
STAT3 Inhibitory Activities of Lignans Isolated from the Stems of Lindera obtusiloba Blume

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Extraction and Isolation
2.3. Cell Culture
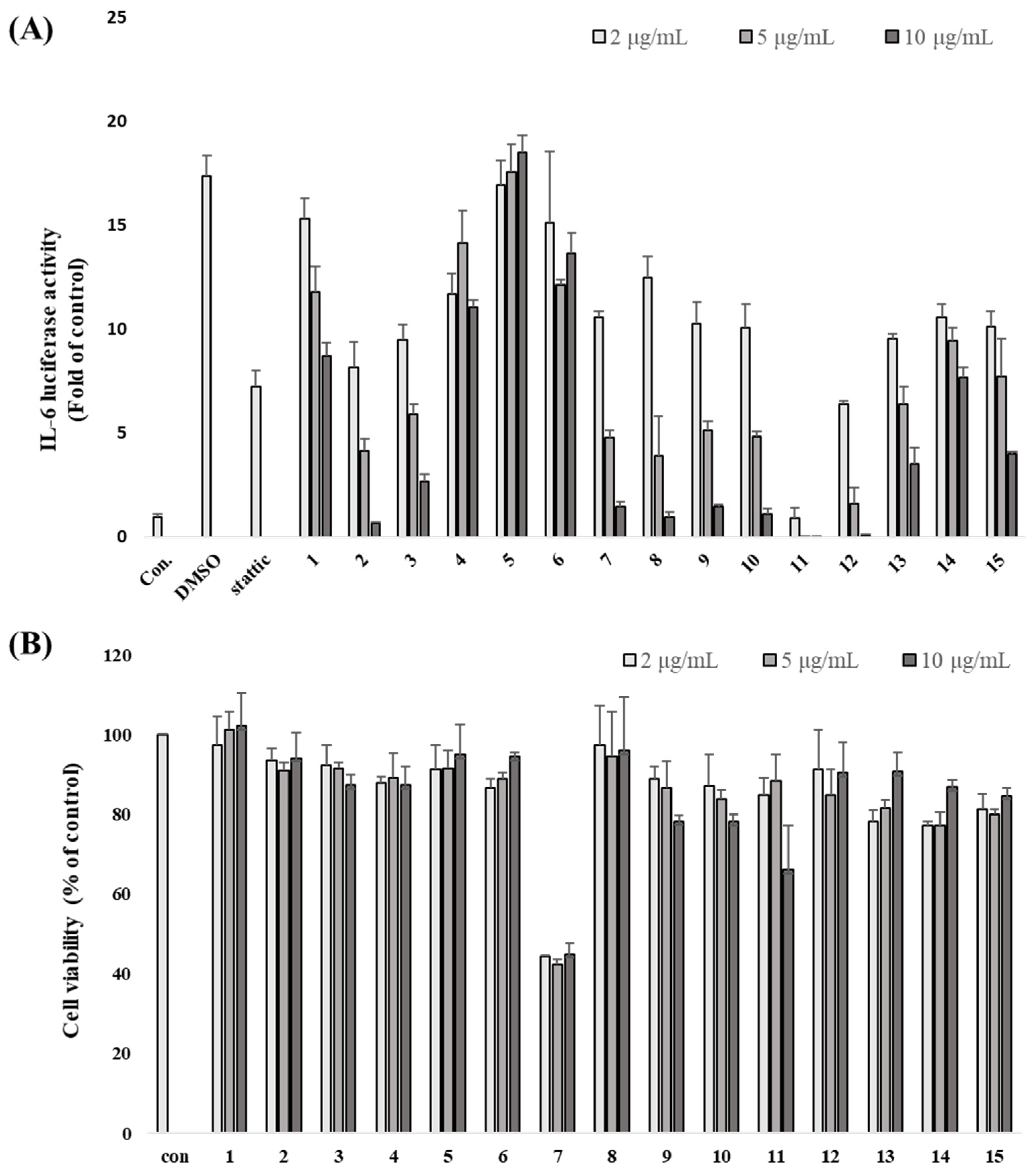
2.4. Cell Viability
2.5. IL-6-Induced STAT3 Luciferase Reporter Assay
2.5.1. Real-Time PCR
2.5.2. Western Blot Analysis
2.6. Statistical Analysis
3. Results and Discussion
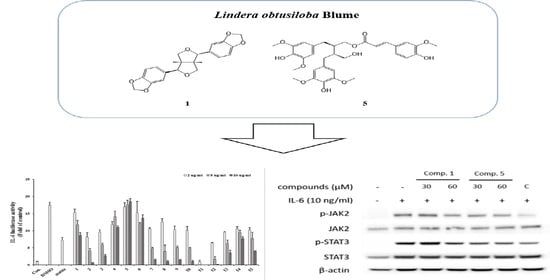
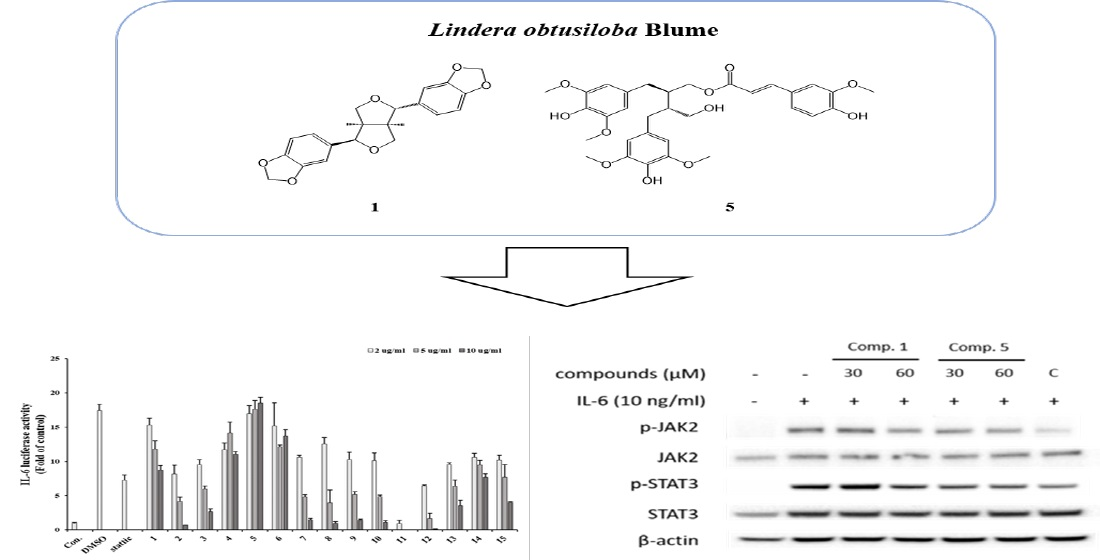
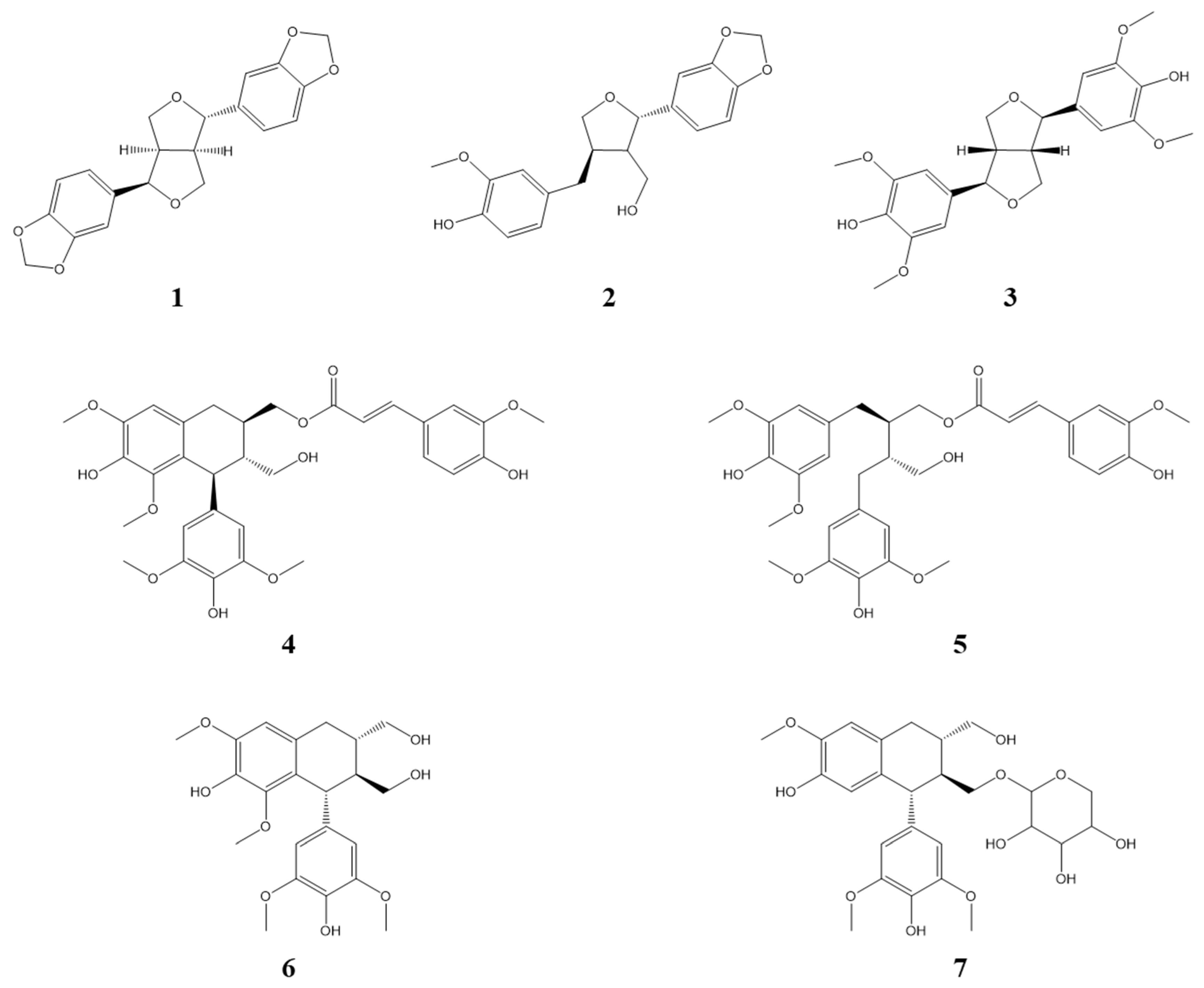
3.1. Identification of Compounds from Lindera obtusiloba
3.2. L. obtusiloba Blume Extract and Fractions Inhibit IL-6-Induced pSTAT3 Luciferase Activity
3.3. Inhibitory Effects of Isolated Compounds on IL-6-Induced pSTAT3 Luciferase Activity
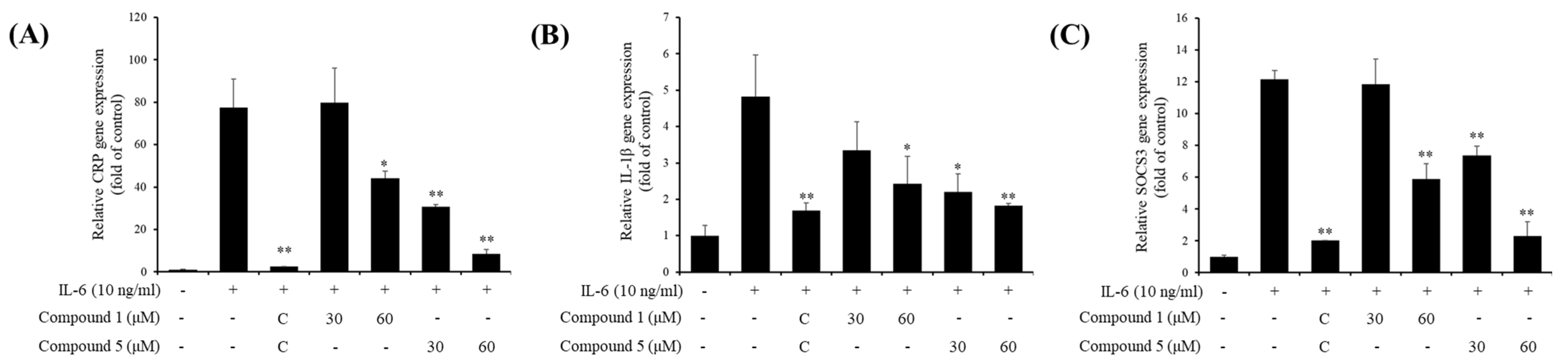
3.4. Inhibitory Effects of Active Compounds on IL-6-Induced Gene Expressions
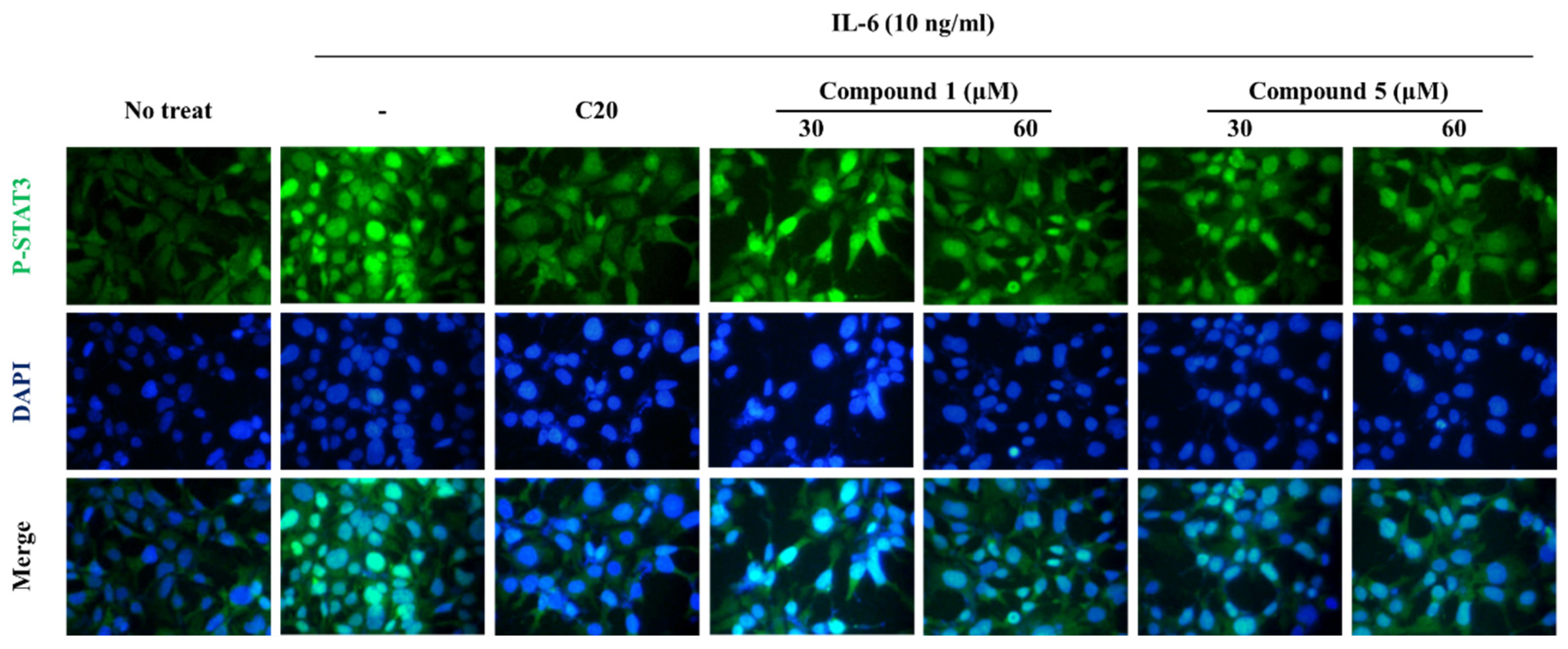
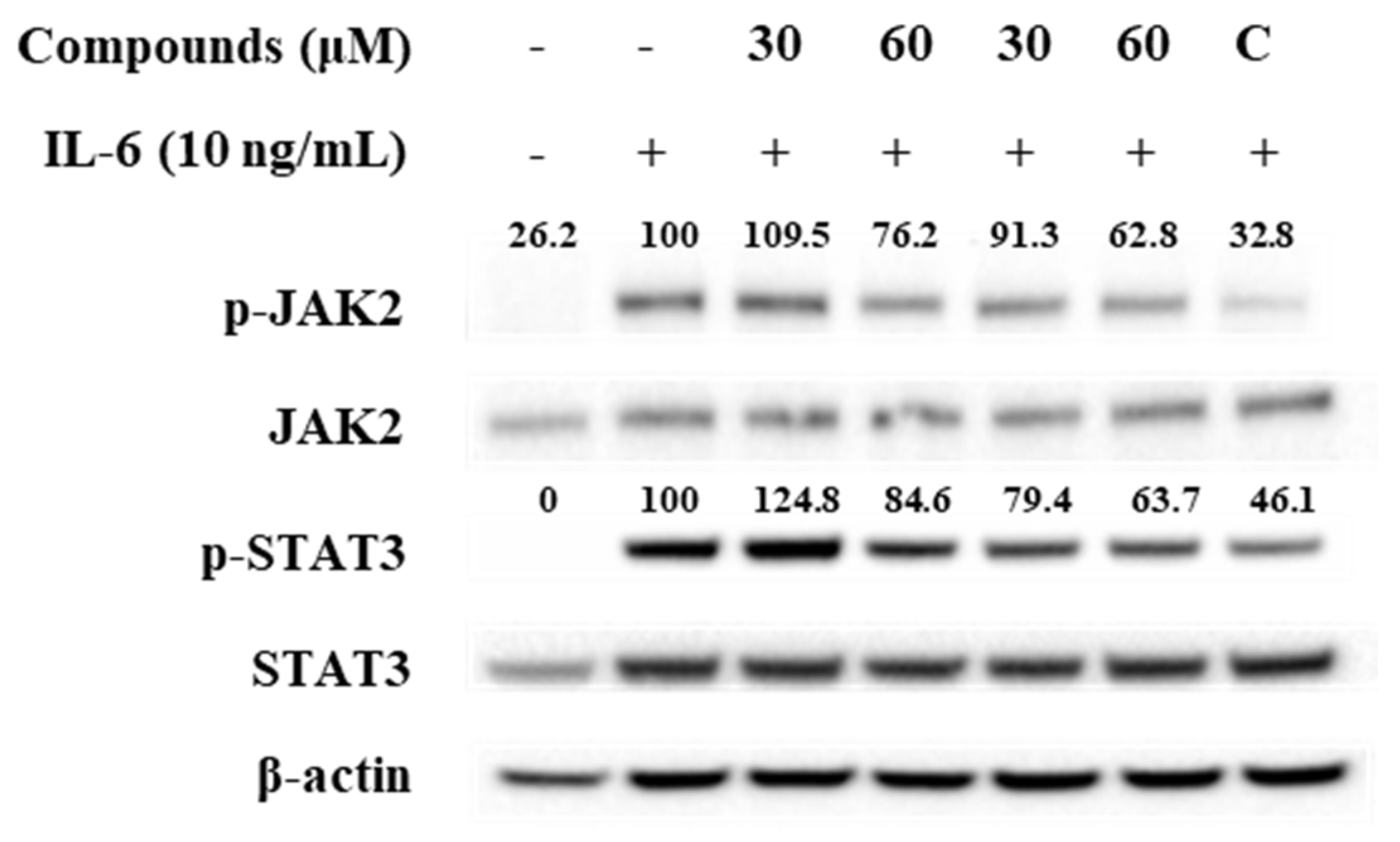
3.5. Effect of Compounds 1 and 5 on IL-6/STAT3 Signaling Molecules
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haque, M.E.; Azam, S.; Balakrishnan, R.; Akther, M.; Kim, I.S. Therapeutic potential of Lindera obtusiloba: Focus on antioxidative and pharmacological properties. Plants 2020, 9, 1765. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.O.; Rhee, C.H.; Won, N.H.; Choi, H.D.; Lee, K.W. Protective effect of 70% ethanolic extract of Lindera obtusiloba Blume on tert-butyl hydroperoxide-induced oxidative hepatotoxicity in rats. Food Chem. Toxicol. 2013, 53, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Park, J.C.; Yu, Y.B.; Lee, J.H. Isolation and structure elucidation of flavonoid glycosides from Lindera obtusiloba BL. J. Korean Soc. Food Nutr. 1996, 25, 76–79. [Google Scholar]
- Kwon, H.C.; Choi, S.U.; Lee, J.O.; Kwon, H.C.; Choi, S.U.; Lee, J.O.; Bae, K.H.; Zee, O.P.; Lee, K.R. Two New Lignan from Lindera obtusiloba blume. Arch. Pharm. Res. 1999, 22, 417–422. [Google Scholar] [CrossRef]
- Kwon, H.C.; Baek, N.I.; Choi, S.U.; Lee, K.R. New cytotoxic butanolides from Lindera obtusiloba BLUME. Chem. Pharm. Bull. 2000, 48, 614–616. [Google Scholar] [CrossRef]
- Suh, W.M.; Park, S.B.; Lee, S.; Kim, H.H.; Suk, K.; Son, J.H.; Kwon, T.K.; Choi, H.G.; Lee, S.H.; Kim, S.H. Suppression of mast-cell-mediated allergic inflammation by Lindera obtusiloba. Exp. Biol. Med. 2011, 236, 240–246. [Google Scholar] [CrossRef]
- Park, K.J.; Park, S.H.; Kim, J.K. Anti-wrinkle activity of Lindera obtusiloba extract. J. Soc. Cosmet. Sci. Korea 2009, 35, 317–323. [Google Scholar]
- Bang, C.Y.; Won, E.K.; Park, K.W.; Lee, G.W.; Choung, S.Y. Antioxidant activites and whitening effect from Lindera obtusiloba BL. Extract. Yakhak Hoeji 2008, 52, 355–360. [Google Scholar]
- Kim, J.H.; Lee, J.; Kang, S.; Moon, H.; Chung, K.H.; Kim, K.R. Antiplatelet and antithrombotic effects of the extract of Lindera obtusiloba leaves. Biomol. Ther. 2016, 24, 659–664. [Google Scholar] [CrossRef]
- Lee, J.O.; Oak, M.H.; Jung, S.H.; Park, D.H.; Auger, C. An ethanolic extract of Lindera obtusiloba stems causes NO-mediated endothelium dependent relaxations in rat aortic rings and prevents angiotensin II-induced hypertension and endothelial dysfunction in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 635–645. [Google Scholar] [CrossRef]
- Seo, K.H.; Baek, M.Y.; Lee, D.Y.; Cho, J.G. Isolation of Flavonoids and Lignans from the Stem Wood of Lindera Obtusioba Blume. J. Appl. Biol. Chem. 2011, 54, 178–183. [Google Scholar] [CrossRef]
- Kwon, D.J.; Kim, J.K.; Bae, Y.S. Essential oils from leaves and twigs of Lindera obtusiloba. J. Korean For. Soc. 2007, 96, 65–69. [Google Scholar]
- Lee, K.Y.; Kim, S.H.; Jeong, E.J.; Park, J.H.; Kim, S.H.; Kim, Y.C.; Sung, S.H. New Secoisolariciresinol Derivatives from Lindera obtusiloba Stems and Their Neuroprotective Activities. Planta Med. 2010, 76, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Freise, C.; Erben, U.; Neuman, U.; Kim, K.; Zeitz, M.; Somasundaram, R.; Ruehl, M. An active extract of Lindera obtusiloba inhibits adipogenesis via sustained Wnt signaling and exerts anti-inflammatory effects in the 3T3-L1 preadipocytes. J. Nutr. Biochem. 2011, 21, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Mark, D.T.; Belinda, N.; Tara, H.; Daniel, J.P. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumour Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef]
- Mihara, M.; Hashizume, M.; Yoshida, H.; Suzuki, M.; Shiina, M. IL-6/IL-6 receptor system and its role in physiological and pathological conditions. Clin. Sci. 2012, 122, 143–159. [Google Scholar] [CrossRef]
- Jang, H.J.; Lim, H.J.; Park, E.J.; Lee, S.J.; Lee, S.; Lee, S.W.; Rho, M.C. STAT3-inhibitory activity of sesquiterpenoids and diterpenoids from Curcuma phaeocaulis. Bioorg. Chem. 2019, 93, 103267. [Google Scholar] [CrossRef]
- Lee, S.J.; Jang, H.J.; Kim, Y.; Oh, H.M.; Lee, S.; Jung, K.; Rho, M.C. Inhibitory effects of IL-6-induced STAT3 activation of bio-active compounds derived from Salvia plebeia R. Br. Process Biochem. 2016, 51, 2222–2229. [Google Scholar] [CrossRef]
- Liu, Z.; Saarinen, N.M.; Thompson, L.U. Sesamin is one of the major precursors of mammalian lignans in sesame seed (Sesamum indicum) as observed in vitro and in rats. J. Nutr. 2006, 136, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xia, H.; Wang, L.; Xia, G.; Qu, Y.; Shang, X.; Lin, S. Lignans from the Twigs of Litsea cubeba and Their Bioactivities. Molecules 2019, 24, 306. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Huang, Y.H.; Lin, J.J. Cytotoxic lignan ester from cinnamomum osmophloeum. Planta Med. 2009, 76, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Sadhu, S.K.; Khatun, A.; Phattanawasin, P.; Ohtsuki, T.; Ishibashi, M. Lignan glycosides and flavonoids from Saraca asoca with antioxidant activity. J. Nat. Med. 2007, 61, 480–482. [Google Scholar] [CrossRef]
- Choi, H.G.; Choi, Y.H.; Kim, J.H.; Kim, H.H.; Kim, S.H.; Kim, J.A. A new neolignan and lignans from the stems of Lindera obtusiloba Blume and their anti-allergic inflammatory effects. Arch. Pharm. Res. 2014, 37, 467–472. [Google Scholar] [CrossRef]
- Rahman, M.A.; Katayama, T.; Suzuki, T.; Nakagawa, T. Stereochemistry and biosynthesis of (+)-lyoniresinol, a syringyl tetrahydronaphthalene lignan in Lyonia ovalifolia var. elliptica I: Isolation and stereochemistry of syringyl lignans and predicted precursors to (+)-lyoniresinol from wood. J. Wood Sci. 2007, 53, 161–167. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| IC50 (µM) | 14.3 ± 2.67 | >50 | >50 | 40.39 ± 3.71 | 10.83 ± 2.40 | >50 | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, E.-J.; Lim, H.J.; Lim, H.J.; Yun, B.-S.; Lee, S.; Lee, S.-J.; Lee, S.W. STAT3 Inhibitory Activities of Lignans Isolated from the Stems of Lindera obtusiloba Blume. Sci. Pharm. 2023, 91, 56. https://doi.org/10.3390/scipharm91040056
Park E-J, Lim HJ, Lim HJ, Yun B-S, Lee S, Lee S-J, Lee SW. STAT3 Inhibitory Activities of Lignans Isolated from the Stems of Lindera obtusiloba Blume. Scientia Pharmaceutica. 2023; 91(4):56. https://doi.org/10.3390/scipharm91040056
Chicago/Turabian StylePark, Eun-Jae, Hee Ju Lim, Hyung Jin Lim, Bong-Sik Yun, Soyoung Lee, Seung-Jae Lee, and Seung Woong Lee. 2023. "STAT3 Inhibitory Activities of Lignans Isolated from the Stems of Lindera obtusiloba Blume" Scientia Pharmaceutica 91, no. 4: 56. https://doi.org/10.3390/scipharm91040056