
Acclimatization of Mediterranean Native Sages (Salvia spp.) and Interspecific Hybrids in an Urban Green Roof under Regular and Reduced Irrigation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Substrate and Experimental Setup
2.2. Irrigation Scheduling
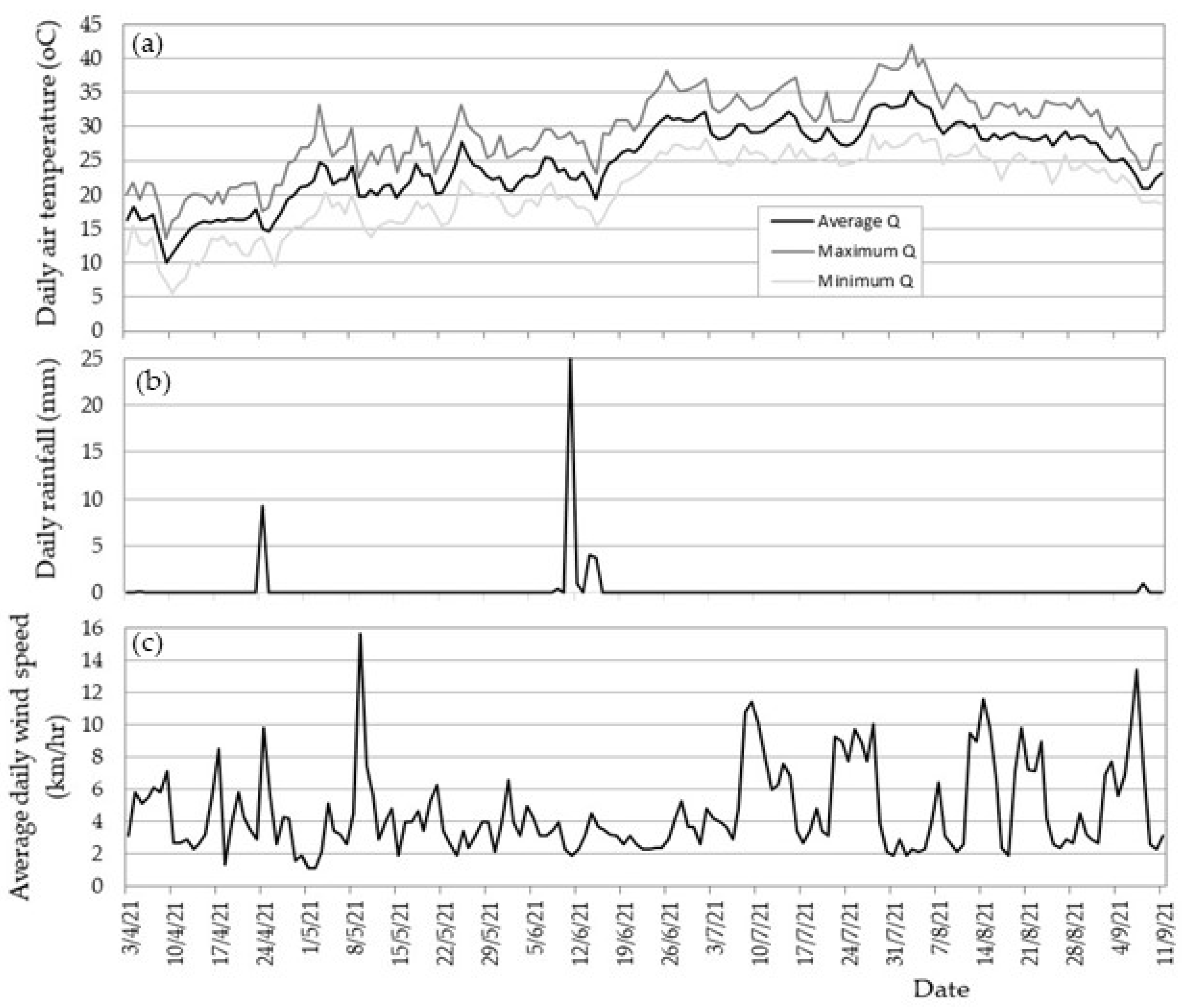
2.3. Meteorological Data
2.4. Plant Growth and Physiological Parameters Evaluation
2.5. Statistical Analysis
3. Results
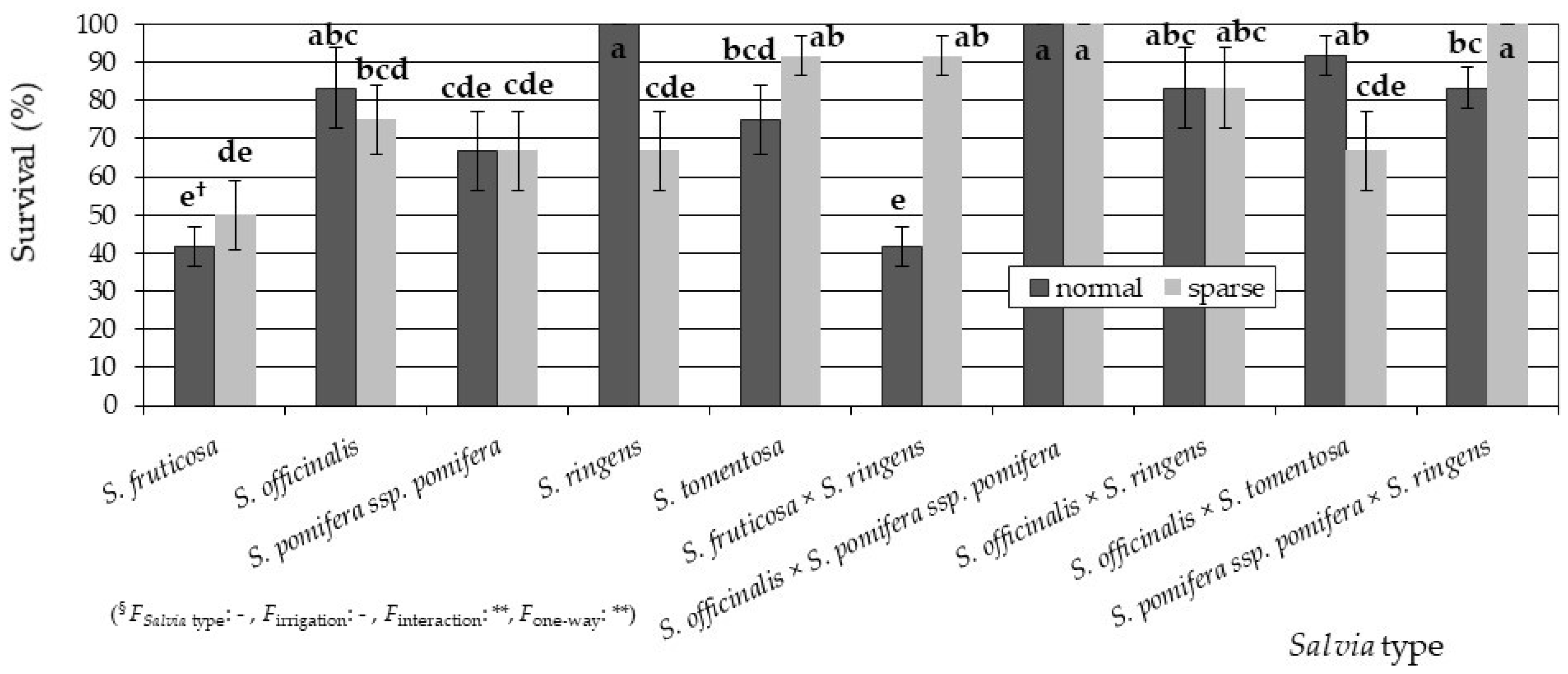
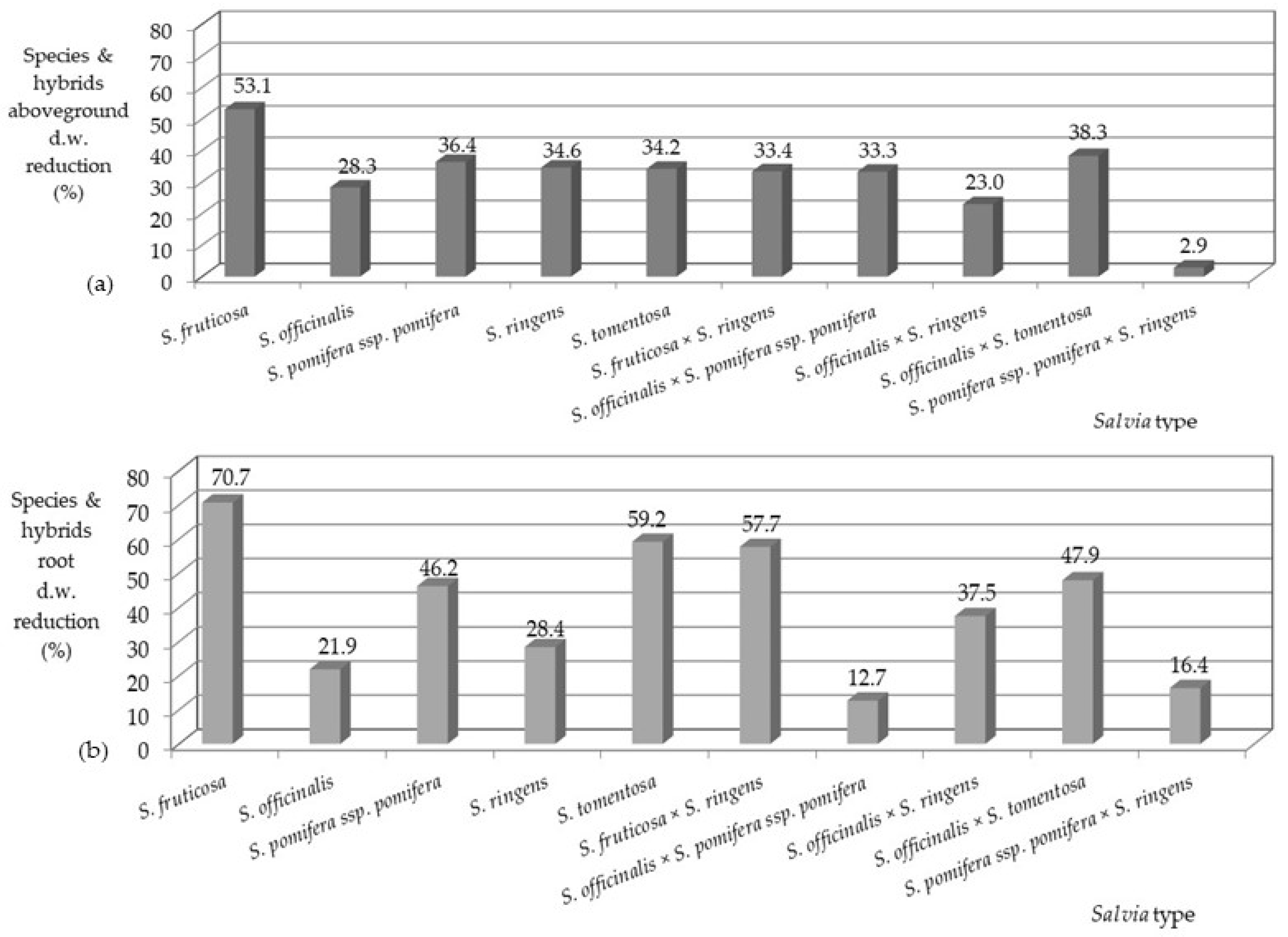
3.1. Plant Growth and Survival
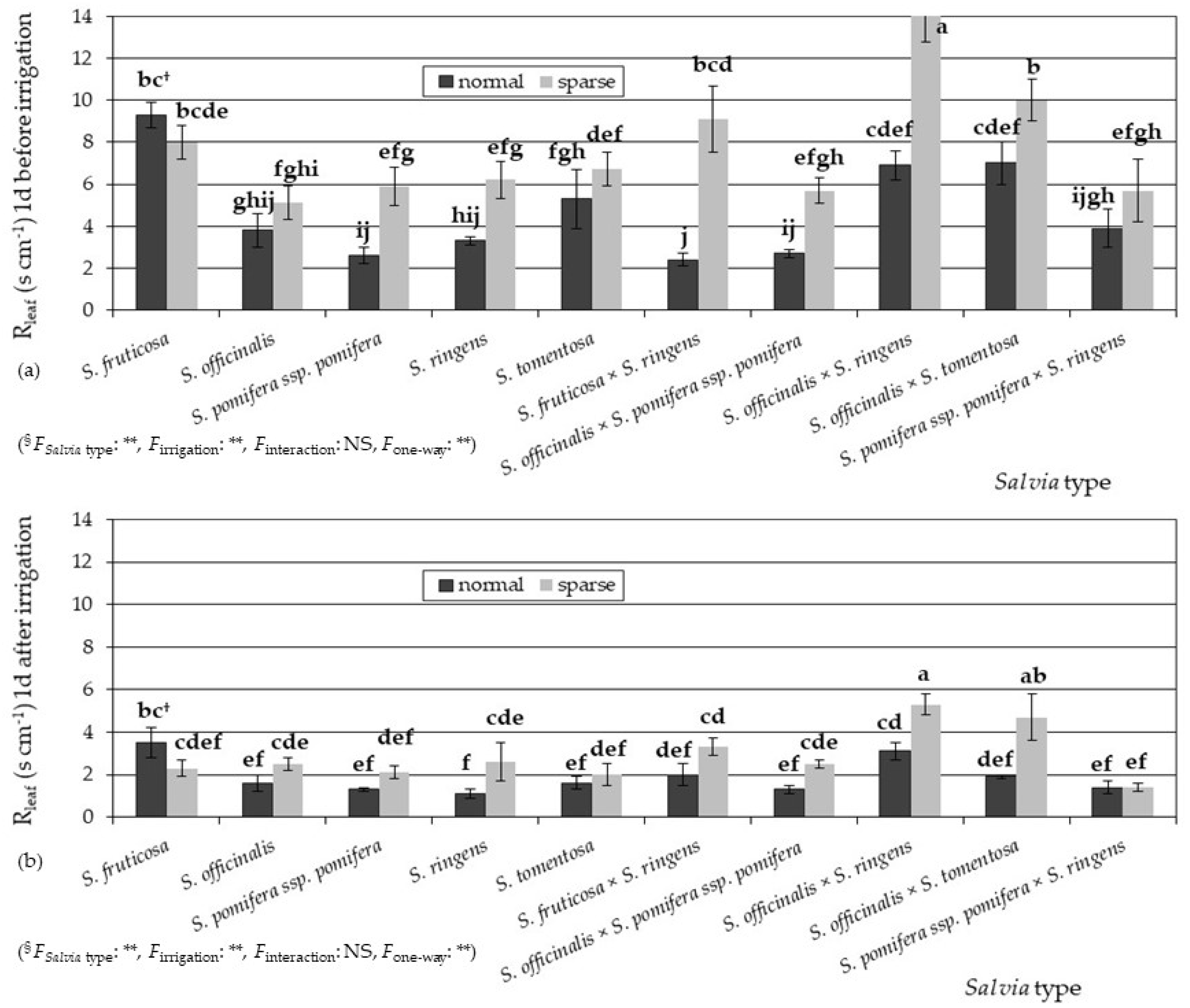
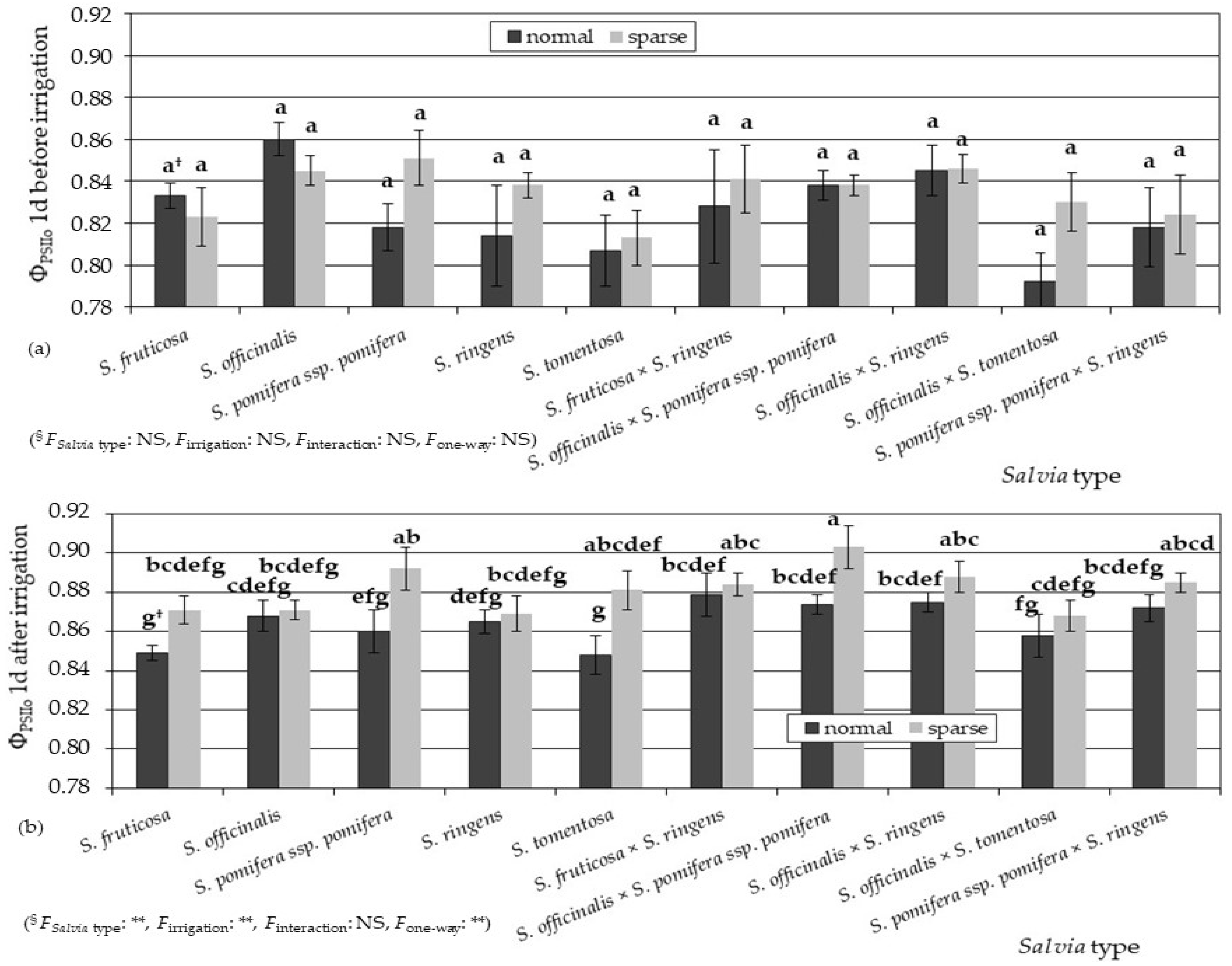
3.2. Physiological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Whittinghill, L.J.; Rowe, D.B. The role of green roof technology in urban agriculture. Ren. Agric. Food Syst. 2012, 27, 314–322. [Google Scholar] [CrossRef]
- Berardi, U.; Ghaffarian Hoseini, A.H.; Ghaffarian Hoseini, A. State-of-the-art analysis of the environmental benefits of green roofs. Appl. Energ. 2014, 115, 411–428. [Google Scholar] [CrossRef]
- Caneva, G.; Kumbaric, A.; Savo, V.; Casalini, R. Ecological approach in selecting extensive green roof plants: A data-set of Mediterranean plants. Plant Biosyst. 2015, 149, 374–383. [Google Scholar] [CrossRef]
- Shafique, M.; Kim, R.; Rafiq, M. Green roof benefits, opportunities and challenges—A review. Renew. Sustain. Energy Rev. 2018, 90, 757–773. [Google Scholar] [CrossRef]
- Wooster, E.I.F.; Fleck, R.; Torpy, F.; Ramp, D.; Irga, P.J. Urban green roofs promote metropolitan biodiversity: A comparative case study. Build. Environ. 2022, 207, 108458. [Google Scholar] [CrossRef]
- Cook, L.M.; Larsen, T.A. Towards a performance-based approach for multifunctional green roofs: An interdisciplinary review. Build. Environ. 2021, 188, 107489. [Google Scholar] [CrossRef]
- Joshi, M.Y.; Teller, J. Urban Integration of Green Roofs: Current Challenges and Perspectives. Sustainability 2021, 13, 12378. [Google Scholar] [CrossRef]
- Kweon, B.S.; Sullivan, W.C.; Wiley, A.R. Green common spaces and the social integration of inner-city older adults. Environ. Behav. 1998, 30, 832–858. [Google Scholar] [CrossRef]
- Savi, T.; Dal Borgo, A.; Love, V.L.; Andri, S.; Tretiach, M.; Nardini, A. Drought versus heat: What’s the major constraint on Mediterranean green roof plants? Sci. Total Environ. 2016, 566, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Koroxenidis, E.; Theodosiou, T. Comparative environmental and economic evaluation of green roofs under Mediterranean climate conditions—Extensive green roofs a potentially preferable solution. J. Clean. Product. 2021, 311, 127563. [Google Scholar] [CrossRef]
- Stathers, R.; Schroders, U.K. The bee and the stock market: An overview of pollinator decline and its economic and corporate significance. In The Business of Bees, 1st ed.; Atkins, J., Atkins, B., Eds.; Routledge: London, UK, 2017; pp. 124–144. [Google Scholar] [CrossRef]
- Benvenuti, S.; Bacci, D. Initial agronomic performances of Mediterranean xerophytes in simulated dry green roofs. Urban Ecosyst. 2010, 13, 349–363. [Google Scholar] [CrossRef]
- Tassoula, L.; Papafotiou, M.; Liakopoulos, G.; Kargas, G. Water use efficiency, growth and anatomic-physiological parameters of Mediterranean xerophytes as affected by substrate and irrigation on a green roof. Notul. Bot. Hort. Agrobot. 2021, 49, 12283. [Google Scholar] [CrossRef]
- Ondoño, S.; Martínez-Sánchez, J.J.; Moreno, J.L. Evaluating the growth of several Mediterranean endemic species in artificial substrates: Are these species suitable for their future use in green roofs? Ecolog. Engin. 2015, 81, 405–417. [Google Scholar] [CrossRef]
- Papafotiou, M.; Pergialioti, N.; Tassoula, L.; Massas, I.; Kargas, G. Growth of native aromatic xerophytes in an extensive Mediterranean green roof as affected by substrate type and depth and irrigation frequency. HortScience 2013, 48, 1327–1333. [Google Scholar] [CrossRef]
- Azeñas, V.; Janner, I.; Medrano, H.; Gulías, J. Performance evaluation of five Mediterranean species to optimize ecosystem services of green roofs under water-limited conditions. J. Environ. Manag. 2018, 212, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Paço, T.A.; Cruz de Carvalho, R.; Arsénio, P.; Martins, D. Green Roof Design Techniques to Improve Water Use under Mediterranean Conditions. Urban Sci. 2019, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Bourgou, S.; Ben Haj Jilani, I.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; et al. Medicinal-Cosmetic Potential of the Local Endemic Plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for Conservation and Sustainable Exploitation of Neglected and Underutilized Phytogenetic Resources. Biology 2021, 10, 1344. [Google Scholar] [CrossRef]
- Krigas, N.; Tsoktouridis; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M.A.; et al. Exploring the Potential of Neglected Local Endemic Plants of Three Mediterranean Regions in the Ornamental Sector: Value Chain Feasibility and Readiness Timescale for Their Sustainable Exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Papafotiou, M.; Martini, A.N.; Papanikolaou, E.; Stylias, E.G.; Kalantzis, A. Hybrids Development between Greek Salvia Species and Their Drought Resistance Evaluation along with Salvia fruticosa, under Attapulgite-Amended Substrate. Agronomy 2021, 11, 2401. [Google Scholar] [CrossRef]
- Savi, T.; Andri, S.; Nardini, A. Impact of different green roof layering on plant water status and drought survival. Ecolog. Engin. 2013, 57, 188–196. [Google Scholar] [CrossRef]
- Raimondo, F.; Trifilò, P.; Lo Gullo, M.A.; Andri, S.; Savi, T.; Nardini, A. Plant performance on Mediterranean green roofs: Interaction of species-specific hydraulic strategies and substrate water relations. AoB Plants 2015, 7, plv007. [Google Scholar] [CrossRef] [PubMed]
- Papafotiou, M.; Koutri, A.; Massas, I. Heavy metal concentration in sage plants cultivated on an urban green roof or roadside location as affected by substrate type and fertilization. Acta Hort. 2017, 1189, 439–442. [Google Scholar] [CrossRef]
- Kemp, S.; Hadley, P.; Blanuša, T. The influence of plant type on green roof rainfall retention. Urban Ecosyst. 2019, 22, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Kokkinou, I.; Ntoulas, N.; Nektarios, P.A.; Varela, D. Response of Native Aromatic and Medicinal Plant Species to Water Stress on Adaptive Green Roof Systems. HortScience 2016, 51, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.E.; Munné-Bosch, S. Salicylic acid may be involved in the regulation of drought-induced leaf senescence in perennials: A case study in field-grown Salvia officinalis L. plants. Environ. Exper. Bot. 2008, 64, 105–112. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Mueller, M.; Schwarz, K.; Alegre, L. Diterpenes and antioxidative protection in drought-stressed Salvia officinalis plants. J. Plant Physiol. 2001, 158, 1431–1437. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Abate, E.; Azzarà, M.; Trifilò, P. When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics. Plants 2021, 10, 1888. [Google Scholar] [CrossRef]
- Abate, E.; Nardini, A.; Petruzzellis, F.; Trifilò, P. Too dry to survive: Leaf hydraulic failure in two Salvia species can be predicted on the basis of water content. Plant Physiol. Bioch. 2021, 166, 215–224. [Google Scholar] [CrossRef]
- Gonceariuc, M. Reducing the impact of drought on productivity by cultivating resistant varieties of medicinal and aromatic plants. Buletinul AŞM Ştiinţele Vieţii 2019, 2, 95–103. [Google Scholar]
- FLL. Guideline for the Planning, Execution and Upkeep of Green-Roof Sites, English ed.; Forschungsgesellschaft Landschaftsentwicklung Landschaftsbau: Bonn, Germany, 2010. [Google Scholar]
- Abobatta, W.F. Drought adaptive mechanisms of plants—A review. Adv. Agr. Environ. Sci. 2019, 2, 42–45. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; Nie, Y.; Bai, S.H.; Zhou, L.; Shao, J.; Cheng, W.; Wang, J.; Hu, F.; Fu, Y. Drought-induced changes in root biomass largely result from altered root morphological traits: Evidence from a synthesis of global field trials. Plant Cell Environ. 2018, 41, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, J.J.; Morales, M.A.; Ferrández, T.; Sánchez-Blanco, M.J. Effects of water and salt stresses on growth, water relations and gas exchange in Rosmarinus officinalis. J. Hortic. Sci. Biotechnol. 2006, 81, 845–853. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Savi, T.; Marin, M.; Luglio, J.; Petruzzellis, F.; Mayr, S.; Nardini, A. Leaf hydraulic vulnerability protects stem functionality under drought stress in Salvia officinalis. Funct. Plant Biol. 2016, 43, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Nagase, A.; Dunnett, N. Drought tolerance in different vegetation types for extensive green roofs: Effects of watering and diversity. Landsc. Urb. Plan. 2010, 97, 318–327. [Google Scholar] [CrossRef]
- Toscano, S.; Branca, F.; Romano, D.; Ferrante, A. An evaluation of different parameters to screen ornamental shrubs for salt spray tolerance. Biology 2020, 9, 250. [Google Scholar] [CrossRef]
- Posch, S.; Bennett, L.T. Photosynthesis, photochemistry and antioxidative defence in response to two drought severities and with re-watering in Allocasuarina luehmannii. Plant Biol. 2009, 11, 83–93. [Google Scholar] [CrossRef]
- Akoumianaki-Ioannidou, A.; Podaropoulou, L.; Liakopoulos, G. Assessment of growth of Mentha pulegium in a green roof system through the study of PSII photochemical parameters. Acta Hortic. 2017, 1189, 257–262. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernández, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins. J. Exper. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2-Way Anova | Aboveground d.w. (g) | Root d.w. (g) | Root d.w./ Aboveground d.w. | Aboveground f.w. (g) | Root f.w. (g) | Root f.w./ Aboveground f.w. |
|---|---|---|---|---|---|---|
| S. fruticosa | 74 cd | 48.1 cd † | 0.6 cd | 209.0 bc | 188.0 | 0.8 |
| S. officinalis | 60.3 e | 85.4 a | 1.4 a | 155.5 e | 268.5 | 1.7 |
| S. pomifera ssp. pomifera | 87.9 b | 39.6 d | 0.4 d | 216.8 bcd | 142.5 | 0.7 |
| S. ringens | 71.6 cde | 52.9 cd | 0.7 bc | 204.8 bcde | 175.8 | 0.9 |
| S. tomentosa | 90.7 ab | 65.5 bc | 0.7 bc | 215.3 bcd | 216.1 | 1.0 |
| S. fruticosa × S. ringens | 69.3 cde | 43.2 d | 0.6 cd | 167.2 de | 167.7 | 1.0 |
| S. officinalis × S. pomifera ssp. pomifera | 86.3 b | 70.1 ab | 0.8 b | 222.3 ab | 287.8 | 1.3 |
| S. officinalis × S. ringens | 103.5 a | 74.2 ab | 0.7 bc | 267.7 a | 266.6 | 1.0 |
| S. officinalis × S. tomentosa | 68.8 de | 47.0 d | 0.7 bc | 175.2 de | 162.2 | 0.9 |
| S. pomifera ssp. pomifera × S. ringens | 71.8 cde | 44.9 d | 0.6 cd | 183.4 cde | 150.6 | 0.8 |
| Normal | 93.6 a | 71.4 a | 0.8 a | 239.1 a | 252.1 | 1.1 |
| Sparse | 63.2 b | 42.8 b | 0.7 b | 164.3 b | 152.9 | 0.9 |
| Significance § | ||||||
| FSalvia type | ** | ** | ** | ** | - | - |
| Firrigation | ** | ** | ** | ** | - | - |
| FSalvia type × irrigation | NS | NS | NS | NS | * | ** |
| Salvia Species | Irrigation Frequency | Aboveground d.w. (g) | Root d.w. (g) | Root d.w./ Aboveground d.w. | Aboveground f.w. (g) | Root f.w. (g) | Root f.w./ Aboveground f.w. |
|---|---|---|---|---|---|---|---|
| S. fruticosa | Normal | 100.8 ± 2.7 ab † | 74.5 ± 11.4 ab | 0.7 ± 0.1 bc | 276.9 ± 8.7 ab | 301.5 ± 29.8 ab | 1.1 ± 0.1 cd |
| Sparse | 47.3 ± 1.5 e | 21.8 ± 1.8 f | 0.5 ± 0.0 bcd | 141.1 ± 10.9 ef | 74.5 ± 5.0 f | 0.5 ± 0.0 f | |
| S. officinalis | Normal | 70.3 ± 5.9 cd | 95.9 ± 11.2 a | 1.4 ± 0.1 a | 174.0 ± 21.2 def | 294.2 ± 31.2 ab | 1.7 ± 0.1 a |
| Sparse | 50.4 ± 6.6 e | 74.9 ± 10.7 ab | 1.5 ± 0.1 a | 137.0 ± 17.4 f | 242.9 ± 32.5 bc | 1.8 ± 0.1 a | |
| S. pomifera ssp. pomifera | Normal | 107.5 ± 8.5 a | 51.5 ± 6.5 cde | 0.5 ± 0.1 bcd | 256.9 ± 22.2 ab | 179.9 ± 20.3 cde | 0.7 ± 0.1 def |
| Sparse | 68.4 ± 4.6 cd | 27.7 ± 1.3 f | 0.4 ± 0.0 d | 176.6 ± 9.9 def | 105.1 ± 5.7 ef | 0.6 ± 0.0 ef | |
| S. ringens | Normal | 86.6 ± 6.0 bc | 61.6 ± 8.0 bc | 0.7 ± 0.1 bc | 244.3 ± 18.4 bc | 201.5 ± 23.9 cde | 0.8 ± 0.1 de |
| Sparse | 56.6 ± 6.0 de | 44.1 ± 6.3 de | 0.8 ± 0.0 b | 165.2 ± 14.4 ef | 148.2 ± 34.0 de | 0.9 ± 0.2 d | |
| S. tomentosa | Normal | 109.4 ± 7.7 a | 93.1 ± 27.7 a | 0.8 ± 0.2 b | 274.1 ± 29.5 ab | 295.5 ± 68.4 ab | 1.0 ± 0.1 cd |
| Sparse | 72.0 ± 5.9 cd | 38.0 ± 9.2 ef | 0.5 ± 0.1 bcd | 156.4 ± 17.3 ef | 136.6 ± 27.7 ef | 0.9 ± 0.1 d | |
| S. fruticosa × S. ringens | Normal | 83.2 ± 3.0 bc | 60.7 ± 2.9 bc | 0.7 ± 0.1 bc | 195.3 ± 0.3 de | 233.7 ± 5.7 bc | 1.2 ± 0.0 bc |
| Sparse | 55.4 ± 2.1 e | 25.7 ± 6.0 f | 0.5 ± 0.1 bcd | 139.1 ± 11.0 f | 101.7 ± 20.8 ef | 0.7 ± 0.1 def | |
| S. officinalis × S. pomifera ssp. pomifera | Normal | 103.6 ± 7.6 ab | 74.8 ± 5.8 ab | 0.7 ± 0.0 bc | 262.9 ± 18.1 ab | 315.0 ± 19.4 ab | 1.2 ± 0.0 bc |
| Sparse | 69.1 ± 4.6 cd | 65.3 ± 7.3 bc | 0.9 ± 0.1 b | 181.8 ± 11.5 def | 260.6 ± 26.0 abc | 1.4 ± 0.1 b | |
| S. officinalis × S. ringens | Normal | 116.9 ± 6.5 a | 91.3 ± 9.5 a | 0.8 ± 0.1 b | 301.2 ± 16.4 a | 322.6 ± 30.4 a | 1.1 ± 0.1 cd |
| Sparse | 90.0 ± 2.6 b | 57.1 ± 3.0 bc | 0.6 ± 0.0 bc | 234.2 ± 5.8 bc | 210.6 ± 6.3 cd | 0.9 ± 0.0 d | |
| S. officinalis × S. tomentosa | Normal | 85.1 ± 4.4 bc | 61.8 ± 6.0 bc | 0.7 ± 0.1 bc | 212.2 ± 9.9 cd | 214.0 ± 17.1 cd | 1.0 ± 0.1 cd |
| Sparse | 52.5 ± 4.9 e | 32.2 ± 4.4 ef | 0.6 ± 0.1 cd | 138.2 ± 9.7 f | 110.4 ± 11.7 ef | 0.8 ± 0.1 de | |
| S. pomifera ssp. pomifera × S. ringens | Normal | 72.9 ± 6.7 cd | 48.9 ± 7.5 cde | 0.7 ± 0.1 bc | 193.4 ± 16.6 de | 162.6 ± 23.0 cde | 0.8 ± 0.1 de |
| Sparse | 70.8 ± 2.3 cd | 40.9 ± 7.6 def | 0.6 ± 0.1 bc | 173.4 ± 5.3 def | 138.5 ± 21.2 ef | 0.8 ± 0.1 de | |
| Significance § | |||||||
| Fone-way ANOVA | ** | ** | ** | ** | ** | * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papafotiou, M.; Martini, A.N.; Tassoula, L.; Stylias, E.G.; Kalantzis, A.; Dariotis, E. Acclimatization of Mediterranean Native Sages (Salvia spp.) and Interspecific Hybrids in an Urban Green Roof under Regular and Reduced Irrigation. Sustainability 2022, 14, 4978. https://doi.org/10.3390/su14094978
Papafotiou M, Martini AN, Tassoula L, Stylias EG, Kalantzis A, Dariotis E. Acclimatization of Mediterranean Native Sages (Salvia spp.) and Interspecific Hybrids in an Urban Green Roof under Regular and Reduced Irrigation. Sustainability. 2022; 14(9):4978. https://doi.org/10.3390/su14094978
Chicago/Turabian StylePapafotiou, Maria, Aikaterini N. Martini, Lamprini Tassoula, Eleftherios G. Stylias, Anastasios Kalantzis, and Eleftherios Dariotis. 2022. "Acclimatization of Mediterranean Native Sages (Salvia spp.) and Interspecific Hybrids in an Urban Green Roof under Regular and Reduced Irrigation" Sustainability 14, no. 9: 4978. https://doi.org/10.3390/su14094978