
Plant Carbon Sources for Denitrification Enhancement and Its Mechanism in Constructed Wetlands: A Review


 ,
,
Abstract
:1. Introduction
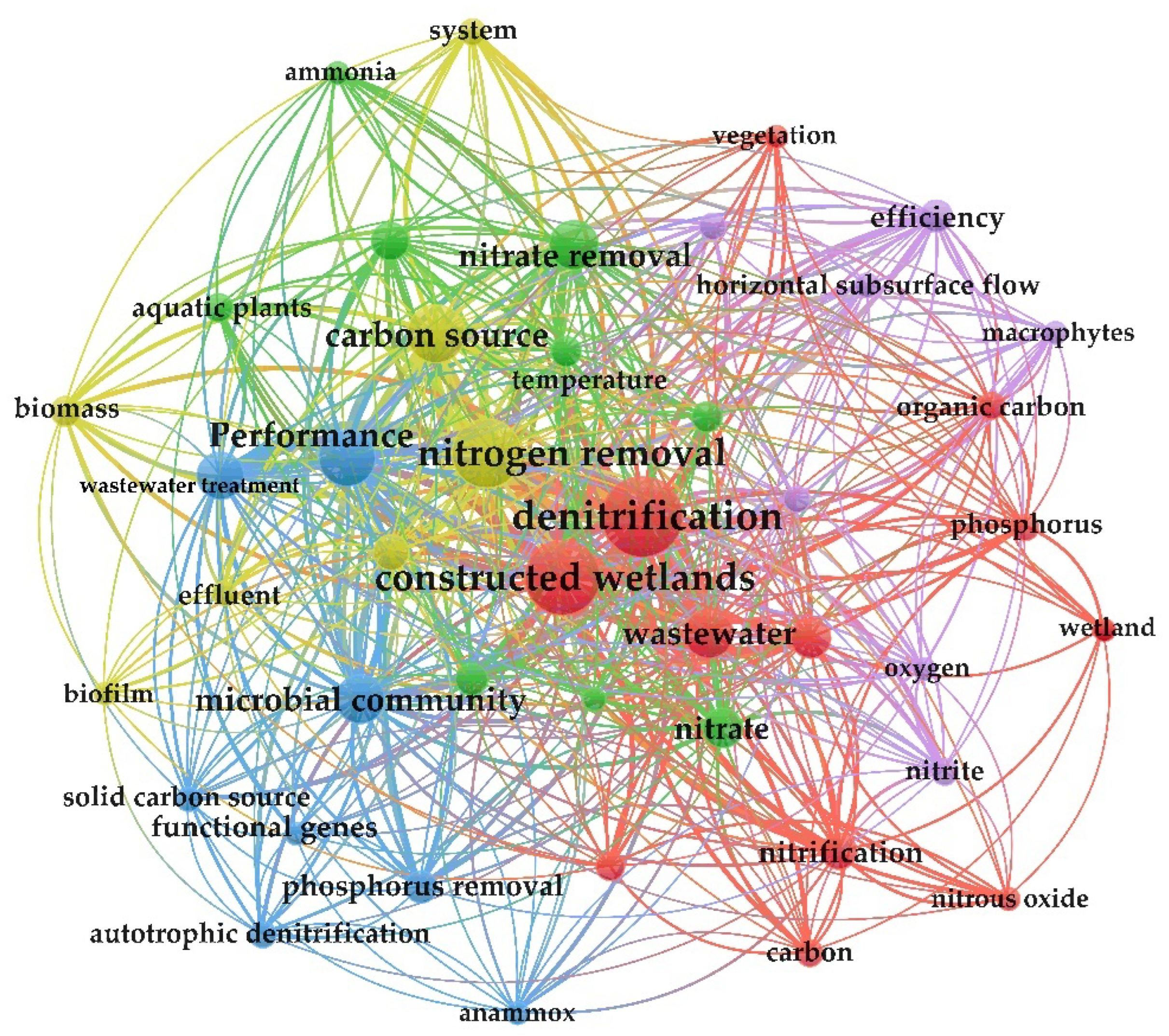
2. Bibliometric Assay Based on Keywords
3. PCS Characteristics
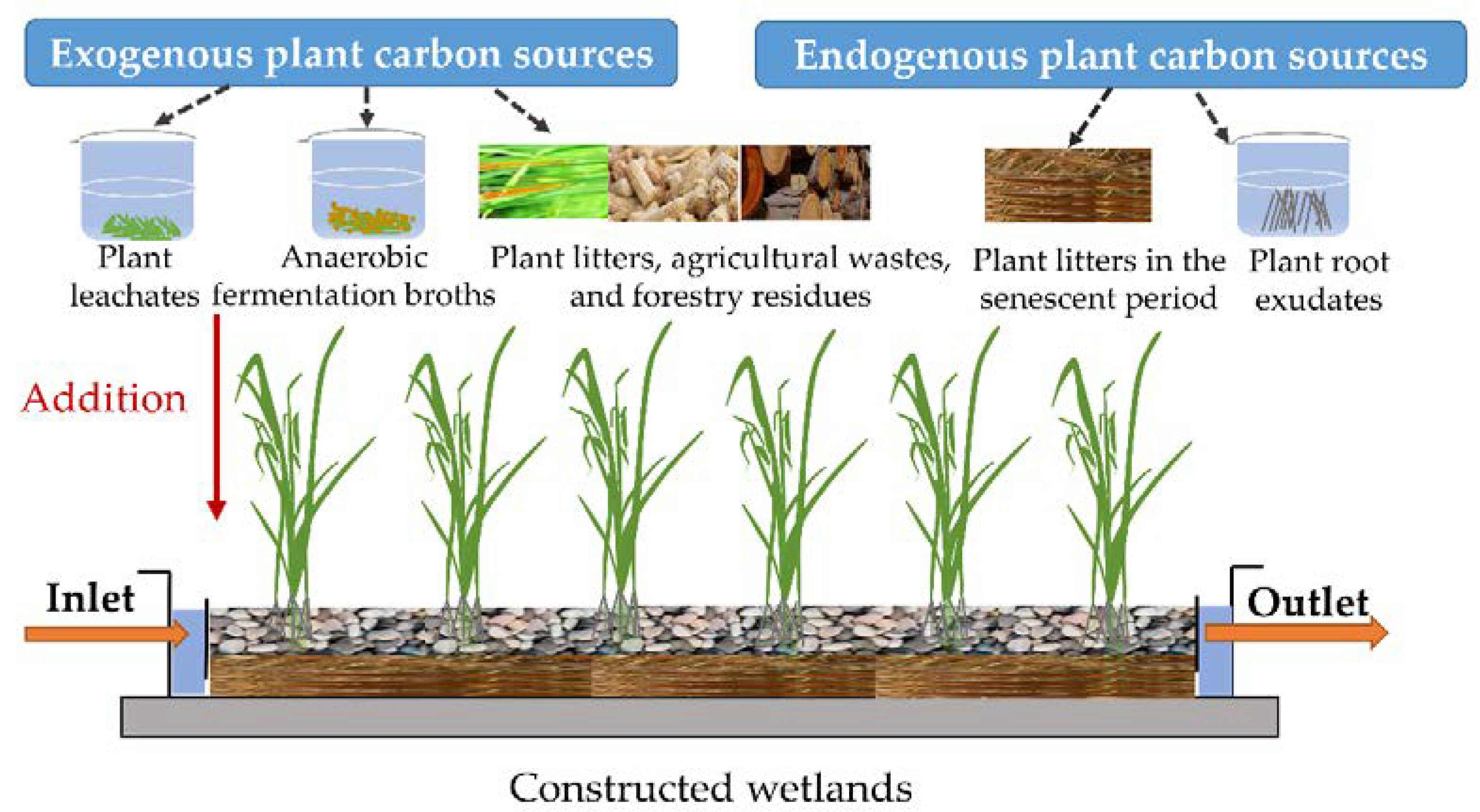
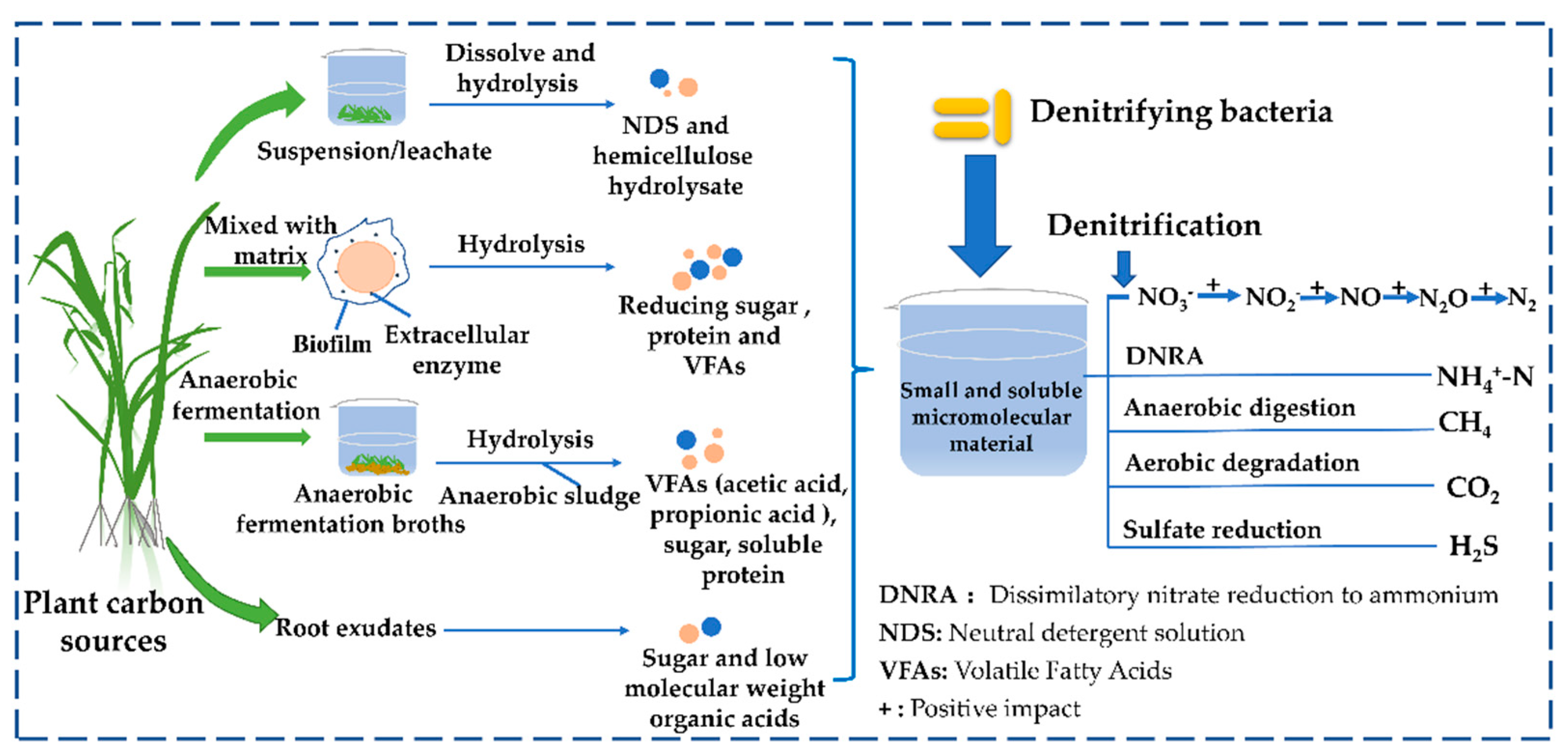
3.1. Classification of PCSs and Available Substances Released by PCSs in CWs
3.2. Comparison of Raw and Pretreated PCSs
4. Enhanced Denitrification by PCSs in CWs
4.1. Denitrification Performance Enhanced by PCSs in CWs
4.2. Secondary Pollutant Release
5. Influencing Factors of Denitrification Rate
5.1. Temperature
5.2. HRT
5.3. Influent Dissolved Oxygen (DO)
5.4. pH
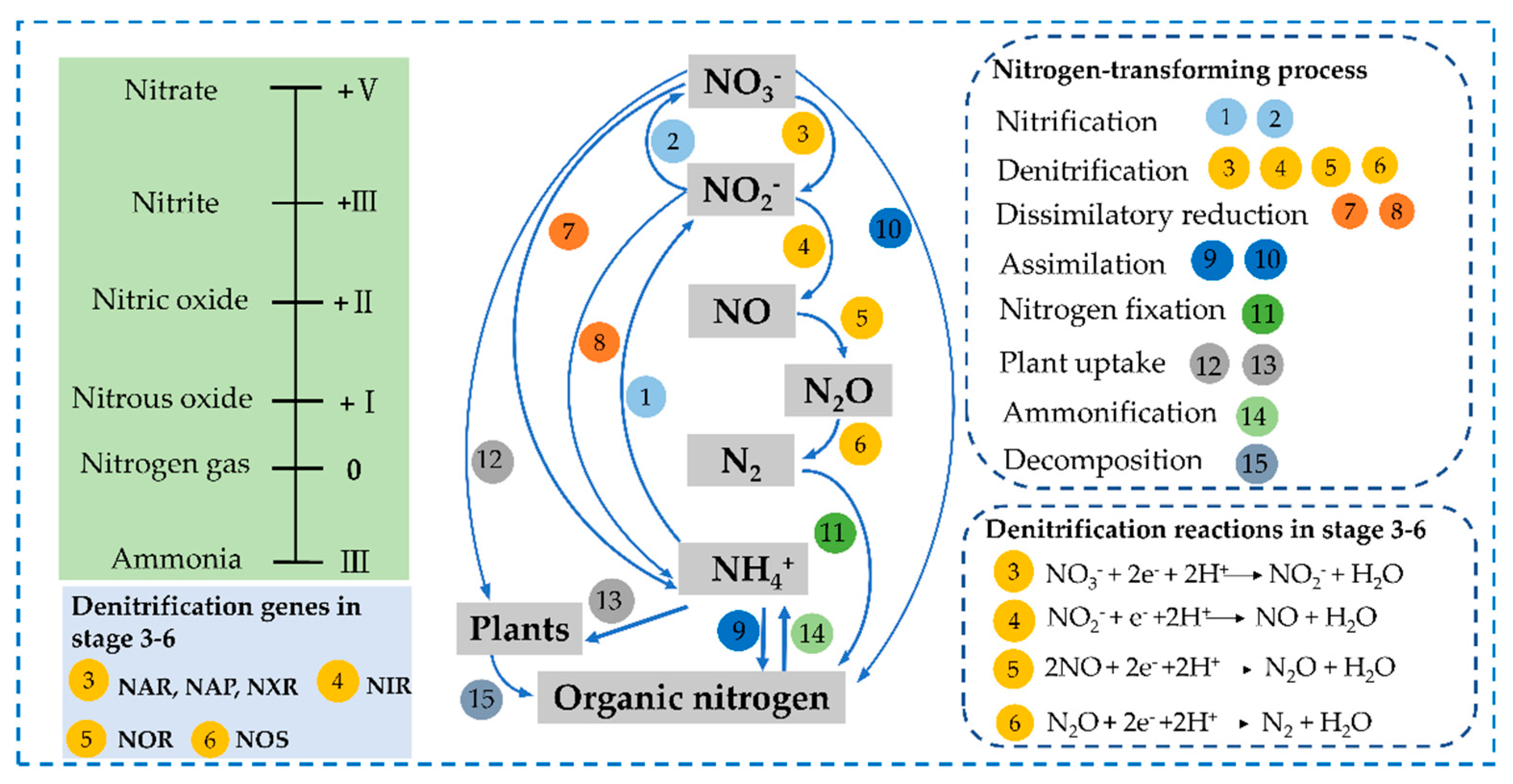
6. Denitrification Mechanism of PCSs
6.1. Physicochemical Mechanism
6.2. Molecular Biology Mechanism
6.2.1. Denitrification Functional Genes
6.2.2. Community Structure and Abundance of Microorganisms
7. Engineering Prospects
8. Existing Problems
9. Conclusions
- Acid or alkali combined with biological pretreatment (laccase, high-redox-potential peroxidase, and oxidase) of PCSs, aiming to make full use of the lignocellulose.
- Denitrification rates reflected in denitrification functional genomes and the influence of PCSs on denitrification functional genes and bacterial community structure in order to better understand the mechanism of denitrification enhancement by PCSs.
- Greenhouse gas (nitrous oxide and methane) emission reduction during the denitrification process.
- Synergistic removal of nitrogen and emerging pollutants such as pesticides and pharmaceutical and personal care products in CWs.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PCSs | Plant carbon sources |
| CWs | Constructed wetlands |
| COD | Chemical oxygen demand |
| COD/N | Chemical oxygen demand to total nitrogen |
| TN | Total nitrogen |
| VFAs | Volatile fatty acids |
| TOC | Total organic carbon |
| C/N | Carbon to nitrogen |
| HRT | Hydraulic retention time |
| SSF CWs | Subsurface-flow CWs |
| SF CWs | Surface-flow CWs |
| DOC | Dissolved organic carbon |
| DO | Dissolved oxygen |
| VF CWs | Vertical-flow CWs |
| NapA | Periplasmic nitrate reductase |
| NarG | Membrane-bound nitrate reductase |
| NirS | Cd1-containing nitrite reductase |
| NirK | Copper-containing nitrite reductase |
| QnorB/cnorB | Nitric oxide reductase |
| NosZ/nosR | Nitrous oxide reductase |
References
- Saeed, T.; Sun, G. A review on nitrogen and organics removal mechanisms in subsurface flow constructed wetlands: Dependency on environmental parameters, operating conditions and supporting media. J. Environ. Manag. 2012, 112, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ma, C.; Huang, X.; Liu, J.; Lu, L.; Peng, K.; Li, S. Research progress in solid carbon source-based denitrification technologies for different target water bodies. Sci. Total Environ. 2021, 782, 146669. [Google Scholar] [CrossRef] [PubMed]
- Maine, M.; Sune, N.; Hadad, H.; Sánchez, G.; Bonetto, C. Nutrient and metal removal in a constructed wetland for wastewater treatment from a metallurgic industry. Ecol. Eng. 2006, 26, 341–347. [Google Scholar] [CrossRef]
- Białowiec, A.; Davies, L.; Albuquerque, A.; Randerson, P.F. The influence of plants on nitrogen removal from landfill leachate in discontinuous batch shallow constructed wetland with recirculating subsurface horizontal flow. Ecol. Eng. 2012, 40, 44–52. [Google Scholar] [CrossRef]
- Lin, Y.F.; Jing, S.R.; Wang, T.W.; Lee, D.Y. Effects of macrophytes and external carbon sources on nitrate removal from groundwater in constructed wetlands. Environ. Pollut. 2002, 119, 413–420. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; Ngo, H.H.; Guo, W.; Hu, Z.; Liang, S.; Fan, J.; Liu, H. A review on the sustainability of constructed wetlands for wastewater treatment: Design and operation. Bioresour. Technol. 2015, 175, 594–601. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Knight, R.L.; Vymazal, J.; Brix, H.; Cooper, P.; Haberl, R. Constructed Wetlands for Pollution Control: Processes, Performance, Design and Operation, 6th ed.; IWA Publishing: London, UK, 2000; p. 156. [Google Scholar]
- Chen, X.; Zhu, H.; Yan, B.; Shutes, B.; Tian, L.; Wen, H. Optimal influent COD/N ratio for obtaining low GHG emissions and high pollutant removal efficiency in constructed wetlands. J. Clean. Prod. 2020, 267, 122003. [Google Scholar] [CrossRef]
- Sun, H.; Yang, Z.; Wei, C.; Wu, W. Nitrogen removal performance and functional genes distribution patterns in solid-phase denitrification sub-surface constructed wetland with micro aeration. Bioresour. Technol. 2018, 263, 223–231. [Google Scholar] [CrossRef]
- Hang, Q.; Wang, H.; Chu, Z.; Ye, B.; Li, C.; Hou, Z. Application of plant carbon source for denitrification by constructed wetland and bioreactor: Review of recent development. Environ. Sci. Pollut. Res. 2016, 23, 8260–8274. [Google Scholar] [CrossRef]
- van Oostrom, A.J. Nitrogen removal in constructed wetlands treating nitrified meat processing effluent. Water Sci. Technol. 1995, 32, 137–147. [Google Scholar] [CrossRef]
- Lu, S.; Hu, H.; Sun, Y.; Yang, J. Effect of carbon source on the denitrification in constructed wetlands. J. Environ. Sci. 2009, 21, 1036–1043. [Google Scholar] [CrossRef]
- Laber, J.; Perfler, R.; Haberl, R. Two strategies for advanced nitrogen elimination in vertical flow constructed wetlands. Water Sci. Technol. 1997, 35, 71–77. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L. Biological nitrate removal from water and wastewater by solid-phase denitrification process. Biotechnol. Adv. 2016, 34, 1103–1112. [Google Scholar] [CrossRef]
- Fan, C.; He, S.; Wu, S.; Huang, J. Improved denitrification in surface flow constructed wetland planted with calamus. J. Clean. Prod. 2021, 291, 125944. [Google Scholar] [CrossRef]
- Fu, G.P.; Huangshen, L.K.; Guo, Z.P.; Zhou, Q.H.; Wu, Z.B. Effect of plant-based carbon sources on denitrifying microorganisms in a vertical flow constructed wetland. Bioresour. Technol. 2017, 224, 214–221. [Google Scholar] [CrossRef]
- Wang, T.; Wang, H.; Chang, Y.; Chu, Z.; Zhao, Y.; Liu, R. Enhanced Nutrients Removal Using Reeds Straw as Carbon Source in a Laboratory Scale Constructed Wetland. Int. J. Environ. Res. Public. Health 2018, 15, 1081. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.Q.; Gao, L.; Gu, J.Y.; Zhou, W.L.; Fan, C.Z.; He, S.B.; Huang, J.C.; Zhang, X.; Cheng, Y.F.; Wu, Z.W.; et al. Enhancement of nitrogen removal via addition of cattail litter in surface flow constructed wetland. J. Clean. Prod. 2018, 204, 205–211. [Google Scholar] [CrossRef]
- Margalef-Marti, R.; Carrey, R.; Merchan, D.; Soler, A.; Causape, J.; Otero, N. Feasibility of using rural waste products to increase the denitrification efficiency in a surface flow constructed wetland. J. Hydrol. 2019, 578, 124035. [Google Scholar] [CrossRef]
- Jia, L.; Wang, R.; Feng, L.; Zhou, X.; Lv, J.; Wu, H. Intensified nitrogen removal in intermittently-aerated vertical flow constructed wetlands with agricultural biomass: Effect of influent C/N ratios. Chem. Eng. J. 2018, 345, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Chen, Y.; Zheng, N.; Yang, D.; Zhou, Q. Effects of plant biomass on nitrate removal and transformation of carbon sources in subsurface-flow constructed wetlands. Bioresour. Technol. 2010, 101, 7286–7292. [Google Scholar] [CrossRef]
- Tao, J.Q.; Zhang, Y.; Zhang, T.; You, Z.Y.; Shah, K.J.; Kim, H. Application of Reeds as Carbon Source for Enhancing Denitrification of Low C/N Micro-Polluted Water in Vertical-Flow Constructed Wetland. Appl. Sci. 2022, 12, 6756. [Google Scholar] [CrossRef]
- Ling, Y.; Yan, G.; Wang, H.; Dong, W.; Wang, H.; Chang, Y.; Chang, M.; Li, C. Release mechanism, secondary pollutants and denitrification performance comparison of six kinds of agricultural wastes as solid carbon sources for nitrate removal. Int. J. Environ. Res. Public. Health 2021, 18, 1232. [Google Scholar] [CrossRef]
- Si, Z.; Song, X.; Wang, Y.; Cao, X.; Zhao, Y.; Wang, B.; Chen, Y.; Arefe, A. Intensified heterotrophic denitrification in constructed wetlands using four solid carbon sources: Denitrification efficiency and bacterial community structure. Bioresour. Technol. 2018, 267, 416–425. [Google Scholar] [CrossRef]
- Song, S.Y.; Wang, P.H.; Liu, Y.X.; Zhao, D.H.; Leng, X.; An, S.Q. Effects of Oenanthe javanica on Nitrogen Removal in Free-Water Surface Constructed Wetlands under Low-Temperature Conditions. Int. J. Environ. Res. Public. Health 2019, 16, 1420. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, J.; Li, J. The removal of microplastics in the wastewater treatment process and their potential impact on anaerobic digestion due to pollutants association. Chemosphere 2020, 251, 126360. [Google Scholar] [CrossRef]
- Wu, H.L.; Wang, X.Z.; He, X.J.; Zhang, S.B.; Liang, R.B.; Shen, J. Effects of root exudates on denitrifier gene abundance, community structure and activity in a micro-polluted constructed wetland. Sci. Total Environ. 2017, 598, 697–703. [Google Scholar] [CrossRef]
- Ravindran, R.; Jaiswal, A.K. A comprehensive review on pre-treatment strategy for lignocellulosic food industry waste: Challenges and opportunities. Bioresour. Technol. 2016, 199, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Sorek, N.; Yeats, T.H.; Szemenyei, H.; Youngs, H.; Somerville, C.R. The Implications of Lignocellulosic Biomass Chemical Composition for the Production of Advanced Biofuels. Bioscience 2014, 64, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Han, L.; Dong, H. An insight to pretreatment, enzyme adsorption and enzymatic hydrolysis of lignocellulosic biomass: Experimental and modeling studies. Renew. Sustain. Energy Rev. 2021, 140, 110758. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Fleming-Singer, M.S.; Horne, A.J. Enhanced Nitrate Removal Efficiency in Wetland Microcosms Using an Episediment Layer for Denitrification. Environ. Sci. Technol. 2002, 36, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Chen, D.; Wu, F.; He, S.; Huang, J. Recycled utilization of Iris pseudacorus in constructed wetlands: Litters self-consumption and nitrogen removal improvement. Chemosphere 2021, 262, 127863. [Google Scholar] [CrossRef] [PubMed]
- Hume, N.P.; Fleming, M.S.; Horne, A.J. Denitrification potential and carbon quality of four aquatic plants in wetland microcosms. Soil Sci. Soc. Am. J. 2002, 66, 1706–1712. [Google Scholar] [CrossRef] [Green Version]
- Fu, G.P.; Yu, T.Y.; Huangshen, L.; Han, J.Y. The influence of complex fermentation broth on denitrification of saline sewage in constructed wetlands by heterotrophic nitrifying/aerobic denitrifying bacterial communities. Bioresour. Technol. 2018, 250, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Tian, Y.; Ding, Y.; Li, Z. The use of fermentation liquid of wastewater primary sedimentation sludge as supplemental carbon source for denitrification based on enhanced anaerobic fermentation. Bioresour. Technol. 2016, 219, 6–13. [Google Scholar] [CrossRef]
- Zhang, C.C.; Yin, Q.; Wen, Y.; Guo, W.R.; Liu, C.; Zhou, Q. Enhanced nitrate removal in self-supplying carbon source constructed wetlands treating secondary effluent: The roles of plants and plant fermentation broth. Ecol. Eng. 2016, 91, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, L.; Zhang, N.; Li, Z.; Zhang, G.; Xu, Y.; Shen, Q.; Zhang, R. Plant-microbe communication enhances auxin biosynthesis by a root-associated bacterium, Bacillus amyloliquefaciens SQR9. Mol. Plant-Microbe Interact. 2016, 29, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Haichar, F.e.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Li, P.; Zuo, J.; Wang, Y.; Zhao, J.; Tang, L.; Li, Z. Tertiary nitrogen removal for municipal wastewater using a solid-phase denitrifying biofilter with polycaprolactone as the carbon source and filtration medium. Water Res. 2016, 93, 74–83. [Google Scholar] [CrossRef]
- Yuan, T.Q.; You, T.T.; Wang, W.; Xu, F.; Sun, R.C. Synergistic benefits of ionic liquid and alkaline pretreatments of poplar wood. Part 2: Characterization of lignin and hemicelluloses. Bioresour. Technol. 2013, 136, 345–350. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, Y.Y.; Kim, T.H. A review on alkaline pretreatment technology for bioconversion of lignocellulosic biomass. Bioresour. Technol. 2016, 199, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Huang, S.; Wu, J.; Cheng, S.P.; Deng, Z.F. The use of microalgal biomass as a carbon source for nitrate removal in horizontal subsurface flow constructed wetlands. Ecol. Eng. 2019, 127, 263–267. [Google Scholar] [CrossRef]
- Ding, Y.; Song, X.; Wang, Y.; Yan, D. Effect of supplying a carbon extracting solution on denitrification in horizontal subsurface flow constructed wetlands. Korean J. Chem. Eng. 2013, 30, 379–384. [Google Scholar] [CrossRef]
- Barua, V.B.; Goud, V.V.; Kalamdhad, A.S. Microbial pretreatment of water hyacinth for enhanced hydrolysis followed by biogas production. Renew. Energy 2018, 126, 21–29. [Google Scholar] [CrossRef]
- Vasić, K.; Knez, Ž.; Leitgeb, M. Bioethanol Production by Enzymatic Hydrolysis from Different Lignocellulosic Sources. Molecules 2021, 26, 753. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R. Current Progress in Production of Building-Block Organic Acids by Consolidated Bioprocessing of Lignocellulose. Fermentation 2021, 7, 248. [Google Scholar] [CrossRef]
- Thakur, V.; Sharma, E.; Guleria, A.; Sangar, S.; Singh, K. Modification and management of lignocellulosic waste as an ecofriendly biosorbent for the application of heavy metal ions sorption. Mater. Today Proc. 2020, 32, 608–619. [Google Scholar] [CrossRef]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. Lignocellulolytic Enzymes in Biotechnological and Industrial Processes: A Review. Sustainability 2020, 12, 7282. [Google Scholar] [CrossRef]
- Zhao, D.H.; Zhang, M.; Liu, Z.; Sheng, J.; An, S.Q. Can cold-season macrophytes at the senescence stage improve nitrogen removal in integrated constructed wetland systems treating low carbon/nitrogen effluent? Bioresour. Technol. 2018, 265, 380–386. [Google Scholar] [CrossRef]
- Yuan, C.; Zhao, F.; Zhao, X.; Zhao, Y. Woodchips as sustained-release carbon source to enhance the nitrogen transformation of low C/N wastewater in a baffle subsurface flow constructed wetland. Chem. Eng. J. 2020, 392, 124840. [Google Scholar] [CrossRef]
- Ghafari, S.; Hasan, M.; Aroua, M.K. Bio-electrochemical removal of nitrate from water and wastewater—A review. Bioresour. Technol. 2008, 99, 3965–3974. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Z.; Dzakpasu, M.; Zhao, L.P.; Wang, Z.Z.; Zhang, D.X.; Qu, M.W.; Chen, R.; Wang, X.C.; Zheng, Y.C. Enhancement of partial denitrification-anammox pathways in constructed wetlands by plant-based external carbon sources. J. Clean. Prod. 2022, 370, 133581. [Google Scholar] [CrossRef]
- Yu, G.L.; Peng, H.Y.; Fu, Y.J.; Yan, X.J.; Du, C.Y.; Chen, H. Enhanced nitrogen removal of low C/N wastewater in constructed wetlands with co-immobilizing solid carbon source and denitrifying bacteria. Bioresour. Technol. 2019, 280, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Yu, T.; Ning, K.; Guo, Z.; Wong, M.-H. Effects of nitrogen removal microbes and partial nitrification-denitrification in the integrated vertical-flow constructed wetland. Ecol. Eng. 2016, 95, 83–89. [Google Scholar] [CrossRef]
- He, Y.; Wang, Y.; Song, X. High-effective denitrification of low C/N wastewater by combined constructed wetland and biofilm-electrode reactor (CW-BER). Bioresour. Technol. 2016, 203, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yi, N.K.; Wang, S.; Lu, L.J.; Huang, X.F. Impact of plant species on spatial distribution of metabolic potential and functional diversity of microbial communities in a constructed wetland treating aquaculture wastewater. Ecol. Eng. 2016, 94, 564–573. [Google Scholar] [CrossRef]
- Sudarno, U.; Winter, J.; Gallert, C. Effect of varying salinity, temperature, ammonia and nitrous acid concentrations on nitrification of saline wastewater in fixed-bed reactors. Bioresour. Technol. 2011, 102, 5665–5673. [Google Scholar] [CrossRef]
- Wu, S.Q.; He, S.B.; Huang, J.C.; Gu, J.Y.; Zhou, W.L.; Gao, L. Decomposition of Emergent Aquatic Plant (Cattail) Litter Under Different Conditions and the Influence on Water Quality. Water Air Soil Pollut. 2017, 228, 70. [Google Scholar] [CrossRef]
- Gao, D.W.; Hu, Q. Bio-contact oxidation and greenhouse-structured wetland system for rural sewage recycling in cold regions: A full-scale study. Ecol. Eng. 2012, 49, 249–253. [Google Scholar] [CrossRef]
- Yan, Y.; Xu, J. Improving Winter Performance of Constructed Wetlands for Wastewater Treatment in Northern China: A Review. Wetlands 2014, 34, 243–253. [Google Scholar] [CrossRef]
- Ji, M.; Hu, Z.; Hou, C.; Liu, H.; Ngo, H.H.; Guo, W.; Lu, S.; Zhang, J. New insights for enhancing the performance of constructed wetlands at low temperatures. Bioresour. Technol. 2020, 301, 122722. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.H.; Chen, C.; Lu, Q.Q.; Zhang, M.; Zhang, H.; An, S.Q. Combined use of cold-season and warm-season macrophytes in floating constructed wetlands to increase nitrogen removal in the early cold season. J. Clean. Prod. 2020, 254, 120054. [Google Scholar] [CrossRef]
- Korom, S.F. Natural denitrification in the saturated zone: A review. Water Resour. Res. 1992, 28, 1657–1668. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; He, F.; Ma, L.; Zhang, Y.; Wu, Z. Microbial nitrogen removal pathways in integrated vertical-flow constructed wetland systems. Bioresour. Technol. 2016, 207, 339–345. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, X.; Fan, Y.; Qiu, T.; Gao, M.; Wang, X. Heterotrophic nitrification and aerobic denitrification by Diaphorobacter polyhydroxybutyrativorans SL-205 using poly (3-hydroxybutyrate-co-3-hydroxyvalerate) as the sole carbon source. Bioresour. Technol. 2017, 241, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wen, Y.; Cheng, J.; Xue, C.; Yang, D.; Zhou, Q. Effects of dissolved oxygen on extracellular enzymes activities and transformation of carbon sources from plant biomass: Implications for denitrification in constructed wetlands. Bioresour. Technol. 2011, 102, 2433–2440. [Google Scholar] [CrossRef]
- Faulwetter, J.L.; Gagnon, V.; Sundberg, C.; Chazarenc, F.; Burr, M.D.; Brisson, J.; Camper, A.K.; Stein, O.R. Microbial processes influencing performance of treatment wetlands: A review. Ecol. Eng. 2009, 35, 987–1004. [Google Scholar] [CrossRef]
- Liu, F.; Sun, L.; Wan, J.; Shen, L.; Yu, Y.; Hu, L.; Zhou, Y. Performance of different macrophytes in the decontamination of and electricity generation from swine wastewater via an integrated constructed wetland-microbial fuel cell process. J. Environ. Sci. 2020, 89, 252–263. [Google Scholar] [CrossRef]
- Thomas, K.L.; Lloyd, D.; Boddy, L. Effects of oxygen, pH and nitrate concentration on denitrification by Pseudomonas species. FEMS Microbiol. Lett. 1994, 118, 181–186. [Google Scholar] [CrossRef]
- Silverstein, J.; Hund, R.; Figueroa, L.; Cook, N.; Cutter, K.; Deters, J. Biological denitrification of high nitrate industrial wastewater. In Proceedings of the 48th Purdue Waste Conference Proceedings, Purdue University, West Lafayette, IN, USA, 10–12 May 1993; pp. 723–732. [Google Scholar]
- Glass, C.; Silverstein, J. Denitrification kinetics of high nitrate concentration water: pH effect on inhibition and nitrite accumulation. Water Res. 1998, 32, 831–839. [Google Scholar] [CrossRef]
- Saleh-Lakha, S.; Shannon, K.E.; Henderson, S.L.; Goyer, C.; Trevors, J.T.; Zebarth, B.J.; Burton, D.L. Effect of pH and Temperature on Denitrification Gene Expression and Activity in Pseudomonas mandelii. Appl. Environ. Microbiol. 2009, 75, 3903–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, D.C.; DeLaune, R.D. Fungal and bacterial mediated denitrification in wetlands: Influence of sediment redox condition. Water Res. 2010, 44, 2441–2450. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.; Yu, X.X.; Zhang, Y.G.; Peng, Z.X.; Yu, L.J.; Cheng, L.L.; Li, T.M. Comparison of agricultural wastes and synthetic macromolecules as solid carbon source in treating low carbon nitrogen wastewater. Sci. Total Environ. 2020, 739, 139885. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wen, Y.; Zhou, Q.; Vymazal, J. Effects of plant biomass on nitrogen transformation in subsurface-batch constructed wetlands: A stable isotope and mass balance assessment. Water Res. 2014, 63, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.X.; Zhang, H.; Zuo, J.; Wang, P.H.; Zhao, D.H.; An, S.Q. Decreasing but still significant facilitation effect of cold-season macrophytes on wetlands purification function during cold winter. Sci. Rep. 2016, 6, 27011. [Google Scholar] [CrossRef]
- Jin, P.; Chen, Y.; Yao, R.; Zheng, Z.; Du, Q. New insight into the nitrogen metabolism of simultaneous heterotrophic nitrification-aerobic denitrification bacterium in mRNA expression. J. Hazard. Mater. 2019, 371, 295–303. [Google Scholar] [CrossRef]
- Luo, Z.; Li, S.; Zhu, X.; Ji, G. Carbon source effects on nitrogen transformation processes and the quantitative molecular mechanism in long-term flooded constructed wetlands. Ecol. Eng. 2018, 123, 19–29. [Google Scholar] [CrossRef]
- Zhi, W.; Yuan, L.; Ji, G.; He, C. Enhanced Long-Term Nitrogen Removal and Its Quantitative Molecular Mechanism in Tidal Flow Constructed Wetlands. Environ. Sci. Technol. 2015, 49, 4575–4583. [Google Scholar] [CrossRef]
- Abell, G.C.; Revill, A.T.; Smith, C.; Bissett, A.P.; Volkman, J.K.; Robert, S.S. Archaeal ammonia oxidizers and nirS-type denitrifiers dominate sediment nitrifying and denitrifying populations in a subtropical macrotidal estuary. ISME J. 2010, 4, 286–300. [Google Scholar] [CrossRef]
- Chen, Y.; Wen, Y.; Zhou, Q.; Vymazal, J. Effects of plant biomass on denitrifying genes in subsurface-flow constructed wetlands. Bioresour. Technol. 2014, 157, 341–345. [Google Scholar] [CrossRef]
- Bawiec, A. Efficiency of nitrogen and phosphorus compounds removal in hydroponic wastewater treatment plant. Environ. Technol. 2019, 40, 2062–2072. [Google Scholar] [CrossRef] [PubMed]
- Hallin, S.; Hellman, M.; Choudhury, M.I.; Ecke, F. Relative importance of plant uptake and plant associated denitrification for removal of nitrogen from mine drainage in sub-arctic wetlands. Water Res. 2015, 85, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L. Denitrifying genes in bacterial and Archaeal genomes. BBA-Gene Struct. Expr. 2002, 1577, 355–376. [Google Scholar] [CrossRef]
- Jones, C.M.; Stres, B.; Rosenquist, M.; Hallin, S. Phylogenetic analysis of nitrite, nitric oxide, and nitrous oxide respiratory enzymes reveal a complex evolutionary history for denitrification. Mol. Biol. Evol. 2008, 25, 1955–1966. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wen, Y.; Tang, Z.R.; Huang, J.G.; Zhou, Q.; Vymazal, J. Effects of plant biomass on bacterial community structure in constructed wetlands used for tertiary wastewater treatment. Ecol. Eng. 2015, 84, 38–45. [Google Scholar] [CrossRef]
- Ji, G.; He, C.; Tan, Y. The spatial distribution of nitrogen removal functional genes in multimedia biofilters for sewage treatment. Ecol. Eng. 2013, 55, 35–42. [Google Scholar] [CrossRef]
- Zhi, W.; Ji, G.D. Quantitative response relationships between nitrogen transformation rates and nitrogen functional genes in a tidal flow constructed wetland under C/N ratio constraints. Water Res. 2014, 64, 32–41. [Google Scholar] [CrossRef]
- Huang, Z.J.; Kong, F.L.; Li, Y.; Xu, G.M.; Yuan, R.Y.; Wang, S. Advanced treatment of effluent from municipal wastewater treatment plant by strengthened ecological floating bed. Bioresour. Technol. 2020, 309, 123358. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, H.; Zuo, J.; Zhao, D.; Zou, X.; Zhu, Z.; Jeelani, N.; Leng, X.; An, S. A Hardy Plant Facilitates Nitrogen Removal via Microbial Communities in Subsurface Flow Constructed Wetlands in Winter. Sci. Rep. 2016, 6, 33600. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L.; Hallin, S.; Schloter, M. Ecology of Denitrifying Prokaryotes in Agricultural Soil. Adv. Agron. 2007, 96, 249–305. [Google Scholar] [CrossRef]
- Shen, Z.; Zhou, Y.; Liu, J.; Xiao, Y.; Cao, R.; Wu, F. Enhanced removal of nitrate using starch/PCL blends as solid carbon source in a constructed wetland. Bioresour. Technol. 2015, 175, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Hou, R.; Yang, P.; Qian, S.; Feng, Z.; Chen, Z.; Wang, F.; Yuan, R.; Chen, H.; Zhou, B. Application of external carbon source in heterotrophic denitrification of domestic sewage: A review. Sci. Total Environ. 2022, 817, 153061. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, J.; Bai, S.; Ma, F.; Wang, L. Microbial population dynamics in response to bioaugmentation in a constructed wetland system under 10°C. Bioresour. Technol. 2016, 205, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Meng, P.; Pei, H.; Hu, W.; Shao, Y.; Li, Z. How to increase microbial degradation in constructed wetlands: Influencing factors and improvement measures. Bioresour. Technol. 2014, 157, 316–326. [Google Scholar] [CrossRef]
- Chen, Y.; Wen, Y.; Zhou, J.; Zhou, Q.; Vymazal, J.; Kuschk, P. Transformation of Chloroform in Model Treatment Wetlands: From Mass Balance to Microbial Analysis. Environ. Sci. Technol. 2015, 49, 6198–6205. [Google Scholar] [CrossRef]
- Hume, N.P.; Fleming, M.S.; Horne, A.J. Plant carbohydrate limitation on nitrate reduction in wetland microcosms. Water Res. 2002, 36, 577–584. [Google Scholar] [CrossRef]
- Thomas, S.C.; Martin, A.R. Carbon Content of Tree Tissues: A Synthesis. Forests 2012, 3, 332–352. [Google Scholar] [CrossRef] [Green Version]
- National Bureau of Statistics. Environmental Statistics: Wetland Conditions in Different Regions from June to September. Available online: http://www.stats.gov.cn/ztjc/ztsj/hjtjzl/2013/201412/t20141219_655911.html (accessed on 4 March 2014).
- Vymazal, J. Constructed Wetlands for Wastewater Treatment: Five Decades of Experience. Environ. Sci. Technol. 2011, 45, 61–69. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, H.; Hang, Q.; Deng, Y.; Liu, K.; Li, C.; Zheng, S. Comparison of the MBBR denitrification carriers for advanced nitrogen removal of wastewater treatment plant effluent. Environ. Sci. Pollut. Res. 2015, 22, 13970–13979. [Google Scholar] [CrossRef]
- Aravani, V.P.; Sun, H.; Yang, Z.; Liu, G.; Wang, W.; Anagnostopoulos, G.; Syriopoulos, G.; Charisiou, N.D.; Goula, M.A.; Kornaros, M.; et al. Agricultural and livestock sector’s residues in Greece & China: Comparative qualitative and quantitative characterization for assessing their potential for biogas production. Renew. Sustain. Energy Rev. 2022, 154, 111821. [Google Scholar] [CrossRef]
- Yang, X.L.; Jiang, Q.; Song, H.L.; Gu, T.T.; Xia, M.Q. Selection and application of agricultural wastes as solid carbon sources and biofilm carriers in MBR. J. Hazard. Mater. 2015, 283, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, F.; Luo, P.; Chen, X.; Chen, J.; Huang, Z.; Peng, J.; Xiao, R.; Wu, J. Stimulation of optimized influent C:N ratios on nitrogen removal in surface flow constructed wetlands: Performance and microbial mechanisms. Sci. Total Environ. 2019, 694, 133575. [Google Scholar] [CrossRef] [PubMed]
- Mosier, A.; Halvorson, A.; Peterson, G.; Robertson, G.; Sherrod, L. Measurement of net global warming potential in three agroecosystems. Nutr. Cycl. Agroecosyst. 2005, 72, 67–76. [Google Scholar] [CrossRef]
- Elgood, Z.; Robertson, W.D.; Schiff, S.L.; Elgood, R. Nitrate removal and greenhouse gas production in a stream-bed denitrifying bioreactor. Ecol. Eng. 2010, 36, 1575–1580. [Google Scholar] [CrossRef]
- Li, M.; Wu, H.; Zhang, J.; Ngo, H.H.; Guo, W.; Kong, Q. Nitrogen removal and nitrous oxide emission in surface flow constructed wetlands for treating sewage treatment plant effluent: Effect of C/N ratios. Bioresour. Technol. 2017, 240, 157–164. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, J.; Jia, W.; Xie, H.; Gu, R.R.; Li, C.; Gao, B. Impact of COD/N ratio on nitrous oxide emission from microcosm wetlands and their performance in removing nitrogen from wastewater. Bioresour. Technol. 2009, 100, 2910–2917. [Google Scholar] [CrossRef]
- Liu, X.; Guo, X.; Liu, Y.; Lu, S.; Xi, B.; Zhang, J.; Wang, Z.; Bi, B. A review on removing antibiotics and antibiotic resistance genes from wastewater by constructed wetlands: Performance and microbial response. Environ. Pollut. 2019, 254, 112996. [Google Scholar] [CrossRef]
- Ramprasad, C.; Philip, L. Surfactants and personal care products removal in pilot scale horizontal and vertical flow constructed wetlands while treating greywater. Chem. Eng. J. 2016, 284, 458–468. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCSs | Pretreatment Method | Raw PCSs | Pretreated PCSs | Ref | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cellulose | Hemicellulose | Lignin | C/N | Cellulose | Hemicellulose | Lignin | C/N | |||
| Mixed Cattail, canna, and rice straw | Water extracted | 34.08 a | 26.19 a | 15.80 a | NA | 37.20 a | 29.38 a | 13.12 a | NA | [44] |
| 2% H2 SO4 extracted | 38.87 a | 31.92 a | 11.02 a | NA | ||||||
| 5% H2SO4 extracted | 41.81 a | 33.95 a | 9.53 a | NA | ||||||
| Cattail litter | 2% NaOH extracted | 29.10 a | 11.10 a | 12.40 a | 60 | 38.01 a | 13.43 a | 10.04 a | 669 | [21] |
| Cattail litter (Typha latifolia) | Anaerobic fermentation | 28.60 a | 13.70 a | 9.30 a | NA | Mainly composed of VFAs | [37] | |||
| PCSs | Influent | Plant Dosage (g L−1) | Influent Nitrate (mg L−1) | Influent COD/N | HRT (d) | Temperature (°C) | Nitrate Removal Efficiency (%) | Denitrification Rate (g N m−3 d−1) | Note | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| Iris pseudacorus | SIS | 0.50–1.00 b | 10 b | 0 b | 2 b | 25–29 b | 45.00–95.00 b | 2.25–4.75 b | PL | [33] |
| Control | SIS | 0 b 2.80 b | 50.00 b | 0.37 b | 5 b | greenhouse | 29.90–100.00 b 56.30–100 b | 0.98–5.97 1.81–29.13 | [21] | |
| Cattail | ||||||||||
| Microalgal | SIS | 0 b | 10 a | 4 b | 6 b | Summer | 51.20 a | 0.28 b | PL | [43] |
| 0.10 b | 65.30–79.10 a | 0.36–0.43 b | ||||||||
| Arundo donax | SIS | influent COD/N = 9 | 10 a | 1.5 b | 0.83 b | 27 a | 70.40 b | 10.87 b | AFbs | [16] |
| Pontederia cordata | 81.00 b | 11.02 b | ||||||||
| Mixed Arundo donax and Pontederia cordata | 94.50 b | 12.20 b | ||||||||
| Control | SW | NA | 15 b | 1.22 b | 4 b | 25 a | 6.60 b | 0.24 b | [37] | |
| Typha latifolia | 20 d | 8.60 b | 0.33 b | Ps | ||||||
| 0.93–3.68 b,c | 25.70–98.70 b | 0.95–3.72 b | Afbs | |||||||
| 20 d and 0.93–3.68 b,c | 30.50–98.70 b | 1.17–3.74 b | Ps and Afbs | |||||||
| Calamus | SIS | 0 b | 13.10 b TN | TOC/TN = 0.62–0.72 b | 1 b | 23.6–31.5 b | 20.20 b TN | 2.65 b TN | [15] | |
| 0.10 b | Winter–Summer | 27.50–61.80 b TN | 3.60–8.10 b TN | |||||||
| Potamogeton crispus | SW | 0 b | 5.27 a | 2.50 | 5 b | 23.6 b | 41.6% b | 0.62 b | [50] | |
| 0.63 b | 68.6% b | 0.85 b | ||||||||
| Cattail litter, leaves, and stems | SIS | 0 b | 10.00 b | TOC/TN = 0.5 b | 1 b | 19.7–26.6 b | 20.50 b TN | 2.63 b TN | [18] | |
| 1.00 b | 43.20 b TN | 5.50 b TN | ||||||||
| Winter–Summer | 24.40–42.20 b TN | 3.50–5.88 b TN | ||||||||
| Corn stubble | SWB | 0 b | 18.00 b | NA | 2.13 b | 16.0 b | 38.50 b | 3.29 b | [19] | |
| 6.00–8.00 b | 21.00–22.00 b | 0.83h b | 15.8–24.6 b | ≥90.00 b | 20.80–22.40 b | 0–7d | ||||
| Reed straw | SIS | 0 b | 16.40 a | 0.80 b | 2–4 b | greenhouse | 14.40–26.80 b | 0.60–2.25 b | [17] | |
| 65.71 b | 62.10–87.40 b | 3.67–5.22 b | ||||||||
| Control | SIS | NA | 43.00 b | 2.44–2.52 b | 1 b | 12.5–24.6 b | 13.95–26.47 b | 6.00–9.00 b | [24] | |
| Wheat straw | 49.16–98.32 b | 21.13–42.28 b | ||||||||
| Cotton | 53.89–70.86 b | 23.17–30.46 b | ||||||||
| Woodchip | SIS | NA | 47.42–99.25 b | 1.87–0.98 b | 2 b | 20.0–29.0 b | 94.50–80.00 b | 22.41–39.70 b | [51] | |
| Only aerated without PCSs | SIS | 0 b | 41.79–44.24 b TN | 0.5–5 b | 3 b | 25.0 b | 3.98–32.60 b TN | 0.59–4.62 b TN | High ammonia concentration | [20] |
| Wheat straw | Carbon:substrate = 1:1 b | 59.20–96.90 b TN | 8.72–14.28 b TN | |||||||
| Apricot pit | 19.40–46.30 b TN | 2.86–6.55 b TN | ||||||||
| Walnut shell | 46.30 –62.90 b TN | 6.55–9.27 b TN | ||||||||
| Oenanthe javanica | SW | NA | 25.00 b TN | 0.6 b | 10 b | <10 b | 8.00 b TN | 0.20 b | [25] | |
| 26 d | 43.00 b TN | 1.08 b |
| PCSs | Major and Related Bacteria after PCS Addition and Their Proportion | Ref. |
|---|---|---|
| Control | Dechloromonasa (NA), Thauera a (NA), Paludibacter a,b (0.23%), Levilinea c (NA), Saccharofermentans d (<0.10%) | [24] |
| Wheat straw | Dechloromonasa (1.97%), Thauera a (1.02%), Paludibacter a,b (2.69%), Leviline c (0.91%), Saccharofermentans d (3.14%) | |
| Cotton | Dechloromonasa (3.92%), Thauera a (2.35%), Paludibacter a,b (NA), Levilinea c (NA) c, Saccharofermentans d (<0.10%) | |
| Control | Halomonasa (18.68%), Flavobacterium a (1.19%), Bacillus a (0.29%), Acidovorax a (0.11%), Azoarcus a (1.26%), Azospira a (0.16%), Azospirillum a (0.03%), Bradyrhizobium a (0.07%), Dechloromonas a (0.23%), Hyphomicrobium a (0.15%), Mesorhizobium a (0.07%), Pseudomonas a (9.64%) | [25] |
| Oenanthe javanica | Halomonasa (14.49%), Flavobacterium a (2.79%), Bacillus a (0.27%), Paenibacillus a (0.01%), Acidovorax a (0.03%), Azoarcus a (1.15%), Azospira a (0.16%), Azospirillum a (0.15%), Bradyrhizobium a (0.09%), Dechloromonas a (0.31%), Hyphomicrobium a (0.09%), Mesorhizobium a (0.15%), Pseudomonas a (5.19%) | |
| Control | Bacillusa (0.23%), Flavobacterium a (1.00%), Hyphomicrobium a (0.12%), Rhodobacter a (0.01%), Comamonas a (0.06%), Hydrogenophaga a (0.05%), Azospira a (0.01%), Dechloromonas a (0.14%), Thauera a (0.08%) | [15] |
| Calamus litter | Bacillusa (4.96%), Flavobacterium a (8.19%), Hyphomicrobium a (0.06%), Rhodobacter a (0.18%), Comamonas a (0.04%), Hydrogenophaga a (0.38%), Azospira a (0.04%), Dechloromonas a (1.63%), Thauera a (0.03%) | |
| Control | Dechloromonasa (1.03%), Rhodobacter a (NA), Thiobacillus a (NA) | [18] |
| Cattail litter | Dechloromonasa (6.67%), Rhodobacter a (NA), Thiobacillus a (NA) | |
| Control | Halomonasa (27.81%) | [91] |
| Iris pseudacorus | Flavobacteriuma (18.28%), Halomonas a (2.91%), Aeromonas a (1.81%), Pseudomonas a (1.68%), Dechloromonas a (1.29%), Azospira a (0.67%), Shewanella a (0.56%), Arcobacter a (0.24%) | |
| Typha orientalis | Flavobacteriuma (19.47%), Halomonas a (12.21%) | |
| starch/ PCL | Bacillusa (24.25%), Thauera a (9.36%), Acidovorax a (5.37%), Chlorobium e (7.06%), Desulfobacter f (6.15%), Desulfobulbus g (6%), Desulfovibrio g (5.26%) | [93] |
| Control | Thaueraa (5.70%), Dechloromonas a (2.20%), Flexibacter a (1.90%), Thiobacillus a (1.50%), Anaerolineaceae a (1.30%), Rhodobacter a (1.20%), Haliangium a (1.20%) | [87] |
| Typha latifolia | Paracoccusa (1.90%), Hyphomicrobium b (1.80%), Novosphingobium b (1.70%), Nocardioides a (1.60%), Pseudonocardia a (1.50%), Sinobacteraceae a (1.40%), Flexibacter a (1.30%), Nocardioides a (1.60%), Pseudonocardia a (1.50%), Sinobacteraceae a (1.40%), Flexibacter a (1.30%) | |
| Cattail litter | Thaueraa (6.20%), Dechloromonas a (4.40%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Dong, W.; Yan, G.; Wang, H.; Wang, H.; Chang, Y.; Yu, S.; Chu, Z.; Ling, Y.; Li, C. Plant Carbon Sources for Denitrification Enhancement and Its Mechanism in Constructed Wetlands: A Review. Sustainability 2022, 14, 12545. https://doi.org/10.3390/su141912545
Zhang Y, Dong W, Yan G, Wang H, Wang H, Chang Y, Yu S, Chu Z, Ling Y, Li C. Plant Carbon Sources for Denitrification Enhancement and Its Mechanism in Constructed Wetlands: A Review. Sustainability. 2022; 14(19):12545. https://doi.org/10.3390/su141912545
Chicago/Turabian StyleZhang, Yanjie, Weiyang Dong, Guokai Yan, Haiyan Wang, Huan Wang, Yang Chang, Shan Yu, Zhaosheng Chu, Yu Ling, and Congyu Li. 2022. "Plant Carbon Sources for Denitrification Enhancement and Its Mechanism in Constructed Wetlands: A Review" Sustainability 14, no. 19: 12545. https://doi.org/10.3390/su141912545