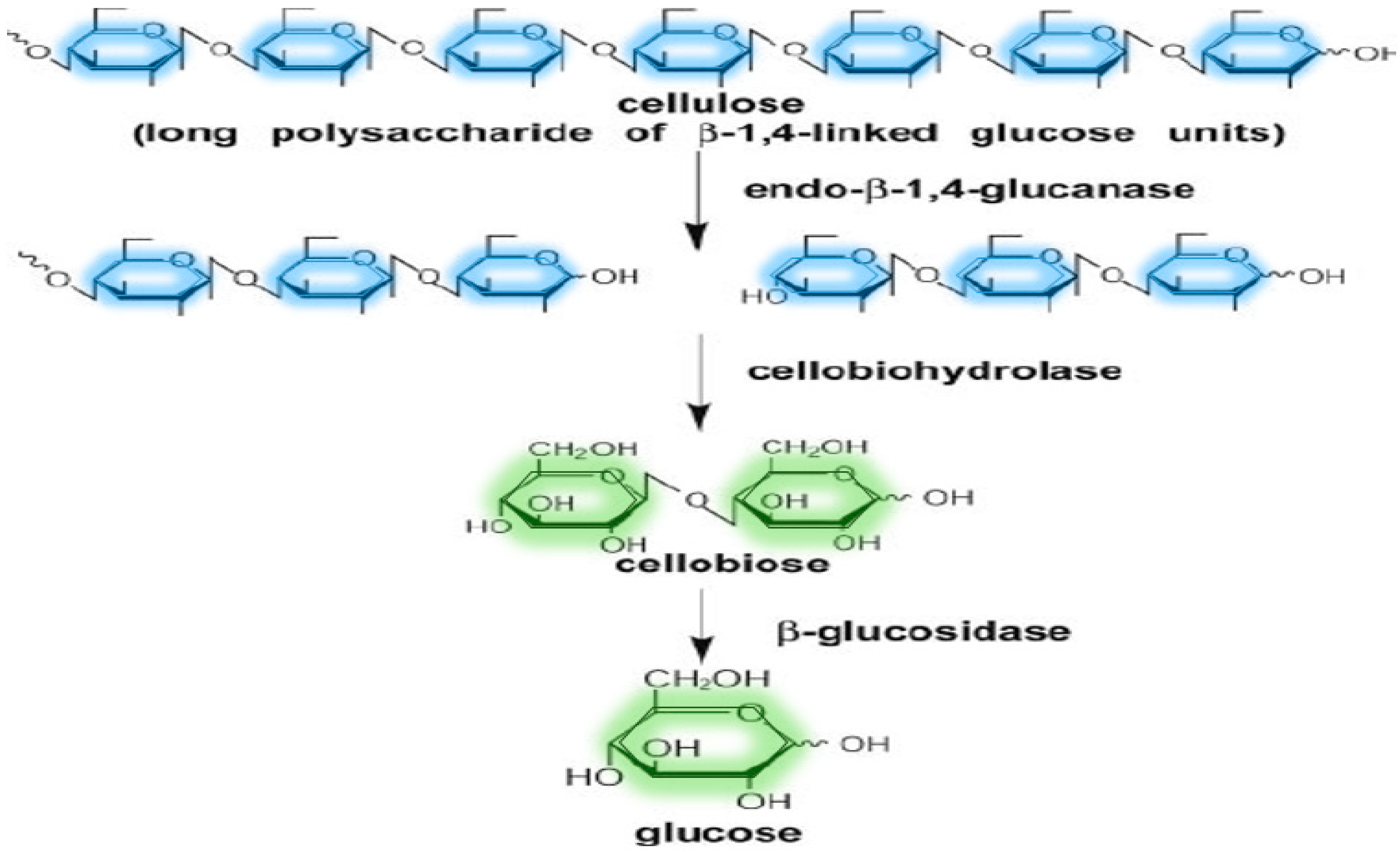{kind=link}
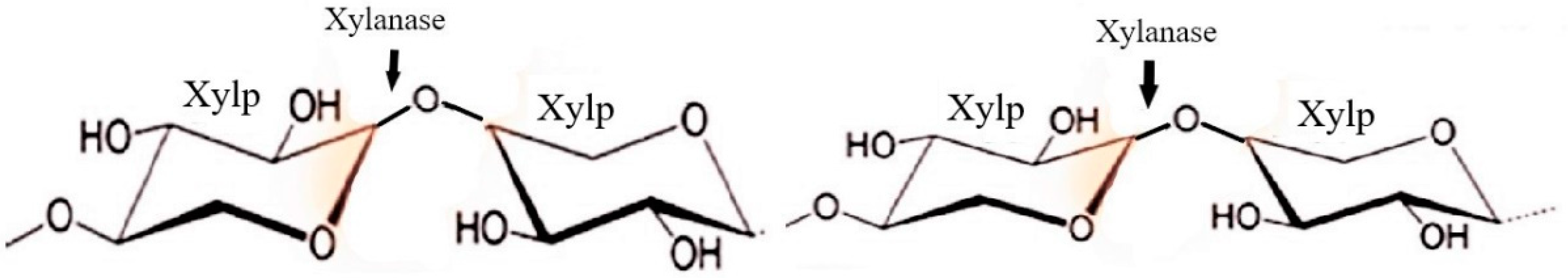{kind=link}
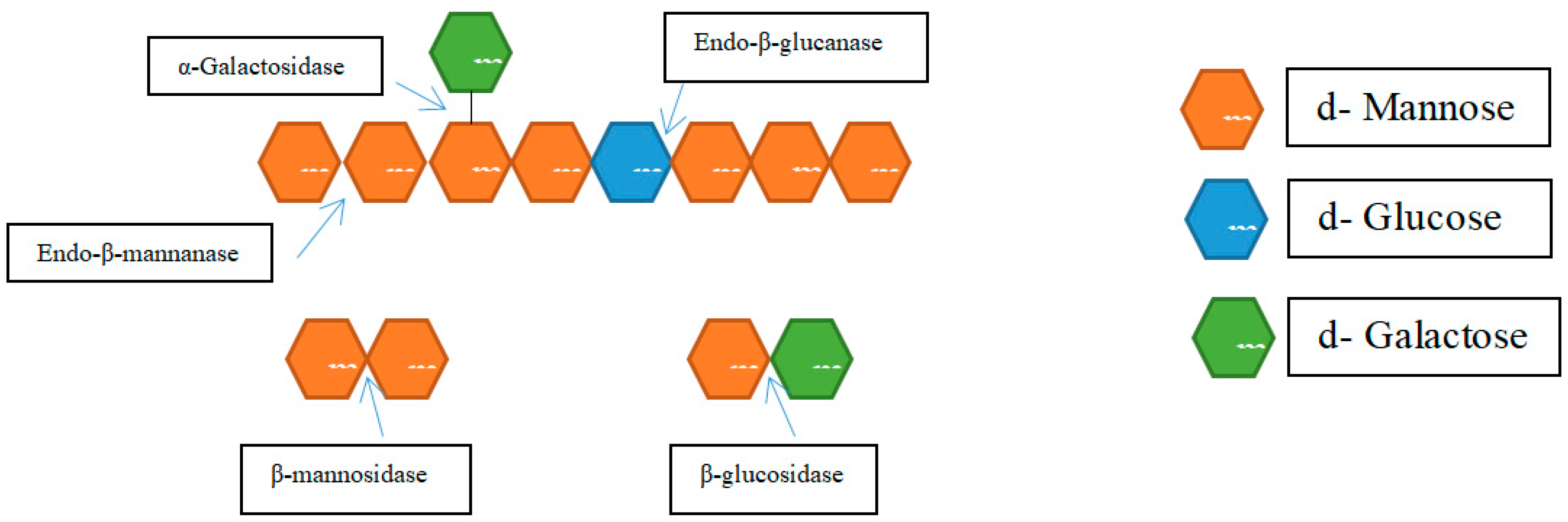{kind=link}
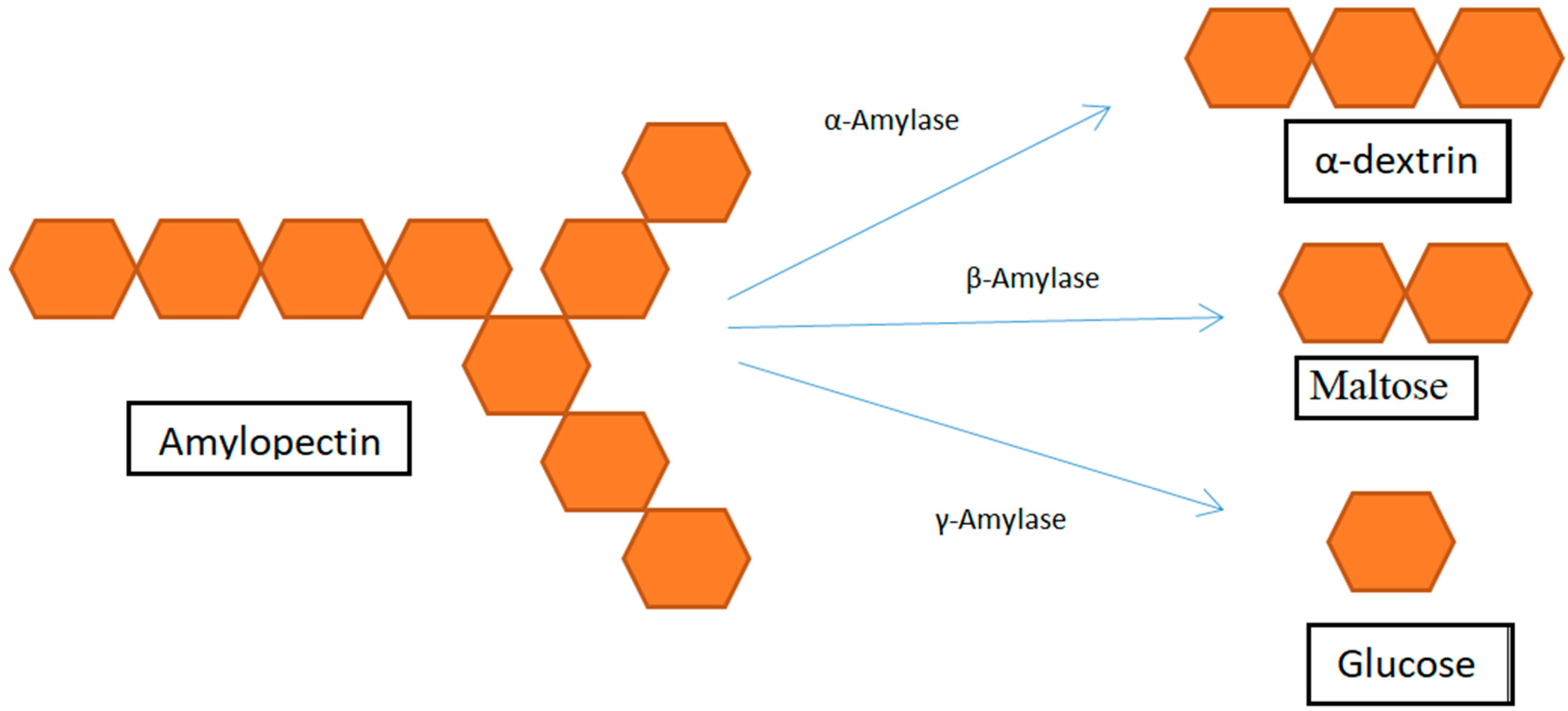{kind=link}
Table 1.
Lignocellulosic composition (%) of various agricultural wastes on a dry basis of the substrate.
Table 1.
Lignocellulosic composition (%) of various agricultural wastes on a dry basis of the substrate.
| Substrate | Cellulose (%) | Hemicellulose (%) | Lignin (%) | Reference |
|---|---|---|---|---|
| Rice straw | 32–47 | 19–27 | 5–24 | [21] |
| Rice husk | 34.40 | 29.30 | 19.20 | [22] |
| Wheat straw | 35–45 | 20–30 | 8–15 | [21] |
| Corn straw | 42.60 | 21.30 | 8.20 | [21] |
| Corn cobs | 45.00 | 35.00 | 15.00 | [21] |
| Corn stover | 38.00 | 26.00 | 19.00 | [23] |
| Wheat bran | 25.30 | 14.60 | 3.20 | [24] |
| Sugarcane bagasse | 42.00 | 25–36 | 19–20 | [16] |
| Sweet sorghum | 48–49 | 20–26 | 19–20 | [25] |
| Coconut fiber | 36–43 | 0.15–0.25 | 41–45 | [21] |
| Cocoa pods husk | 35 | 10 | 14 | [26] |
| Soft wood | 40–44 | 25–29 | 25–31 | [21] |
| Banana fiber | 60–65 | 6–8 | 5–10 | [21] |
| Switch grass | 36–38 | 27 | 17–19 | [27] |
| De-oiled rice bran | 9.80 | 20.60 | 3.90 | [28] |
| Barley straw | 31–45 | 27–38 | 14–19 | [21] |
Table 2.
Various methods, processes, and their impact on lignocellulosic biomass during the pretreatment.
Table 2.
Various methods, processes, and their impact on lignocellulosic biomass during the pretreatment.
| Nature of Pretreatment | Method | Process | Impact | Reference |
|---|---|---|---|---|
| Mechanical or physical | Milling | Roll, ball, hammer, disk, and colloid milling | Decreases polymerization and crystalline structure of cellulose, increases specific surface area | [47] |
| Extrusion | Mixing, heating, and shearing of biomass | Alterations in the physical and chemical structure. Defibrillation and fiber shortening | [48] | |
| Pulse electric field | A sudden burst of high voltage between 5.0–20.0 kV/cm for nano to milliseconds | Disruption of the cell wall and electroporation | [49] | |
| Microwave | Irradiation with 2450 MHz microwaves (170–200 °C) | Alterations in the ultra-structure of cellulose, partially removes hemicelluloses and lignin | [50,51] | |
| Chemical | Acidic | Treatment with dilute HCl, H3PO4, HNO3, H2SO4, acetic acid, citric acid, oxalic acid, maleic acid, fumaric acid, etc | Hydrolysis of hemicellulose | [9] |
| Alkaline | Treatment with dilute NaOH, KOH, Ca(OH)2, NH4OH | Efficient removal of lignin | [9] | |
| Physicochemical | Wet Oxidation | Treatment with oxidative agents such as peracetic acid, sodium chlorite, KMnO4, and H2O2 at high temperatures | Higher lignin and hemicelluloses solubilization | [52] |
| Organosolv | Treatment with organic or aqueous–organic solvent systems with or without added catalysts in the temperature range of 100–250 °C | Hydrolysis of lignin and hemicellulose | [53] | |
| Ammonia Fibre Expansion treatment (AFEX) | Treatment with anhydrous or liquid ammonia at a temperature ranging from 90 to 100 °C followed by a successive lowering of pressure | Lignin removal | [54] | |
| Steam Explosion | Exposure to saturated steam under high pressure followed by a sudden lowering of pressure | Lignin removal and hemicellulose solubilization | [55] | |
| Liquid hot water | Use of high temperature of 170°–230 °C and pressure more than 5 MPa | Removal of hemicelluloses | [56] | |
| Biological | Enzymes or microorganisms | Acton of lignin-degrading enzymes such as peroxidases and laccases | Lignin degradation | [46,57] |
Table 3.
Application of various hydrolytic enzymes in the degradation of polysaccharides in various biomass residues.
Table 3.
Application of various hydrolytic enzymes in the degradation of polysaccharides in various biomass residues.
| Hydrolytic Enzyme | Classification | Mode of Action | Common Lignocellulosic Biomass | References |
|---|---|---|---|---|
| Cellulases | Endoglucanase or Endo-β-1,4-glucanase | Random hydrolysis of the interior glycosidic bonds in cellulolytic biomass | Wheat straw, rice straw, corn cobs, wheat bran, oat bran, Arundo donax, Populus tremuloides, deoiled rice bran, kitchen waste | [9,73,75,106] |
| Cellobiohydrolase or Exo-β-1,4-glucanase | Hydrolysis of beta-D-glucosidic linkages by releasing mainly cellobiose | |||
| Cellobiase or β-glucosidase (BG) | Cleavage of cellobiose | |||
| Hemicellulases | Endo-β-1,4-xylanase | Release of xylose from xylan by Endohydrolysis of (1 → 4)-beta-D-xylosidic linkages | Wheat bran, kitchen waste, Banana peels, Peanut oil cake, Brewer’s spent grain | [9,80,81,84,106,107,108] |
| Exo-β-1,4-xylanase or β-1,4-xylan xylohydrolase | Release monomeric xylose from the non-reducing end of xylan | |||
| β-1,4-xylosidase or Xylobiase | hydrolyzes disaccharides such as xylobiose and the higher xylooligosaccharides | |||
| Endo-β-1,4-mannanase | Randomly cleaving the mannan’s β-1,4-linkage internal links | |||
| Exo-β-mannosidase | Releases mannose sugar moieties by cleaving β-1,4-linked mannosides from the non-reducing ends of mannan | |||
| β-glucosidase | Hydrolyzes the 1,4-β-D-glucopyranose found at the non-reducing ends of the oligosaccharides | |||
| α-galactosidase | breaks down the α-1,6-linked D-galactopyranosyl side chains of the oligosaccharides | |||
| Acetyl mannan esterase | The debranching enzyme releases acetyl groups. | |||
| Pectinases | Protopectinases | Liberate soluble form polymerized pectin | Wheat bran, mango peel, banana peel, kitchen waste, Orange peels, exhausted sugar beet cassettes | [84,85,109,110] |
| Pectin Methyl Esterases | Deesterify the methyl group of pectin, releasing pectic acid and methanol | |||
| Pectin Acetyl Esterases | Hydrolysis of the acetyl esters found in pectin | |||
| Polymethylgalacturonases | Breaks α-1,4-glycosidic linkages in pectin | |||
| Polygalacturonases | Cleaves the polygalacturonic acid’s α-1,4-glycosidic linkages | |||
| Pectate Lyases | Release α-4,5-D-galacturonate from the glycosidic bonds in polygalacturonic acid | |||
| Pectin Lyases | randomly break the esterified pectin and create unsaturated methyloligogalacturonates. | |||
| Amylase | Endoamylases or α-amylase | Cleaves the α-1,4-bonds present in the inner regions of amylose and amylopectin | Rice bran, wheat bran, black gram bran, Soybean husk, flour mill waste | [101,105,111] |
| Exoamylase or β-amylase | Release limit dextrins and β-maltose | |||
| ƴ-amylase or Amyloglucosidase or Glucoamylase | Debranching enzyme releases glucose |
Table 4.
Examples of the production of hydrolytic enzyme systems by solid-state fermentations using various substrates and breakthroughs.
Table 4.
Examples of the production of hydrolytic enzyme systems by solid-state fermentations using various substrates and breakthroughs.
| Substrate | Microorganism | Enzymes | Major Breakthrough | References |
|---|---|---|---|---|
| Wheat straw, rice straw, corn cobs, wheat bran, oat bran, Arundo donax, Populus tremuloides | Thermoascus aurantiacus | Cellulases | Thermostable cellulolytic components production | [121] |
| Wheat bran | Aspergillus awamori Nakazawa (MTCC 6652) | Glucoamylase | Optimization of extraction and purification of glucoamylase | [122] |
| Wheat bran | Aspergillus niger NS-2 | Cellulases xylanase, mannanase, pectinase, amylases | Co-production of multiple enzymes for Bioethanol Production | [123] |
| Deoiled rice bran | Aspergillus niger, Aspergillus oryzae, Trichoderma reesei | Cellulase, amylase | Co-production of the thermostable multi-enzyme system for ethanol production | [9,124] |
| Kitchen waste | Aspergillus niger CJ-5 | Cellulases, xylanase, mannanase, pectinase, amylases | Co-production of multiple enzymes for Bioethanol Production from kitchen waste residues | [73] |
| Brewer’s spent grain | Fusarium oxysporum SS-25 | Cellulases | Production of cellulases for the production of ethanol from brewer’s spent grain | [125] |
| wheat straw, paddy straw, sugarcane waste, maize straw | Bacillus licheniformis | α-amylase | Production of amylase from the mixture of agricultural residue waste | [126] |
| Rice bran, wheat bran, black gram bran | Achromobacter xylosoxidans | Amylase, cellulase, xylanase | Co-production of multiple enzymes from various agro waste | [127] |
| Peanut oil cake | Aspergillus oryzae | Cellulase, xylanase, amylase | Enhancement in various functional properties during fermentation in addition to enzyme activities | [107] |
| Brewer’s spent grain | Aspergillus niger CECT2088 | Cellulase, xylanase | Simultaneous production of lignocellulolytic enzymes | [108] |
| Orange peel, apple pomace, and rice fiber | Compost from Municipal Solid Waste as inoculum | Cellulases | Development of a framework for a zero-waste enzyme production process | [128] |
| Coffee husk and wood chips | Compost from MSW as inoculum | Cellulases | Enhanced cellulase production | [129] |
| Orange peels and exhausted sugar beet cassettes | Aspergillus awamori 2B.361 U2/1 | Cellulase, xylanase, pectinase | Enhanced sugar production | [109] |
| Sugarcane bagasse | Penicillium sp., Rhizomucor sp., Trichoderma sp. | Cellulases | Use of sugarcane bagasse as an inducer for cellulase | [130] |
| Grape pomace with wheat bran | Aspergillus niger 3T5B8 | Cellulase, xylanase | Production of a cocktail of hydrolytic enzymes using Grape pomace with wheat bran | [131] |
| Wheat bran, banana peel, orange peel, rice bran, pine apple peel | Bacillus subtilis D19 | Amylase | Enhanced amylase production on various agro-waste residues | [132] |
| Mango peels | Aspergillus tamarii | Pectinase | Enhanced polygalacturonase and pectin lyase | [110] |
| Wheat chaff | Trichoderma reesei QM 9414 | Cellulases and xylanase | Simultaneous production of cellulase and xylanase | [133] |
| Rice straw | Aspergillus niger P-19 | Cellulases, hemicellulases | Enhanced sugars and ethanol from rice straw | [119] |
| Rice straw | Penicillium spp. | Cellulase | Potent cellulase cocktail production for lignocellulosic degradation | [69] |
| Soybean husk and flour mill waste | Aspergillus oryzae | Amylase | Production and purification of alpha-amylase | [111] |
| Wheat bran | Bacillus sp. TC-DT13 | Xylanase | Optimized production of extracellular xylanase | [134] |
| Wheat bran | Trichoderma reesei, Neurospora crassa | Cellulases | Optimization and standardization of various factors for cellulase production | [24] |
| Banana peels | Aspergillus fumigatus | Pectinase and xylanase | Coproduction of pectinase and xylanase | [84] |
| Kitchen waste | Aspergillus niger S-30 | Cellulases, Hemicellulases, Pectinases, Amylases | 19 hydrolytic enzymes from a single substrate and organism | [120] |
Table 5.
Examples of the production of hydrolytic enzyme systems by liquid-state fermentations using various substrates and breakthroughs.
Table 5.
Examples of the production of hydrolytic enzyme systems by liquid-state fermentations using various substrates and breakthroughs.
| Substrate | Microorganism | Enzymes | Major Breakthrough | References |
|---|---|---|---|---|
| Rice bran | Aspergillus niger | Pectinase | Enhanced Polygalacturonase and Pectinmethylesterase activity | [142] |
| Solka-Floc cellulose | Penicillium brasilianum IBT 20888 | Cellulases, xylanase | Coinduction of cellulolytic and xylanolytic | [143] |
| Mandarin peels and tree leaves | Pleurotus dryinus | Cellulases, xylanase, laccase, manganese peroxidase | Enhanced activity of cellulases, xylanase, laccase, manganese peroxidase | [144] |
| Starch | Bacillus sp. | Amylase | Optimization of enhanced amylase production | [145] |
| Partially delignified cellulignin | Trichoderma harzianum IOC-4038 | Cellulases | Simultaneous saccharification and fermentation process development using partially delignified cellulignin | [146] |
| Sugarcane bagasse, corn stover | Acremonium sp. | Cellulases, xylanase | Enhanced reducing sugar conversion | [147] |
| Wheat bran | Aspergillus tamarii MTCC5152 | Amylase | Production of a cellulase-free and alkali-stable xylanase | [148] |
| Corn cob | Aspergillus fumigatus SD5A | xylanase | Use of eight fungal strains in xylanase production | [140] |
| Pineapple stem | Bacillus subtilis BKDS1 | Pectinase | Economical production of the enzyme, pectinase using pineapple stem extract (PSE) medium | [149] |
| Coffee waste | Penicillium humicola | Mannanase | Statistical experimental designs to enhance the β-mannanase production | [150] |
| Wheat bran and citrus peel waste | Bacillus pumilus | Xylanase and pectinase | Maximum production of xylanase and pectinase in a short submerged fermentation cycle | [151] |
| Banana peels | Bacillus subtilis TYg4-3 and Bacillus amyloliquefaciens SW106 | Pectinase | Optimization of bacterial pectinsae | [152] |
| Coffee residue powder, date seeds powder, prickly pear seeds | Bacillus subtilis US191 | Mannanase | Statistical experimental designs to enhance the bacterial β-mannanase production | [90] |
| Peanut shells | Bacillus paralichniformis | Cellulases | Utilization of peanut shells for cellulase production through Box-Behnken Design | [141] |
| Wheat chaff | Trichoderma reesei QM 9414 | Cellulases and xylanase | Simultaneous production of cellulase and xylanase | [133] |
| Wheat bran, rice husk | Aspergillus niger | Amylase | Production and purification of amylase using an aqueous two-phase system | [153] |
| Corn stover | Phanerochaete chrysosporium PC2 | Cellulases and hemicellulases | Revealed the importance of carbohydrate-binding module in the hydrolysis process of lignocellulose | [154] |
| Corn bran | Aspergillus niger | Xylanase | Use of UV- rays for enhanced xylanase | [155] |
| Wheat bran and citrus peel waste | Bacillus safensis M35, Bacillus altitudinis J208 | Xylanase and pectinase | Concentration values for wheat bran and citrus peel substrates are to be amended in one single production medium for enhanced xylanase and pectinase | [156] |
| Banana peels | Aspergillus fumigatus | Pectinase and xylanase | Coproduction of pectinase and xylanase | [84] |
| Kitchen waste | Aspergillus niger S-30 | Cellulases, Hemicellulases, Pectinases, Amylases | 19 hydrolytic enzymes from a single substrate and organism | [120] |
Table 6.
Salient features of various types of fermentation technology involved in second-generation bioethanol production.
Table 6.
Salient features of various types of fermentation technology involved in second-generation bioethanol production.
| Fermentation Technology | Steps Involved | Advantages | Disadvantages | Reference |
|---|---|---|---|---|
| Separate hydrolysis and fermentation (SHF) |
| The conditions can be optimized separately for each step | End product inhibitionRequire separate reactors for each step High energy and time consumption | [161] |
| Simultaneous saccharification and fermentation (SSF) |
|
| Differences in the optimum condition for hydrolytic enzymes and fermenting microorganisms | [162] |
| Simultaneous Saccharification and Co-Fermentation (SSCF) |
|
| Differences in the optimum condition for hydrolytic enzymes and fermenting microorganisms | [163] |
| Consolidated Bio-Processing (CBP) | Pretreatment, enzyme production, Saccharification, and Fermentation |
| Differences in the optimal conditions for enzymes or microorganisms involved in the process | [164] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Soni, S.K.; Sharma, A.; Soni, R. Microbial Enzyme Systems in the Production of Second Generation Bioethanol. Sustainability 2023, 15, 3590. https://doi.org/10.3390/su15043590
AMA Style
Soni SK, Sharma A, Soni R. Microbial Enzyme Systems in the Production of Second Generation Bioethanol. Sustainability. 2023; 15(4):3590. https://doi.org/10.3390/su15043590
Chicago/Turabian StyleSoni, Sanjeev Kumar, Apurav Sharma, and Raman Soni. 2023. "Microbial Enzyme Systems in the Production of Second Generation Bioethanol" Sustainability 15, no. 4: 3590. https://doi.org/10.3390/su15043590
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.