
Semaglutide as a Possible Calmodulin Binder: Ligand-Based Computational Analyses and Relevance to Its Associated Reward and Appetitive Behaviour Actions
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
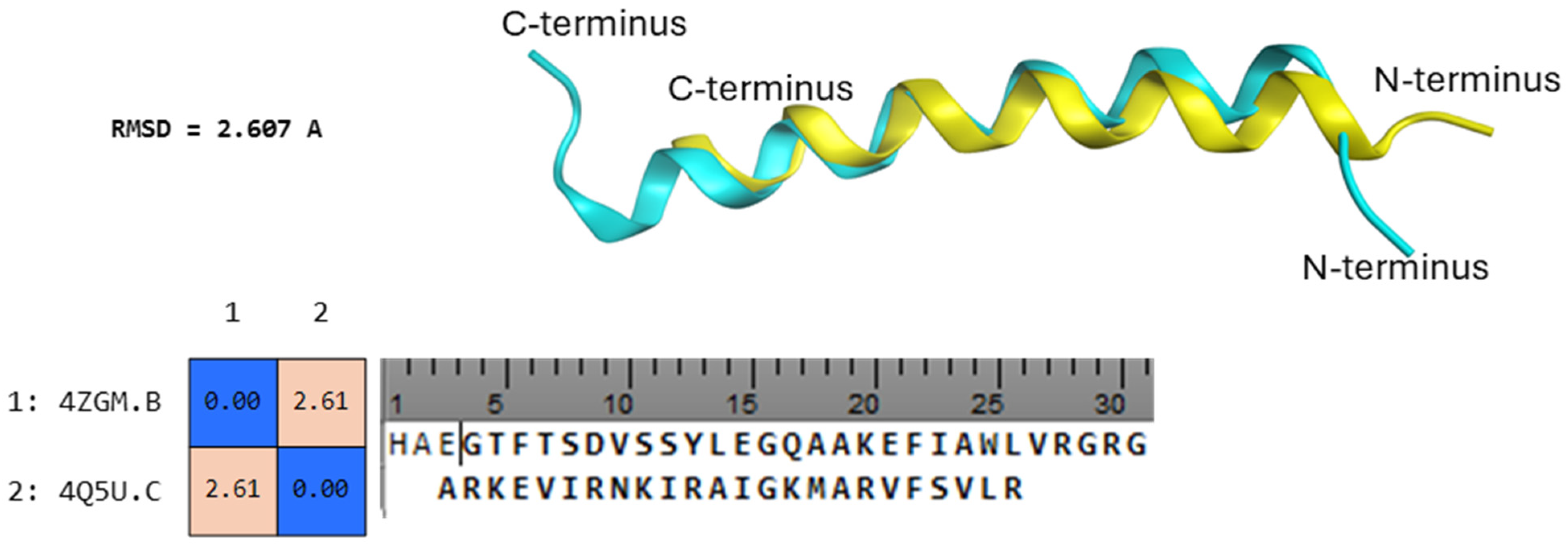
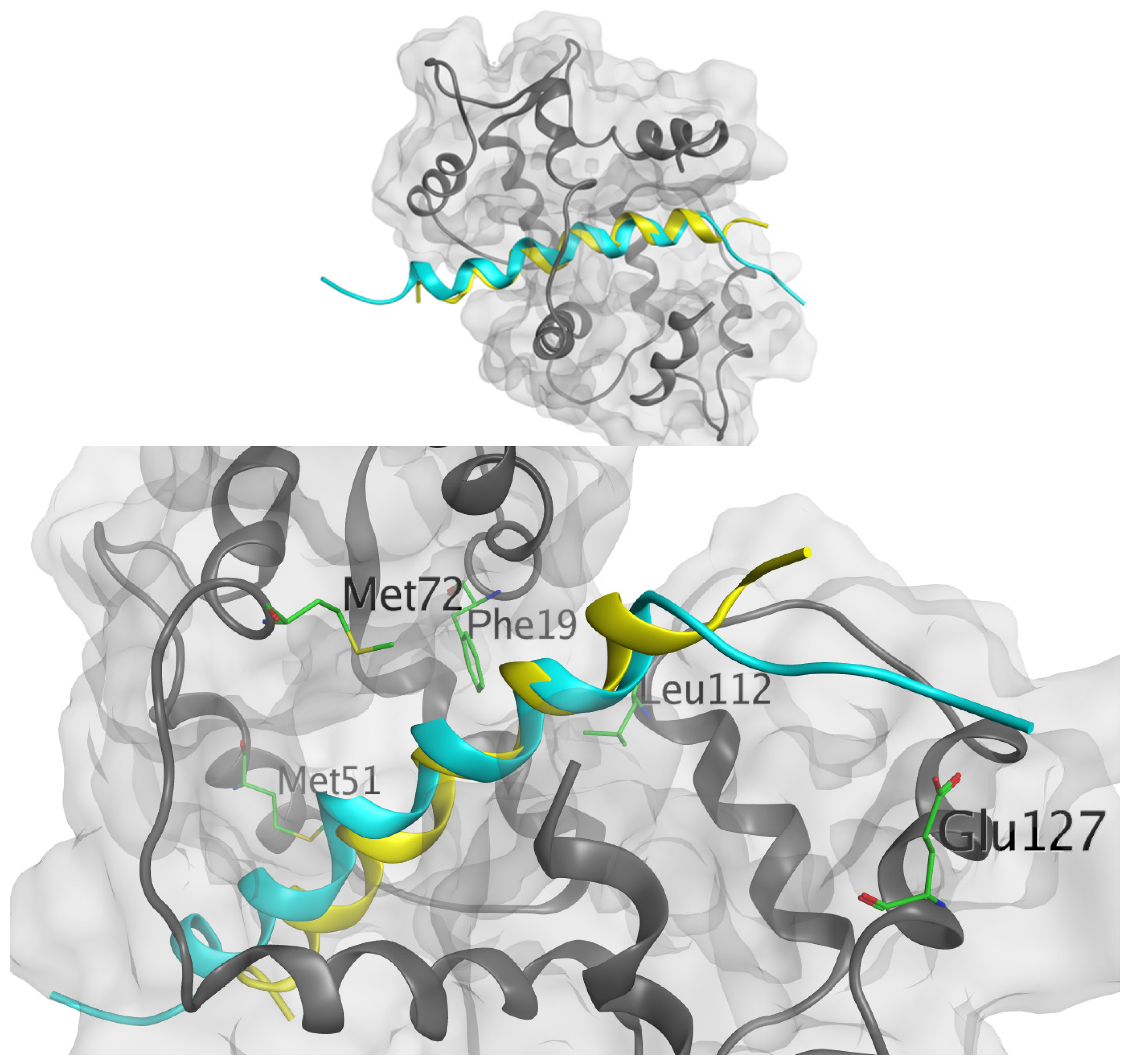
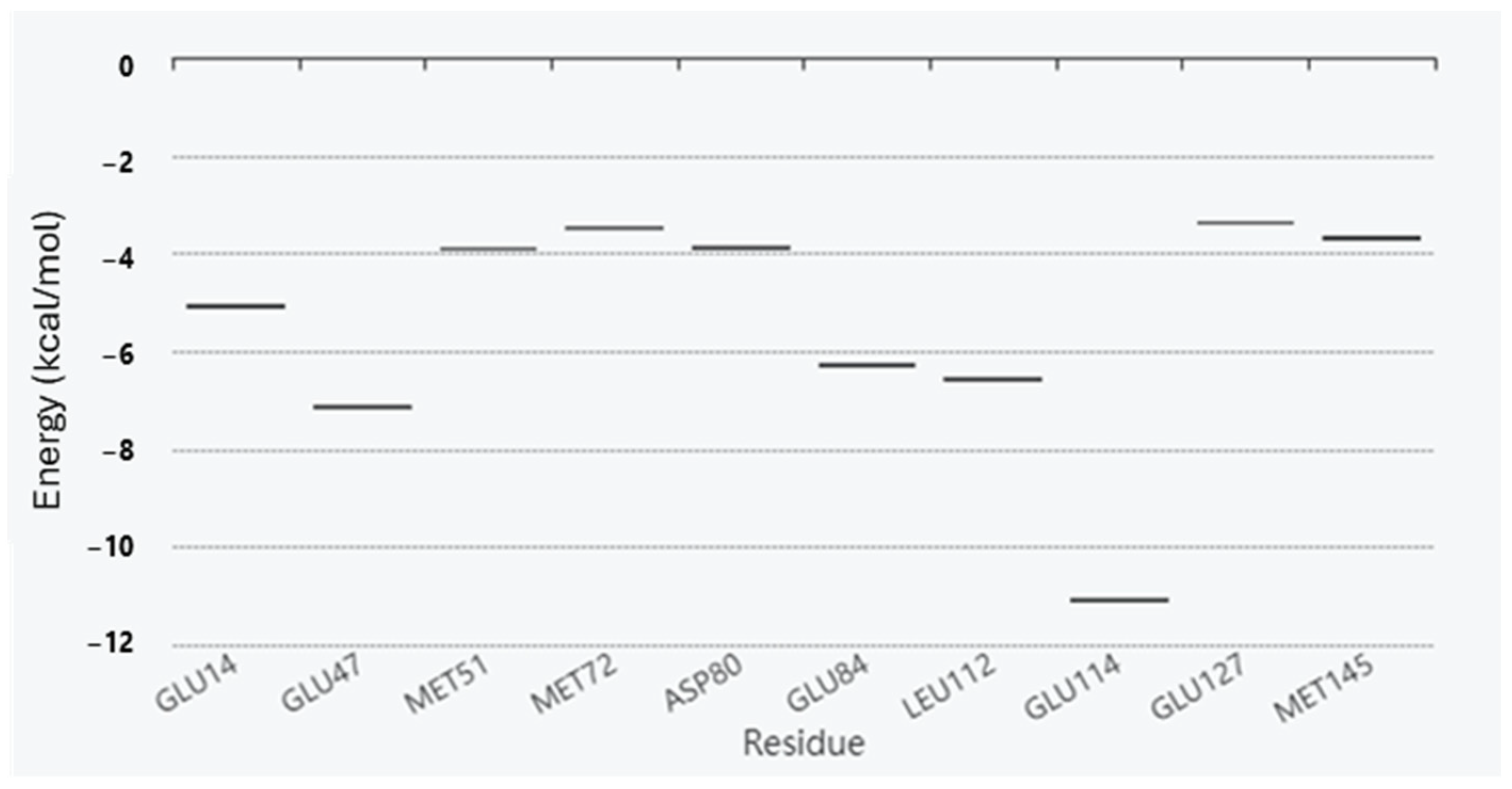
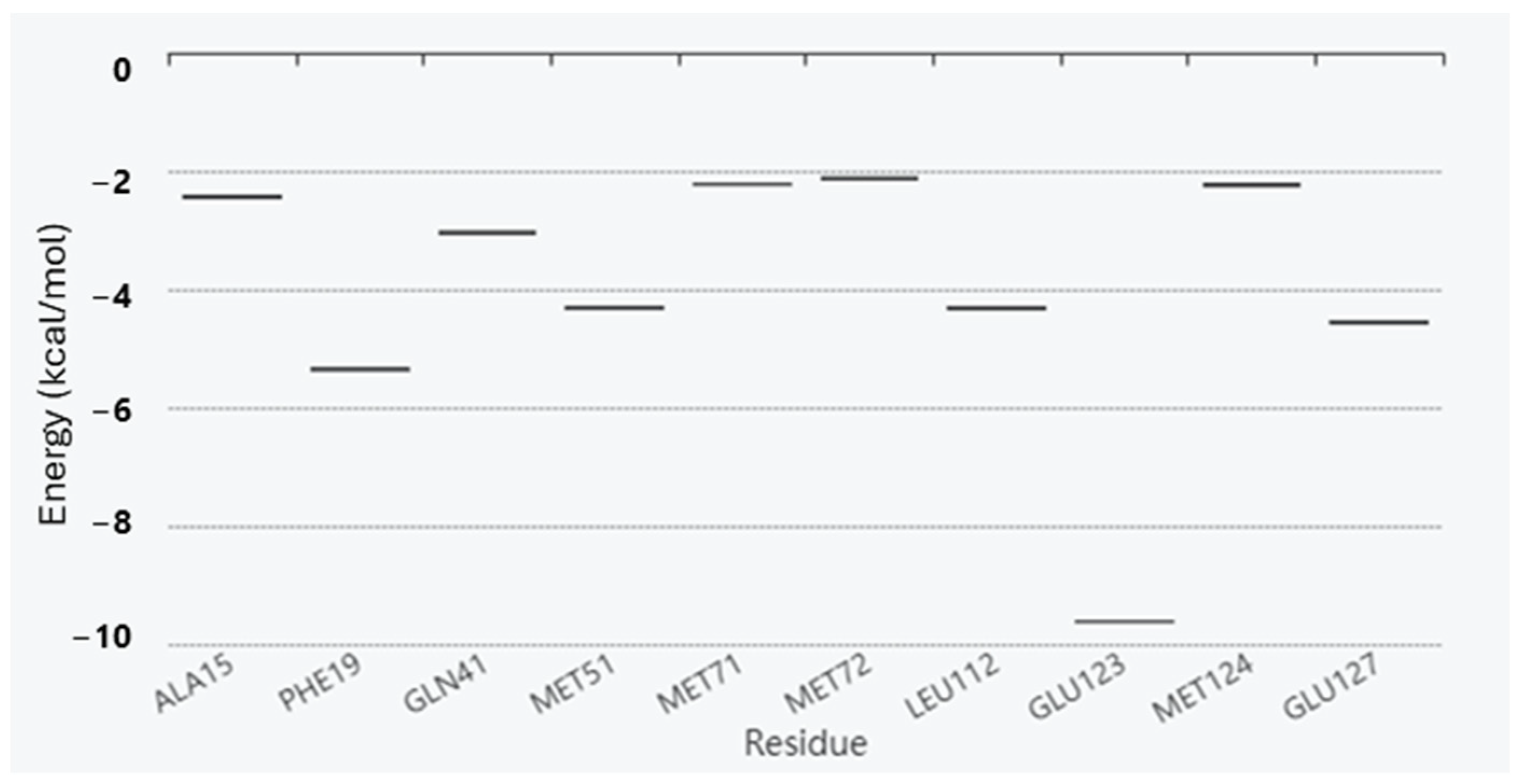
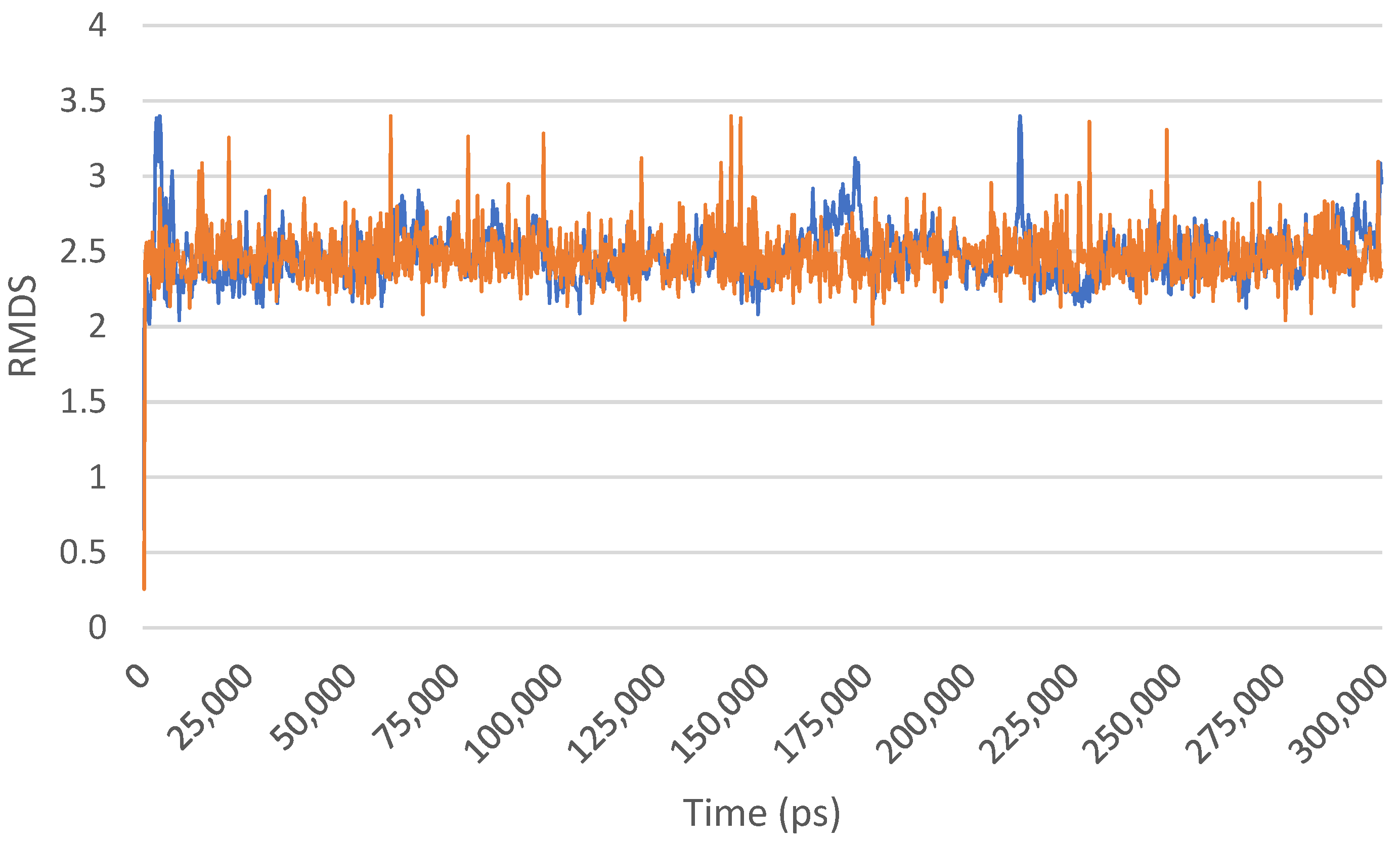
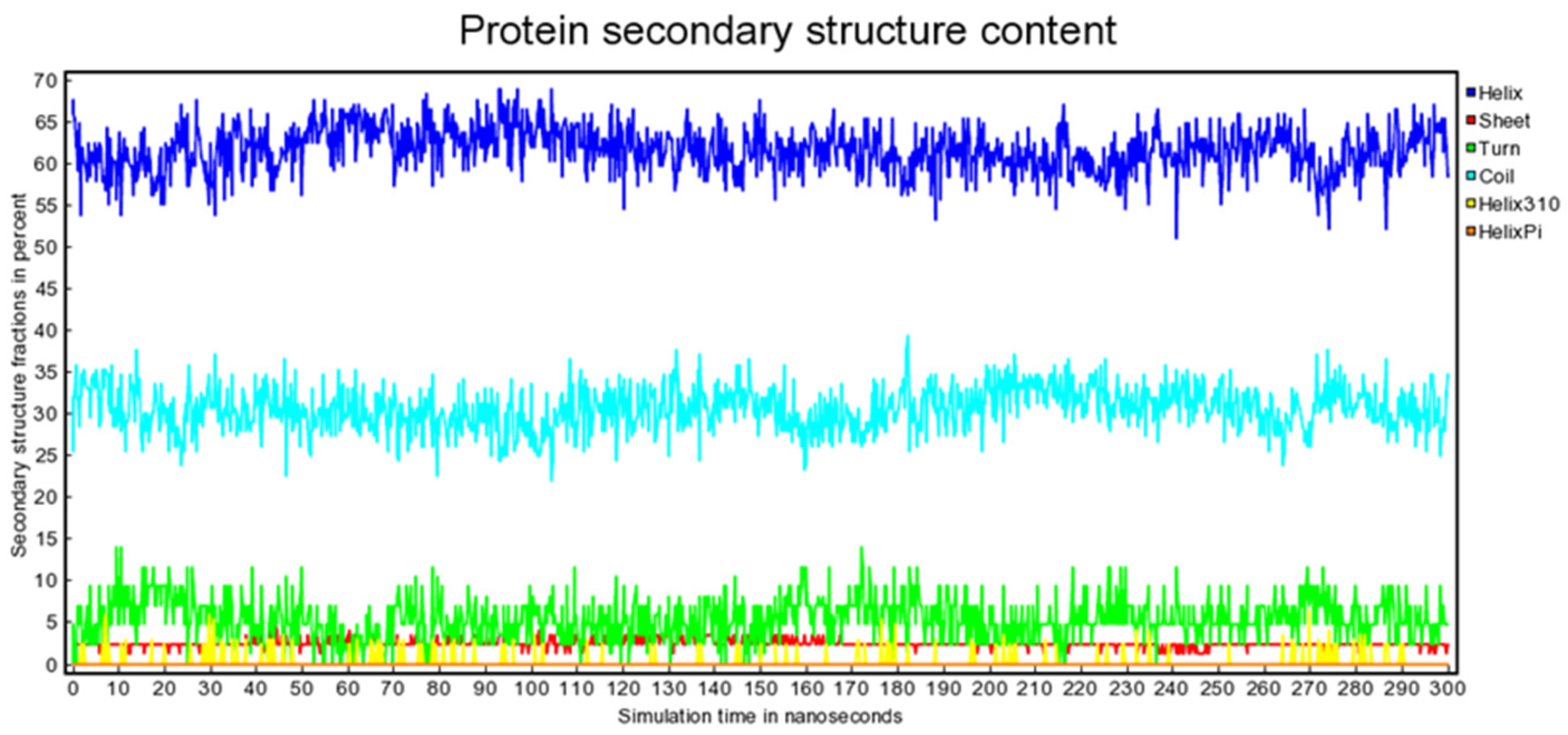
2. Results
3. Materials and Methods
4. Discussion
4.1. Semaglutide Actions on Reward and Appetitive Behaviour; Clinical Relevance of Current Study Findings—Semaglutide; DAergic Reward Pathways; and Related Weight Loss
4.2. Calmodulin Involvement in Both Drug and Natural Rewards
4.3. Semaglutide and Calmodulins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, X.; Hua, X.; Wang, X.; Xu, W.; Zhang, Y.; Shi, C.; Gu, M. Efficacy and safety of semaglutide on weight loss in obese or overweight patients without diabetes: A systematic review and meta-analysis of randomized controlled trials. Front. Pharmacol. 2022, 13, 935823. [Google Scholar] [CrossRef]
- Xie, Z.; Yang, S.; Deng, W.; Li, J.; Chen, J. Efficacy and Safety of Liraglutide and Semaglutide on Weight Loss in People with Obesity or Overweight: A Systematic Review. Clin. Epidemiol. 2022, 14, 1463–1476. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, M.; Wen, Z.; Lu, Z.; Cui, L.; Fu, C.; Xue, H.; Liu, Y.; Zhang, Y. GLP-1 Receptor Agonists: Beyond Their Pancreatic Effects. Front. Endocrinol. 2021, 12, 721135. [Google Scholar] [CrossRef] [PubMed]
- Floresta, G.; Amata, E.; Gentile, D.; Romeo, G.; Marrazzo, A.; Pittalà, V.; Salerno, L.; Rescifina, A. Fourfold Filtered Statistical/Computational Approach for the Identification of Imidazole Compounds as HO-1 Inhibitors from Natural Products. Mar. Drugs 2019, 17, 113. [Google Scholar] [CrossRef]
- Floresta, G.; Gentile, D.; Perrini, G.; Patamia, V.; Rescifina, A. Computational Tools in the Discovery of FABP4 Ligands: A Statistical and Molecular Modeling Approach. Mar. Drugs 2019, 17, 624. [Google Scholar] [CrossRef]
- Floresta, G.; Patamia, V.; Gentile, D.; Molteni, F.; Santamato, A.; Rescifina, A.; Vecchio, M. Repurposing of FDA-Approved Drugs for Treating Iatrogenic Botulism: A Paired 3D-QSAR/Docking Approach(†). ChemMedChem 2020, 15, 256–262. [Google Scholar] [CrossRef]
- Wen, Z.; He, J.; Tao, H.; Huang, S.Y. PepBDB: A comprehensive structural database of biological peptide-protein interactions. Bioinformatics 2019, 35, 175–177. [Google Scholar] [CrossRef]
- Hoeflich, K.P.; Ikura, M. Calmodulin in action: Diversity in target recognition and activation mechanisms. Cell 2002, 108, 739–742. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, K.T.; Degrado, W.F. How calmodulin binds its targets: Sequence independent recognition of amphiphilic α-helices. Trends Biochem Sci. 1990, 15, 59–64. [Google Scholar] [CrossRef]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef]
- Ataman, Z.A.; Gakhar, L.; Sorensen, B.R.; Hell, J.W.; Shea, M.A. The NMDA receptor NR1 C1 region bound to calmodulin: Structural insights into functional differences between homologous domains. Structure 2007, 15, 1603–1617. [Google Scholar] [CrossRef]
- Dunlap, T.B.; Guo, H.-F.; Cook, E.C.; Holbrook, E.; Rumi-Masante, J.; Lester, T.E.; Colbert, C.L.; Vander Kooi, C.W.; Creamer, T.P. Stoichiometry of the Calcineurin Regulatory Domain–Calmodulin Complex. Biochemistry 2014, 53, 5779–5790. [Google Scholar] [CrossRef]
- Cheng, A.; Best, S.A.; Merz, K.M., Jr.; Reynolds, C.H. GB/SA water model for the Merck molecular force field (MMFF). J. Mol. Graph. Model. 2000, 18, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J. Optimization of parameters for semiempirical methods IV: Extension of MNDO, AM1, and PM3 to more main group elements. J. Mol. Model. 2004, 10, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J. MOPAC: A semiempirical molecular orbital program. J. Comput. Aided Mol. Des. 1990, 4, 1–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Pan, H.; Sun, H.; Kang, Y.; Liu, H.; Cao, D.; Hou, T. fastDRH: A webserver to predict and analyze protein-ligand complexes based on molecular docking and MM/PB(GB)SA computation. Brief. Bioinform. 2022, 23, 201. [Google Scholar] [CrossRef]
- Cardullo, N.; Catinella, G.; Floresta, G.; Muccilli, V.; Rosselli, S.; Rescifina, A.; Bruno, M.; Tringali, C. Synthesis of Rosmarinic Acid Amides as Antioxidative and Hypoglycemic Agents. J. Nat. Prod. 2019, 82, 573–582. [Google Scholar] [CrossRef]
- Krieger, E.; Dunbrack, R.L.; Hooft, R.W.; Krieger, B. Assignment of protonation states in proteins and ligands: Combining pK a prediction with hydrogen bonding network optimization. In Computational Drug Discovery and Design; Springer: Berlin/Heidelberg, Germany, 2012; pp. 405–421. [Google Scholar]
- Krieger, E.; Nielsen, J.E.; Spronk, C.A.; Vriend, G. Fast empirical pKa prediction by Ewald summation. J. Mol. Graph. Model. 2006, 25, 481–486. [Google Scholar] [CrossRef]
- Maier, J.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Jakalian, A.; Jack, D.B.; Bayly, C.I. Fast, efficient generation of high-quality atomic charges. AM1-BCC model: II. Parameterization and validation. J. Comput. Chem. 2002, 23, 1623–1641. [Google Scholar] [CrossRef]
- Hornak, V.; Abel, R.; Okur, A.; Strockbine, B.; Roitberg, A.; Simmerling, C. Comparison of multiple Amber force fields and development of improved protein backbone parameters. Proteins Struct. Funct. Bioinform. 2006, 65, 712–725. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Fuochi, V.; Floresta, G.; Emma, R.; Patamia, V.; Caruso, M.; Zagni, C.; Ronchi, F.; Ronchi, C.; Drago, F.; Rescifina, A.; et al. Heparan Sulfate and Enoxaparin Interact at the Interface of the Spike Protein of HCoV-229E but Not with HCoV-OC43. Viruses 2023, 15, 663. [Google Scholar] [CrossRef]
- Crocetti, L.; Floresta, G.; Zagni, C.; Merugu, D.; Mazzacuva, F.; de Oliveira Silva, R.R.; Vergelli, C.; Giovannoni, M.P.; Cilibrizzi, A. Ligand Growing Experiments Suggested 4-amino and 4-ureido pyridazin-3(2H)-one as Novel Scaffold for FABP4 Inhibition. Pharmaceuticals 2022, 15, 1335. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M. Hot weight loss drugs tested against addiction. Science 2023, 381, 930–931. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Wen, S.; Zhou, L. The Relationship Between the Blood-Brain-Barrier and the Central Effects of Glucagon-Like Peptide-1 Receptor Agonists and Sodium-Glucose Cotransporter-2 Inhibitors. Diabetes Metab. Syndr. Obes. 2022, 15, 2583–2597. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, M.K.; Karuppasamy, M.; Sahoo, B.M. Semaglutide, a glucagon like peptide-1 receptor agonist with cardiovascular benefits for management of type 2 diabetes. Rev. Endocr. Metab. Disord. 2022, 23, 521–539. [Google Scholar] [CrossRef]
- Gabery, S.; Salinas, C.G.; Paulsen, S.J.; Ahnfelt-Ronne, J.; Alanentalo, T.; Baquero, A.F.; Buckley, S.T.; Farkas, E.; Fekete, C.; Frederiksen, K.S.; et al. Semaglutide lowers body weight in rodents via distributed neural pathways. JCI Insight 2020, 5, 133429. [Google Scholar] [CrossRef]
- Jones, B.; Buenaventura, T.; Kanda, N.; Chabosseau, P.; Owen, B.M.; Scott, R.; Goldin, R.; Angkathunyakul, N.; Corrêa, I.R., Jr.; Bosco, D.; et al. Targeting GLP-1 receptor trafficking to improve agonist efficacy. Nat. Commun. 2018, 9, 1602. [Google Scholar] [CrossRef] [PubMed]
- Rusnak, F.; Mertz, P. Calcineurin: Form and function. Physiol. Rev. 2000, 80, 1483–1521. [Google Scholar] [CrossRef]
- Novikoff, A.; O’Brien, S.L.; Bernecker, M.; Grandl, G.; Kleinert, M.; Knerr, P.J.; Stemmer, K.; Klingenspor, M.; Zeigerer, A.; DiMarchi, R.; et al. Spatiotemporal GLP-1 and GIP receptor signaling and trafficking/recycling dynamics induced by selected receptor mono- and dual-agonists. Mol. Metab. 2021, 49, 101181. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.G.; Kitasato, H.; Matsuura, H. Involvement of calmodulin in glucagon-like peptide 1(7-36) amide-induced inhibition of the ATP-sensitive K+ channel in mouse pancreatic beta-cells. Exp. Physiol. 2001, 86, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Le, T.; Xing, G.; Johnson, L.R.; Ursano, R.J. Analysis of kinase gene expression in the frontal cortex of suicide victims: Implications of fear and stress. Front. Behav. Neurosci. 2011, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, L.; Gan, K.; Pan, X.; Chen, S. Phosphorylated proteomics-based analysis of the effects of semaglutide on hippocampi of high-fat diet-induced-obese mice. Diabetol. Metab. Syndr. 2023, 15, 63. [Google Scholar] [CrossRef]
- Greenwood, M.P.; Greenwood, M.; Barez-Lopez, S.; Hawkins, J.W.; Short, K.; Tatovic, D.; Murphy, D. Osmoadaptive GLP-1R signalling in hypothalamic neurones inhibits antidiuretic hormone synthesis and release. Mol. Metab. 2023, 70, 101692. [Google Scholar] [CrossRef]
- Dawson, R.A.; Mac Neil, S. Mitogenic role for extracellular calmodulin-like activity in normal human umbilical vein endothelial cells. Br. J. Haematol. 1992, 82, 151–160. [Google Scholar] [CrossRef]
- Goberdhan, N.J.; Dawson, R.A.; Freedlander, E.; Mac Neil, S. A calmodulin-like protein as an extracellular mitogen for the keratinocyte. Br. J. Dermatol. 1993, 129, 678–688. [Google Scholar] [CrossRef]
- Woodward, B.J.; Lenton, E.A.; Mac Neil, S. Requirement of preimplantation human embryos for extracellular calmodulin for development. Hum. Reprod. 1993, 8, 272–276. [Google Scholar] [CrossRef]
- Patel, P.N.; Fox, C.K.; Bensignor, M.O.; Bomberg, E.M. Weight Loss from Combination Anti-Obesity Medication Regimens Can Approach that Achieved from Bariatric Surgery. JCEM Case Rep. 2023, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Gribble, F.M.; O’Rahilly, S. Obesity therapeutics: The end of the beginning. Cell Metab. 2021, 33, 705–706. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Kawahata, I.; Cheng, A.; Fukunaga, K. The Role of CaMKII and ERK Signaling in Addiction. Int. J. Mol. Sci. 2021, 22, 3189. [Google Scholar] [CrossRef] [PubMed]
- Di Chiara, G.; Acquas, E.; Tanda, G.; Cadoni, C. Drugs of abuse: Biochemical surrogates of specific aspects of natural reward? Biochem. Soc. Symp. 1993, 59, 65–81. [Google Scholar] [PubMed]
- Lutas, A.; Kucukdereli, H.; Alturkistani, O.; Carty, C.; Sugden, A.U.; Fernando, K.; Diaz, V.; Flores-Maldonado, V.; Andermann, M.L. State-specific gating of salient cues by midbrain dopaminergic input to basal amygdala. Nat. Neurosci. 2019, 22, 1820–1833. [Google Scholar] [CrossRef] [PubMed]
- Arillotta, D.; Floresta, G.; Guirguis, A.; Corkery, J.M.; Catalani, V.; Martinotti, G.; Sensi, S.L.; Schifano, F. GLP-1 Receptor Agonists and Related Mental Health Issues; Insights from a Range of Social Media Platforms Using a Mixed-Methods Approach. Brain Sci. 2023, 13, 1503. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Anefors, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. Dopamine signaling in the amygdala, increased by food ingestion and GLP-1, regulates feeding behavior. Physiol. Behav. 2014, 136, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Athauda, D.; Maclagan, K.; Skene, S.S.; Bajwa-Joseph, M.; Letchford, D.; Chowdhury, K.; Hibbert, S.; Budnik, N.; Zampedri, L.; Dickson, J.; et al. Exenatide once weekly versus placebo in Parkinson’s disease: A randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 1664–1675. [Google Scholar] [CrossRef]
- van Bloemendaal, L.; RG, I.J.; Ten Kulve, J.S.; Barkhof, F.; Konrad, R.J.; Drent, M.L.; Veltman, D.J.; Diamant, M. GLP-1 receptor activation modulates appetite- and reward-related brain areas in humans. Diabetes 2014, 63, 4186–4196. [Google Scholar] [CrossRef]
- Chiappini, S.; Vickers-Smith, R.; Harris, D.; Papanti Pelletier, G.D.; Corkery, J.M.; Guirguis, A.; Martinotti, G.; Sensi, S.L.; Schifano, F. Is There a Risk for Semaglutide Misuse? Focus on the Food and Drug Administration’s FDA Adverse Events Reporting System (FAERS) Pharmacovigilance Dataset. Pharmaceuticals 2023, 16, 994. [Google Scholar] [CrossRef]
- Hirst, N.L.; Lawton, S.P.; Walker, A.J. CaMKII regulates neuromuscular activity and survival of the human blood fluke Schistosoma mansoni. Sci. Rep. 2022, 12, 19831. [Google Scholar] [CrossRef] [PubMed]
- Faccidomo, S.; Reid, G.T.; Agoglia, A.E.; Ademola, S.A.; Hodge, C.W. CaMKII inhibition in the prefrontal cortex specifically increases the positive reinforcing effects of sweetened alcohol in C57BL/6J mice. Behav. Brain Res. 2016, 298, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.J.; Purohit, D.C.; Kharidia, K.M.; Mandyam, C.D. Transient Chemogenetic Inhibition of D1-MSNs in the Dorsal Striatum Enhances Methamphetamine Self-Administration. Brain Sci. 2019, 9, 330. [Google Scholar] [CrossRef]
- Jackson, K.J.; Muldoon, P.P.; Walters, C.; Damaj, M.I. Neuronal calcium/calmodulin-dependent protein kinase II mediates nicotine reward in the conditioned place preference test in mice. Behav. Pharmacol. 2016, 27, 50–56. [Google Scholar] [CrossRef]
- Jackson, K.J.; Damaj, M.I. Calcium/calmodulin-dependent protein kinase IV mediates acute nicotine-induced antinociception in acute thermal pain tests. Behav. Pharmacol. 2013, 24, 689–692. [Google Scholar] [CrossRef]
- Rosen, L.G.; Zunder, J.; Renard, J.; Fu, J.; Rushlow, W.; Laviolette, S.R. Opiate Exposure State Controls a D2-CaMKIIα-Dependent Memory Switch in the Amygdala-Prefrontal Cortical Circuit. Neuropsychopharmacology 2016, 41, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.J.; Sanjakdar, S.S.; Chen, X.; Damaj, M.I. Nicotine reward and affective nicotine withdrawal signs are attenuated in calcium/calmodulin-dependent protein kinase IV knockout mice. PLoS ONE 2012, 7, e51154. [Google Scholar] [CrossRef]
- Licata, S.C.; Schmidt, H.D.; Pierce, R.C. Suppressing calcium/calmodulin-dependent protein kinase II activity in the ventral tegmental area enhances the acute behavioural response to cocaine but attenuates the initiation of cocaine-induced behavioural sensitization in rats. Eur. J. Neurosci. 2004, 19, 405–414. [Google Scholar] [CrossRef]
- Steinkellner, T.; Mus, L.; Eisenrauch, B.; Constantinescu, A.; Leo, D.; Konrad, L.; Rickhag, M.; Sørensen, G.; Efimova, E.V.; Kong, E.; et al. In vivo amphetamine action is contingent on αCaMKII. Neuropsychopharmacology 2014, 39, 2681–2693. [Google Scholar] [CrossRef]
- Amaral, I.M.; Scheffauer, L.; Hofer, A.; El Rawas, R. Protein kinases in natural versus drug reward. Pharmacol. Biochem. Behav. 2022, 221, 173472. [Google Scholar] [CrossRef]
- Harda, Z.; Dzik, J.M.; Nalberczak-Skóra, M.; Meyza, K.; Łukasiewicz, K.; Łęski, S.; Radwanska, K. Autophosphorylation of αCaMKII affects social interactions in mice. Genes. Brain Behav. 2018, 17, e12457. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, X.D.; Zhang, J.J.; Yu, L.C. Increases in αCaMKII phosphorylated on Thr286 in the nucleus accumbens shell but not the core during priming-induced reinstatement of morphine-seeking in rats. Neurosci. Lett. 2012, 526, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, S.; Li, Z.; Zhu, R.; Jia, Z.; Ban, J.; Zhen, R.; Chen, X.; Pan, X.; Ren, Q.; et al. Effect of semaglutide and empagliflozin on cognitive function and hippocampal phosphoproteomic in obese mice. Front. Pharmacol. 2023, 14, 975830. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Parameswaran, S. Calmodulin-binding proteins: A journey of 40 years. Cell Calcium 2018, 75, 89–100. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank Docking Score | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| RSC | −235.47 | −229.10 | −221.02 | −209.48 | −209.39 | −208.42 | −208.34 | −205.57 | −201.08 | −200.44 |
| Sem | −253.10 | −235.02 | −229.27 | −227.23 | −221.67 | −221.52 | −218.32 | −217.55 | −215.91 | −210.96 |
| 0 ns | 50 ns | 100 ns | 150 ns | 200 ns | 250 ns | 300 ns |
|---|---|---|---|---|---|---|
| ALA-15 | GLN-8 | GLN-8 | GLN-8 | GLN-8 | GLN-8 | GLN-8 |
| PHE-19 | GLN-41 | GLN-41 | GLN-41 | PHE-12 | PHE-12 | PHE-12 |
| GLN-41 | MET-51 | GLU-47 | GLU-47 | GLU-47 | GLN-41 | GLU-47 |
| MET-51 | GLU-54 | MET-51 | ASP-50 | ASP-50 | GLU-47 | MET-51 |
| MET-71 | MET-71 | GLU-54 | MET-51 | MET-51 | MET-51 | GLU-54 |
| MET-72 | MET-72 | MET-72 | GLU-54 | GLU-54 | GLU-54 | MET-71 |
| LEU-112 | LYS-75 | LYS-75 | MET-72 | MET-72 | MET-71 | MET-72 |
| GLU-123 | MET-144 | LEU-112 | LEU-112 | LYS-75 | MET-72 | LYS-75 |
| MET-124 | MET-145 | MET-145 | MET-144 | ARG-90 | LYS-75 | LEU-112 |
| GLU-127 | ALA-147 | ALA-147 | MET-145 | MET-145 | MET-145 | MET-145 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floresta, G.; Arillotta, D.; Catalani, V.; Papanti Pelletier, G.D.; Corkery, J.M.; Guirguis, A.; Schifano, F. Semaglutide as a Possible Calmodulin Binder: Ligand-Based Computational Analyses and Relevance to Its Associated Reward and Appetitive Behaviour Actions. Sci. Pharm. 2024, 92, 17. https://doi.org/10.3390/scipharm92020017
Floresta G, Arillotta D, Catalani V, Papanti Pelletier GD, Corkery JM, Guirguis A, Schifano F. Semaglutide as a Possible Calmodulin Binder: Ligand-Based Computational Analyses and Relevance to Its Associated Reward and Appetitive Behaviour Actions. Scientia Pharmaceutica. 2024; 92(2):17. https://doi.org/10.3390/scipharm92020017
Chicago/Turabian StyleFloresta, Giuseppe, Davide Arillotta, Valeria Catalani, Gabriele Duccio Papanti Pelletier, John Martin Corkery, Amira Guirguis, and Fabrizio Schifano. 2024. "Semaglutide as a Possible Calmodulin Binder: Ligand-Based Computational Analyses and Relevance to Its Associated Reward and Appetitive Behaviour Actions" Scientia Pharmaceutica 92, no. 2: 17. https://doi.org/10.3390/scipharm92020017