
Effect of Dietary Inclusion of Alpha-Monolaurin on the Growth Performance, Lipid Peroxidation, and Immunity Response in Broilers

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Growth Performance and Digestibility
2.3. Biochemical Analysis
2.4. Fatty Acid Profile and Malondialdehyde
2.5. Infection Trial
2.6. Statistical Analysis
3. Results
3.1. Growth Efficiency, Digestibility, and Organ Weight
3.2. Blood Biochemical Traits
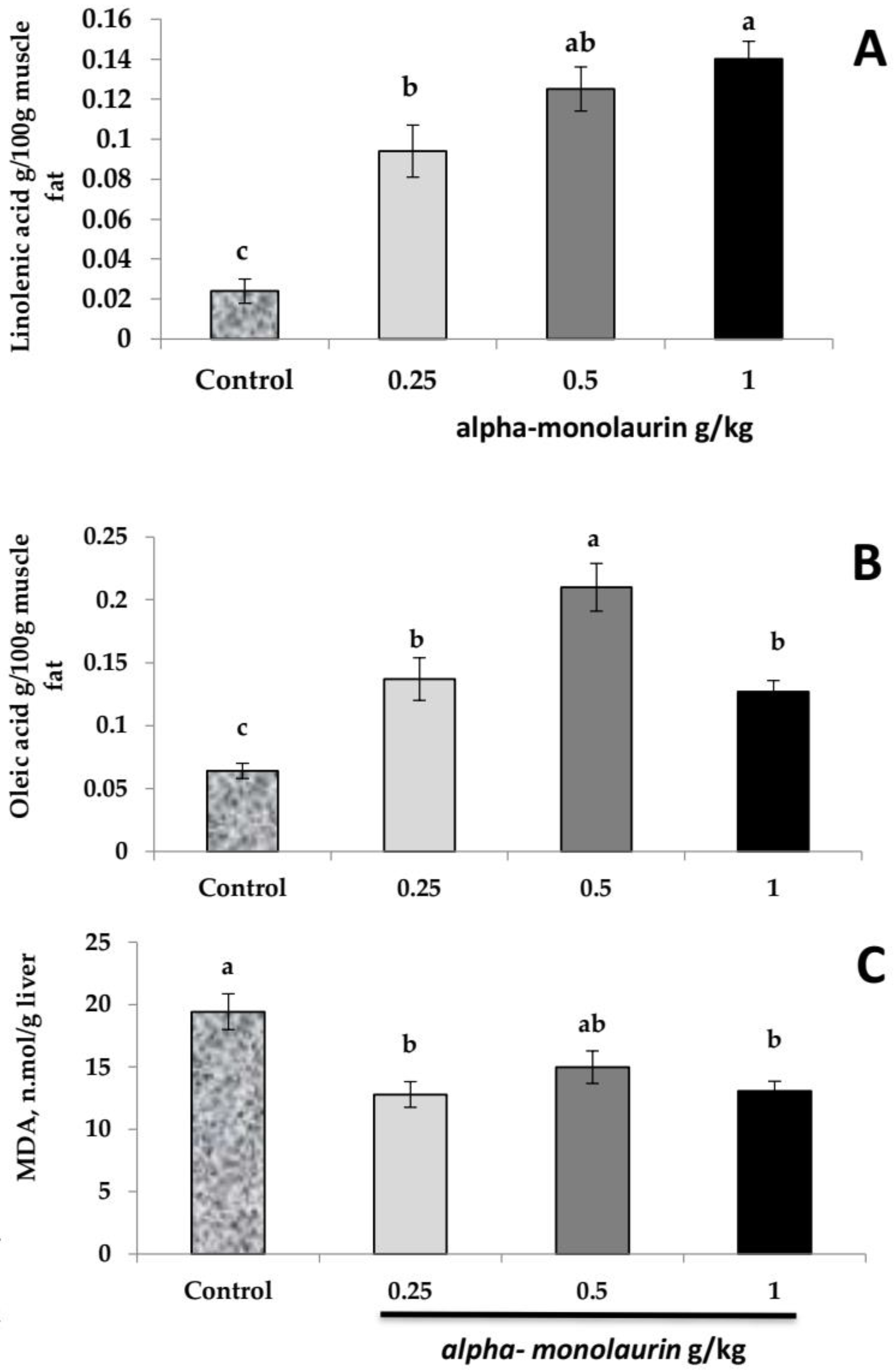
3.3. Fatty Acid Composition and Lipid Peroxidation
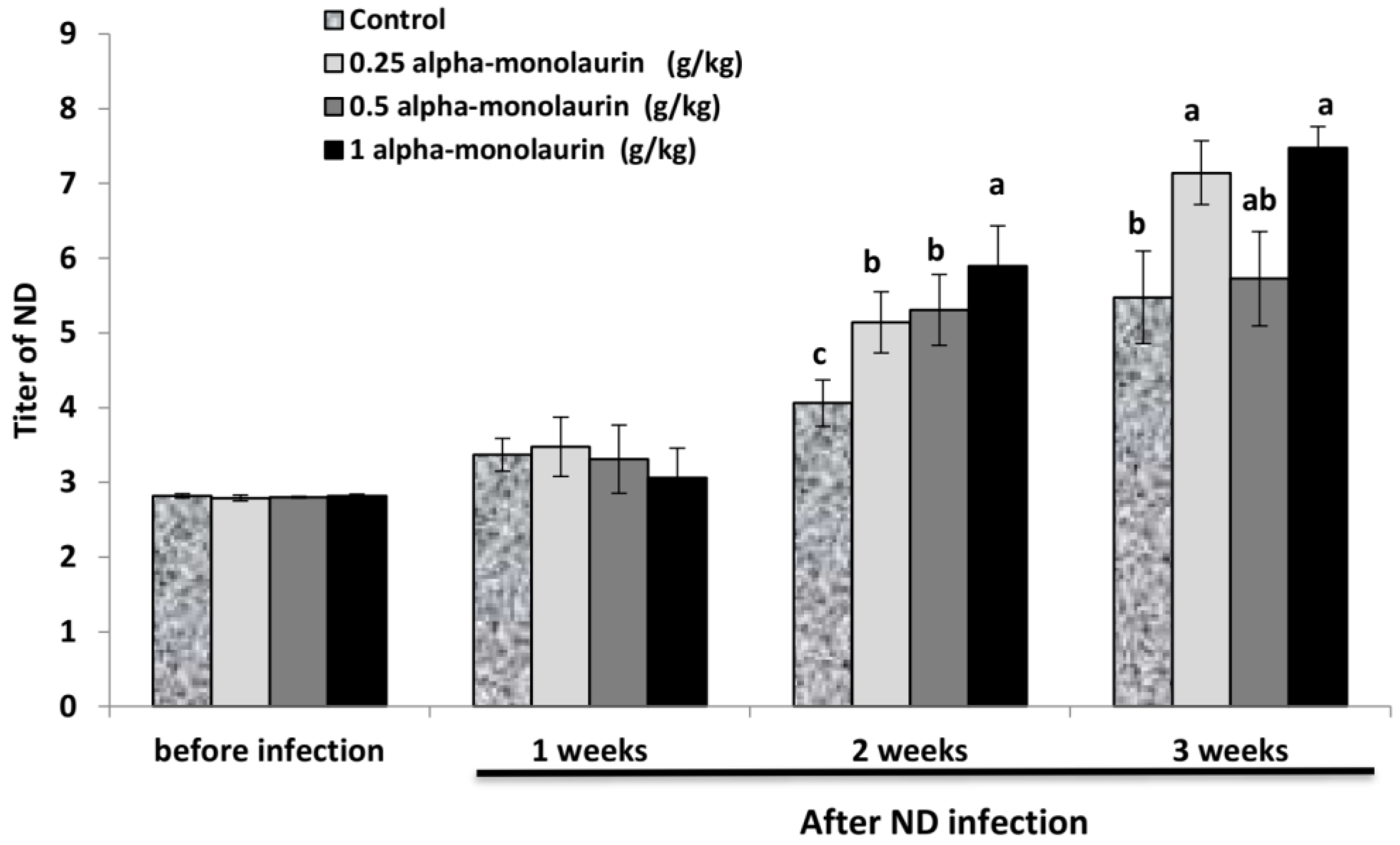
3.4. Resistance to ND
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira, L.S.; Balbino, E.M.; Silva, T.N.S.; Ily, L.; da Rocha, T.C.; de Oliveira Strada, E.S.; Pinheiro, A.M.; de Brito, J.A.G. Use of emulsifier and lipase in feeds for broiler chickens. Semin. Ciências Agrárias 2019, 40, 3181–3196. [Google Scholar] [CrossRef]
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. World Poult. 2015, 31, 16–17. [Google Scholar]
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, B.; Greko, C. Antibiotic resistance—Consequences for animal health, welfare, and food production. Upsala J. Med Sci. 2014, 119, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Elnagar, A.M.; Eid, Y.Z.; Ebeid, T.A.; Amber, K.A. Effect of feeding wheat middlings and calcium lignosulfonate as pellet binders on pellet quality growth performance and lipid peroxidation in broiler chickens. Vet. Med. Sci. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.C.; Kwakkel, R.P.; Soede, J.; Williams, B.A.; Verstegen, M.W.A. Effect of a chinese herb medicine formulation, as an alternative for antibiotics, on performance of broilers. Br. Poult. Sci. 2004, 45, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Paray, B.A.; Dawood, M.A.O. Olive cake meal and bacillus licheniformis impacted the growth performance, muscle fatty acid content, and health status of broiler chickens. Animals 2020, 10, 695. [Google Scholar] [CrossRef]
- El-Deep, M.H.; Amber, K.A.; Elgendy, S.; Dawood, M.A.O.; Zidan, A. In ovo injection of nano-selenium spheres mitigates the hatchability, histopathology image and immune response of hatched chicks. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1392–1400. [Google Scholar] [CrossRef]
- Fahmy, Z.H.; Aly, E.; Mohamed, A.H. The effect of medium chain saturated fatty acid (monolaurin) on levels of the cytokines on experimental animal in entamoeba histolytica and giardia lamblia infection. Afr. J. Pharm. Pharmacol. 2014, 8, 106–114. [Google Scholar]
- Zentek, J.; Buchheit-Renko, S.; Ferrara, F.; Vahjen, W.; Van Kessel, A.; Pieper, R. Nutritional and physiological role of medium-chain triglycerides and medium-chain fatty acids in piglets. Anim. Health Res. Rev. 2011, 12, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Ettinger, M. Antiviral and antibacterial actions of monolaurin and lauric acid. Mol. Cell Biochem. 2005, 272, 29–34. [Google Scholar]
- Londok, J.J.M.R.; Sumiati, S.; Wiryawan, I.K.G.; Manalu, W. Antioxidant enzyme activity and malondialdehyde concentration on broiler fed contain lauric acid and areca vestiaria giseke. Bul. Peternak. 2018, 42. [Google Scholar] [CrossRef]
- Lieberman, S.; Enig, M.G.; Preuss, H.G. A review of monolaurin and lauric acid: Natural virucidal and bactericidal agents. Altern. Complementary Ther. 2006, 12, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, N.G. Biochemical trails associated with different doses of alpha-monolaurin in chicks. Adv. Anim. Vet. Sci. 2019, 7, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, C.E.; Thormar, H.; Pessolano, T. Membrane-disruptive effect of human milk: Inactivation of enveloped viruses. J. Infect. Dis. 1986, 154, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Preuss, H.G.; Echard, B.; Enig, M.; Brook, I.; Elliott, T.B. Minimum inhibitory concentrations of herbal essential oils and monolaurin for gram-positive and gram-negative bacteria. Mol. Cell. Biochem. 2005, 272, 29–34. [Google Scholar] [CrossRef]
- Fortuoso, B.F.; Dos Reis, J.H.; Gebert, R.R.; Barreta, M.; Griss, L.G.; Casagrande, R.A.; de Cristo, T.G.; Santiani, F.; Campigotto, G.; Rampazzo, L.; et al. Glycerol monolaurate in the diet of broiler chickens replacing conventional antimicrobials: Impact on health, performance and meat quality. Microb. Pathog. 2019, 129, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Witcher, K.J.; Novick, R.P.; Schlievert, P.M. Modulation of Immune Cell Proliferation by Glycerol Monolaurate. Clin. Diagn. Lab. Immunol. 1996, 3, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.S.; Sandouk, A.; Houtman, J.C.D. Glycerol Monolaurate (GML) inhibits human T-cell signaling and function by disrupting lipid dynamics. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bartolotta, S.; Garc, C.C.; Candurra, N.A.; Damonte, E.B. Effect of fatty acids on arenavirus replication: Inhibition of virus production by lauric acid. Arch. Virol. 2001, 146, 777–790. [Google Scholar] [CrossRef]
- Arora, R.; Chawla, R.; Marwah, R.; Arora, P.; Sharma, R.K.; Kaushik, V.; Goel, R.; Kaur, A.; Silambarasan, M.; Tripathi, R.P.; et al. Potential of complementary and alternative medicine in preventive management of novel H1N1 flu (swine flu) pandemic: Thwarting potential disasters in the bud. Evidence-Based Complementary Altern. Med. 2011, 586506. [Google Scholar] [CrossRef] [PubMed]
- Hilmarsson, H.; Traustason, B.S.; Kristmundsdóttir, T.; Thormar, H. Virucidal activities of medium- and long-chain fatty alcohols and lipids against respiratory syncytial virus and parainfluenza virus type 2: Comparison at different pH levels. Arch. Virol. 2007, 152, 2225–2236. [Google Scholar] [CrossRef]
- Ruzin, A.; Novick, R.P. Equivalence of Lauric Acid and Glycerol Monolaurate as Inhibitors of Signal Transduction in Staphylococcus aureus. J. Bacteriol. 2000, 182, 2668–2671. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 19th ed.; Association of Official Agricultural Chemists: Arlington, VA, USA, 1994. [Google Scholar]
- Saleh, A.A. Effects of fish oil on the production performances, polysaturated fatty acids and cholesterol levels of yolk in hens. Emir. J. Food Agric. 2013, 25, 605–612. [Google Scholar] [CrossRef] [Green Version]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2008; pp. 1092–1106. [Google Scholar]
- Londok, J.; Manalu, W.; Wiryawan, K.G. Growth performance, carcass characteristics and fatty acids profile of broilers supplemented with lauric acid and natural antioxidant from areca vestiaria giseke. Pak. J. Nutr. 2017, 16, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.; Ahmed, E.; Ebeid, A. The impact of phytoestrogen source supplementation on reproductive performance, plasma profile, yolk fatty acids and antioxidative status in aged laying hens. Reprod. Domest. Anim. 2019, 54, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Zeitz, J.; Fennhoff, J.; Kluge, H.; Stangl, G.; Eder, K. Effects of dietary fats rich in lauric and myristic acid on performance, intestinal morphology, gut microbes, and meat quality in broilers. Poult. Sci. 2015, 94, 2404–2413. [Google Scholar] [CrossRef]
- Çenesiz, A.; Çiftci, İ. Modulatory effects of medium chain fatty acids in poultry nutrition and health. World’s Poult. Sci. J. 2020, 27, 1–15. [Google Scholar] [CrossRef]
- Khosravinia, H. Effect of dietary supplementation of medium-chain fatty acids on growth performance and prevalence of carcass defects in broiler chickens raised in different stocking densities. J. Appl. Poult. Res. 2015, 24, 1–9. [Google Scholar] [CrossRef]
- Mountzouris, K.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef]
- Vieira, S.L.; Oyarzabal, O.; Freitas, D.; Berres, J.; Pena, J.; Torres, C.; Coneglian, J. Performance of broilers fed diets supplemented with sanguinarine-like alkaloids and organic acids. J. Appl. Poult. Res. 2008, 17, 128–133. [Google Scholar] [CrossRef]
- Guillot, E.; Vaugelade, P.; Lemarchali, P.; Rat, A.R. Intestinal absorption and liver uptake of medium-chain fatty acids in non-anaesthetized pigs. Biritish J. Nutr. 1993, 69, 431–442. [Google Scholar] [CrossRef]
- Takase, S.; Goda, T. Effects of medium-chain triglycerides on brush border membrane-bound enzyme activity in rat small intestine. J. Nutr. 1990, 120, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, C.; Zhong, H.; Feng, F. Dietary medium-chain α-monoglycerides increase BW, feed intake, and carcass yield in broilers with muscle composition alteration. Poult. Sci. 2021, 100, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Livingston, M.L.; Cowieson, A.J.; Crespo, R.; Hoang, V.; Nogal, B.; Browning, M.; Livingston, K.A. Effect of broiler genetics, age, and gender on performance and blood chemistry. Heliyon 2020, 6, e04400. [Google Scholar] [CrossRef] [PubMed]
- Seleem, D.; Chen, E.; Benso, B.; Pardi, V.; Murata, R.M.J.P. In vitro evaluation of antifungal activity of monolaurin against candida albicans biofilms. PeerJ 2016, 4, e2148. [Google Scholar] [CrossRef] [Green Version]
- Baltić, B.; Ćirić, J.; Šefer, D.; Radovanović, A.; Đorđević, J.; Glišić, M.; Bošković, M.; Baltić, M.Ž.; Đorđević, V.; Marković, R. Effect of dietary supplementation with medium chain fatty acids on growth performance, intestinal histomorphology, lipid profile and intestinal microflora of broiler chickens. South Afr. J. Anim. Sci. 2018, 48, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Projan, S.J.; Brown-Skrobot, S.; Schlievert, P.M.; Vandenesch, F.; Novick, R.P. Glycerol monolaurate inhibits the production of beta-lactamase, toxic shock toxin-1, and other staphylococcal exoproteins by interfering with signal transduction. J. Bacteriol. 1994, 176, 4204–4209. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Y.; Liu, Y.; Wang, J.; Xu, Q.; Yu, X.; Yang, X.; Liu, Z.; Xue, C. Medium-chain triglycerides promote macrophage reverse cholesterol transport and improve atherosclerosis in ApoE-deficient mice fed a high-fat diet. Nutr. Res. 2016, 36, 964–973. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, Y.; Jacoby, J.r.J.; Jiang, Y.; Zhang, Y.; Yu, L.L. Effects of medium-and long-chain triacylglycerols on lipid metabolism and gut microbiota composition in C57BL/6J mice. J. Agric. Food Chem. 2017, 65, 6599–6607. [Google Scholar] [CrossRef]
- Yao, J.; Wang, J.-Y.; Liu, L.; Li, Y.-X.; Xun, A.-Y.; Zeng, W.-S.; Jia, C.-H.; Wei, X.-X.; Feng, J.-L.; Zhao, L. Anti-oxidant effects of resveratrol on mice with dss-induced ulcerative colitis. Arch. Med. Res. 2010, 41, 288–294. [Google Scholar] [CrossRef]
- Moradi, M.; Tajik, H.; Razavi Rohani, S.M.; Mahmoudian, A. Antioxidant and antimicrobial effects of zein edible film impregnated with zataria multiflora boiss. Essential oil and monolaurin. LWT Food Sci. Technol. 2016, 72, 37–43. [Google Scholar] [CrossRef]
- Ladeira, M.M.; Santarosa, L.C.; Chizzotti, M.L.; Ramos, E.M.; Neto, O.R.M.; Oliveira, D.M.; Carvalho, J.R.R.; Lopes, L.S.; Ribeiro, J.S. Fatty acid profile, color and lipid oxidation of meat from young bulls fed ground soybean or rumen protected fat with or without monensin. Meat Sci. 2014, 96, 597–605. [Google Scholar] [CrossRef]
- Oviedo-Rondón, E.O. Holistic view of intestinal health in poultry. Anim. Feed Sci. Technol. 2019, 250, 1–8. [Google Scholar] [CrossRef]
- Saleh, A.A.; Amber, K.A.; Mousa, M.M.; Nada, A.L.; Awad, W.; Dawood, M.A.O.; El-Moneim, A.E.-M.E.A.; Ebeid, T.A.; Abdel-Daim, M.M. A mixture of exogenous emulsifiers increased the acceptance of broilers to low energy diets: Growth performance, blood chemistry, and fatty acids traits. Animals 2020, 10, 437. [Google Scholar] [CrossRef] [Green Version]
- Kirrella, A.A.; Abdo, S.E.; El-Naggar, K.; Soliman, M.M.; Aboelenin, S.M.; Dawood, M.A.O.; Saleh, A.A. Use of Corn Silk Meal in Broiler Diet: Effect on Growth Performance, Blood Biochemistry, Immunological Responses, andGrowth-Related Gene Expression. Animals 2021, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Gawish, E.; Mahmoud, S.F.; Amber, K.; Awad, W.; Alzawqari, M.H.; Shukry, M.; Abdel-Moneim, A.-M.E. Effect of Natural and Chemical Colorant Supplementation on Performance, Egg-Quality Characteristics, Yolk Fatty-Acid Profile, and BloodConstituents in Laying Hens. Sustainability 2021, 13, 4503. [Google Scholar] [CrossRef]
- Saleh, A.A.; El-Awady, A.; Amber, K.; Eid, Y.Z.; Alzawqari, M.H.; Selim, S.; Soliman, M.M.; Shukry, M. Effects of Sunflower Meal Supplementation as a Complementary Protein Source in the Laying Hen’s Diet on Productive Performance, Egg Quality, and Nutrient Digestibility. Sustainability 2021, 13, 3557. [Google Scholar] [CrossRef]
- Kumar, S.; Ciraci, C.; Redmond, S.; Chuammitri, P.; Andreasen, C.; Palić, D.; Lamont, S. Immune response gene expression in spleens of diverse chicken lines fed dietary immunomodulators. Poult. Sci. 2011, 90, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Desormeaux, A.; Bergeron, M.G. Sodium Lauryl Sulfate, a Microbicide Effective Against Enveloped and Nonenveloped Viruses. Curr. Drug Targets 2005, 3, 17–30. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredient, % | Starter 0–14 Days | Grower 15–24 Days | Finisher 25–33 Days |
|---|---|---|---|
| Yellow corn | 55.551 | 59.553 | 62.0 |
| Soybean meal, 46% | 38.9 | 34.268 | 31.3 |
| Corn gluten meal, 60% | 0.764 | 0.962 | 0.556 |
| Soya oil | 1.0 | 1.795 | 2.974 |
| Calcium carbonate | 1.465 | 1.325 | 1.231 |
| Mono calcium phosphate | 1.028 | 0.838 | 0.715 |
| Salt | 0.261 | 0.289 | 0.291 |
| Sodium bicarbonate | 0.131 | 0.095 | 0.095 |
| DL methionine, 99% | 0.299 | 0.262 | 0.237 |
| L-Lysine HCl, 98% | 0.154 | 0.17 | 0.162 |
| L-Threonine | 0.062 | 0.058 | 0.054 |
| Premix * | 0.2 | 0.2 | 0.2 |
| Anticoccidia (dicalzoril) | 0.02 | 0.02 | 0.02 |
| Antimycotoxin | 0.15 | 0.15 | 0.15 |
| Phytase 500 FTU | 0.015 | 0.015 | 0.015 |
| Total | 100 | 100 | 100 |
| Chemical Analysis on DM basis | |||
| AME kcal | 2899.678 | 2999.676 | 3099.678 |
| Crude protein, %. | 22.868 | 21.109 | 19.646 |
| Fat, % | 3.883 | 4.761 | 5.95 |
| Digestible LYS, % | 1.4 | 1.29 | 1.2 |
| Digestible methionine and cysteine, % | 1.008 | 929 | 0.864 |
| Digestible THR, % | 0.952 | 0.929 | 0.864 |
| Digestible ARG, % | 1.486 | 1.354 | 1.26 |
| Digestible ILE, % | 0.997 | 0.91 | 0.842 |
| Digestible LEU, % | 1.849 | 1.734 | 1.607 |
| Digestible VAL, % | 1.106 | 1.019 | 0.948 |
| Calcium, % | 0.96 | 870 | 0.81 |
| Available P, % | 0.48 | 0.435 | 0.405 |
| Sodium, % | 0.16 | 0.16 | 0.16 |
| Chloride, % | 0.25 | 0.25 | 0.23 |
| Parameters | Alpha-Monolaurin (g/kg) | |||
|---|---|---|---|---|
| 0 | 0.25 | 0.5 | 1 | |
| Initial body weight, g | 47.42 ± 0.13 | 47.38 ± 0.075 | 47.55 ± 0.062 | 47.44 ± 0.13 |
| Body weight at 7 d, g | 202.2 ± 2.7 | 206.3 ± 4.2 | 206.7 ± 4.1 | 205.5 ± 4.0 |
| Feed intake, g/7 d | 172.3 ± 2.5 | 170.2 ± 2.1 | 171.8 ± 2.9 | 173.8 ± 3.1 |
| FCR, 7 d | 0.852 ± 0.02 | 0.825 ± 0.03 | 0.831 ± 0.03 | 0.846 ± 0.05 |
| Body weight at 14 d, g | 514.2 ± 9.4 | 518.5 ± 8.9 | 520.2 ± 8.2 | 517.8 ± 9.6 |
| Feed intake, g/14 d | 542.4 ± 11.5 | 538.7 ± 12.1 | 539.5 ± 11.8 | 540.7 ± 12.2 |
| FCR, 14 d | 1.055 ± 0.017 a | 1.038 ± 0.015 b | 1.037 ± 0.013 b | 1.044 ± 0.015 a,b |
| Body weight at 21 d, g | 977.2 ± 19.4 | 986.5 ± 18.6 | 984.5 ± 15.8 | 980.3 ± 17.4 |
| Feed intake, g/21 d | 1185.2 ± 23.2 | 1175.5 ± 26.2 | 1172.3 ± 25.2 | 1180.4 ± 24.8 |
| FCR, 21 d | 1.213 ± 0.04 a | 1.192 ± 0.05 b | 1.191 ± 0.06 b | 1.204 ± 0.015 a,b |
| Body weight at 28 d, g | 1549.6 ± 28.4 b | 1576.2 ± 31.2 a | 1570.2 ± 25.5 a | 1568.0 ± 30.2 a,b |
| Feed intake, g/28 d | 2113.7 ± 37.1 | 2110.2 ± 36.9 | 2104.7 ± 37.6 | 2118.1 ± 31.2 |
| FCR, 28 d | 1.364 ± 0.017 a | 1.339 ± 0.015 b | 1.340 ± 0.013 b | 1.351 ± 0.015 a,b |
| Body weight at 33 d, g | 1979.9 ± 18.9 | 2052.5 ± 41.6 | 2020.7 ± 15.1 | 2013.3 ± 40.05 |
| Body weight gain, g/33 d | 1932.6 ± 18.8 | 2005.2 ± 41.6 | 1973.2 ± 15.1 | 1965.8 ± 40.04 |
| Feed intake, g/33 d | 2966.5 ± 27.5 | 2954.5 ± 56.1 | 2902.3 ± 27.9 | 2896.3 ± 54.1 |
| FCR, 33 d | 1.499 ± 0.017 a | 1.440 ± 0.015 a,b | 1.436 ± 0.013 b | 1.439 ± 0.015 a,b |
| EPEF | 380.91 ± 4.2 b | 411.42 ± 3.9 a | 405.98 ± 4.5 a,b | 403.6 ± 4.8 a,b |
| Mortality, % | 1.66 ± 0.23 | 1.66 ± 0.31 | 1.66 ± 0.12 | 1.66 ± 0.19 |
| CPU (%) | 81.6 ± 0.6 | 84.6 ± 0.7 | 82.7 ± 0.7 | 82.6 ± 0.7 |
| CFU (%) | 42.9 ± 0.8 | 48.5 ± 0.7 | 45.5 ± 1.1 | 45.8 ± 0.9 |
| EEU (%) | 35.6 ± 0.9 c | 54.3 ± 0.8 a | 48.1 ± 0.9 b | 44.4 ± 0.5 b |
| ME kcal/kg | 65.5 ± 0.9 | 67.3 ± 0.6 | 66.6 ± 0.5 | 66.7 ± 0.9 |
| Parameters | Alpha-Monolaurin (g/kg) | |||
|---|---|---|---|---|
| 0 | 0.25 | 0.5 | 1 | |
| Carcass weight, g/100 g BW | 64.8 ± 1.51 | 63.2 ± 3.02 | 64.4 ± 1.1 | 61.7 ± 4.9 |
| Breast muscle weight, g/100 g BW | 25.0 ± 1.8 | 24.9 ± 2.2 | 23.9 ± 2.1 | 22.6 ± 2.3 |
| Thigh muscle weight, g/100 g BW | 17.6 ± 1.3 | 16.2 ± 1.6 | 16.7 ± 1.6 | 17.4 ± 1.02 |
| Liver weight, g/100 g BW | 2.6 ± 0.16 | 2.9 ± 0.82 | 2.6 ± 0.69 | 2.5 ± 0.37 |
| Gizzard weight, g/100 g BW | 0.94 ± 0.32 | 0.93 ± 0.14 | 0.99 ± 0.12 | 1.09 ± 0.17 |
| Abdominal fat, g/100 g BW | 1.68 ± 0.33 | 1.88 ± 0.77 | 1.71 ± 0.40 | 0.59± 1.9 |
| Spleen weight, g/100 g BW | 0.06 ± 0.01 b | 0.15 ± 0.05 a | 0.14 ± 0.03 a | 0.10 ± 0.05 a,b |
| Heart weight, g/100 g BW | 0.534 ± 0.09 | 0.464 ± 0.13 | 0.523 ± 0.10 | 0.433 ± 0.03 |
| Parameters | Alpha-Monolaurin (g/kg) | |||
|---|---|---|---|---|
| 0 | 0.25 | 0.5 | 1 | |
| TC, mg/dL | 167 ± 5.5 | 166.5 ± 3.6 | 172 ± 6.5 | 165 ± 3.4 |
| TG, mg/dL | 6.6 ± 0.16 | 6.25 ± 0.14 | 5.17 ± 0.86 | 6.0 ± 0.21 |
| HDL, mg/dL | 63.3 ± 2.1 | 70.8 ± 3.2 | 68.0 ± 4.3 | 65.0 ± 3.6 |
| LDL, mg/dL | 58.0 ± 3.1 | 51.6 ± 4.9 | 47.5 ± 4.1 | 52.5 ± 2.4 |
| Glucose, mg/dL | 145 ± 9.6 | 125 ± 7.9 | 148 ± 7.1 | 150 ± 10.4 |
| AST (IU/I) | 178.8 ± 25 | 168.5 ± 45 | 146 ± 26 | 193.5 ± 24 |
| ALT (IU/I) | 6.4 ± 1.1 a | 4.4 ± 0.49 b | 4.6 ± 0.54 b | 5.6 ± 0.64 a,b |
| Creatinine, mg/dL | 0.622 ± 0.11 | 0.713 ± 0.03 | 0.693 ± 0.06 | 0.683 ± 0.02 |
| TP, mg/dL | 3.25 ± 0.26 b | 3.37 ± 0.25 b | 5.13± 0.58 a | 3.68 ± 0.23 b |
| Albumin, mg/dL | 1.73 ± 0.08 b | 1.85 ± 0.18 b | 3.75 ± 0.27 a | 2.09 ± 0.43 b |
| Globulin, mg/dL | 1.52 ± 0.23 | 1.52 ± 0.24 | 1.38 ± 0.37 | 1.52 ± 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleh, A.A.; El-Gharabawy, B.; Hassan, A.; Badawi, N.; Eid, Y.; Selim, S.; Shukry, M.; Dawood, M. Effect of Dietary Inclusion of Alpha-Monolaurin on the Growth Performance, Lipid Peroxidation, and Immunity Response in Broilers. Sustainability 2021, 13, 5231. https://doi.org/10.3390/su13095231
Saleh AA, El-Gharabawy B, Hassan A, Badawi N, Eid Y, Selim S, Shukry M, Dawood M. Effect of Dietary Inclusion of Alpha-Monolaurin on the Growth Performance, Lipid Peroxidation, and Immunity Response in Broilers. Sustainability. 2021; 13(9):5231. https://doi.org/10.3390/su13095231
Chicago/Turabian StyleSaleh, Ahmed Ali, Bahaa El-Gharabawy, Aziza Hassan, Nemat Badawi, Yahya Eid, Shaimaa Selim, Mustafa Shukry, and Mahmoud Dawood. 2021. "Effect of Dietary Inclusion of Alpha-Monolaurin on the Growth Performance, Lipid Peroxidation, and Immunity Response in Broilers" Sustainability 13, no. 9: 5231. https://doi.org/10.3390/su13095231