Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment

Department of Microbiology, Soochow University, Taipei 11101, Taiwan
*
Author to whom correspondence should be addressed.
Sustainability 2020, 12(13), 5410; https://doi.org/10.3390/su12135410
Submission received: 10 May 2020
/
Revised: 25 June 2020
/
Accepted: 2 July 2020
/
Published: 4 July 2020
(This article belongs to the Special Issue Constructed and Floating Wetlands for Sustainable Water Reclamation)
Abstract
:Acetaminophen (ACE) is a widely used medicine. Currently, concerns regarding its potential adverse effects on the environments are raised. The aim of this study was to evaluate ACE biodegradation in mangrove sediments under aerobic and anaerobic conditions. Three ACE biodegradation strategies in mangrove sediments were tested. The degradation half-lives (t1/2) of ACE in the sediments with spent mushroom compost under aerobic conditions ranged from 3.24 ± 0.16 to 6.25 ± 0.31 d. The degradation half-lives (t1/2) of ACE in sediments with isolated bacterial strains ranged from 2.54 ± 0.13 to 3.30 ± 0.17 d and from 2.62 ± 0.13 to 3.52 ± 0.17 d under aerobic and anaerobic conditions, respectively. The degradation half-lives (t1/2) of ACE in sediments amended with NaNO3, Na2SO4 and NaHCO3 under anaerobic conditions ranged from 1.16 ± 0.06 to 3.05 ± 0.15 d, 2.39 ± 0.12 to 3.84 ± 0.19 d and 2.79 ± 0.14 to 10.75 ± 0.53 d, respectively. The addition of the three electron acceptors enhanced ACE degradation in mangrove sediments, where NaNO3 yielded the best effects. Sixteen microbial genera were identified as the major members of microbial communities associated in anaerobic ACE degradation in mangrove sediments with addition of NaNO3 and Na2SO4. Three (Arthrobacter, Enterobacter and Bacillus) of the sixteen microbial genera were identified in the isolated ACE-degrading bacterial strains.
1. Introduction
Acetaminophen (N-acetyl-p-aminophenol; ACE) is a widely used representative medicine. As a nonsteroidal anti-inflammatory drugs (NSAIDs), ACE exhibits analgesic and antipyretic properties and acts via inhibition of cyclooxygenase enzymes [1]. The reported concentrations of ACE range from 0.003 to 30 mg L−1 in stream water, sewage treatment plant influents and effluents [2,3,4]. The frequent detection of ACE in aquatic environments has raised concerns regarding its potential deleterious effects on the environments [5,6,7,8].
Mangrove ecosystems along the coastlines of tropical and subtropical regions are important intertidal estuary wetlands that considered to be significant sinks of pollutants from contaminated tidal water and discharges from freshwater [9]. The mangroves of Guandu and Bali are located on the banks of the Danshui River, one of the most polluted rivers in northern Taiwan. The concentrations and degradation of nonylphenol, sulfonamides and polycyclic aromatic hydrocarbons in the mangrove sediments have been reported [10,11,12,13].
Physicochemical methods such as primarily advanced oxidation processes (AOPs), homogeneous and heterogeneous photocatalysis, Fenton and Fenton like reactions, ozonation, and methods involving ultrasound and microwave treatments, or electrochemical processes are not appropriate for treating ACE in mangrove sediments [14]. Biodegradation is an effective strategy to remove organic pollutants from sediments. Most studies on pharmaceutical biodegradation are focused on their removal during wastewater treatment processes [15]. Some investigations evaluated the microbial degradation of drugs and artificial compounds in freshwater [16] as well as in ocean and estuary waters [17]. The microbial degradation of drugs depends on the prevailing oxygen availability and redox conditions in sediments. However, little is known regarding the aerobic and anaerobic biodegradation potential of ACE in mangrove sediments.
Biodegradation is believed to be an effective strategy for eliminating contaminating ACE in environments. To enhance the efficiency of degradation, three remedial strategies have been proposed: natural attenuation, bioaugmentation and biostimulation [18]. Microbial degradation of ACE has been observed in bacterial strains. Pseudomonas aeruginosa strain HJ1012 was isolated on ACE as a sole carbon [19]. Pseudomonas moorei KB4 can metabolize ACE, with p-aminophenol and hydroquinone identified as degradation products [14]. Two ACE-metabolizing strains, Delftia tsuruhatensis and Pseudomonas aeruginosa, were isolated from the membrane bioreactor [20]. White rot fungus Pleurotus eryngii is one of the most widespread mushrooms consumed in the world. spent mushroom compost (SMC) is a mushroom industry waste which contains extracellular enzymes with organic pollutant degradation ability [21]. The addition of enzyme containing microcapsules (MC) was found to be effective for aerobic degradation of organic pollutants [12] and the aerobic degradation of tetracyclines in the river sediments [22]. Addition of NaHCO3, Na2SO4, andNaNO3 could create methanogenic conditions, sulfate-reducing conditions and nitrate-reducing conditions, respectively, as well as enhance anaerobic degradation of organic pollutants [13]. Therefore, the addition of ACE-degrading bacteria, MC and electron acceptors such as NaNO3, Na2SO4 and NaHCO3 were used to promote ACE degradation in this study.
The aim of this study is to evaluate strategies for enhancing biodegradation of ACE in mangrove sediments under aerobic and anaerobic conditions. Three strategies, including addition of MC, addition of ACE-degrading bacterial strains isolated from sediments and addition of electron acceptors (NaNO3, Na2SO4 or NaHCO3). The microbial communities involved in aerobic and anaerobic degradation of ACE in the mangrove sediments were investigated.
2. Materials and Methods
2.1. Chemicals
The ACE (99.0% analytical standard) used in this study was purchased from the Aldrich Chemical Co. Solvents were purchased from Mallinckrodt, while all other chemicals were purchased from Sigma Chemical Co. (USA).
2.2. Sample Collection
Samples were taken from the Guandu sampling site (25.11°68.43′ N, 121.46°41.53′ E) and Bali sampling site (25.15°86.13′ N, 121.43°55.75′ E) at Tamsui, northern Taiwan. Figure S1 shows the locations of the two sample collection sites. Sediments (0–15 cm) were collected in spring (March 2015). All samples from each sampling site were randomly collected, in triplicate, from an area approximately 1 m2. Adaptation was performed by adding 100 mg kg−1 ACE to 500 g of sediment at 14-d intervals at 30 °C for 6 months under aerobic or anaerobic conditions. The sediment samples were analyzed for salinity, temperature, TOC, bacterial count, and ACE concentration. The salinity and temperature were measured by salinity/temperature meter (model 30, YSl, USA). The sediment samples were mixed in a ratio of 1:1 with distilled water in a beaker before inserting the probe. Readings were taken after allowing the instrument to stabilize. The TOC was measured using a TOC Analyzer (OI Analytical 1030 W, USA). The bacterial counts were enumerated by pour plate method and grown on R2A agar. Sediments properties for the Guandu and Bali sampling sites were listed in Table 1.
2.3. Medium
The aerobic medium contained the followings (mg L−1): K2HPO4, 65.3; KH2PO4, 25.5; Na2HPO4·12 H2O, 133.8; NH4Cl, 5.1; CaCl2, 82.5; MgSO4·7H2O, 67.5; and FeCl3·6H2O, 0.75. The anaerobic medium contained the followings (mg L−1): NH4Cl, 2.7; MgCl2·6H2O, 0.1; CaCl2·2H2O, 0.1; FeCl2·4H2O, 0.02; K2HPO4, 0.27; KH2PO4, 0.35; and resazurin, 0.001. The pH of the medium was adjusted to 7.0 using 0.9-mM titanium citrate as a reducing reagent before autoclaving. The aerobic and anaerobic medium were used under aerobic and anaerobic conditions, respectively.
2.4. Preparation of Enzyme Extract-Containing Microcapsules (MC)
The SMC of Pleurotus eryngii was obtained from a mushroom cultivation farm in Chiayi, Taiwan. The enzyme extract was extracted from the 120 g SMC with 600 mL sodium acetate buffer (pH 5.0) for 3 h at 4 °C. Alginate solution was made by dissolving sodium alginate (4 wt%) in 0.9-wt% sodium chloride with stirring for 1 h. Enzyme extract solution was then added into the alginate solution. An electrostatic droplet generator was used to prepare the MC. The mixture was drawn into a 10-mL syringe fitted with a needle and attached to a syringe pump that provided a steady solution flow rate of 25.2 mL/h and fixed voltage (12 kV) into a gently agitated aqueous solution of calcium chloride 1.5 wt% to form MC of 250 nm in diameter for experiments [12].
2.5. Experimental Setting
Experiments were performed under aerobic and anaerobic conditions. Aerobic experiments were performed using 125 mL serum bottles that contained 40 mL of aerobic medium, 5 g of sediment, 5 mL of the MC and 50 mg kg−1 of ACE, which were incubated on a rotary shaker (120 rpm) at 30 °C in the dark. Anaerobic experiments were performed using serum bottles containing 45 mL of anaerobic medium, 5 g of sediment, 20 mM of electron acceptors (NaNO3, Na2SO4 or NaHCO3) and 50 mg kg−1 of ACE. All experiments were conducted in an anaerobic glove box (Forma Scientific, model 1025 S/N, USA) filled with N2 (85%), H2 (10%) and CO2 (5%) gases. The bottles were capped with butyl rubber stoppers and crimp seals, wrapped in aluminum foil, and then incubated without shaking at 30 °C. Inoculated controls containing 45 mL of aerobic or anaerobic medium, 5 g of sediment and 50 mg kg−1 of ACE (without the addition of MC or electron acceptors) were incubated at 30 °C. Sterile controls containing 45 mL of aerobic or anaerobic medium and 5 g of sediment were autoclaved at 121 °C for 30 min. The duration of aerobic or anaerobic experiments was 24 d. Each experiment was repeated 3 times.
The ACE concentrations in the sterile controls were examined for a 24-d incubation period. The remaining amount of ACE in the sediment ranged from 98.5% to 96.8%, indicating that the aerobic and anaerobic ACE degradation in all of the experiments were due to microbial activity.
2.6. Isolation, Identification and Tests of the ACE-Degrading Bacteria
ACE-degrading bacteria were isolated from sediments under aerobic or anaerobic conditions. The enrichment procedure was performed using aerobic or anaerobic medium agar plates containing 10 mg L−1 of ACE to isolate bacterial clones. To confirm the ACE-degrading ability of isolated bacterial strains, aerobic degradation experiments were performed using 5 mL cultures of the aerobic bacterial strains, 45 mL of aerobic medium and 2 mg L−1 of ACE on a shaker (120 rpm) at 30 °C in the dark. Anaerobic experiments were performed using serum bottles containing 45 mL of anaerobic medium, 5 g of sediment and 2 mg L−1 of ACE and were conducted in an anaerobic glove box. The bottles were capped with butyl rubber stoppers and crimp seals, wrapped in aluminum foil, and then incubated without shaking at 30 °C. Samples were periodically taken to analyze residual ACE. Each individual colony was purified and then identified by 16S rRNA gene sequencing after PCR amplification with the primers F8 and R1510. The PCR products were sequenced using an ABI Prism automatic sequencer. The 16S rRNA gene database was searched using BLAST of the National Center for Biotechnology Information (NCBI). Phylogenetic analysis was performed using ClustalX 2.0 with 1000 bootstraps [23].
Isolated bacterial strains were added to Guandu and Bali sediments under aerobic and anaerobic conditions to test their ACE-degrading ability. ACE degradation was assessed following the addition of aerobic and anaerobic ACE-degrading bacteria into the sediment. The experiment was performed using 1 mL of ACE-degrading bacteria, 5 g sediment, 45 mL of medium and 2 mg L−1 of ACE.
2.7. Analytical Methods
ACE was extracted from sediment samples twice by water (with 0.1% formic acid): acetonitrile: methanol (10:3:1), and then extracted using a C18 solid phase extraction cartridge. The SPE cartridges were Chromabond® HR-X (500 mg, 6 mL), and SPE was conducted at a sample pH of 3.0.
Extracts were analyzed using an Agilent 1260 HPLC system equipped with a 4.6 × 250 mm column (Zorbax Eclipse Plus C18, Agilent) and a photodiode array detector with monitoring at 270 nm. The mobile phase consisted of acetonitrile and water (containing 0.1% formic acid) at a ratio of 30%:70%. The recovery percentage was 96.4%, and the detection limit (LOD) was 0.05 mg L−1, respectively. The remaining percentage of ACE (Rp) was calculated using the formula:
where RCACE is the residual ACE concentration and ICACE is the initial ACE concentration. The ACE degradation data collected in this study well fit first-order kinetics
where C0 is the initial ACE concentration, C is the ACE concentration, t is the time period, t1/2 is the half-live, and k is the degradation rate constant).
Rp = (RCACE/ICACE) × 100%
t = −ln(C/C0)/k
2.8. Microbial Community Analysis
Total DNA was extracted from experimental samples using a PowerSoil DNA Isolation kit (QIAGEN). DNA from three experiments (bottles) of each treatment were extracted and pooled together to perform NGS. The V5–V8 variable region of the 16S rRNA gene was amplified as described previously [12,13]. Next-generation sequencing (NGS) was performed at the Genome Center of the National Yang-Ming University, Taiwan using a MiSeq platform (Illumina, Inc.). Chimeric sequences in the 16S rRNA gene sequence data were removed using Chimera Check. The classifier from the RPD pipeline was used to assign taxonomic groups with a 95% sequence similarity. Cluster analysis of the microbial community compositions in the experimental samples was performed using the heatmap3 package of R. The differences in microbial composition between experimental samples were identified using the prop.test (two proportion test) function in the package MASS of R (https://www.r-project.org/). A p-value of less than 0.05 was considered to be significant. Specific microbial communities (such as those involved in the nitrogen cycle and aromatic compound degradation) were identified by integration of NGS data and the microbial list in the KEGG modules [24].
3. Results and Discussion
3.1. Aerobic ACE Degradation
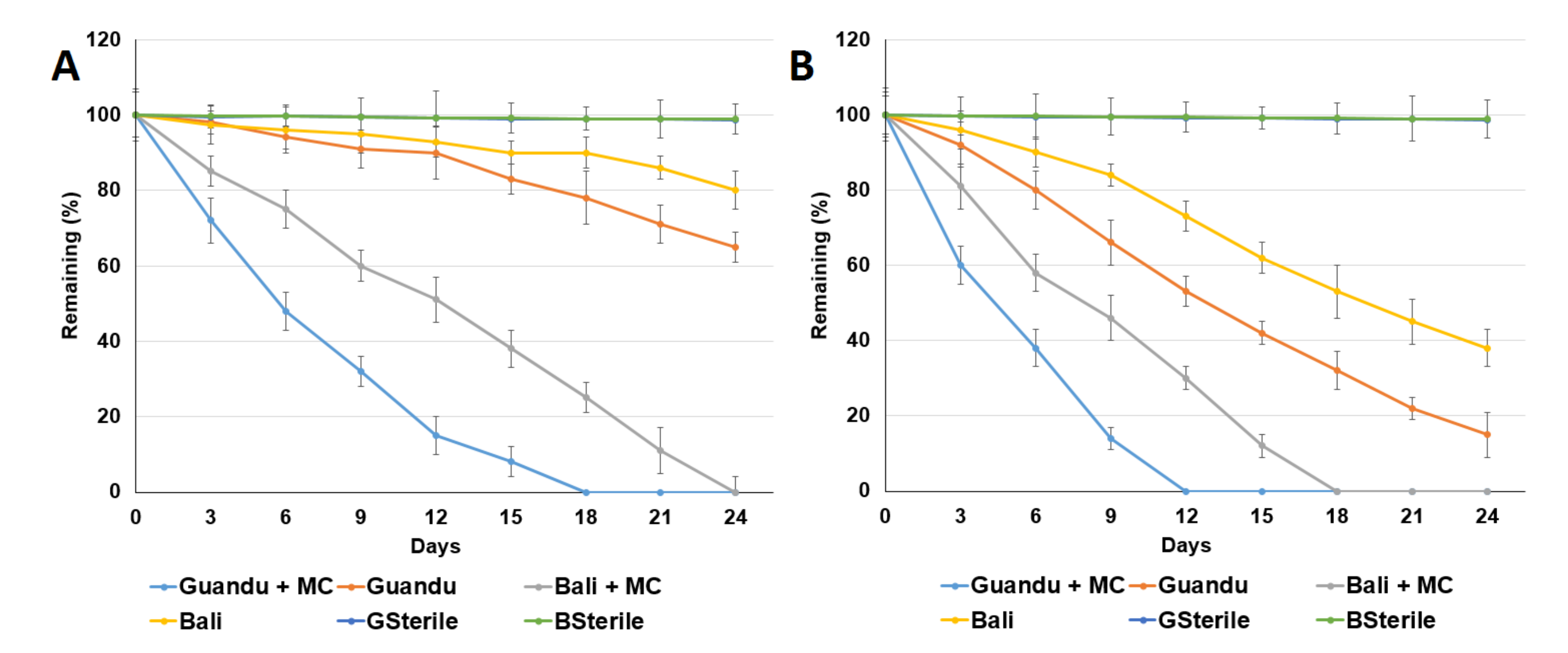
The aerobic degradation of the ACE in the Guandu and Bali sediments are shown in Figure 1. We first found that the remaining percentage of ACE observed after 24 d in the Guandu and Bali non-ACE-adapted sediments without the addition of MC was 65.1% ± 4.1% and 80.8% ± 3.4%, respectively. The remaining ACE in the Guandu and Bali ACE-adapted sediments without added MC was 15.4% ± 3.7% and 38.4% ± 4.1%, respectively. The remaining ACE after 24 d in the Guandu and Bali ACE-adapted sediments without added MC was 15.4% ± 3.7% and 38.4% ± 4.1%, respectively. For the Guandu and Bali ACE-adapted sediment supplemented with MC, ACE was completely degraded after 12 and 18 d, respectively.
The ACE degradation data in Figure 1 were fitted to first-order kinetics. For the Guandu and Bali non-ACE-adapted sediments supplemented with or without MC, the observed ACE half-lives were 3.24 ± 0.06 and 33.55 ± 0.16 d and 6.25 ± 0.31 and 66.66 ± 3.33 d, respectively. For the Guandu and Bali ACE-adapted sediments supplemented with or without MC, the observed ACE half-lives were 2.52 ± 0.12 and 7.42 ± 0.37 d and 4.33 ± 0.31 and 14.20 ± 0.71 d, respectively.
The results indicate that the rate of ACE degradation was higher in the Guandu sediments than in the Bali sediments. As shown in Table 1, the Guandu sediments exhibit higher TOC, bacterial counts and lower salinity and ACE concentrations than those of the Bali sediments. The salinities at the Guandu sediment and Bali sediment were 11.1‰ ± 0.3‰ and 16.5‰ ± 1.5‰, respectively. Salinity may affect organic pollutants degradation in the environment [25]. This result is consistent with biodegradation of phenanthrene by bacteria isolated from mangrove sediments [26].
A comparison of ACE degradation in sediments with or without ACE adaptation indicated that the adaptation process enhanced aerobic ACE degradation. The adaptation of microbial populations occurred by the induction of enzymes necessary for degradation followed by an increase in the population of degrading organisms [18]. The results also revealed the degradation of ACE was enhanced with the addition of MC in the sediments. Similar results were observed in a previous study which reported that the addition of MC enhanced sulfonamide degradation in sediments [12].
3.2. ACE Degradation by ACE-Degrading Bacteria
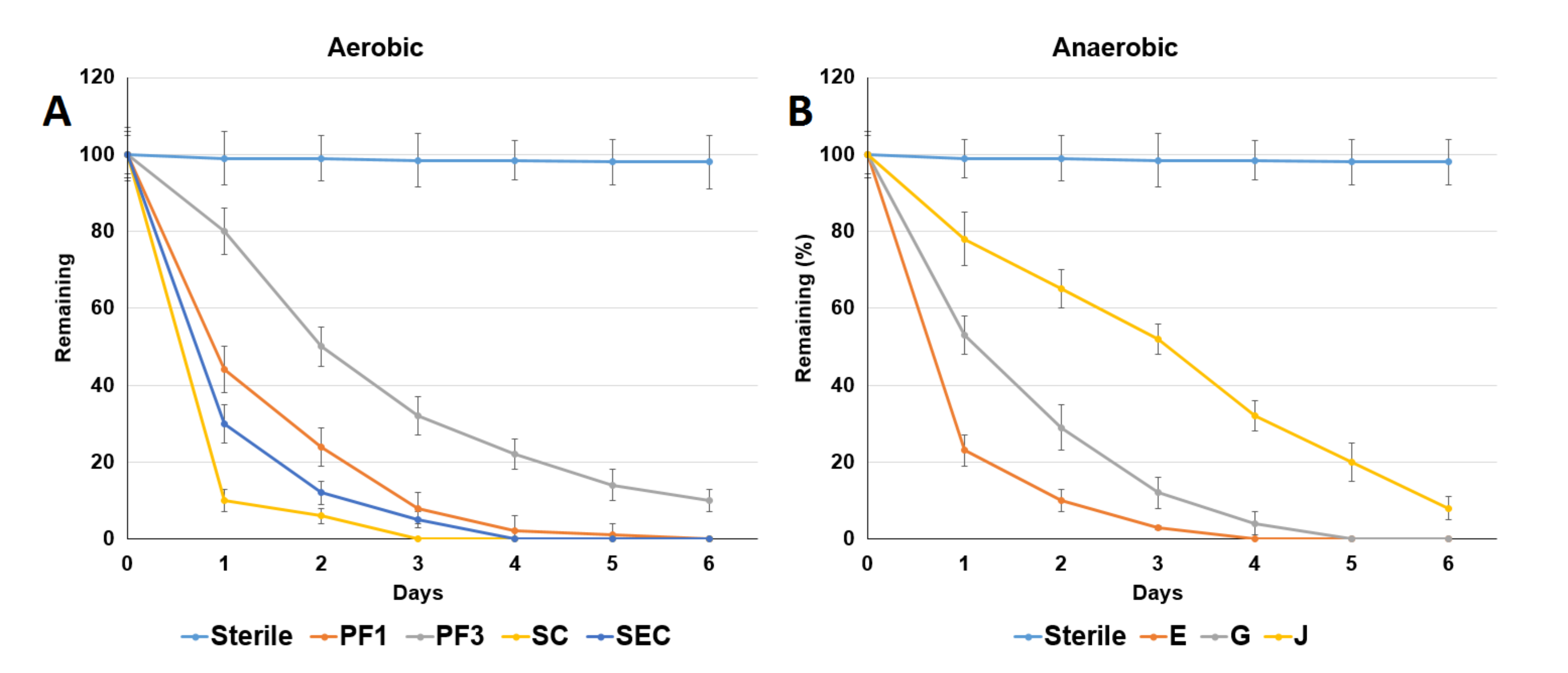
Ten bacterial strains with the capability to using ACE as a carbon source were isolated from the ACE-adapted sediments under aerobic conditions. Four of the ten strains (PF1, PF3, SC and SEC) exhibited higher ACE-degrading activities than the other strains (Figure 2A). The ACE degradation data collected in this study well fit first-order kinetics. The observed half-lives of ACE in the presence of strains SC, SEC, PF1 and PF3 were 0.32 ± 0.01, 0.48 ± 0.02, 0.55 ± 0.03 and 1.31 ± 0.06 d, respectively. The ACE-degrading ability of the strains exhibited the following order: strain SC > strain SEC > strain PF1 > strain PF3.
Nine bacterial strains with the ability to use ACE as a carbon source were isolated from the ACE-adapted sediments under anaerobic conditions. Three of the nine strains (E, G and J) exhibited a greater ACE-degrading capability than the other strains (Figure 2B). The ACE degradation data collected in this study well fit first-order kinetics. The observed degradation half-lives of ACE in the presence of strains E, G and J were 0.42 ± 0.02, 0.56 ± 0.03 and 0.67 ± 0.03 d, respectively. The ACE-degrading ability of the strains exhibited the following order: strain E > strain G > strain J.
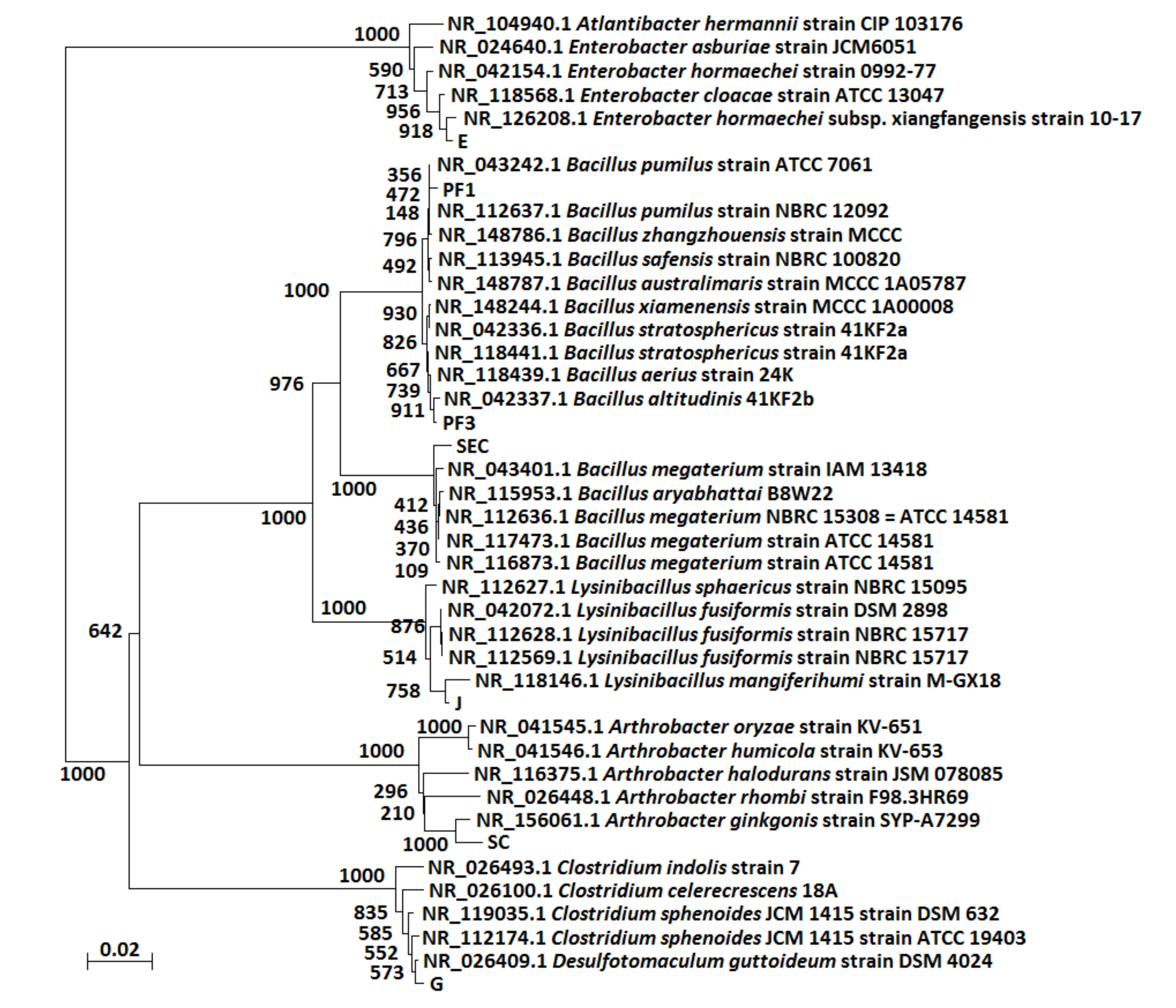
A phylogenetic analysis of the 16S rRNA gene sequences of the isolated bacterial strains is shown in Figure 3. The aerobic strains PF1, PF3, SC and SEC were observed to be closely associated with Bacillus pumilus, Bacillus aerius, Arthrobacter ginkgonis and Bacillus aryabhattai, with similarities of 99%, 100%, 99% and 100%, respectively. In addition, the anaerobic strains E, G and J were observed be closely related to Enterobacter hormechei, Clostridium sphenoides and Lysinibacillus sp., with similarities of 99%, 99% and 99%, respectively (Table 2).
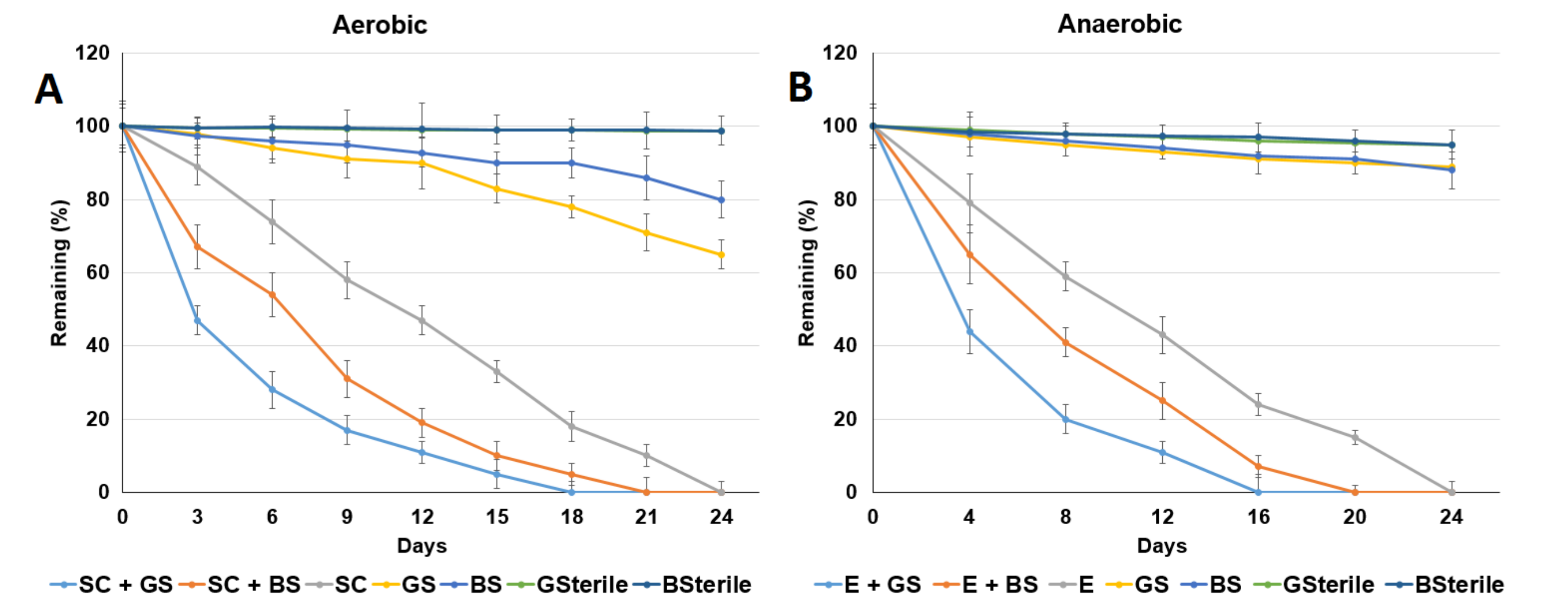
As shown in Figure 4, ACE degradation in the sediments was enhanced with the addition of the bacterial strains SC and E under aerobic and anaerobic conditions, respectively. The ACE degradation data in Figure 4 were fitted to first-order kinetics. The observed ACE half-lives (d) were 2.54 and 3.30 d in the Guandu and Bali sediments with the additions of bacterial strain SC, while values of 2.62 and 3.52 d were obtained in the Guandu and Bali sediments with the addition of bacterial strain E. These results indicated that ACE degradation in the Guandu and Bali sediments was enhanced by the addition of bacterial strains SC and E under aerobic and anaerobic conditions, respectively.
Bacillus pumilus has been previously shown to degrade total petroleum hydrocarbons in contaminated soil [27], while Bacillus aerius has been isolated, characterized and identified as a potential diuron-degrading bacterium [28]. Bacillus aryabhattai was previously isolated from pulp and paper mill wastewater and characterized as having lignin-degrading potential [29]. Arthrobacter ginkgonis was a Gram-positive, aerobic strain which displayed a rod–coccus growth lifecycle, was isolated from the rhizosphere of Ginkgo biloba L [30]. Enterobacter hormechei has been isolated from activated sludge and shown to transform diclofenac [31] as well as to bioconvert lutein into a new compound, 8-methyl-alpha-ionone [32]. Clostridium sphenoides is able to metabolize citrate as a sole carbon source, and a Clostridium sp. was previously shown to be capable of fermenting glucose, but not citric acid [33]. Lysinibacillus fusiformis has been shown to decolorize a selected azo dye [34]. These reports showed that Bacillus pumilus, Bacillus aerius, Bacillus aryabhattai, Enterobacter hormechei, Clostridium sphenoides and Lysinibacillus sp. had the degradation ability. However, there is no information about the degradation properties of Arthrobacter ginkgo.
3.3. Addition of Electron Acceptors Improved Anaerobic ACE Degradation
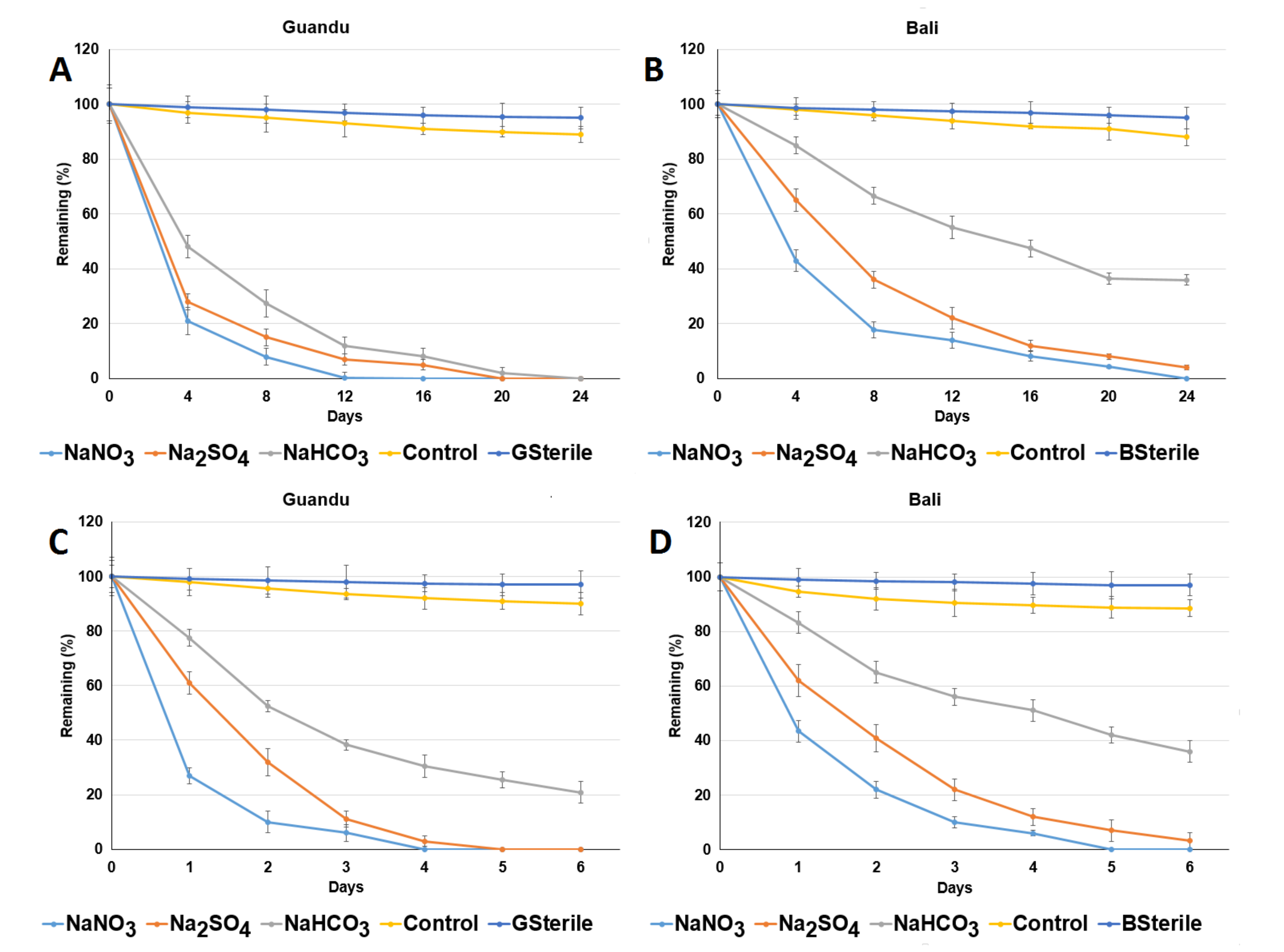
As shown in Figure 5A, ACE was completely degraded after 12, 20 and 24 d in the Guandu non-ACE-adapted sediment supplemented with NaNO3, Na2SO4 and NaHCO3, respectively. For the Bali ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the remaining amount of ACE detected after 24 d was ND (non-detected), 4.1% ± 0.6% and 35.9% ± 1.8%, respectively (Figure 5B). The ACE anaerobic degradation data in Figure 5 were fitted to first-order kinetics. For the Guandu ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the observed ACE degradation half-lives were 1.16 ± 0.06, 2.39 ± 0.12 and 2.79 ± 0.14 d, respectively. For the Bali non-ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the observed ACE half-lives were 3.05 ± 0.15, 3.84 ± 0.19 and 10.75 ± 0.53 d, respectively. In contrast, for the Guandu non-ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the remaining amount of ACE detected after 6 d was ND, ND and 20.8% ± 1.1%, respectively (Figure 5C). For the Bali ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the remaining amount of ACE detected after 6 d was ND, 3.3% ± 0.6% and 35.9% ± 1.8%, respectively (Figure 5D). For the Guandu non-ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the observed ACE half-lives were 0.48 ± 0.02, 0.64 ± 0.3 and 1.79 ± 0.09 d, respectively. For the Bali ACE-adapted sediments supplemented with NaNO3, Na2SO4 or NaHCO3, the observed ACE half-lives were 0.55 ± 0.03, 0.93 ± 0.05 and 2.85 ± 0.14 d, respectively.
These results indicate that the rate of ACE degradation was higher in the Guandu sediments than in the Bali sediments with or without ACE adaptation. A comparison of ACE degradation in the sediments with or without ACE adaptation revealed that the ACE adaptation process enhanced ACE anaerobic degradation. Microorganisms with ACE-degrading activity may increase in response to ACE adaptation, and these results are similar to those of a previous study regarding the effects of nonylphenol adaptation on nonylphenol anaerobic degradation in sediments [11]. Compared with the control treatment (sediments without NaHCO3, Na2SO4 or NaNO3), the anaerobic degradation of ACE was enhanced under methanogenic, sulfate-reducing and nitrate-reducing conditions in the sediments supplemented with NaNO3, Na2SO4 or NaHCO3. The ACE degradation rates observed under the three conditions exhibited the following order: nitrate reducing conditions > sulfate-reducing conditions > methanogenic conditions. The results indicate that nitrate-reducing bacteria and sulfate-reducing bacteria, and methane may be the major contributors to anaerobic ACE degradation in all of the tested anaerobic mesocosms.
Results of acetaminophen degradation half-lives (t1/2, day) using different treatments (addition of ACE-degrading bacterial strains, MC and electron acceptors) were summarized in Table 3. All of the treatments can enhance the ACE biodegradation in mangrove sediments. Addition of NaNO3 exhibited the best effects for ACE biodegradation in mangrove sediments.
3.4. Analysis of Microbial Communities Associated with ACE Degradation
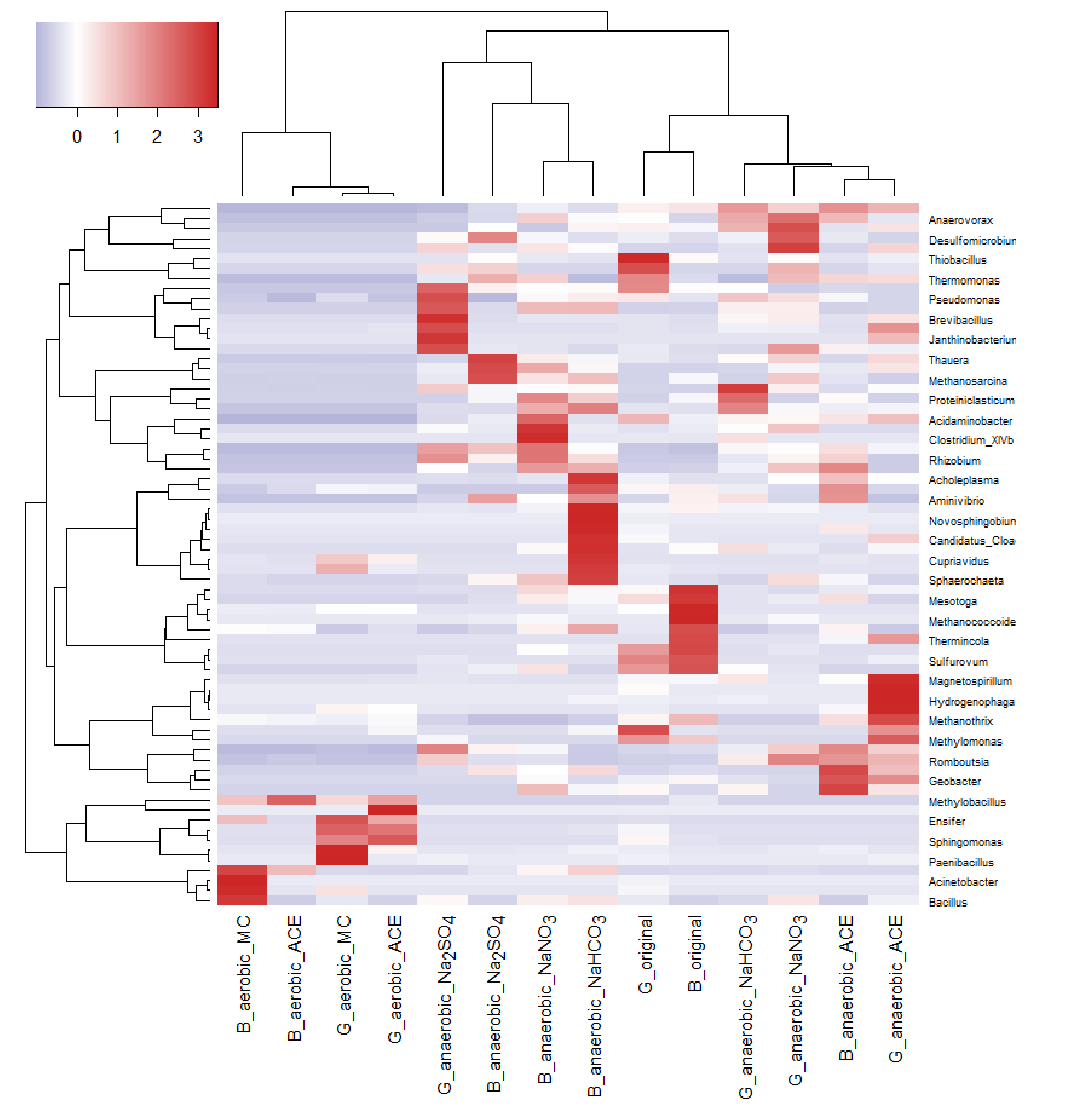
To gain deeper insights into ACE degradation, NGS was performed to analyze the microbial community compositions of aerobic sediments supplemented with MC and anaerobic sediments supplemented with NaNO3, Na2SO4 or NaHCO3. As shown in Figure 6, the microbial community compositions of the aerobic sediments supplemented with MC were highly different from those of anaerobic sediments supplemented with NaNO3, Na2SO4 or NaHCO3.
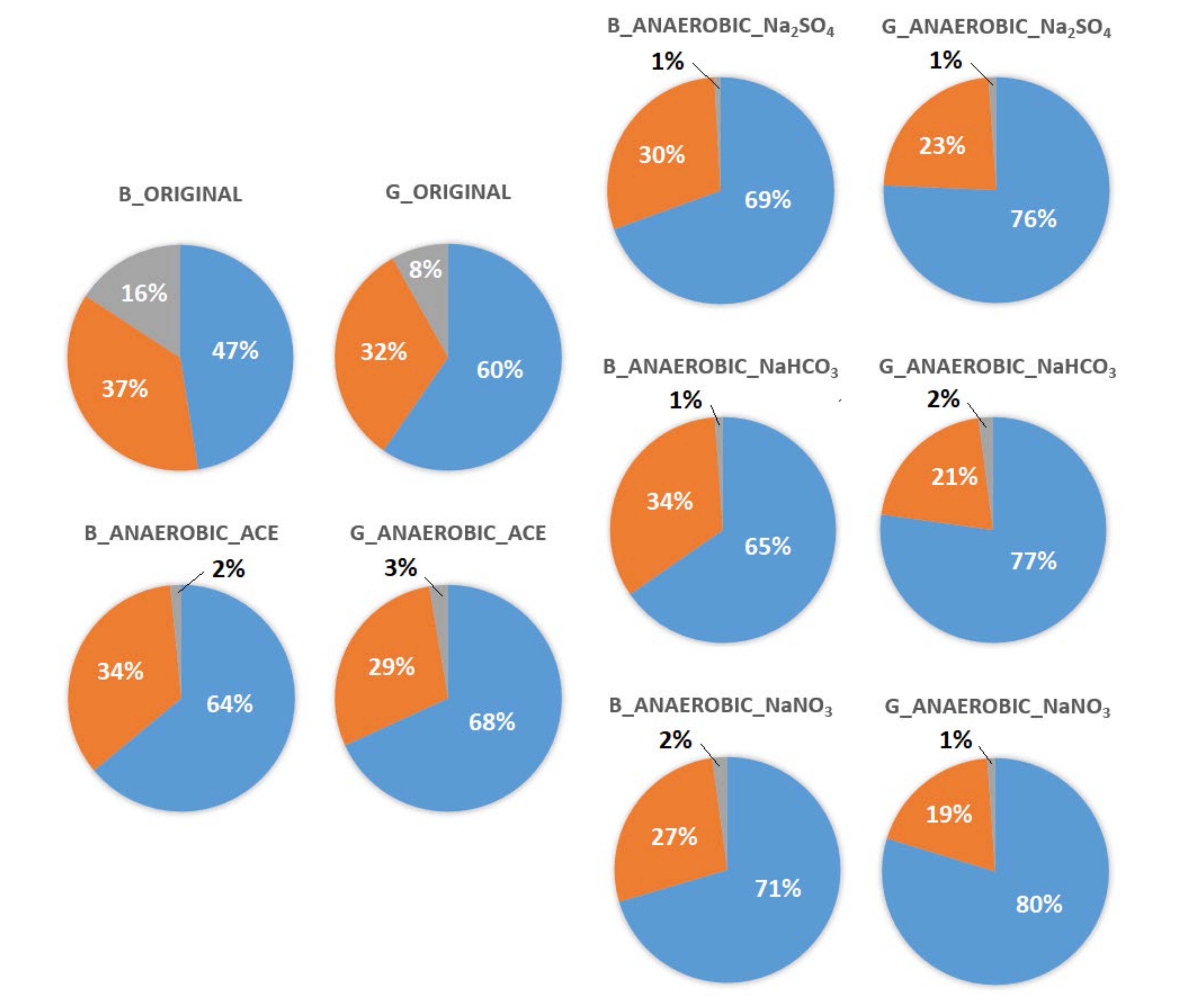
The effects of the addition of NaNO3, Na2SO4 or NaHCO3 are shown in Figure 7. Under anaerobic conditions and with the addition of the electron acceptors, a large increase in the proportion of detected bacteria in the Guandu and Bali sediments (from 60% to 80% and from 47% to 71%, respectively) was observed, with a simultaneous reduction in the proportion of archaea (from 32% to 19% and from 37% to 27%, respectively).
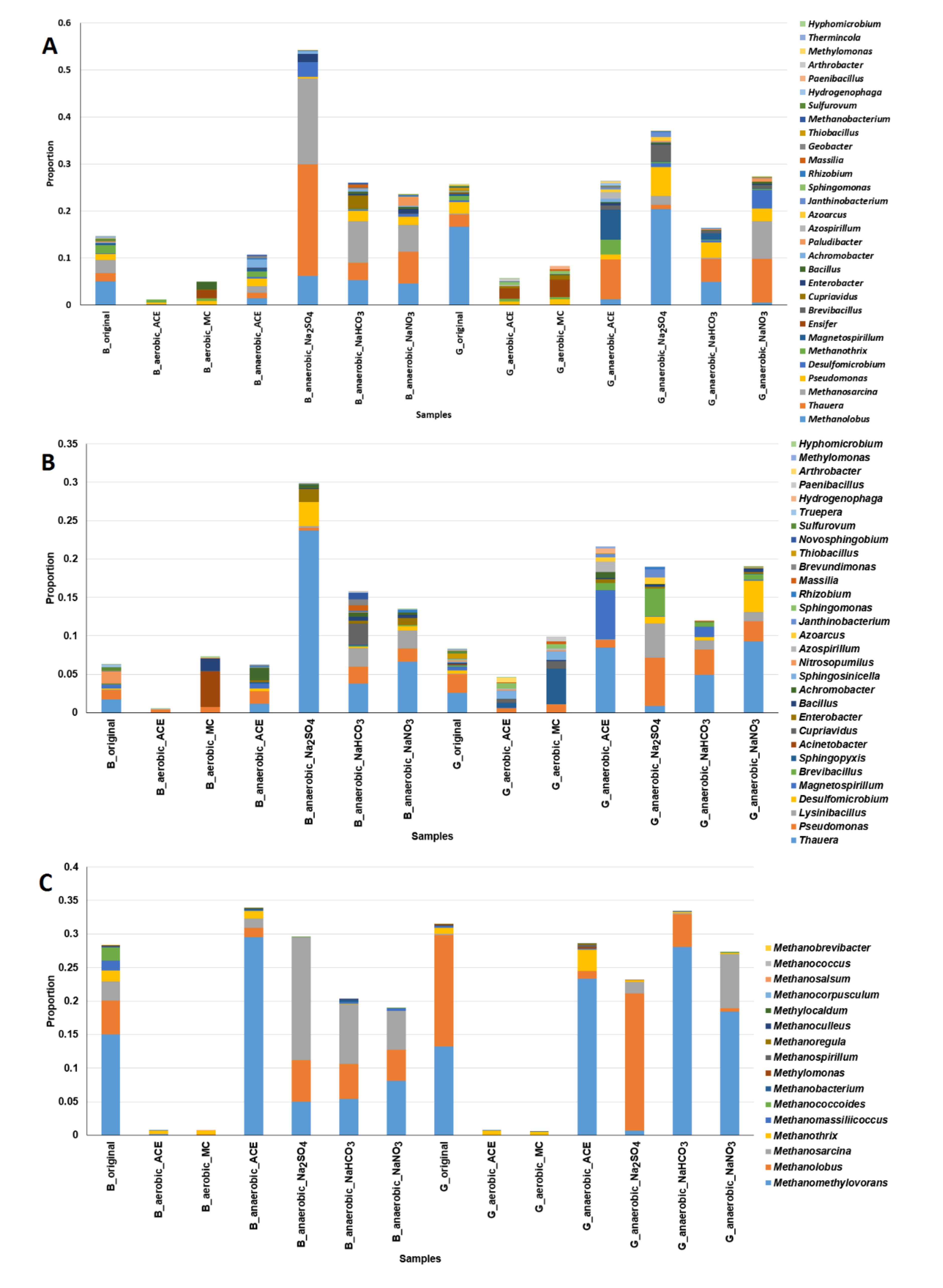
The microbial communities associated with the sediments supplemented with NaNO3, Na2SO4 or NaHCO3 are shown in Figure 8. For the Bali sediments, the overall proportions of nitrogen metabolism bacteria (Figure 8A), sulfate–sulfur metabolism bacteria (Figure 8B) and methanogens (Figure 8C) observed under anaerobic conditions were greater than those detected under aerobic conditions (two proportion test, p < 0.05). Similar results were observed for the Guandu sediments.
Furthermore, for the Bali sediments, the overall proportions of microbes associated with aromatic compound degradation under anaerobic conditions were greater than observed under aerobic conditions (multiproportion test, p < 0.05) (Figure 9). Similar results were observed for the Guandu sediments.
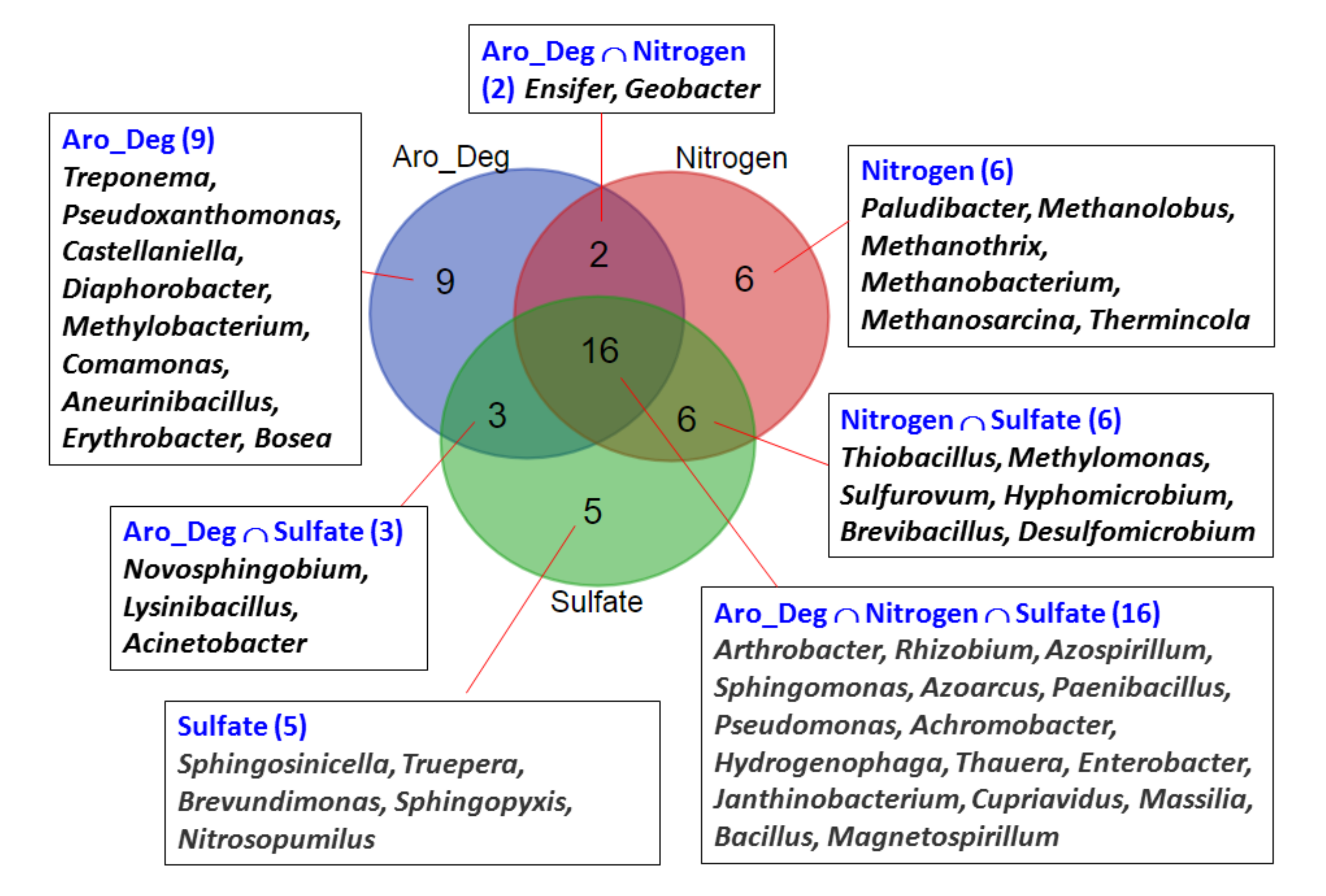
The results presented in Figure 5 indicate that the addition of NaNO3 or Na2SO4 had better ACE degradation effects than the addition of NaHCO3 in both the Bali and Guandu sediments. To identify common and differential microbial genera associated with ACE degradation in the sediments with different additives, a Venn diagram analysis was performed. In Figure 10, “Aro_Deg” indicates the aromatic compound-degrading bacteria, while “Nitrogen” or “Sulfate” indicate the nitrogen or sulfate–sulfur metabolism bacteria in the sediments supplemented with NaNO3 or Na2SO4, respectively.
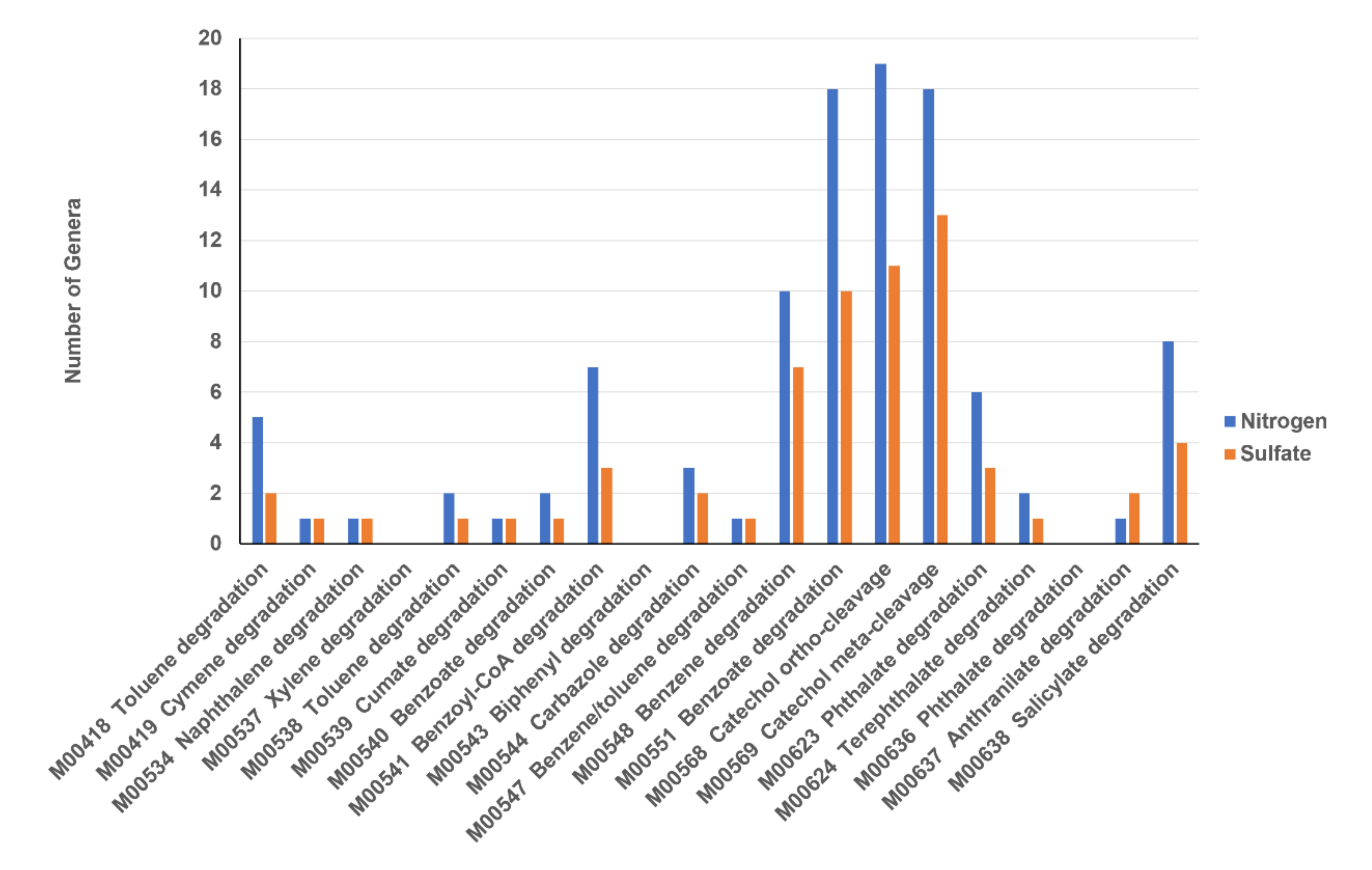
Sixteen microbial genera (Arthrobacter, Rhizobium, Azospirillum, Sphingomonas, Azoarcus, Paenibacillus, Pseudomonas, Achromobacter, Hydrogenophaga, Thauera, Enterobacter, Janthinobacterium, Cupriavidus, Massilia, Bacillus and Magnetospirillum) were detected in all of the sediments supplemented with NaNO3 or Na2SO4. Four of the sixteen microbial genera (Bacillus, Hydrogenophaga, Pseudomonas and Sphingomonas) have been reported to be associated with ACE (and paracetamol compounds) degradation [14,35,36,37]. These results suggested that sixteen microbial genera may be involved in anaerobic ACE degradation in the sediments supplemented with NaNO3 or Na2SO4. Three of the sixteen microbial genera (Arthrobacter, Enterobacter and Bacillus) were identified among the ACE-degrading bacterial isolates. Another two bacterial genera (Ensifer and Geobacter) represent additional aromatic compound-degrading bacteria that were not sulfate–sulfur metabolism bacteria. Three bacterial genera (Novosphingobium, Lysinibacillus and Acinetobacter) represent additional aromatic compound-degrading bacteria that were not nitrogen metabolism bacteria. These five bacterial genera can explain the different effects of the addition of NaNO3 or Na2SO4 on ACE degradation in sediments observed in Figure 5. The distributions of number of major microbial genera with different aromatic compound degradation pathways are shown in Figure 11. Most of the microbial genera associated with six reaction modules, M00551 Benzoate degradation, benzoate => catechol/methyl benzoate => methyl catechol, M00568 Catechol ortho-cleavage, catechol => 3-oxoadipate, M00569 Catechol meta-cleavage, catechol => acetyl-CoA/4-methylcatechol => propionyl-CoA, M00548 Benzene degradation, benzene => catechol, M00541 Benzoyl-CoA degradation, benzoyl-CoA => 3-hydroxypimeloyl-CoA and M00638 Salicylate degradation, salicylate => gentisate. Moreover, more microbial genera associated with aromatic compound degradation were found in the sediment supplemented with NaNO3 than that of the sediment supplemented with Na2SO4. These results can explain the different effects of the addition of NaNO3 or Na2SO4 on ACE degradation in sediments observed in Figure 5.
The sampling sites of mangrove sediments in this study are mangrove (Kandelia obovata) swamps. The study of Weng et al. investigated nitrogen cycle in rhizosphere of Kandelia obovata under nitrogen input shown that the potential nitrification intensity increased 200%–1500% compared with control under ammonium addition. The potential denitrification intensity increased more than 200% under nitrate addition. The abundance of ammonia-oxidizing archaea, ammonia-oxidizing bacteria, nirK and nifH containing microbes increased around the area of Kandelia obovata rhizosphere. The nirS containing microbes were decreased. Total nitrogen in tissues of Kandelia obovata increased more than 200% [38]. Results of another study indicated that addition of NH4+ significantly increased the growth rate of Kandelia obovata in mesocosm experiments [39]. A study of a 10-year-old constructed mangrove wetland planted with Kandelia obovata shown that the abundance of nitrogen cycle related bacteria, including ammonifiers, nitrifiers and denitrifiers, in planted belts were higher than that in unplanted control [40]. A study by Jian et al. demonstrated that addition of Na2SO4 enhanced the acid-volatile sulfide, crystalline Fe and exchangeable P contents, prompted iron plaque formation on root surfaces of Kandelia obovata and increased P accumulation in plant tissues. Sulfate and Kandelia obovata root activities had a confederate influence on the spatial distribution and geochemical cycling of Fe and P in the sediments. They enhanced Kandelia obovata resistance to the rugged surroundings in the mangrove environment and can prompt Kandelia obovata growth [41]. Therefore, additions of nitrate and sulfate to enhance ACE degradation (proposed in this study) are suitable to be applied at mangrove wetlands composed of Kandelia obovata. Moreover, these methods may be useful in a constructed wetland composed of Kandelia obovata.
4. Conclusions
In this study, the feasibility of bioremediation strategies for ACE removal in mangrove sediments were tested. All of the treatments, addition of ACE-degrading bacterial strains, MC and electron acceptors (NaNO3, Na2SO4 or NaHCO3) can enhance the ACE biodegradation in mangrove sediments. Addition of NaNO3 exhibited the best effects for ACE biodegradation in mangrove sediments. Different treatments result in different bacterial communities and exhibited different ACE-degrading effectiveness. Sixteen microbial genera identified may be major microbes involved in anaerobic ACE degradation in mangrove sediments with electron acceptor addition. These results provide useful solutions for removal of ACE in mangrove sediments.
Supplementary Materials
The following are available online at https://www.mdpi.com/2071-1050/12/13/5410/s1. Figure S1: Mangrove sediment sampling sites Bali (25°09′48″ N 121°25′04″ E) and Guandu (25°07′21″ N 121°27′40″ E) in northern Taiwan.
Author Contributions
Data curation, C.-W.Y.; formal analysis, Y.-E.C.; funding acquisition, B.-V.C.; investigation, C.-W.Y.; methodology, Y.-E.C.; project administration, B.-V.C.; visualization, C.-W.Y.; writing—original draft, C.-W.Y. and B.-V.C.; writin—review & editing, C.-W.Y. and B.-V.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Ministry of Science and Technology, Republic of China, Taiwan (grant no. MOST 107-2313-B-031-002).
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Ouellet, M.; Percival, M.D. Acetaminophen inhibition of cyclooxygenase isoforms. Arch. Biochem. Biophys. 2001, 387, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.D.; Bartkow, M.; Blockwell, S.J.; Coleman, H.M.; Khan, S.J.; Lim, R.; McDonald, J.A.; Nice, H.; Nugegoda, D.; Pettigrove, V.; et al. A national survey of trace organic contaminants in Australian Rivers. J. Environ. Qual. 2014, 43, 1702–1712. [Google Scholar] [CrossRef] [PubMed]
- Campanha, M.B.; Awan, A.T.; De Sousa, D.N.R.; Grosseli, G.M.; Mozeto, A.A.; Fadini, P.S. A 3-year study on occurrence of emerging contaminants in an urban stream of So Paulo State of Southeast Brazil. Environ. Sci. Pollut. Res. 2015, 22, 7936–7947. [Google Scholar] [CrossRef]
- Tran, N.H.; Reinhard, M.; Khan, E.; Chen, H.; Nguyen, V.T.; Li, Y.; Goh, S.G.; Nguyen, Q.B.; Saeidi, N.; Gin, K.Y.H. Emerging contaminants in wastewater, stormwater runoff, and surface water: Application as chemical markers for diffuse sources. Sci. Total Environ. 2019, 676, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Zhang, M.; Yuan, S.; Feng, J.; Wang, Q.; Wang, W.; Hu, Z. Transformation of acetaminophen during water chlorination treatment: Kinetics and transformation products identification. Environ. Sci. Pollut. Res. Int. 2016, 23, 12303–12311. [Google Scholar] [CrossRef]
- Sun, C.; Dudley, S.; McGinnis, M.; Trumble, J.; Gan, J. Acetaminophen detoxification in cucumber plants via induction of glutathione S-transferases. Sci. Total Environ. 2019, 649, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Philips, P.J.; Smith, S.G.; Koplin, D.W.; Zaugg, S.D.; Buxton, H.T.; Furlong, E.T.; Esposito, K.; Stinson, B. Pharmaceutical formulation facilities as sources of opioids and other pharmaceuticals to wastewater treatment plant effluents. Environ. Sci. Technol. 2010, 44, 4910–4916. [Google Scholar] [CrossRef]
- Escher, B.I.; Baumgartner, R.; Koller, M.; Treyer, K.; Lienert, J.; McArdell, C.S. Environmental toxicology and risk assessment of pharmaceuticals from hospital wastewater. Water Res. 2011, 45, 75–92. [Google Scholar] [CrossRef] [Green Version]
- Bernard, D.; Pascaline, H.; Jeremie, J.J. Distribution and origin of hydrocarbons in sediments from lagoons with fringing mangrove communities. Mar. Pollut. Bull. 1996, 32, 734–739. [Google Scholar] [CrossRef]
- Chang, B.V.; Chang, I.T.; Yuan, S.Y. Anaerobic degradation of phenanthrene and pyrene in mangrove sediment. Bull. Environ. Contam. Toxicol. 2008, 80, 145–149. [Google Scholar] [CrossRef]
- Chang, B.V.; Lu, Z.J.; Yuan, S.Y. Anaerobic degradation of nonylphenol in subtropical mangrove sediments. J. Hazard. Mater. 2009, 165, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Tsai, L.L.; Chang, B.V. Fungi extracellular enzyme-containing microcapsules enhance degradation of sulfonamide antibiotics in mangrove sediments. Environ. Sci. Pollut. Res. 2017, 25, 10069–10079. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.W.; Tsai, L.L.; Chang, B.V. Anaerobic degradation of sulfamethoxazole in mangrove sediments. Sci. Total Environ. 2018, 634, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Żur, J.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Marchlewicz, A.; Guzik, U. Paracetamol—Toxicity and microbial utilization. Pseudomonas moorei KB4 as a case study for exploring degradation pathway. Chemosphere 2018, 20, 192–202. [Google Scholar]
- Joss, A.; Keller, E.; Alder, A.C.; Gobel, A.; McArdell, C.S.; Ternes, T.; Siegrist, H. Removal of pharmaceuticals and fragrances in biological wastewater treatment. Water Res. 2005, 39, 3139–3152. [Google Scholar] [CrossRef]
- Groning, J.; Held, C.; Garten, C.; Claussnitzer, U.; Kaschabek, S.R.; Schlomann, M. Transformation of diclofenac by the indigenous microflora of river sediments and identification of a major intermediate. Chemosphere 2007, 69, 509–516. [Google Scholar] [CrossRef]
- Ying, G.G.; Kookana, R.S. Degradation of five selected endocrine-disrupting chemicals in seawater and marine sediment. Environ. Sci. Technol. 2003, 37, 1256–1260. [Google Scholar] [CrossRef]
- Pepper, L.L.; Gerba, C.P.; Gentry, T.J. Environmental Microbiology, 3rd ed.; Elsevier BV: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hu, J.; Zhang, L.L.; Chen, J.M.; Liu, Y. Degradation of paracetamol by Pseudomonas aeruginosa strain HJ1012. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 2013, 48, 791–799. [Google Scholar] [CrossRef]
- De Gusseme, B.; Vanhaecke, L.; Verstraete, W.; Boon, N. Degradation of acetaminophen by Delftia tsuruhatensis and Pseudomonas aeruginosa in a membrane bioreactor. Water Res. 2011, 45, 1829–1837. [Google Scholar] [CrossRef]
- Li, X.; Lin, X.; Zhang, J.; Wu, Y.; Yin, R.; Feng, Y.; Wang, Y. Degradation of polycyclic aromatic hydrocarbons by crude extracts from spent mushroom substrate and its possible mechanisms. Curr. Microbiol. 2010, 60, 336–342. [Google Scholar] [CrossRef]
- Chang, B.V.; Ren, Y.L. Biodegradation of three tetracyclines in river sediment. Ecol. Eng. 2015, 75, 272–277. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Carvajal, L.C.; Sanz-Martín, J.L.; Barragán-Huerta, B.E. Biodegradation of organic pollutants in saline wastewater by halophilic microorganisms: A review. Environ. Sci. Pollut. Res. 2014, 21, 9578–9588. [Google Scholar] [CrossRef]
- Tam, N.F.; Guo, C.L.; Yau, W.Y.; Wong, Y.S. Preliminary study on biodegradation of phenanthrene by bacteria isolated from mangrove sediments in Hong Kong. Mar. Pollut. Bull. 2002, 45, 316–324. [Google Scholar] [CrossRef]
- Varma, S.S.; Lakshmi, M.B.; Rajagopal, P.; Velan, M. Degradation of total petroleum hydrocarbon (TPH) in contaminated soil using Bacillus pumilus MVSV3. Biocontrol. Sci. 2017, 22, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanapiah, M.; Zulkifli, S.Z.; Mustafa, M.; Mohamat-Yusuff, F.; Ismail, A. Isolation, characterization, and identification of potential diuron-degrading bacteria from surface sediments of Port Klang, Malaysia. Mar. Pollu. Bull. 2018, 127, 453–457. [Google Scholar] [CrossRef]
- Zainith, S.; Purchase, D.; Saratale, G.D.; Ferreira, L.F.R.; Bilal, M.; Bharagava, R.N. Isolation and characterization of lignin-degrading bacterium Bacillus aryabhattai from pulp and paper mill wastewater and evaluation of its lignin-degrading potential. 3 Biotech. 2019, 9, 92. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.Y.; Zhao, J.C.; Xu, J.; Zhang, Y.; Zhang, T.Y.; Wu, Y.Y.; Zhang, Y.X. Arthrobacter ginkgonis sp. nov., an actinomycete isolated from rhizosphere of Ginkgo biloba L. Int. J. Syst. Evol. Microbiol. 2017, 67, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Aissaoui, S.; Ouled-Haddar, H.; Sifour, M.; Harrouche, K.; Sghaier, H. Metabolic and co-metabolic transformation of diclofenac by Enterobacter hormaechei D15 isolated from activated sludge. Curr. Microbiol. 2017, 74, 381–388. [Google Scholar] [CrossRef]
- Zhong, G.F.; Wang, F.F.; Sun, J.H.; Ye, J.B.; Mao, D.B.; Ma, K.; Yang, X.P. Bioconversion of lutein by Enterobacter hormaechei to form a new compound, 8-methyl-alpha-ionone. Biotech. Lett. 2017, 39, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.J.; Dodge, C.J.; Meinken, G.E. Biotransformation of pertechnetate by Clostridia. Radiochim. Acta 2009, 90, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Sari, I.P.; Simarani, K. Decolorization of selected azo dye by Lysinibacillus fusiformis W1B6: Biodegradation optimization, isotherm, and kinetic study biosorption mechanism. Adsorp. Sci. Technol. 2019, 37, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.J.; Chang, Y.H.; Chao, W.L.; Jane, W.N.; Chang, Y.T. Effect of hydraulic retention time on electricity generation using a solid plain-graphite plate microbial fuel cell anoxic/oxic process for treating pharmaceutical sewage. J. Environ. Sci. Health Pert A 2018, 53, 1185–1197. [Google Scholar] [CrossRef]
- Chang, Y.T.; Yang, C.W.; Chang, Y.J.; Chang, T.C.; Wei, D.J. The treatment of PPCP-containing sewage in an anoxic/aerobic reactor coupled with a novel design of solid plain graphite-plates microbial fuel cell. Biomed. Res. Int. 2014, 2014, 765652. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, J.; Zhu, R.; Zhou, Q.; Chen, J. Degradation of paracetamol by pure bacterial cultures and their microbial consortium. Appl. Microbiol. Biotechnol. 2013, 97, 3687–3698. [Google Scholar] [CrossRef]
- Weng, B.; Xie, X.; Yang, J.; Liu, J.; Lu, H.; Yan, C. Research on the nitrogen cycle in rhizosphere of Kandelia obovata under ammonium and nitrate addition. Mar. Pollut. Bull. 2013, 76, 227–240. [Google Scholar] [CrossRef]
- Cui, X.; Song, W.; Feng, J.; Jia, D.; Guo, J.; Wang, Z.; Wu, H.; Qi, F.; Liang, J.; Lin, G. Increased nitrogen input enhances Kandelia obovata seedling growth in the presence of invasive Spartina alterniflora in subtropical regions of China. Biol. Lett. 2017, 13, 20160760. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Tam, N.F.Y.; Zan, Q.; Cheung, S.G.; Shin, P.K.S.; Wong, Y.S.; Zhang, L.; Chen, Z. Performance and bacterial community structure of a 10-years old constructed mangrove wetland. Mar. Pollut. Bull. 2017, 124, 1096–1105. [Google Scholar] [CrossRef]
- Jian, L.; Junyi, Y.; Jingchun, L.; Chongling, Y.; Haoliang, L.; Spencer, K.L. The effects of sulfur amendments on the geochemistry of sulfur, phosphorus and iron in the mangrove plant (Kandelia obovata (S. L.)) rhizosphere. Mar. Pollut. Bull. 2017, 114, 733–741. [Google Scholar] [CrossRef]
Figure 1.
Aerobic degradation of acetaminophen with MC in non-acetaminophen (ACE)-adapted sediment (A) and ACE-adapted sediment; (B) Guandu: Guandu sediment. Bali: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. MC: enzyme extract-containing microcapsules. Data from three independent experiments are presented as the means ± SE.
Figure 1.
Aerobic degradation of acetaminophen with MC in non-acetaminophen (ACE)-adapted sediment (A) and ACE-adapted sediment; (B) Guandu: Guandu sediment. Bali: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. MC: enzyme extract-containing microcapsules. Data from three independent experiments are presented as the means ± SE.

Figure 2.
Comparison of the acetaminophen degradation abilities of the isolated bacterial strains. (A) Bacterial strains PF1, PF3, SC and SEC (without sediments) under aerobic conditions; (B) bacterial strains E, G and J (without sediments) under anaerobic conditions. Sterile: sterilized medium without the addition of bacteria. GS: Guandu sediment. BS: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Data from three independent experiments are presented as the means ± SE.
Figure 2.
Comparison of the acetaminophen degradation abilities of the isolated bacterial strains. (A) Bacterial strains PF1, PF3, SC and SEC (without sediments) under aerobic conditions; (B) bacterial strains E, G and J (without sediments) under anaerobic conditions. Sterile: sterilized medium without the addition of bacteria. GS: Guandu sediment. BS: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Data from three independent experiments are presented as the means ± SE.

Figure 3.
Phylogenetic analysis of 16S rRNA sequences of the seven bacterial strains (PF1, PF3, SEC, SC, E, G and J). Seven isolated bacterial strains: PF1, PF3, SC, SEC, E, G and J.
Figure 3.
Phylogenetic analysis of 16S rRNA sequences of the seven bacterial strains (PF1, PF3, SEC, SC, E, G and J). Seven isolated bacterial strains: PF1, PF3, SC, SEC, E, G and J.

Figure 4.
Comparison of the acetaminophen degradation abilities of the isolated bacterial strains. (A) Bacterial strain SC (Arthrobacter ginkgonis) with sediments under aerobic conditions; (B) bacterial strain E (Enterobacter hormechei) with sediments under anaerobic conditions. Sterile: sterilized medium without the addition of bacteria. GS: Guandu sediment. BS: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Data from three independent experiments are presented as the means ± SE.
Figure 4.
Comparison of the acetaminophen degradation abilities of the isolated bacterial strains. (A) Bacterial strain SC (Arthrobacter ginkgonis) with sediments under aerobic conditions; (B) bacterial strain E (Enterobacter hormechei) with sediments under anaerobic conditions. Sterile: sterilized medium without the addition of bacteria. GS: Guandu sediment. BS: Bali sediment. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Data from three independent experiments are presented as the means ± SE.

Figure 5.
Anaerobic degradation of acetaminophen in (A) Guandu and (B) Bali non-ACE-adapted sediments and in (C) Guandu and (D) Bali ACE-adapted sediments. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Control: sediment of the sampling site. Data from three independent experiments are presented as the means ± SE. NaNO3: addition of NaNO3 in sediment. Na2SO4: addition of Na2SO4 in sediment. NaHCO3: addition of NaHCO3 in sediment.
Figure 5.
Anaerobic degradation of acetaminophen in (A) Guandu and (B) Bali non-ACE-adapted sediments and in (C) Guandu and (D) Bali ACE-adapted sediments. GSterile: sterilized Guandu sediment. BSterile: sterilized Bali sediment. Control: sediment of the sampling site. Data from three independent experiments are presented as the means ± SE. NaNO3: addition of NaNO3 in sediment. Na2SO4: addition of Na2SO4 in sediment. NaHCO3: addition of NaHCO3 in sediment.

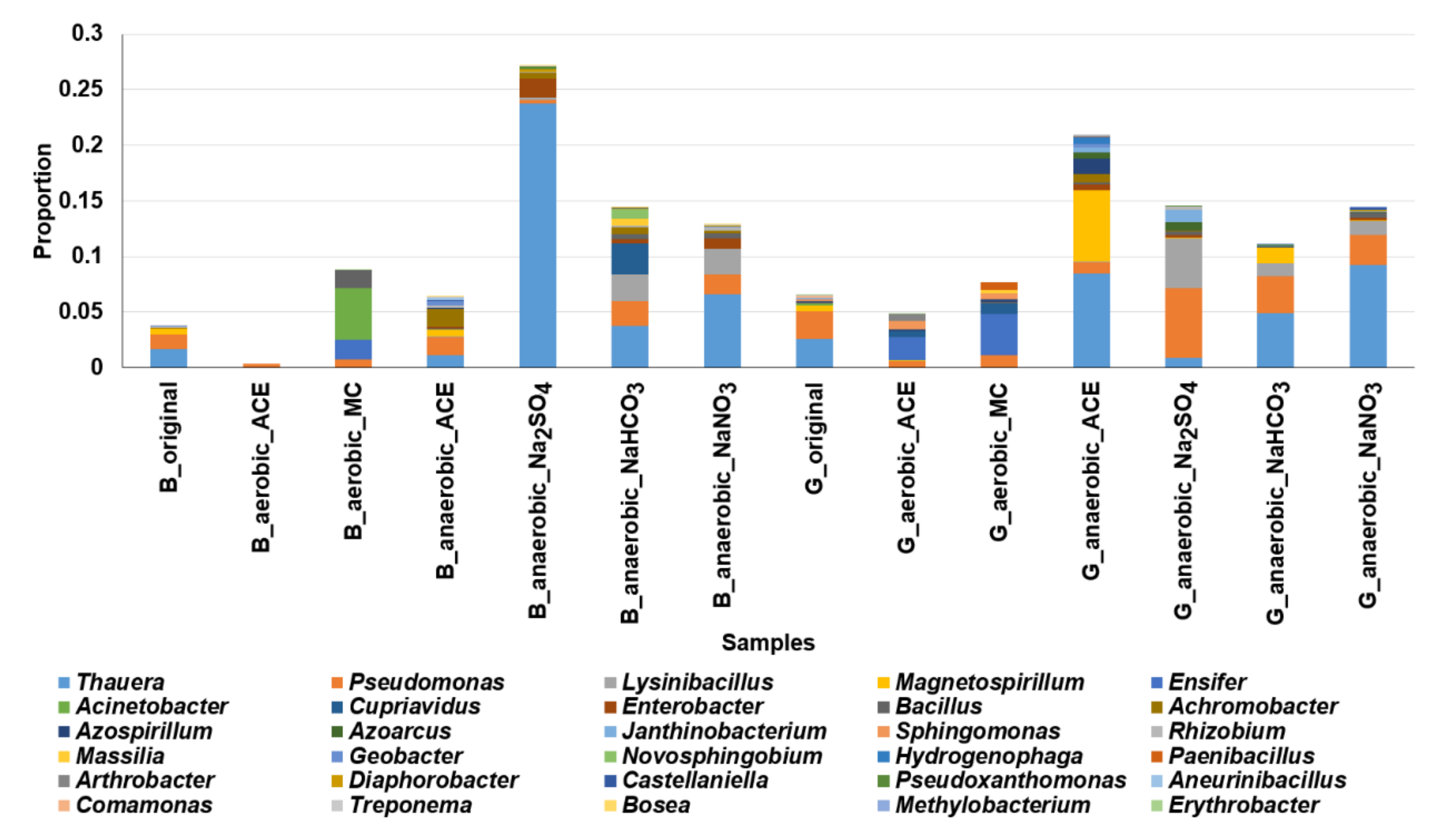
Figure 6.
Cluster analysis of microbial communities (at genus level) involved in aerobic and anaerobic degradation of acetaminophen in mangrove sediments. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.
Figure 6.
Cluster analysis of microbial communities (at genus level) involved in aerobic and anaerobic degradation of acetaminophen in mangrove sediments. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.

Figure 7.
Compositions of Bacteria and Archaea in anaerobic experiments. ORIGINAL: original sediment. ACE: sediments with ACE. Na2SO4: sediments with ACE + Na2SO4. NaHCO3: sediments with ACE + NaHCO3. NaNO3: sediments with ACE + NaNO3.
Figure 7.
Compositions of Bacteria and Archaea in anaerobic experiments. ORIGINAL: original sediment. ACE: sediments with ACE. Na2SO4: sediments with ACE + Na2SO4. NaHCO3: sediments with ACE + NaHCO3. NaNO3: sediments with ACE + NaNO3.

Figure 8.
Microbial communities involved in the anaerobic degradation of acetaminophen in mangrove sediments supplemented with NaNO3, Na2SO4 and NaHCO3. (A) Major 30 microbial genera associated with nitrogen cycle; (B) major 30 microbial genera associated with sulfate cycle; (C) methanogens. The differences in microbial compositions between the aerobic and anaerobic experimental samples were assessed by the two-proportion test. The p-values of all of these tests are less than 0.05. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.
Figure 8.
Microbial communities involved in the anaerobic degradation of acetaminophen in mangrove sediments supplemented with NaNO3, Na2SO4 and NaHCO3. (A) Major 30 microbial genera associated with nitrogen cycle; (B) major 30 microbial genera associated with sulfate cycle; (C) methanogens. The differences in microbial compositions between the aerobic and anaerobic experimental samples were assessed by the two-proportion test. The p-values of all of these tests are less than 0.05. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.

Figure 9.
Major 30 microbial genera that have been reported to be involved in the degradation of aromatic compounds. The differences in the microbial compositions between the aerobic and anaerobic experimental samples were identified using the two-proportion test. The p-values of all of these tests are less than 0.05. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.
Figure 9.
Major 30 microbial genera that have been reported to be involved in the degradation of aromatic compounds. The differences in the microbial compositions between the aerobic and anaerobic experimental samples were identified using the two-proportion test. The p-values of all of these tests are less than 0.05. G: Guandu. B: Bali. Original: sediments without additives. aerobic_ACE: sediments with ACE under aerobic conditions. anaerobic_ACE: sediments with ACE under anaerobic conditions. aerobic_MC: aerobic with addition of spent mushroom compost. anaerobic_Na2SO4: anaerobic with addition of Na2SO4. anaerobic_NaNO3: anaerobic with addition of NaNO3. anaerobic_NaHCO3: anaerobic with addition of NaHCO3.

Figure 10.
Identification of common and differential microbial genera detected between the different experiments by Venn diagram analysis. Aro_Deg: Major 30 microbial genera identified in the experiments using electron acceptors, which are associated with aromatic compound degradation. Nitrogen: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with nitrogen metabolism. Sulfate: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with sulfate–sulfur metabolism.
Figure 10.
Identification of common and differential microbial genera detected between the different experiments by Venn diagram analysis. Aro_Deg: Major 30 microbial genera identified in the experiments using electron acceptors, which are associated with aromatic compound degradation. Nitrogen: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with nitrogen metabolism. Sulfate: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with sulfate–sulfur metabolism.

Figure 11.
Distributions of number of major microbial genera with different aromatic compound degradation pathways. Mxxxxx are KEGG database module ID. Nitrogen: 30 major microbial genera identified in the experiments using electron acceptors that are associated with nitrogen metabolism. Sulfate: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with sulfate–sulfur metabolism.
Figure 11.
Distributions of number of major microbial genera with different aromatic compound degradation pathways. Mxxxxx are KEGG database module ID. Nitrogen: 30 major microbial genera identified in the experiments using electron acceptors that are associated with nitrogen metabolism. Sulfate: Major 30 microbial genera identified in the experiments using electron acceptors that are associated with sulfate–sulfur metabolism.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sediments properties for the two sampling sites.
| Parameters | Guandu Sampling Sites | Bali Sampling Sites |
|---|---|---|
| Bacterial counts | (1.9 ± 0.2) × 106 CFU g−1 | (2.6 ± 0.3) × 105 CFU g−1 |
| Temperature | 18.1 ± 1.1 °C | 20.5 ± 1.9 °C |
| Salinities | 11.1‰ ± 0.3‰ | 16.5‰ ± 1.5‰ |
| TOC | 3.6 wt% ± 0.2 wt% | 2.5 wt% ± 0.4 wt% |
| ACE concentrations | 0.89 mg kg−1 | 0.45 mg kg−1 |
Table 2.
The 16S rRNA gene-sequence blast results of the seven bacterial strains.
| Strain | Name | Identity | Accession Number |
|---|---|---|---|
| PF1 | Bacillus pumilus strain NBRC 12092 | 1251/1255 (99%) | NR_112637 |
| PF3 | Bacillus aerius strain 24K | 1326/1326 (100%) | NR_118439 |
| SC | Arthrobacter ginkgonis strain SYP-A7299 | 1283/1302 (99%) | NR_156061 |
| SEC | Bacillus aryabhattai B8W22 | 1330/1330 (100%) | NR_115953 |
| E | Enterobacter hormechei subsp. xiangfangensis strain 10–17 | 1292/1299 (99%) | NR_126208 |
| G | Clostridium sphenoides JCM 1415 strain ATCC 19403 | 1260/1268 (99%) | NR_026409 |
| J | Lysinibacillus fusiformis strain NBRC15717 | 1301/1305 (99%) | NR_112569 |
Table 3.
Comparison of acetaminophen degradation half-lives (t1/2, day) using different treatments.
| Treatment | Guandu Sediment | Bali Sediment | ||
|---|---|---|---|---|
| Without Additives | With Additives | Without Additives | With Additives | |
| Aerobic conditions | ||||
| MC | 33.55 ± 1.67 | 3.24 ± 0.16 | 66.66 ± 3.33 | 6.25 ± 0.31 |
| Arthrobacter sp. | 33.55 ± 1.67 | 2.54 ± 0.13 | 66.66 ± 3.33 | 3.30 ± 0.17 |
| Anaerobic conditions | ||||
| Enterobacter sp. | 95.59 ± 4.63 | 2.62 ± 0.13 | 98.04 ± 4.90 | 3.52 ± 0.17 |
| NaNO3 | 95.59 ± 4.63 | 1.16 ± 0.06 | 98.04 ± 4.90 | 3.05 ± 0.15 |
| Na2SO4 | 95.59 ± 4.63 | 2.39 ± 0.12 | 98.04 ± 4.90 | 3.84 ± 0.19 |
| NaHCO3 | 95.59 ± 4.63 | 2.79 ± 0.14 | 98.04 ± 4.90 | 10.75 ± 0.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, C.-W.; Chen, Y.-E.; Chang, B.-V. Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment. Sustainability 2020, 12, 5410. https://doi.org/10.3390/su12135410
AMA Style
Yang C-W, Chen Y-E, Chang B-V. Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment. Sustainability. 2020; 12(13):5410. https://doi.org/10.3390/su12135410
Chicago/Turabian StyleYang, Chu-Wen, Yi-En Chen, and Bea-Ven Chang. 2020. "Microbial Communities Associated with Acetaminophen Biodegradation from Mangrove Sediment" Sustainability 12, no. 13: 5410. https://doi.org/10.3390/su12135410
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.