
Optimal Mowing Regime in Enhancing Biodiversity in Seasonal Floodplains along Engineered Channels


1
Division of Ecology & Biodiversity, School of Biological Sciences, The University of Hong Kong, Pok Fu Lam, Hong Kong
2
Outdoor Wildlife Learning Hong Kong, Hong Kong
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(7), 4002; https://doi.org/10.3390/su14074002
Submission received: 3 March 2022
/
Revised: 23 March 2022
/
Accepted: 25 March 2022
/
Published: 28 March 2022
Abstract
:The vegetation grown on grasscretes along channelized rivers have been regularly mown in Hong Kong. However, no baseline information on the relationship between different mowing regimes and the biodiversity of such riverbed vegetation is available. We therefore carried out a manipulative experiment along a channelized river to test the effect of mowing frequency and intensity on the abundance and species richness of terrestrial biodiversity. We conducted point counts and transect counts to survey birds, butterflies and Odonates, night surveys for amphibians and reptiles, quadrat surveys for vegetation and sweep net and pan traps for other macroinvertebrates. The results from Generalized Linear Mixed Models (GLMMs) showed a taxon-specific effect of mowing regime. Bird species richness was significantly suppressed in plots mown with high frequency. Both butterfly abundance and species richness were greatly enhanced by low intensity and low frequency mowing. Odonate abundance, and the abundance and species richness of other macroinvertebrates remained high whenever a portion of vegetation was retained as refuge. Amphibians and reptiles did not prefer to utilize the vegetation grown on grasscretes, and thus showed no impact from different mowing regimes. The overall species richness of vegetation was not affected by mowing regimes, but the domination of tall invasive Brachiaria mutica was suppressed by any mowing activity. To cater for the need of most taxa, we propose a mosaic mowing regime, in which most parts along the channelized river could be mown infrequently to 600 mm tall while some of the patches remain unmown.
1. Introduction
In response to the rising awareness of biodiversity conservation, green channel designs such as rip-rap lining or grasscrete have been incorporated into drainage projects as alternatives to traditional concrete cover in many channelized lowland rivers, including those in Hong Kong [1,2]. These would enable vegetation to grow on the engineered riverbed, which supports other wildlife. Meanwhile, regular mowing along the channelized rivers is required to maintain the vegetation height for regulating flood prevention capacity and aesthetic enhancement. It would be ideal to further enhance biodiversity by implementing an optimal mowing regime (that is, with the optimal mowing frequency and height) without compromising the aforementioned goals.
Currently, there is no baseline information on the relationship between different mowing regimes and biodiversity of such riverbed vegetation. Most research on the effect of mowing regimes on biodiversity were conducted on agricultural, urbanized or natural grasslands [3,4,5], in which numerous environmental factors such as the association with river and soil properties are totally different from those along engineered channels. Furthermore, around 90% of these research projects on mowing regimes were conducted in temperate regions in Europe and North America [5]. Given the great differences in composition and adaptations of the species and the climate between Hong Kong (a subtropical city) and those from the temperate Holoarctic region, the recommended mowing regimes in these regions may not be suitable for Hong Kong and other non-temperate areas. A study is therefore needed to test different mowing regimes for providing useful results in the formulation of future mowing practices benefiting biodiversity along engineered channels. Moreover, since very limited information on the fauna and flora in riverbed vegetation along channelized rivers are available, opportunities could be taken to perform a post-establishment ecological study on the selected green channel.
By carrying out a manipulative experiment at a channelized river site in the New Territories of Hong Kong, this study was commissioned to provide ecological data of riverbed vegetation under different mowing height and frequency for developing future zonal manipulation schemes. Specifically, this study has the following aims: (1) to generate the baseline information on the biodiversity of channelized riverbed vegetation, (2) to study the changes in fauna and flora communities in the riverbed vegetation under different mowing regimes (and controls) and (3) to recommend mowing considerations that optimize biodiversity in the tested channelized river habitat.
2. Materials and Methods
2.1. Study Site
A section of Tan Shan River next to Ko Po North (22.51453° N, 114.16455° E) in the New Territories of Hong Kong SAR was selected as the study site for the zonal manipulative experiment from June 2020 to April 2021 (Figure 1A). The site has a subtropical monsoonal climate composed of a wet summer and a dry winter, with a monthly minimum mean temperature of around 15 °C in January (winter) and a monthly maximum mean temperature of around 28 °C in July (summer). The annual rainfall is around 1900 mm. Rip-rap lining, grassed cellular concrete paving (grasscrete) and hydroseeding were applied on the riverbed at both sides of the low-flow channel in 2011. The seed mixture used in hydroseeding was dominated by Cynodon dactylon (native), Paspalum notatum (exotic) and Lolium perenne (exotic) [6]. The bottom of the low-flow channel was converted into concrete, whereas the riverbed was covered by rip-rap lining and grasscrete paving, which are now dominated by exotic weeds such as Bidens alba, Brachiaria mutica and Wedelia trilobata, together with some native herbaceous plants such as Commelina diffusa and Callipteris esculenta.
2.2. Experimental Set Up
The study site was divided into three zones, each with four treatment plots (i.e., Treatment A to D) and a control plot arranged randomly (Figure 1C). The plot area was on average 139 m2 (±SE 14 m2). To investigate the effect of mowing frequency, Treatments A and B, representing treatments with higher mowing frequencies, were mowed once every month in the wet season from June to October and once every two months in the dry season from November to April. On the other hand, Treatments C and D, representing treatments with low mowing frequencies, were mowed once every three months throughout the study period (Table 1). To examine the effect of mowing intensity, Treatment A and C were mowed to the ground level while Treatment B and D were mowed to a higher vegetation height at around 600 mm. No mowing was applied to the control plots. A minimum of 1 m vegetation free buffer strip was maintained between all field plots.
2.3. Biodiversity Monitoring
Monthly ecological surveys were conducted for each plot during the study period from June 2020 to April 2021, targeting birds, amphibians and reptiles, butterflies, Odonates, terrestrial macroinvertebrates and plants. Two additional surveys were conducted for plots that had been mowed in the respective months (see Section 2.4 for details, and see Table S1 for more details in survey dates). To enhance the understanding of biodiversity within the study area, casual observations for all taxa except terrestrial macroinvertebrates and plants made outside the treatment plots during the surveys were recorded as ‘general records’ for each survey.
2.4. Monthly Surveys
Monthly surveys were conducted in all plots targeting the taxa mentioned above (Table 2). In each month, both vegetation and fauna surveys were conducted. Five survey methods were employed for fauna—(a) point count, (b) transect count, (c) night survey, (d) sweep net and (e) pan-trap (Table 2). Two additional fauna surveys, including methods (a), (b), (c) and (d), were conducted in plots that had been mowed in the respective months. All the fauna surveys using (a) to (e) were conducted within 7 days after mowing.
2.5. Vegetation Surveys
The monthly vegetation survey was conducted at least three weeks after mowing. During each survey, three 1 m × 1 m quadrats were set randomly in each plot, where the percentage coverage and height of each species, except for climbers, were recorded. In addition, rare, protected or threatened plant species were recorded with GPS locations whenever they were discovered within the study, as they were throughout the study.
2.6. Fauna Surveys
2.6.1. Point Count (Birds, Butterflies and Odonates)
All point count surveys were conducted between 9 am and 11 am. All plots were monitored simultaneously by two groups of surveyors with binoculars for 30 min. Only target taxa using the plots (e.g., foraging, resting, hiding or landing) were identified and recorded. During the surveys, surveyors were positioned high on top of the gabion wall to ensure visual coverage of all plots.
2.6.2. Transect Count (Birds, Butterflies and Odonates)
A five-minute random walk within each plot was used to flush out the target taxa, which were identified and recorded with the aid of binoculars. As in the point count, only target taxa using the plots were recorded.
2.6.3. Sweep Net (Butterflies, Odonates and Other Terrestrial Macroinvertebrates)
A total of six sweeps were made at the height of the vegetation of each plot with a circular sweep net (diameter: 30 cm; mesh size: 1 mm) while walking at a constant speed. Captured macroinvertebrates were transferred into a zip-lock bag and placed into an ice bag immediately to inhibit sample loss caused by predation. The sweep net samples were brought back to a laboratory for further identification.
2.6.4. Night Surveys (Amphibians and Reptiles)
Call survey and transect walk were conducted during the night survey for amphibians and reptiles during the wet season (June–September). In each survey, surveyors stood at the center of the study site with the light off and listened to frog calls for 10 min prior to the transect count. Calls heard were identified and recorded for each plot. Afterwards, a random transect walk within each plot was conducted, and the target taxa encountered were recorded.
2.6.5. Pan Trap (Other Terrestrial Macroinvertebrates)
Four colored pan traps 13 cm in diameter and 6 cm deep, painted with UV-bright fluorescent paints (red, yellow, blue and white), were installed randomly on two metal poles within each plot at the height of the vegetation. The variety of colors used aimed at collecting a wider range of floral-visiting arthropods [7,8]. Each pan trap contained soap water as the killing agent, and holes were drilled near the upper margin to prevent overflow of specimens during rain. In each survey, the trap was left in the field for 24–48 h, and specimens inside the traps were then sieved through a kitchen strainer (mesh size of 0.5 mm). Next, the sieved materials were transferred into a 100 mL container using 70% ethanol. The samples were then brought back to the laboratory for further identification.
2.7. Data Analysis
To investigate the effects of mowing regimes on fauna and flora biodiversity across seasons, Generalized Linear Mixed Models (GLMMs) were constructed with species richness and abundance of each taxon as the dependent variables, and ‘Treatment’, ‘Season’ and their interaction term as independent variables using the R package glmmTMB. ‘Season’ was defined as below: May–August was regarded as the ‘Wet season’, September–October and March–April as the ‘Transitional season’ and ‘November–February’ as the ‘Dry season’. ‘Zone’ was incorporated into the model as covariate and ‘Month’ was used as the random factor. For macroinvertebrates, the sampling methods, i.e., sweep net vs. pan trap, were added to the model as an independent variable. Whenever treatment effect was significant, comparison across treatments was carried out by calculating the least-squares means using the R package emmeans.
3. Results
3.1. Overview
A total of 85 taxa of amphibians and reptiles, birds, butterflies and Odonates were recorded utilizing treatments or control plots between June 2020 and April 2021, which included four amphibians, three reptiles, 15 birds, 43 butterflies and 20 Odonates, 12 of which (out of the 85 taxa) were species of conservation concern (Table S1). These included one bird species, Cisticola juncidis, which was listed as being of “Local Concern” by Fellowes et al. [9]. Nine butterfly species are considered by the Agriculture, Fisheries and Conservation Department (AFCD) [10] of Hong Kong either as “Uncommon” (Ampittia dioscorides etura, Danaus chrysippus chrysippus, Euchrysops cnejus cnejus and Hypolimnas misippus (also listed as “Local Concern” by Fellowes et al. [9])), “Rare” (Aeromachus jhora, Pieris rapae crucivora and Udaspes folus), or “Very Rare” (Catochrysops strabo strabo and Polygonia c-aureum c-aureum (also classified as species of conservation concern by AFCD [10]). Lastly, two Odonate species, Onychothemis tonkinensis (also listed as “Uncommon by AFCD [10]) and Paracercion calamorum, were listed as “Local Concern” [9].
For other terrestrial macroinvertebrates, a total of 673 morphospecies in 181 families and 19 orders were recorded from samples from the sweep net (509 morphospecies; Table S2) and the pan trap (372 morphospecies; Table S3). Hymenoptera had the highest diversity, with 229 species recorded. In terms of abundance, the majority of macroinvertebrates collected were Hemiptera (16,660 individuals; 107 species), followed by Diptera (10,684; 119 species).
For the vegetation survey, thirty-nine species of plants were recorded; 19 of them were native and 20 were exotic. All plots were mainly dominated by four species of plants (Table S4), which are Bidens alba (exotic), Brachiaria mutica (exotic), Callipteris esculenta (native) and Wedelia trilobata (exotic).
3.2. The Effects of Mowing Regimes on Birds
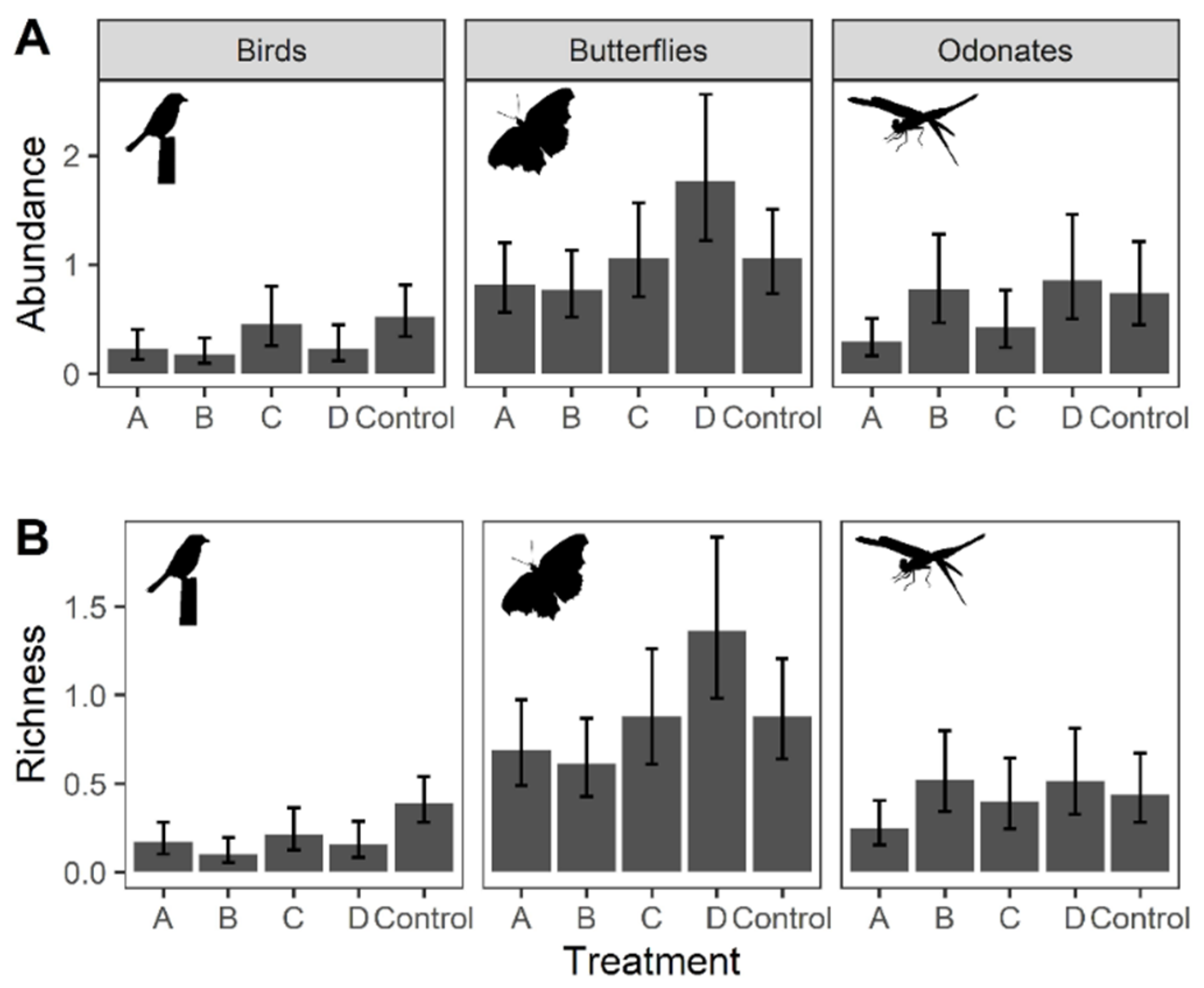
A total of 154 individuals of birds were recorded from the point and transect count surveys. The monthly mean abundance for each treatment (including the control) ranged from 0 to 3, while that of the mean richness ranged from 0 to 1. The results of the GLMMs suggested that abundance (Chisq = 13.151, p = 0.011) and richness of birds (Chisq = 20.865, p < 0.001) were significantly affected by treatment with Control having significantly higher bird richness than frequently mown plots (GLMMs: A–Control: t.ratio = −0.842, p = 0.043; B–Control: t.ratio = −1.353, p = 0.002) (Figure 2 and Table 3). In terms of abundance, Control was only significantly higher than that of Treatment B (B–Control: t.ratio = −1.097, p = 0.031). While season itself was significantly affecting bird species richness (Chisq = 13.094, p = 0.001), the effect of treatment was consistent across the seasons as the interaction term between season and treatment was not significant.
3.3. The Effects of Mowing Regimes on Butterflies
A total of 616 individual butterflies were recorded from the point and transect count surveys. The monthly mean abundance for each treatment (including the control) ranged from 0 to 7, while that of the mean richness ranged from 0 to 4.67. The results of the GLMMs suggested that abundance (Chisq = 27.033, p < 0.001) and species richness of butterflies (Chisq = 27.474, p < 0.001) were significantly affected by treatment with treatment D (i.e., low mowing intensity and frequency), having significant higher butterfly abundance than all other treatments and Control (GLMMs: A–D: t.ratio = −4.336, p < 0.001; B–D: t.ratio = −4.615, p < 0.001; C–D: t.ratio = −2.812, p = 0.041; Control–D: t.ratio = −3.187, p = 0.013) (Figure 2 and Table 3). Butterfly richness in Treatment D was also significantly higher than that in other treatments and Control except for Treatment C (GLMMs: A–D: t.ratio = −4.155, p < 0.001; B–D: t.ratio = −4.734, p < 0.001; Control–D: t.ratio = −2.993, p = 0.024). The effect of treatment was consistent across seasons.
3.4. The Effects of Mowing Regimes on Odonates
A total of 501 individual Odonates were recorded from the point and transect count surveys. The monthly mean abundance for each treatment (including the control) ranged from 0 to 7.1, while that of the mean richness ranged from 0 to 2.2. The results of the GLMMs suggested that the abundance of Odonates was significantly higher for Treatment B, D and Control than Treatment A (GLMMs: A–B: t.ratio = −4.237, p < 0.001; A–D: t.ratio = −4.296, p < 0.001; A–Control: t.ratio = −4012, p < 0.001. Table 3). Species richness of Treatment B and D were significantly higher than Treatment A (A–B: t.ratio = −3.693, p = 0.002; A–D: t.ratio = −3.291, p = 0.010), but were not significantly different from those of Control and Treatment C (Figure 2 and Table 3). No interaction was detected between the effects of treatment and season.
3.5. The Effects of Mowing Regimes on Amphibians and Reptiles
A total of 16 individual amphibians and reptiles were recorded from only eight plots (out of the 15 treatment and control plots) during the night surveys between June and September 2020. The monthly mean abundance of amphibians and reptiles (mean across the three zones) for each treatment (including the control) ranged from 0 to 1.33, while that of the mean richness ranged from 0 to 1. Given that amphibians and reptiles were not found at almost half of the plots, the data were insufficient for any meaningful statistical analysis. Thus, statistical analyses were not conducted for amphibians and reptiles.
3.6. The Effect of Mowing Regimes on Other Terrestrial Macroinvertebrates
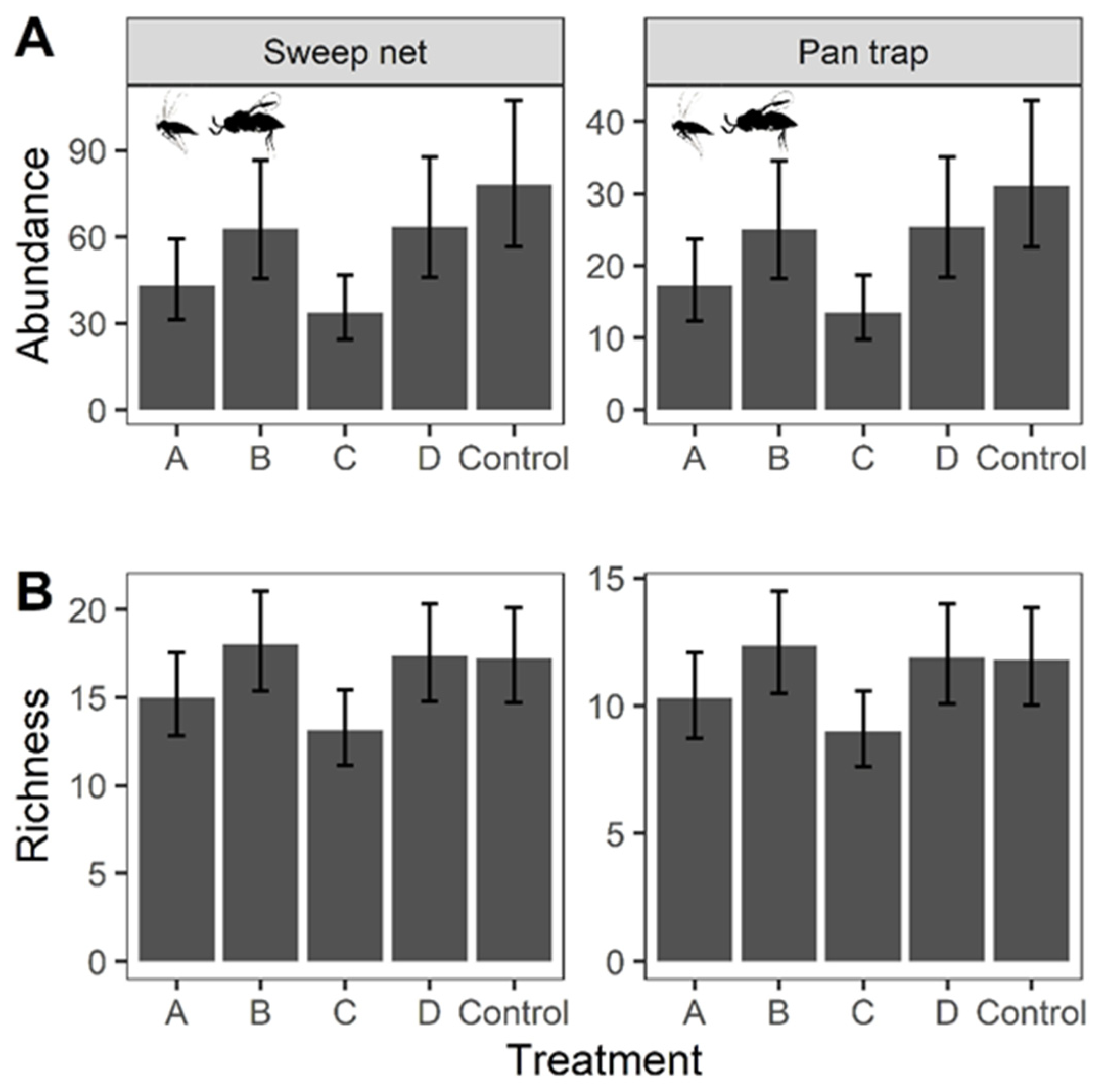
A total of 29,091 individual terrestrial macroinvertebrates were collected from the sweep net samples, representing 509 morphospecies in 155 families and 19 orders. Species abundance per sample ranged from 2 to 972 individuals with an average of 82.2, while species richness per sample ranged from 1 to 56 species, with an average of 18.2. For pan traps, 4852 individuals macroinvertebrates were collected, and 372 morphospecies in 136 families and 17 orders were identified. The species abundance per trap ranged from 0 to 275, with an average of 8.11 individuals, while the species richness recorded per trap ranged from 0 to 33 species, with an average of 3.98.
The results of the GLMMs suggested that Control plots had the highest macroinvertebrate abundance (GLMMs: A–Control: t.ratio = −20.573, p < 0.001; B–Control: t.ratio = −14.410, p < 0.001; C–Control: t.ratio = −42.208, p < 0.001; Control–D: t.ratio = 12.794, p < 0.001), followed by Treatment B and D (i.e., low mowing intensity), and then Treatment A followed by C (i.e., high mowing intensity; Figure 3 and Table 3). In terms of species richness, Treatment B, D and Control harbored significantly higher diversity of macroinvertebrates than Treatment A and lastly C.
3.7. The Effect of Mowing Regimes on Vegetation
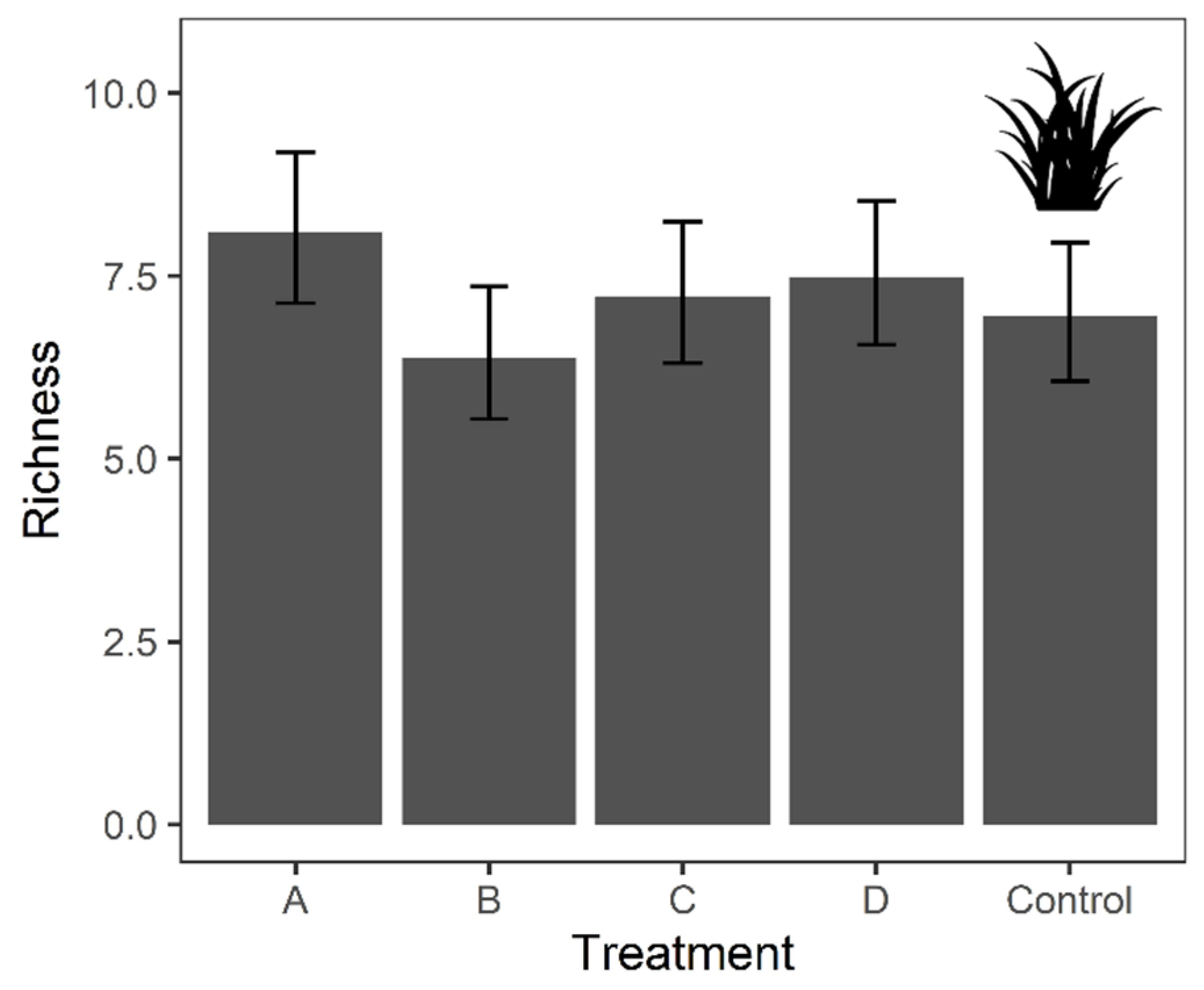
The overall species richness (i.e., cumulative species list) was higher for Treatment A (26 species), Treatment C (24 species) and Control (24 species) than Treatment B (18 species) and D (19 species). However, the results of the GLMMs suggested that flora richness was not significantly affected by treatment and season (Figure 4 and Table 3). Nevertheless, in terms of vegetation height, the height of the dominating and invasive tall grass Brachiaria mutica was suppressed in any mown plots (Table S5).
4. Discussion
4.1. Overall Response of Local Biodiversity to Mowing Regimes
Overall, the response of local biodiversity to different mowing regimes was taxon-specific. None of the treatment was favourable to all studied taxa. However, all taxa were either not affected (butterflies) or suppressed (birds, Odonates and other macroinvertebrates) by high intensity mowing in Treatment A and C. It was clear that high intensity mowing at the ground level, which is the current management strategy in Hong Kong, was not a favourable management practice from a biodiversity point of view. We acknowledge the potential effects of the close-proximity of the experimental plots and the relatively small size of plots. Despite these confounding effects that are expected to minimize the effect of treatments, significantly different responses from different taxa were still detected. The response of each taxon to various mowing regimes are discussed as follows.
4.2. Birds’ Preference for Infrequent Mowing Regime
Bird diversity was negatively impacted by frequent mowing, regardless of the mowing intensity. This suggested that excessive human disturbance from frequent mowing may deter birds from utilizing resources from the riverbed vegetation. As in many other studies, bird species richness has been shown to be negatively affected by human disturbance, especially for migrant species [11,12]. Infrequent mowing regimes shall therefore be implemented to retain bird diversity along the channelized rivers.
In addition, frequent mowing was particularly detrimental to birds nesting among tall grass. For instance, we have seen Prinia flaviventris carrying nest-building materials into the long grass, implying their use of the long grass along the engineered channel as a breeding site. Frequent mowing not only directly destroyed the nests and offspring, but also prevented these grass-dependent birds from accessing the breeding habitat [13]. Thus, retention of some uncut patches may provide breeding grounds for these grass-dependent small passerines.
4.3. Odonates’ Preference for Low Intensity Mowing Regime
Regardless of the mowing frequency, Odonate diversity was higher in low mowing intensity and unmown plots. This can be explained by the need for shelter and perching sites. As shown in Harabiš [14], when the vegetation was mown intensively (i.e., to ground level in Treatments A and C), Odonates were deprived from shelter and vegetative hunting platforms, and thus Odonates showed a lower preference for bare plots.
4.4. Enhancement of Butterfly Diversity by Mowing Regime with Low Mowing Intensity and Frequency
Plots with low mowing intensity and low mowing frequency harbored the highest butterfly diversity. This elevated diversity includes a number of species with conservation concerns. Seven out of the nine butterfly species with conservation concerns recorded in this study were recorded in plots of Treatment D (Table S1). The rare Udaspes folus, for example, was only recorded in a plot of Treatment D. In addition, most of the records (3/5) of the uncommon Hypolimnas misippus with Local Concern were also taken from plots of Treatment D. These results are similar to the findings of Uchida and Ushimaru [15], who found the highest butterfly diversity at intermediate mowing frequency (mown every six to four months). The higher butterfly diversity might be due to the larger supply of nectary source provided by the increased flower and host plant densities after mowing. As more time was available for growth and regeneration from the previous mow, flowers are more likely to reach maturity and thus more nectar is available [15]. We observed more flowers, particularly for the exotic weed Bidens alba, in plots with low mowing frequency and intensity, thus attracting more butterflies. Our study is the first to show that retention of a certain vegetation height may further enhance butterfly diversity in addition to low mowing frequency.
Meanwhile, the domination by tall and dense invasive Brachiaria mutica in unmown plots has deprived flowering plants of access to sunlight and growing space, and has caused a reduction in detection chance by floral visitors including butterflies. Therefore, lower butterfly diversity was recorded from unmown plots than in plots with low mowing intensity and frequency.
4.5. Low Utility of Grasscrete Vegetation by Amphibians and Reptiles
Due to the low number of observations for amphibians and reptiles across all plots (only 19 individuals from eight plots), no conclusion can be drawn on the effect of mowing regimes on these taxa. This low number of observations suggests that amphibians and reptiles living along the engineered channels do not prefer to use the vegetation grown on rip-rap lining and grasscretes. Instead, they were more often observed staying amongst submerged vegetation along the edge of the channelized river (for frogs), amongst crevices of exposed boulders (for geckos and skinks) and around mud pools.
4.6. Macroinvertebrates’ Preference for Low Intensity Mowing Regime and Unmown Areas
Higher macroinvertebrate species richness was recorded in plots with low intensity of mowing, as well as in unmown plots. When vegetation was mown to ground level, most macroinvertebrates living on grasscretes lost the vegetation that served as shelter, food and breeding sites [16]. Consequently, a significantly lower macroinvertebrate diversity was found in plots with high intensity of mowing. In contrast, the retention of a layer of vegetation under low mowing intensity regimes provides refuge for various macroinvertebrates to survive [17], and thus boosts macroinvertebrate diversity.
When comparing low mowing intensity plots with unmown plots, the macroinvertebrates diversity did not differ. Yet, the abundance of macroinvertebrates in unmown plots was significantly higher. This may be due to the higher vegetation biomass and surface area available as food, shelter and breeding ground [16].
4.7. Effect of Mowing Regimes on Vegetation
The overall effect of treatment on vegetation diversity, if taking only the species richness into account, was inconclusive because of the great variation in vegetation composition within and across treatments. The difference in vegetation, especially for the dominating species, was likely to be caused by the vegetation history specific to each plot. Once the dominating species have settled in each plot before the start of the study, the dominating plant species within each plot remained competitive across the ten-month study period. Noticeably, the dominating species differed across zones. For instance, plots in Zone 2 (except for the Control plot) have been particularly dominated by Wedelia trilobata throughout the study regardless of the mowing treatment, resulting in an average coverage of 23.83% across the Zone 2 plots, whereas those in Zone 1 and 3 were 5.00% and 2.30%, respectively. The huge variation in vegetation composition across plots has masked the effect of mowing regimes on vegetation diversity, leading to inconclusive results from our study. It is also worth noticing that none of the predominant species used during hydroseeding, which are Cynodon dactylon, Paspalum notatum and Lolium perenne, respectively [6], has been established in the plots throughout the experimental period (Table S5). Thus, hydroseeding performed before the start of experiment is believed to have no effect on the vegetation community composition in our study.
Nevertheless, it was revealed that total removal of vegetation is not favourable for diversity of butterflies, Odonates and macroinvertebrates because these fauna groups require vegetation for food source, shelter, perching sites and breeding grounds. Having said that, it was observed that mowing would favour some flowering groundcover by inhibiting domination by Brachiaria mutica, hence attracting butterflies and some macroinvertebrates that rely on flowering plants for food.
4.8. Recommended Mowing Regime
Based on the discussions above, with regards to the higher diversity of Odonates, butterflies and macroinvertebrates in plots with low mowing intensity, it is suggested to retain a certain vegetation height during mowing. In addition, since butterfly diversity was highly enhanced by the lower mowing frequency through promotion of flowering groundcover, it is recommended to mow the vegetation infrequently.
Although all taxa except for amphibians and reptiles are favored by the mowing at low frequency and intensity, we propose a mosaic mowing regime to cater for the needs of grass-dependent taxa such as some bush-nesting passerines. Apart from using low intensity and low frequency mowing to manage most parts of the vegetation along the engineered channels, patches of unmown riverbed vegetation could be maintained to provide suitable habitats and breeding sites for tall grass specialist birds and macroinvertebrates. Retention of patches of unmown grass within mown areas has been shown to benefit macroinvertebrate diversity by increasing habitat heterogeneity and providing refuge for grass-dependent specialists [17,18,19]. Thus, patches of unmown area could be preserved within the mown vegetation.
The mowing regimes at different areas of the riverbed could be decided depending on the land use beyond the riverbanks. Areas nearer to orchards, farmland or natural vegetation may be more suitable for unmown or low intensity/frequency mowing as wildlife could move between the riparian and the riverbed vegetation. Areas nearer to houses may be mowed more frequently with higher intensity, as complaints often come from residents along the riverbanks.
4.9. Limitations
The lack of treatment representing further lower mowing frequency is one of the biggest limitations of our study. The ‘low mowing frequency’ defined in our study was considered relatively frequent in most other research (e.g., [15,20]). A few studies from the temperate regions have revealed an optimal mowing frequency of once to twice a year [3,20]. However, whether the optimal mowing frequency used in the temperate region is applicable in subtropical areas such as Hong Kong remains unknown. Future research is therefore needed to test the effect of mowing regimes with lower mowing frequency on local biodiversity.
In addition, with regards to the close-proximity between plots and the relatively small size of plots in this study, future research should increase the plot size and separate the experimental plots further from each other. A longer monitoring period is recommended for capturing essential yearly fluctuation in biodiversity in vegetation along the channelized riverbed.
4.10. Other Recommendations for Enhancing Biodiversity along Engineered Channels
Other than the mosaic mowing regime suggested above, a few recommendations for further boosting biodiversity along engineered channels are listed as follows.
Adding poles or dead branches amongst or next to the grasscrete vegetation may attract predatory birds to utilize the resources within the vegetation. Lanius schach and Saxicola stejnegeri, which made up 6.5% and 41.8% of the total records of birds visiting the experimental plots, respectively (Table S1), were observed perching on the bamboo poles for marking the boundaries of each plot before they dived into the riverbed vegetation in search for food. Inserting a few poles or any similar structure along the engineered channels can therefore attract more birds.
The spread of invasive weeds on the water surfaces such as Eichhornia crassipes has substantially reduced the utility of nearby vegetation for Odonates. It is strongly recommended to remove surface-covering invasive plants regularly to maintain the open hunting ground above the river surface for Odonates.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su14074002/s1, Table S1: Mowing and survey dates; Table S2: Species list of fauna surveys; Tables S3 and S4: Species list of sweep net and pan trap; Table S5: Species list of vegetation survey; Table S6: Changes in vegetation height of the six dominating species in different seasons and treatment plots.
Author Contributions
B.C.H.H., K.Y.K.S. and B.T.W.S. contributed to the conception and design of the study. T.K.C.L., K.Y.K.S. and B.T.W.S. collected field data for fauna surveys (except for night surveys). In addition, B.T.W.S. conducted vegetation and night surveys. T.K.C.L. was responsible for sorting and identification of macroinvertebrates collected by pan traps and sweep net. K.Y.K.S. and T.K.C.L. performed data analysis for the study. The first draft of the manuscript was written by T.K.C.L. and all authors provided comments for the revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Drainage Services Department of Hong Kong (Agreement No. MN 01/2019).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The survey dataset is available at https://doi.org/10.5281/zenodo.6388990 (accessed on 6 September 2021).
Acknowledgments
This project was funded by the Drainage Services Department of the Hong Kong SAR Government (Agreement No. MN 01/2019) and we thank the Director of Drainage Services for endorsing this manuscript. We sincerely thank Hoi Yan Mak and Maria Lo for the preparation of field equipment. We also thank Chi Chiu Pang for providing valuable advice on the design of the experiment. We also acknowledge the crucial comments from Chui Yu Cheung for refining the manuscript on this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ellis, J.B.; Scholes, L.; Revitt, D.M.; Oldham, J. Sustainable urban development and drainage. Proc. Inst. Civ. Eng. Munic Eng. 2004, 157, 245–250. [Google Scholar] [CrossRef]
- Yu, C. Sustainable Urban Drainable Systems for Management of Surface Water. In Design and Management of Sustainable Built Environments; Yao, R., Ed.; Springer: London, UK, 2013; pp. 119–140. [Google Scholar]
- Valkó, O.; Török, P.; Matus, G.; Tóthmérész, B. Is regular mowing the most appropriate and cost-effective management maintaining diversity and biomass of target forbs in mountain hay meadows? Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 303–309. [Google Scholar] [CrossRef]
- Mazalová, M.; Šipoš, J.; Rada, S.; Kašák, J.; Šarapatka, B.; Kuras, T. Responses of grassland arthropods to various biodiversity-friendly management practices: Is there a compromise? Eur. J. Entomol. 2015, 112, 734–746. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, S.; Bernes, C.; Bullock, J.M.; Verheyen, K.; Lindborg, R. How does roadside vegetation management affect the diversity of vascular plants and invertebrates? A systematic review. Environ. Evid. 2018, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Civil Engineering and Development Department. General Specifications for Engineering Works (GS), 2020 ed.; Civil Engineering and Development Department: Hong Kong, China, 2020; Volume 1, pp. 121–122.
- Leksono, A.S.; Takada, K.; Koji, S.; Nakagoshi, N.; Anggraeni, T.; Nakamura, K. Vertical and seasonal distribution of flying beetles in a suburban temperate deciduous forest collected by water pan trap. Insect Sci. 2005, 12, 199–206. [Google Scholar] [CrossRef]
- Saunders, M.E.; Luck, G.W. Pan trap catches of pollinator insects vary with habitat. Aust. J. Entomol. 2013, 52, 106–113. [Google Scholar] [CrossRef]
- Fellowes, J.R.; Lau, M.W.; Dudgeon, D.; Reels, G.T.; Ades, G.W.; Carey, G.J.; Chan, B.P.L.; Kendrick, R.C.; Lee, K.S.; Leven, M.R.; et al. Wild animals to watch: Terrestrial and freshwater fauna of conservation concern in Hong Kong. Mem. Hong Kong Nat. Hist. Soc. 2002, 25, 123–159. [Google Scholar]
- Agriculture, Fisheries and Conservation Department. Hong Kong Biodiversity Database. 2020. Available online: https://www.afcd.gov.hk/english/conservation/hkbiodiversity/database/search.php (accessed on 6 September 2021).
- Zhou, D.; Chu, L.M. How would size, age, human disturbance, and vegetation structure affect bird communities of urban parks in different seasons? J. Ornithol. 2012, 153, 1101–1112. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.R.; Lee, D. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Frawley, B.J.; Best, L.B. Effects of mowing on breeding bird abundance and species composition in alfalfa fields. Wildl Soc. Bull. 1991, 19, 135–142. [Google Scholar]
- Harabiš, F. Does the management of surrounding terrestrial habitats increase the tendency of odonates to leave aquatic habitats? Biodivers. Conserv. 2017, 26, 2155–2167. [Google Scholar] [CrossRef]
- Uchida, K.; Ushimaru, A. Biodiversity declines due to abandonment and intensification of agricultural lands: Patterns and mechanisms. Ecol. Monogr. 2014, 84, 637–658. [Google Scholar] [CrossRef]
- Cizek, O.; Zamecnik, J.; Tropek, R.; Kocarek, P.; Konvicka, M. Diversification of mowing regime increases arthropods diversity in species-poor cultural hay meadows. J. Insect Conserv. 2012, 16, 215–226. [Google Scholar] [CrossRef]
- Buri, P.; Arlettaz, R.; Humbert, J.Y. Delaying mowing and leaving uncut refuges boosts orthopterans in extensively managed meadows: Evidence drawn from field-scale experimentation. Agric. Ecosyst. Environ. 2013, 181, 22–30. [Google Scholar] [CrossRef]
- Humbert, J.Y.; Ghazoul, J.; Walter, T. Meadow harvesting techniques and their impacts on field fauna. Agric. Ecosyst. Environ. 2009, 130, 1–8. [Google Scholar] [CrossRef]
- Köhler, B.; Gigon, A.; Edwards, P.J.; Krüsi, B.; Langenauer, R.; Lüscher, A.; Ryser, P. Changes in the species composition and conservation value of limestone grasslands in Northern Switzerland after 22 years of contrasting managements. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 51–67. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Römermann, C.; Sperlich, S.; Schmidt, W. Explaining grassland biomass–the contribution of climate, species and functional diversity depends on fertilization and mowing frequency. J. Appl. Ecol. 2011, 48, 1088–1097. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
(A) Location of the study site. (B) Overview of the study site. (C) Layout of the experimental plots in the study site. A minimum of 1 m of cut-down-to-ground strip will be maintained between each plot as a buffer.
Figure 1.
(A) Location of the study site. (B) Overview of the study site. (C) Layout of the experimental plots in the study site. A minimum of 1 m of cut-down-to-ground strip will be maintained between each plot as a buffer.

Figure 2.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on (A) abundance and (B) richness of birds, butterflies and Odonates across treatments. All models used abundance or richness as dependent variables, with ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate, and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.
Figure 2.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on (A) abundance and (B) richness of birds, butterflies and Odonates across treatments. All models used abundance or richness as dependent variables, with ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate, and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.

Figure 3.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on abundance and richness of terrestrial macroinvertebrates across Treatments (A,B). The models used abundance or richness as dependent variables, with ‘collection methods’, ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.
Figure 3.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on abundance and richness of terrestrial macroinvertebrates across Treatments (A,B). The models used abundance or richness as dependent variables, with ‘collection methods’, ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.

Figure 4.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on the species richness of vegetation across treatments. Species richness was as dependent variables, with ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.
Figure 4.
Predicted value from the Generalized Linear Mixed Models (GLMMs) on the species richness of vegetation across treatments. Species richness was as dependent variables, with ‘Treatment’, ‘Season’ and their interaction term set as independent variables. ‘Zone’ was incorporated into the model as covariate and ‘Month’ was used as the random factor. Error bar indicates the 95% confident intervals.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The four experimental treatments with different mowing regimes. Each with three replicates (i.e., three zones).
Table 1.
The four experimental treatments with different mowing regimes. Each with three replicates (i.e., three zones).
| High Mowing Intensity | Low Mowing Intensity | |
|---|---|---|
| High mowing frequency | Treatment A Mowed once every month (March to October) or once every two months (November to February) to the ground level. | Treatment B Mowed once every month (March to October) or once every two months (November to February) to 600 mm vegetation height. |
| Low mowing frequency | Treatment C Mowed once every three months throughout the study period to the ground level. | Treatment D Mowed once every three months throughout the study period to 600 mm vegetation height. |
Table 2.
Target taxa of each surveying method employed for monthly surveys during the study period.
| Methods/Target Taxa | Birds | Amphibians | Reptiles | Butterflies | Odonates | Other Terrestrial Macroinvertebrates | Plants |
|---|---|---|---|---|---|---|---|
| Point count * | ✓ | ✓ | ✓ | ||||
| Transect count * | ✓ | ✓ | ✓ | ||||
| Sweep net * | ✓ | ✓ | ✓ | ||||
| Pan trap | ✓ | ||||||
| Night survey #,* | ✓ | ✓ | |||||
| Vegetation survey & | ✓ |
* Apart from monthly surveys, two additional surveys were conducted for plots that had been mowed in the respective months. # Only conducted in wet season (June–September). & A vegetation survey was conducted at the early beginning of this project prior to any mowing, during which a complete inventory of plant species was recorded for the whole study site.
Table 3.
Summary of the results of the Generalized Linear Mixed Models (GLMMs) and the post-hoc test calculated using least-square means. Post-hoc test for a particular taxon was done only if significant effect of ‘Treatment’ was detected in the respective GLMMs. The analysis for ‘Other macroinvertebrates’ was done by combining the data collected from the sweep net and the pan traps, and the effect of the capturing method was represented in the model as ‘Method’. ‘*’ indicates 0.05 > p ≥ 0.01, ‘**’ represents 0.01 > p ≥ 0.001, and ‘***’ indicates p > 0.001.
Table 3.
Summary of the results of the Generalized Linear Mixed Models (GLMMs) and the post-hoc test calculated using least-square means. Post-hoc test for a particular taxon was done only if significant effect of ‘Treatment’ was detected in the respective GLMMs. The analysis for ‘Other macroinvertebrates’ was done by combining the data collected from the sweep net and the pan traps, and the effect of the capturing method was represented in the model as ‘Method’. ‘*’ indicates 0.05 > p ≥ 0.01, ‘**’ represents 0.01 > p ≥ 0.001, and ‘***’ indicates p > 0.001.
| GLMMs | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abundance | Species Richness | |||||||||||||
| Taxa | Chisq | df | p-value | Chisq | df | p-value | ||||||||
| Birds | Treatment | 13.151 | 4 | 0.011 | * | Treatment | 20.865 | 4 | <0.001 | *** | ||||
| Season | 3.379 | 2 | 0.157 | Season | 13.094 | 2 | 0.001 | ** | ||||||
| Butterflies | Treatment | 27.033 | 4 | <0.001 | *** | Treatment | 27.474 | 4 | <0.001 | *** | ||||
| Season | 18.958 | 2 | <0.001 | *** | Season | 17.356 | 2 | <0.001 | *** | |||||
| Odonates | Treatment | 27.561 | 4 | <0.001 | *** | Treatment | 15.645 | 4 | 0.003 | ** | ||||
| Season | 33.220 | 2 | <0.001 | *** | Season | 28.059 | 2 | <0.001 | *** | |||||
| Other macro-invertebrates | Treatment | 2487.390 | 4 | <0.001 | *** | Treatment | 94.571 | 4 | <0.001 | *** | ||||
| Season | 15.826 | 2 | <0.001 | *** | Season | 28.819 | 2 | <0.001 | *** | |||||
| Method | 3478.240 | 1 | <0.001 | *** | Method | 204.763 | 1 | <0.001 | *** | |||||
| Vegetation | Treatment | 6.617 | 4 | 0.158 | Treatment | / | ||||||||
| Season | 5.284 | 2 | 0.071 | Season | ||||||||||
| Post-hoc test: Least square means | ||||||||||||||
| Estimate | SE | df | t.ratio | p-value | Estimate | SE | df | t.ratio | p-value | |||||
| Birds | A–B | 0.272 | 0.422 | 358 | 0.647 | 0.967 | A–B | 0.511 | 0.422 | 359 | 1.212 | 0.745 | ||
| A–C | −0.686 | 0.403 | 358 | −1.699 | 0.436 | A–C | −0.239 | 0.372 | 359 | −0.643 | 0.968 | |||
| A–D | <0.001 | 0.439 | 358 | <0.001 | 1.000 | A–D | 0.097 | 0.409 | 359 | 0.237 | 0.999 | |||
| A–Control | −0.825 | 0.352 | 358 | −2.341 | 0.135 | A–Control | −0.842 | 0.301 | 359 | −2.796 | 0.043 | * | ||
| B–C | −0.958 | 0.424 | 358 | −2.259 | 0.161 | B–C | −0.750 | 0.428 | 359 | −1.753 | 0.403 | |||
| B–D | −0.272 | 0.459 | 358 | −0.593 | 0.976 | B–D | −0.413 | 0.460 | 359 | −0.899 | 0.897 | |||
| B–Control | −1.097 | 0.377 | 358 | −2.912 | 0.031 | * | B–Control | −1.353 | 0.368 | 359 | −3.680 | 0.002 | ** | |
| C–D | 0.685 | 0.433 | 358 | 1.584 | 0.509 | C–D | 0.336 | 0.414 | 359 | 0.813 | 0.927 | |||
| C–Control | −0.139 | 0.352 | 358 | −0.395 | 0.995 | C–Control | −0.603 | 0.308 | 359 | −1.959 | 0.288 | |||
| Control–D | 0.825 | 0.391 | 358 | 2.108 | 0.219 | Control–D | 0.940 | 0.351 | 359 | 2.676 | 0.060 | |||
| Butterflies | A–B | 0.064 | 0.181 | 358 | 0.352 | 0.997 | A–B | 0.121 | 0.174 | 359 | 0.696 | 0.957 | ||
| A–C | −0.250 | 0.191 | 358 | −1.310 | 0.685 | A–C | −0.243 | 0.183 | 359 | −1.328 | 0.674 | |||
| A–D | −0.768 | 0.177 | 358 | −4.336 | <0.001 | *** | A–D | −0.683 | 0.164 | 359 | −4.155 | <0.001 | *** | |
| A–Control | −0.253 | 0.165 | 358 | −1.532 | 0.542 | A–Control | −0.244 | 0.155 | 359 | −1.576 | 0.514 | |||
| B–C | −0.314 | 0.194 | 358 | −1.618 | 0.489 | B–C | −0.364 | 0.188 | 359 | −1.939 | 0.299 | |||
| B–D | −0.832 | 0.180 | 358 | −4.615 | <0.001 | *** | B–D | −0.805 | 0.170 | 359 | −4.734 | <0.001 | *** | |
| B–Control | −0.317 | 0.168 | 358 | −1.885 | 0.327 | B–Control | −0.365 | 0.161 | 359 | −2.274 | 0.156 | |||
| C–D | −0.518 | 0.184 | 358 | −2.812 | 0.041 | * | C–D | −0.441 | 0.171 | 359 | −2.572 | 0.078 | ||
| C–Control | −0.003 | 0.177 | 358 | −0.015 | 1.000 | C–Control | −0.001 | 0.167 | 359 | −0.006 | 1.000 | |||
| Control–D | −0.515 | 0.162 | 358 | −3.187 | 0.013 | * | Control–D | −0.440 | 0.147 | 359 | −2.993 | 0.024 | * | |
| Odonates | A–B | −0.969 | 0.229 | 358 | −4.237 | <0.001 | *** | A–B | −0.747 | 0.202 | 359 | −3.693 | 0.002 | ** |
| A–C | −0.377 | 0.275 | 358 | −1.369 | 0.648 | A–C | −0.473 | 0.236 | 359 | −2.001 | 0.268 | |||
| A–D | −1.073 | 0.250 | 358 | −4.296 | <0.001 | *** | A–D | −0.733 | 0.223 | 359 | −3.291 | 0.010 | * | |
| A–Control | −0.923 | 0.230 | 358 | −4.012 | <0.001 | *** | A–Control | −0.566 | 0.208 | 359 | −2.714 | 0.054 | ||
| B–C | 0.592 | 0.249 | 358 | 2.375 | 0.125 | B–C | 0.275 | 0.203 | 359 | 1.354 | 0.658 | |||
| B–D | −0.104 | 0.215 | 358 | −0.482 | 0.989 | B–D | 0.014 | 0.187 | 359 | 0.077 | 1.000 | |||
| B–Control | 0.046 | 0.195 | 358 | 0.237 | 0.999 | B–Control | 0.182 | 0.170 | 359 | 1.070 | 0.822 | |||
| C–D | −0.696 | 0.263 | 358 | −2.644 | 0.065 | C–D | −0.260 | 0.219 | 359 | −1.190 | 0.757 | |||
| C–Control | −0.546 | 0.247 | 358 | −2.209 | 0.179 | C–Control | −0.093 | 0.209 | 359 | −0.445 | 0.992 | |||
| Control–D | −0.150 | 0.217 | 358 | −0.693 | 0.958 | Control–D | −0.167 | 0.193 | 359 | −0.865 | 0.910 | |||
| Other macro-invertebrates | A–B | −0.380 | 0.018 | 494 | −21.110 | <0.001 | *** | A–B | −0.183 | 0.034 | 494 | −5.465 | <0.001 | *** |
| A–C | 0.239 | 0.022 | 494 | 10.657 | <0.001 | *** | A–C | 0.135 | 0.039 | 494 | 3.422 | 0.006 | ** | |
| A–D | −0.392 | 0.019 | 494 | −35.337 | <0.001 | *** | A–D | −0.145 | 0.036 | 494 | −3.969 | <0.001 | *** | |
| A–Control | −0.595 | 0.017 | 494 | −20.573 | <0.001 | *** | A–Control | −0.137 | 0.033 | 494 | −4.139 | <0.001 | *** | |
| B–C | 0.619 | 0.021 | 494 | 29.463 | <0.001 | *** | B–C | 0.317 | 0.038 | 494 | 8.363 | <0.001 | *** | |
| B–D | −0.012 | 0.017 | 494 | −0.696 | 0.957 | B–D | 0.039 | 0.035 | 494 | 1.103 | 0.805 | |||
| B–Control | −0.215 | 0.015 | 494 | −14.410 | <0.001 | *** | B–Control | 0.047 | 0.031 | 494 | 1.485 | 0.573 | ||
| C–D | −0.631 | 0.021 | 494 | −29.614 | <0.001 | *** | C–D | −0.279 | 0.040 | 494 | −7.023 | <0.001 | *** | |
| C–Control | −0.834 | 0.020 | 494 | −42.208 | <0.001 | *** | C–Control | −0.271 | 0.037 | 494 | −7.273 | <0.001 | *** | |
| Control–D | 0.203 | 0.016 | 494 | 12.794 | <0.001 | *** | Control–D | −0.008 | 0.034 | 494 | −0.235 | 0.999 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Leung, T.K.C.; So, K.Y.K.; Shum, B.T.W.; Hau, B.C.H. Optimal Mowing Regime in Enhancing Biodiversity in Seasonal Floodplains along Engineered Channels. Sustainability 2022, 14, 4002. https://doi.org/10.3390/su14074002
AMA Style
Leung TKC, So KYK, Shum BTW, Hau BCH. Optimal Mowing Regime in Enhancing Biodiversity in Seasonal Floodplains along Engineered Channels. Sustainability. 2022; 14(7):4002. https://doi.org/10.3390/su14074002
Chicago/Turabian StyleLeung, Tsz Kin Calvin, Ken Ying Kin So, Bond Ting Wing Shum, and Billy Chi Hang Hau. 2022. "Optimal Mowing Regime in Enhancing Biodiversity in Seasonal Floodplains along Engineered Channels" Sustainability 14, no. 7: 4002. https://doi.org/10.3390/su14074002
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.