Symbiotic Co-Culture of Scenedesmus sp. and Azospirillum brasilense on N-Deficient Media with Biomass Production for Biofuels

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions and Inoculum Preparation
2.2. Experimental Design and Statistical Analysis
2.3. Growth and Biomass Analysis
2.3.1. Biomass Growth
2.3.2. Cell Size
2.3.3. Biomass Analysis
3. Results and Discussion
3.1. Growth Analysis
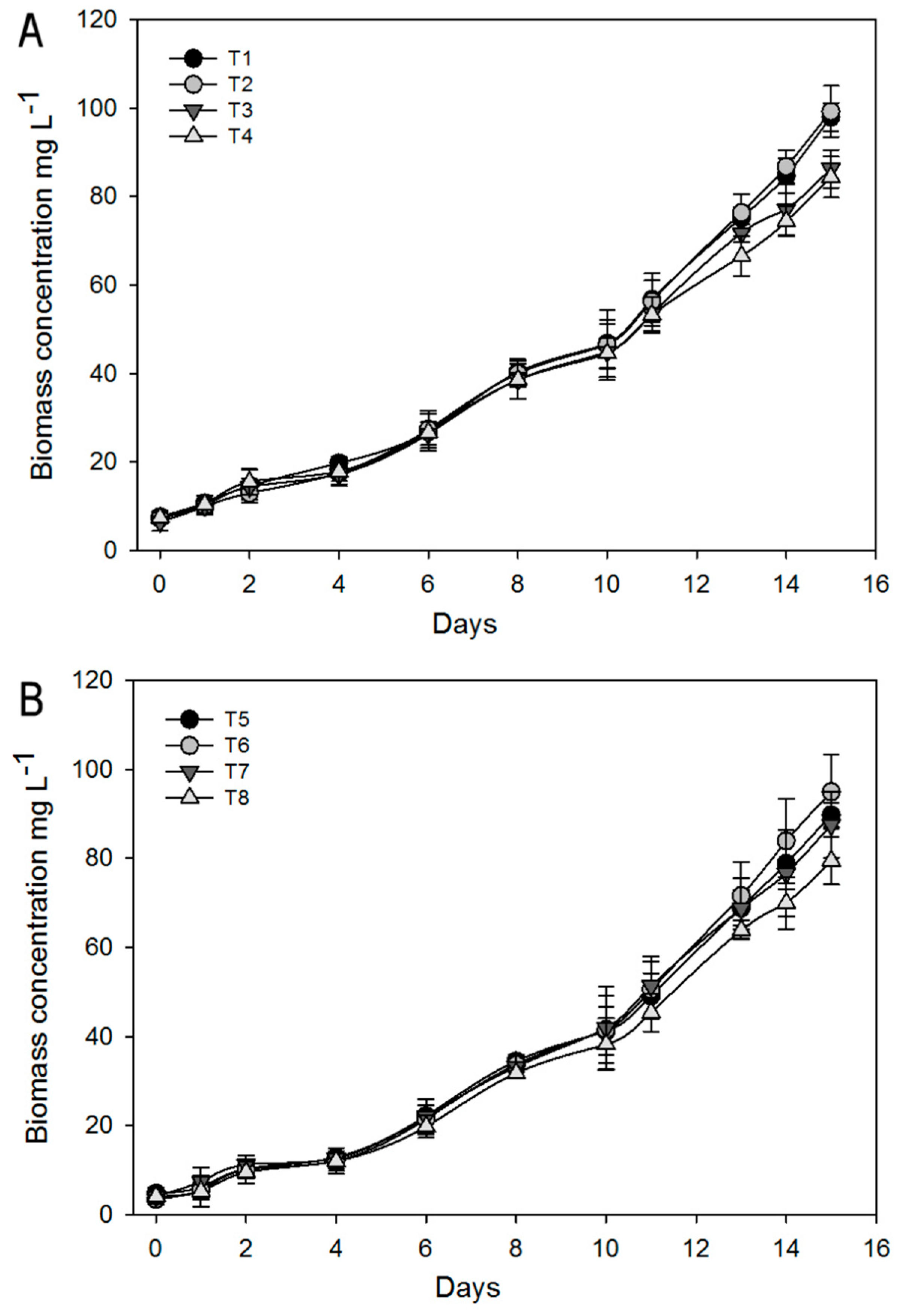
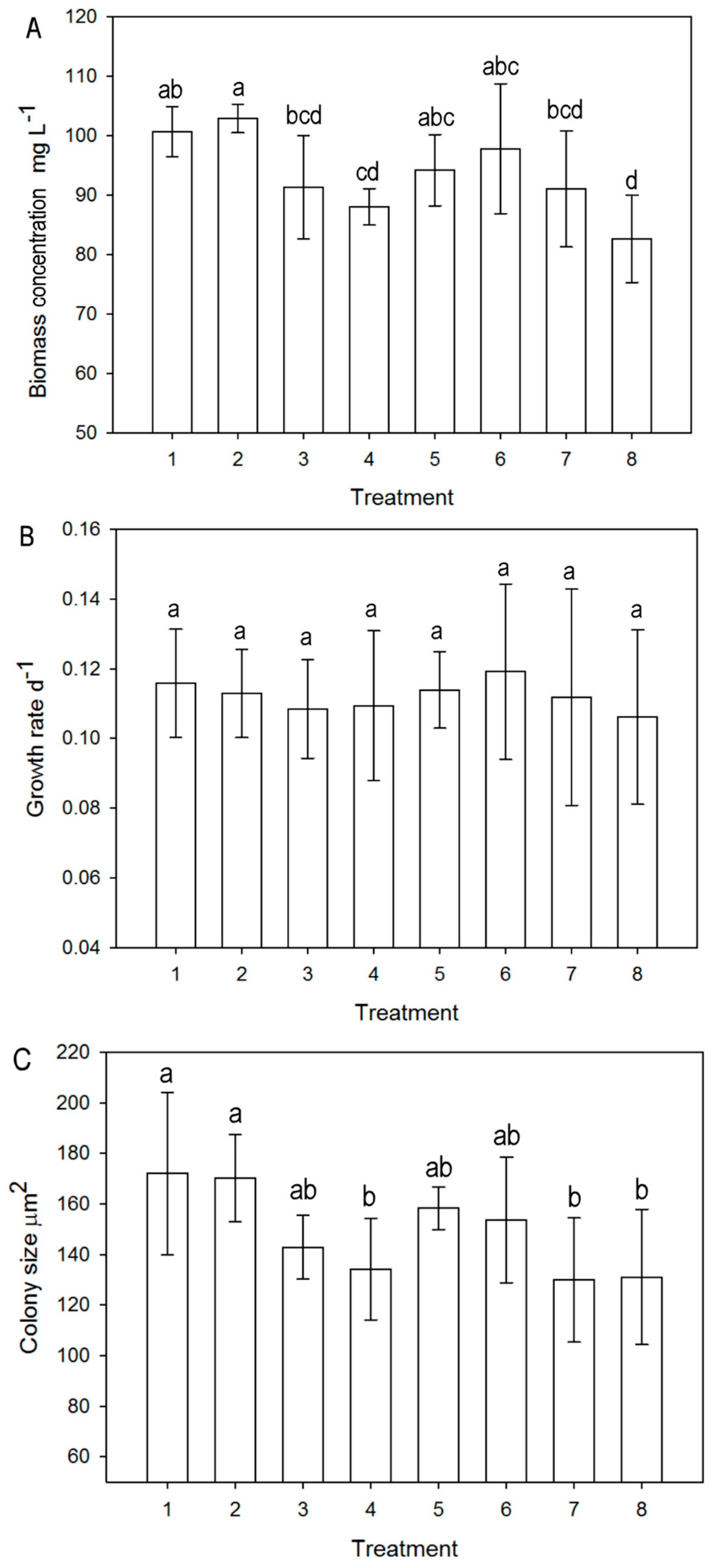
3.1.1. Biomass and Growth Analysis
3.1.2. Growth in terms of Colony Size
3.2. Analysis of Biomass Composition
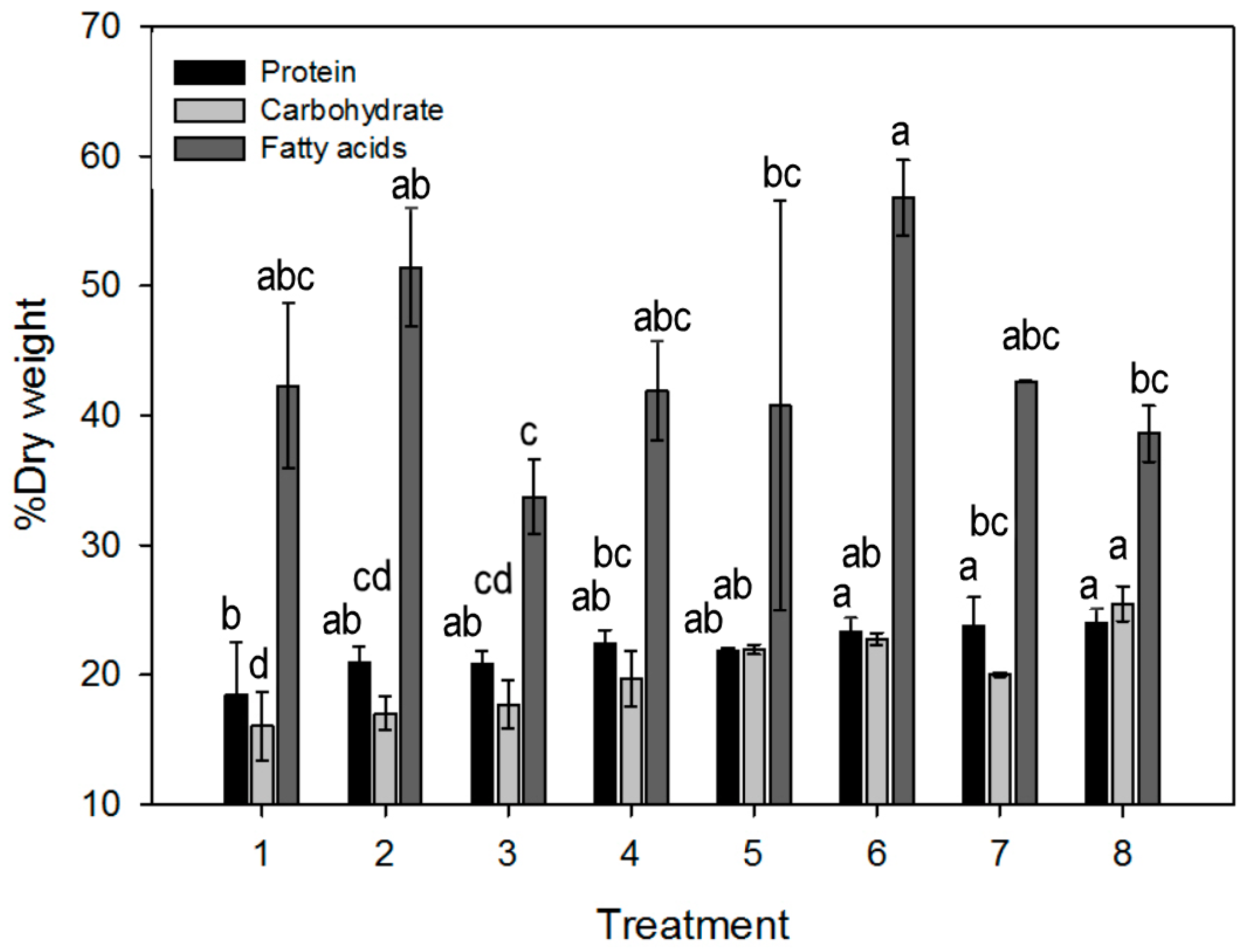
3.2.1. Protein Content
3.2.2. Carbohydrate Content
3.2.3. Fatty Acid Content and Profile
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mata, T.M.; Caetano, N.S.; Martins, A.A. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and purification of high-value metabolites from microalgae: Essential lipids, astaxanthin and phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L. Biorefinery as a promising approach to promote microalgae industry: An innovative framework. Renew. Sustain. Energy Rev. 2015, 41, 1376–1384. [Google Scholar] [CrossRef]
- Gupta, S.; Pawar, S.B. An integrated approach for microalgae cultivation using raw and anaerobic digested wastewaters from food processing industry. Bioresour. Technol. 2018, 269, 571–576. [Google Scholar] [CrossRef]
- Broderick, T.A.; Sherrard, J.H. Treatment of nutrient deficient wastewaters. J. Water Pollut. Control Fed. 1985, 57, 1178–1182. [Google Scholar]
- Esvelt, L.A.; Hart, H.H. Treatment of fruit processing waste by aeration. J. Water Pollut. Contr. Fed. 1970, 42, 1305–1326. [Google Scholar]
- Kargi, F.; Özmihçi, S. Improved biological treatment of nitrogen-deficient wastewater by incorporation of N2-fixing bacteria. Biotechnol. Lett. 2002, 24, 1281–1284. [Google Scholar] [CrossRef]
- Cuellar-Bermudez, S.P.; Aleman-Nava, G.S.; Chandra, R.; Garcia-Perez, J.S.; Contreras-Angulo, J.R.; Markou, G.; Muylaert, K.; Rittmann, B.E.; Parra-Saldivar, R. Nutrients utilization and contaminants removal. A review of two approaches of algae and cyanobacteria in wastewater. Algal Res. 2017, 24, 438–449. [Google Scholar] [CrossRef]
- Lananan, F.; Abdul Hamid, S.H.; Din, W.N.S.; Ali, N.; Khatoon, H.; Jusoh, A.; Endut, A. Symbiotic bioremediation of aquaculture wastewater in reducing ammonia and phosphorus utilizing effective microorganism (EM-1) and microalgae (Chlorella sp.). Int. Biodeterior. Biodegrad. 2014, 95, 127–134. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Hu, C. Culture modes and financial evaluation of two oleaginous microalgae for biodiesel production in desert area with open raceway pond. Bioresour. Technol. 2016, 218, 571–579. [Google Scholar] [CrossRef]
- Jorquera, O.; Kiperstok, A.; Sales, E.A.; Embiruçu, M.; Ghirardi, M.L. Comparative energy life-cycle analyses of microalgal biomass production in open ponds and photobioreactors. Bioresour. Technol. 2010, 101, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Muylaert, K.; Vandamme, D.; Foubert, I.; Brady, P.V. Harvesting of microalgae by means of flocculation. In Biomass and Biofuels from Microalgae; Springer International Publishing: Cham, Switzerland, 2015; pp. 251–273. [Google Scholar]
- Psycha, M.; Pyrgakis, K.; Kokossis, A.C. Process design analysis for the valorisation and selection of integrated micro-algae biorefineries. Comput. Aided Chem. Eng. 2014, 33, 1543–1548. [Google Scholar]
- Tijani, H.; Abdullah, N.; Yuzir, A. Integration of microalgae biomass in biomethanation systems. Renew. Sustain. Energy Rev. 2015, 52, 1610–1622. [Google Scholar] [CrossRef]
- Bai, X.; Lant, P.; Pratt, S. The contribution of bacteria to algal growth by carbon cycling. Biotechnol. Bioeng. 2015, 112, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Mujtaba, G.; Lee, K. Advanced treatment of wastewater using symbiotic co-culture of microalgae and bacteria. Appl. Chem. Eng. 2016, 27, 1–9. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Mayali, X.; Bebout, B.M.; Weber, P.K.; Detweiler, A.M.; Hernandez, J.-P.; Prufert-Bebout, L.; Bashan, Y. Establishment of stable synthetic mutualism without co-evolution between microalgae and bacteria demonstrated by mutual transfer of metabolites (NanoSIMS isotopic imaging) and persistent physical association (Fluorescent in situ hybridization). Algal Res. 2016, 15, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Plant Growth-promoting bacteria: Mechanisms and applications. Scientifica (Cairo) 2012, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Antoun, H.; Bashan, Y. Involvement of Indole-3-Acetic acid produced by the growth-promoting bacterium Azospirillum ssp. in promoting growth of Chlorella vulgaris. J. Phycol. 2008, 44, 938–947. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae–bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- BPDB: Bio-Pesticides DataBase, Azospirillum brasilense. University of Hertfordshire. Available online: http://sitem.herts.ac.uk/aeru/bpdb/Reports/1947.htm (accessed on 10 December 2018).
- Gonzalez, L.E.; Bashan, Y. Increased Growth of the Microalga Chlorella vulgaris when Coimmobilized and Cocultured in Alginate Beads with the Plant-Growth-Promoting Bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 2000, 66, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Bashan, Y.; Moreno, M.; Lebsky, V.K.; Bustillos, J.J. Increased pigment and lipid content, lipid variety, and cell and population size of the microalgae Chlorella spp. when co-immobilized in alginate beads with the microalgae-growth-promoting bacterium Azospirillum brasilense. Can. J. Microbiol. 2002, 48, 514–521. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Moreno, M.; Hernandez, J.-P.; Bashan, Y. Removal of ammonium and phosphorus ions from synthetic wastewater by the microalgae Chlorella vulgaris coimmobilized in alginate beads with the microalgae growth-promoting bacterium Azospirillum brasilense. Water Res. 2002, 36, 2941–2948. [Google Scholar] [CrossRef]
- Choix, F.J.; de-Bashan, L.E.; Bashan, Y. Enhanced accumulation of starch and total carbohydrates in alginate-immobilized Chlorella spp. induced by Azospirillum brasilense: II. Heterotrophic conditions. Enzyme Microb. Technol. 2012, 51, 300–309. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Hernandez, J.P.; Bashan, Y. Interaction of Azospirillum spp. with Microalgae: A Basic Eukaryotic–Prokaryotic Model and Its Biotechnological Applications. In Handbook for Azospirillum; Springer International Publishing: Cham, Switzerland, 2015; pp. 367–388. [Google Scholar]
- Ruiz-Güereca, D.A.; del Pilar Sánchez-Saavedra, M. Growth and phosphorus removal by Synechococcus elongatus co-immobilized in alginate beads with Azospirillum brasilense. J. Appl. Phycol. 2016, 28, 1501–1507. [Google Scholar] [CrossRef]
- Eustance, E.; Wray, J.T.; Badvipour, S.; Sommerfeld, M.R. The effects of cultivation depth, areal density, and nutrient level on lipid accumulation of Scenedesmus acutus in outdoor raceway ponds. J. Appl. Phycol. 2016, 28, 1459–1469. [Google Scholar] [CrossRef]
- Mata, T.M.; Melo, A.C.; Simões, M.; Caetano, N.S. Parametric study of a brewery effluent treatment by microalgae Scenedesmus obliquus. Bioresour. Technol. 2012, 107, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, C.; Morales, S.P.; Acosta, A. Scenedesmus sp. growth in different culture mediums for microalgal protein production. VitaeRevista La Fac. Quim. Farm. 2008, 15, 25–31. [Google Scholar]
- Shen, X.-F.; Liu, J.-J.; Chu, F.-F.; Lam, P.K.S.; Zeng, R.J. Enhancement of FAME productivity of Scenedesmus obliquus by combining nitrogen deficiency with sufficient phosphorus supply in heterotrophic cultivation. Appl. Energy 2015, 158, 348–354. [Google Scholar] [CrossRef]
- Mata, T.M.; Melo, A.C.; Meireles, S.; Mendes, A.M.; Martins, A.A.; Caetano, N.S. Potential of microalgae Scenedesmus obliquus grown in brewery wastewater for biodiesel production. Chem. Eng. Trans. 2013, 32. [Google Scholar] [CrossRef]
- Limón, P.; Malheiro, R.; Casal, S.; Acién-Fernández, F.G.; Fernández-Sevilla, J.M.; Rodrigues, N.; Cruz, R.; Bermejo, R.; Pereira, J.A. Improvement of stability and carotenoids fraction of virgin olive oils by addition of microalgae Scenedesmus almeriensis extracts. Food Chem. 2015, 175, 203–211. [Google Scholar] [CrossRef]
- Rengifo-Gallego, A.L.; Salamanca, E.J.P. Interaction Algae–Bacteria Consortia: A new application of heavy metals bioremediation. In Phytoremediation; Springer International Publishing: Cham, Switzerland, 2015; pp. 63–73. [Google Scholar]
- Tang, X.; He, L.Y.; Tao, X.Q.; Dang, Z.; Guo, C.L.; Lu, G.N.; Yi, X.Y. Construction of an artificial microalgal-bacterial consortium that efficiently degrades crude oil. J. Hazard. Mater. 2010, 181, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Burris, R.H. Regulation of nitrogenase activity by oxygen in Azospirillum brasilense and Azospirillum lipoferum. J. Bacteriol. 1987, 169, 944–948. [Google Scholar] [CrossRef]
- Hartmann, A. Ecophysiological aspects of growth and nitrogen fixation in Azospirillum spp. In Nitrogen Fixation with Non-Legumes; Springer: Dordrecht, The Netherlands, 1989; pp. 123–136. [Google Scholar]
- Stein, J.R. Handbook of Phycological Methods, Culture Methods and Growth Measurements; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Griffiths, M.J.; Garcin, C.; Van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an analytical procedure for extraction of lipids from microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Ruiz-Marin, A.; Mendoza-Espinosa, L.G.; Stephenson, T. Growth and nutrient removal in free and immobilized green algae in batch and semi-continuous cultures treating real wastewater. Bioresour. Technol. 2010, 101, 58–64. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Hernandez, J.-P.; Morey, T.; Bashan, Y. Microalgae growth-promoting bacteria as “helpers” for microalgae: A novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 2004, 38, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Yao, L.; Huang, Q. Aeration and mass transfer optimization in a rectangular airlift loop photobioreactor for the production of microalgae. Bioresour. Technol. 2015, 190, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, L.; Hu, H.; Ke, G.; Sun, Y. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Cabanelas, I.T.D.; Ruiz, J.; Arbib, Z.; Chinalia, F.A.; Garrido-Pérez, C.; Rogalla, F.; Nascimento, I.A.; Perales, J.A. Comparing the use of different domestic wastewaters for coupling microalgal production and nutrient removal. Bioresour. Technol. 2013, 131, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Tian-Yuan, Z.; Yin-Hu, W.; Shu-Feng, Z.; Feng-Min, L.; Hong-Ying, H. Isolation and heterotrophic cultivation of mixotrophic microalgae strains for domestic wastewater treatment and lipid production under dark condition. Bioresour. Technol. 2013, 149, 586–589. [Google Scholar]
- Meza, B.; de-Bashan, L.E.; Bashan, Y. Involvement of indole-3-acetic acid produced by Azospirillum brasilense in accumulating intracellular ammonium in Chlorella vulgaris. Res. Microbiol. 2015, 166, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Grobbelaar, J.U. Inorganic algal nutrition. In Handbook of Microalgal Culture; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 123–133. [Google Scholar]
- Beuckels, A.; Smolders, E.; Muylaert, K. Nitrogen availability influences phosphorus removal in microalgae-based wastewater treatment. Water Res. 2015, 77, 98–106. [Google Scholar] [CrossRef]
- Anand, J.; Arumugam, M. Enhanced lipid accumulation and biomass yield of Scenedesmus quadricauda under nitrogen starved condition. Bioresour. Technol. 2015, 188, 190–194. [Google Scholar] [CrossRef]
- Apandi, N.M.; Radin Mohamed, R.M.S.; Latiffi, N.A.A.; Rozlan, N.F.M.; Al-Gheethi, A.A.S. Protein and Lipid Content of Microalgae Scenedesmus sp. Biomass Grown in Wet Market Wastewater. MATEC Web Conf. 2017, 103, 06011. [Google Scholar] [CrossRef]
- De-Bashan, L.E.; Schmid, M.; Rothballer, M.; Hartmann, A.; Bashan, Y. Cell-cell interaction in the eukaryote-prokaryote model of the microalgae Chlorella vulgaris and the bacterium Azospirillum brasilense immobilized in polymer beads. J. Phycol. 2011, 47, 1350–1359. [Google Scholar] [CrossRef]
- Palacios, O.A.; Gomez-Anduro, G.; Bashan, Y.; De-Bashan, L.E. Tryptophan, thiamine and indole-3-acetic acid exchange between Chlorella sorokiniana and the plant growth-promoting bacterium Azospirillum brasilense. FEMS Microbiol. Ecol. 2016, 92, fiw077. [Google Scholar] [CrossRef]
- Ho, S.-H.; Chen, C.-Y.; Chang, J.-S. Effect of light intensity and nitrogen starvation on CO2 fixation and lipid/carbohydrate production of an indigenous microalga Scenedesmus obliquus CNW-N. Bioresour. Technol. 2012, 113, 244–252. [Google Scholar] [CrossRef]
- Yoo, C.; Jun, S.-Y.; Lee, J.-Y.; Ahn, C.-Y.; Oh, H.-M. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour. Technol. 2010, 101, S71–S74. [Google Scholar] [CrossRef]
- Kozlova, T.A.; Hardy, B.P.; Krishna, P.; Levin, D.B. Effect of phytohormones on growth and accumulation of pigments and fatty acids in the microalgae Scenedesmus quadricauda. Algal Res. 2017, 27, 325–334. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | A. brasilense | Mixing System | Nitrogen |
|---|---|---|---|
| 1 | Yes (+) | Aeration (+) | 41.17 mg·L−1 (+) |
| 2 | Yes (+) | Aeration (+) | 8.23 mg·L−1 (-) |
| 3 | Yes (+) | Magnetic stirred (-) | 41.17 mg·L 1 (+) |
| 4 | Yes (+) | Magnetic stirred (-) | 8.23 mg·L −1 (-) |
| 5 | No (-) | Aeration (+) | 41.17 mg·L−1 (+) |
| 6 | No (-) | Aeration (+) | 8.23 mg·L −1 (-) |
| 7 | No (-) | Magnetic stirred (-) | 41.17 mg·L−1 (+) |
| 8 | No (-) | Magnetic stirred (-) | 8.23 mg·L−1 (-) |
| Fatty Acid (FA) | Treatments | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| C14:0 | 0.3% ± 0.3 | 0.8% ± 0.1 | 0.5% ± 0.3 | 0.3% ± 0.1 | 0.4% ± 0.1 | 0.8% ± 0.3 | 0.6% ± 0.2 | 0.9% ± 0.3 |
| C16:0 | 4.1% ± 1.0 | 4.8% ± 0.6 | 5.0% ± 1.7 | 4.7% ± 0.7 | 4.9% ± 0.3 | 5.4% ± 0.4 | 5.7% ± 0.4 | 5.8% ± 0.3 |
| C16:1 | 0.6% ± 0.4 | 0.2% ± 0.1 | 0.7% ± 0.9 | 0.2% ± 0.1 | 0.6% ± 0.6 | 0.7% ± 0.7 | 0.6% ± 0.7 | 0.5% ± 0.4 |
| C18:1 | 2.0% ± 0.7 | 2.4% ± 0.3 | 3.5% ± 1.8 | 3.0% ± 0.2 | 2.4% ± 0.2 | 2.8% ± 0.4 | 3.5% ± 0.8 | 3.7% ± 1.1 |
| C18:2 | 2.9% ± 1.0 | 3.9% ± 0.5 | 4.8% ± 1.7 | 4.8% ± 0.4 | 3.7% ± 0.2 | 4.4% ± 0.1 | 5.4% ± 0.3 | 5.2% ± 0.2 |
| C18:3 n3 | 30.9% ± 3.1 | 37.2% ± 6.2 | 17.6% ± 3.6 | 27.4% ± 2.1 | 26.9% ± 15.9 | 40.5% ± 2.4 | 25.1% ± 1.9 | 20.6% ± 0.8 |
| Others | 1.5% ± 0.7 | 2.1% ± 0.2 | 1.6% ± 0.2 | 1.5% ± 0.3 | 1.8% ± 0.1 | 2.2% ± 0.3 | 1.7% ± 0.1 | 1.9% ± 0.4 |
| Total FA (%) | 42.3% ± 6.3 | 51.4%± 4.5 | 33.7%± 2.9 | 41.9%± 3.8 | 40.7% ± 15.8 | 56.8% ± 2.9 | 42.6% ± 0.1 | 38.6% ± 2.2 |
| SFA (%) | 10.3 | 10.9 | 16.5 | 11.9 | 13.2 | 10.9 | 14.8 | 17.2 |
| UFA (%) | 89.7 | 89.1 | 83.5 | 88.1 | 86.8 | 89.1 | 85.2 | 82.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Angulo, J.R.; Mata, T.M.; Cuellar-Bermudez, S.P.; Caetano, N.S.; Chandra, R.; Garcia-Perez, J.S.; Muylaert, K.; Parra-Saldivar, R. Symbiotic Co-Culture of Scenedesmus sp. and Azospirillum brasilense on N-Deficient Media with Biomass Production for Biofuels. Sustainability 2019, 11, 707. https://doi.org/10.3390/su11030707
Contreras-Angulo JR, Mata TM, Cuellar-Bermudez SP, Caetano NS, Chandra R, Garcia-Perez JS, Muylaert K, Parra-Saldivar R. Symbiotic Co-Culture of Scenedesmus sp. and Azospirillum brasilense on N-Deficient Media with Biomass Production for Biofuels. Sustainability. 2019; 11(3):707. https://doi.org/10.3390/su11030707
Chicago/Turabian StyleContreras-Angulo, Jose R., Teresa M. Mata, Sara P. Cuellar-Bermudez, Nidia S. Caetano, Rashmi Chandra, J. Saul Garcia-Perez, Koenraad Muylaert, and Roberto Parra-Saldivar. 2019. "Symbiotic Co-Culture of Scenedesmus sp. and Azospirillum brasilense on N-Deficient Media with Biomass Production for Biofuels" Sustainability 11, no. 3: 707. https://doi.org/10.3390/su11030707