

Analysis of Differences in Single-Joint Movement of Dominant and Non-Dominant Hands for Human-like Robotic Control

Abstract
:1. Introduction
2. Methods
2.1. Participants
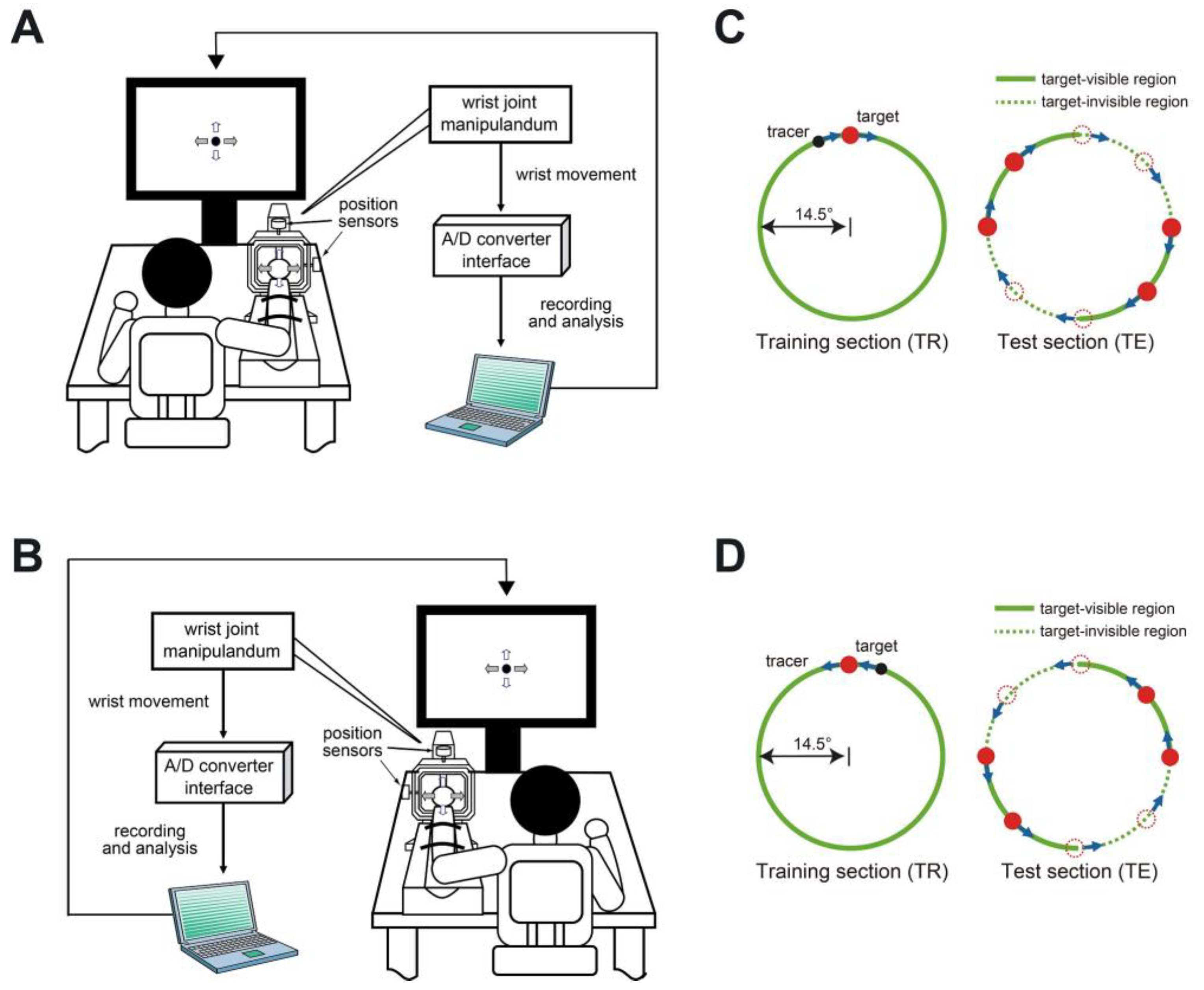
2.2. Experimental Procedure and Movement Task
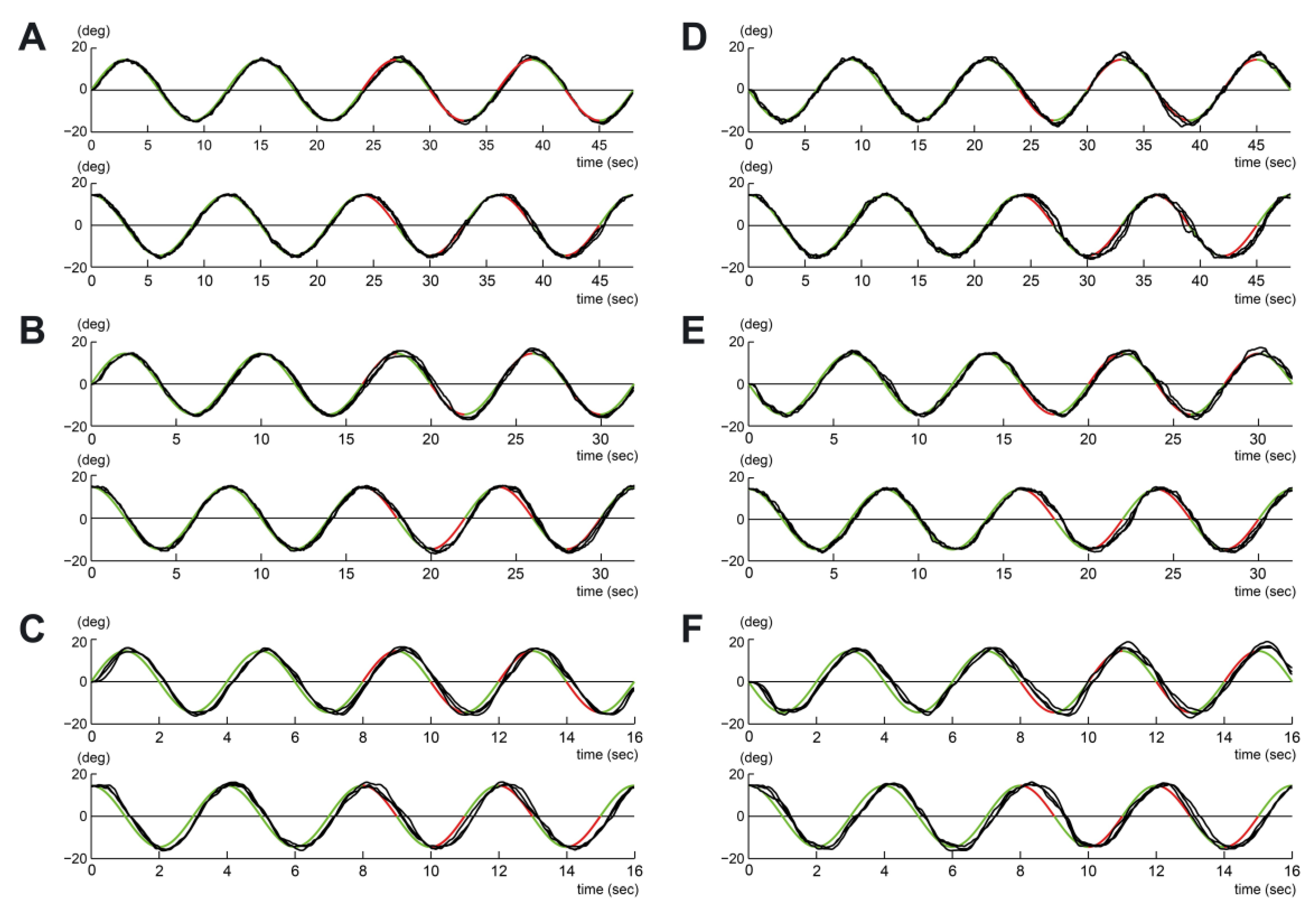
2.3. Data Analysis
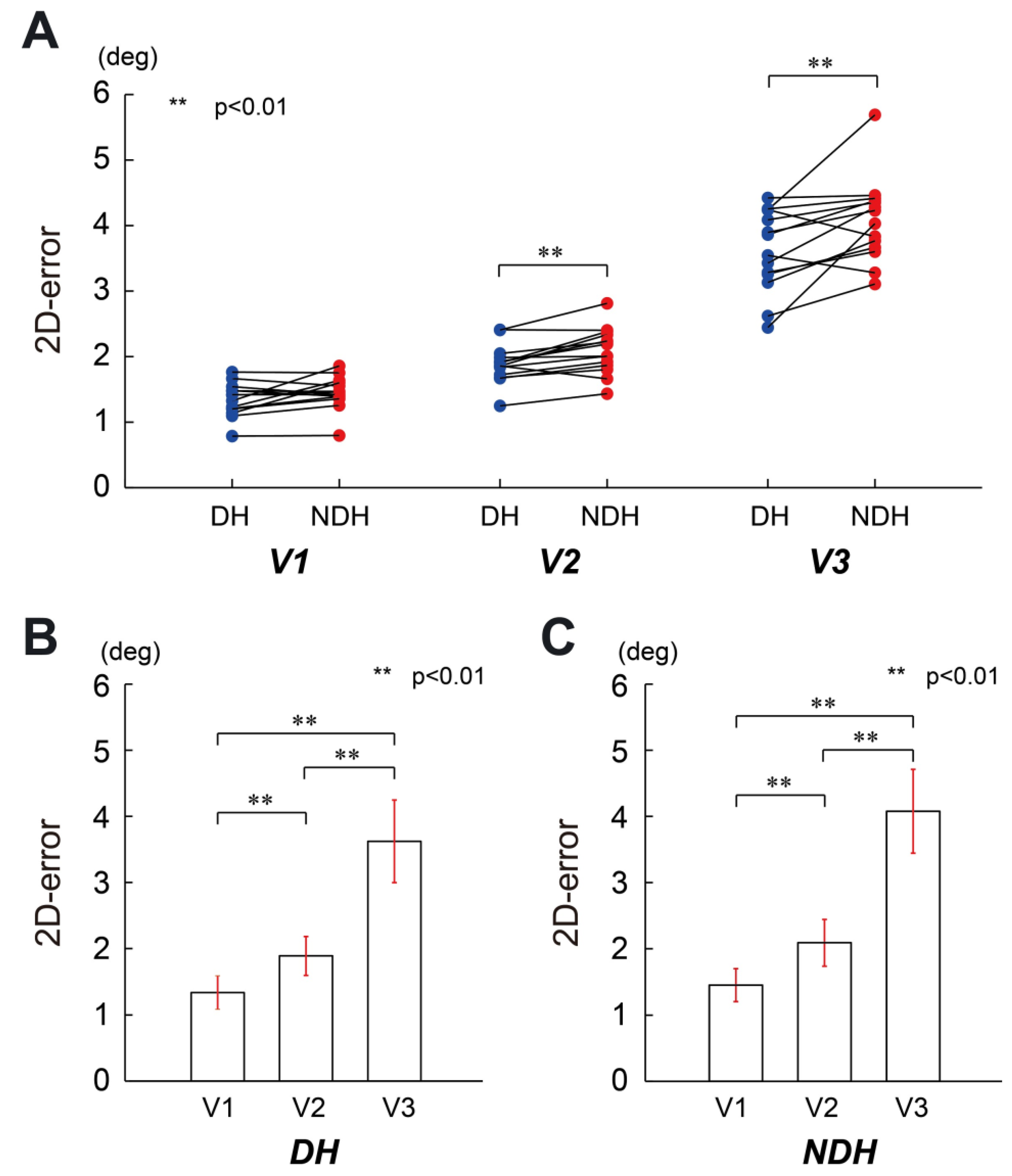
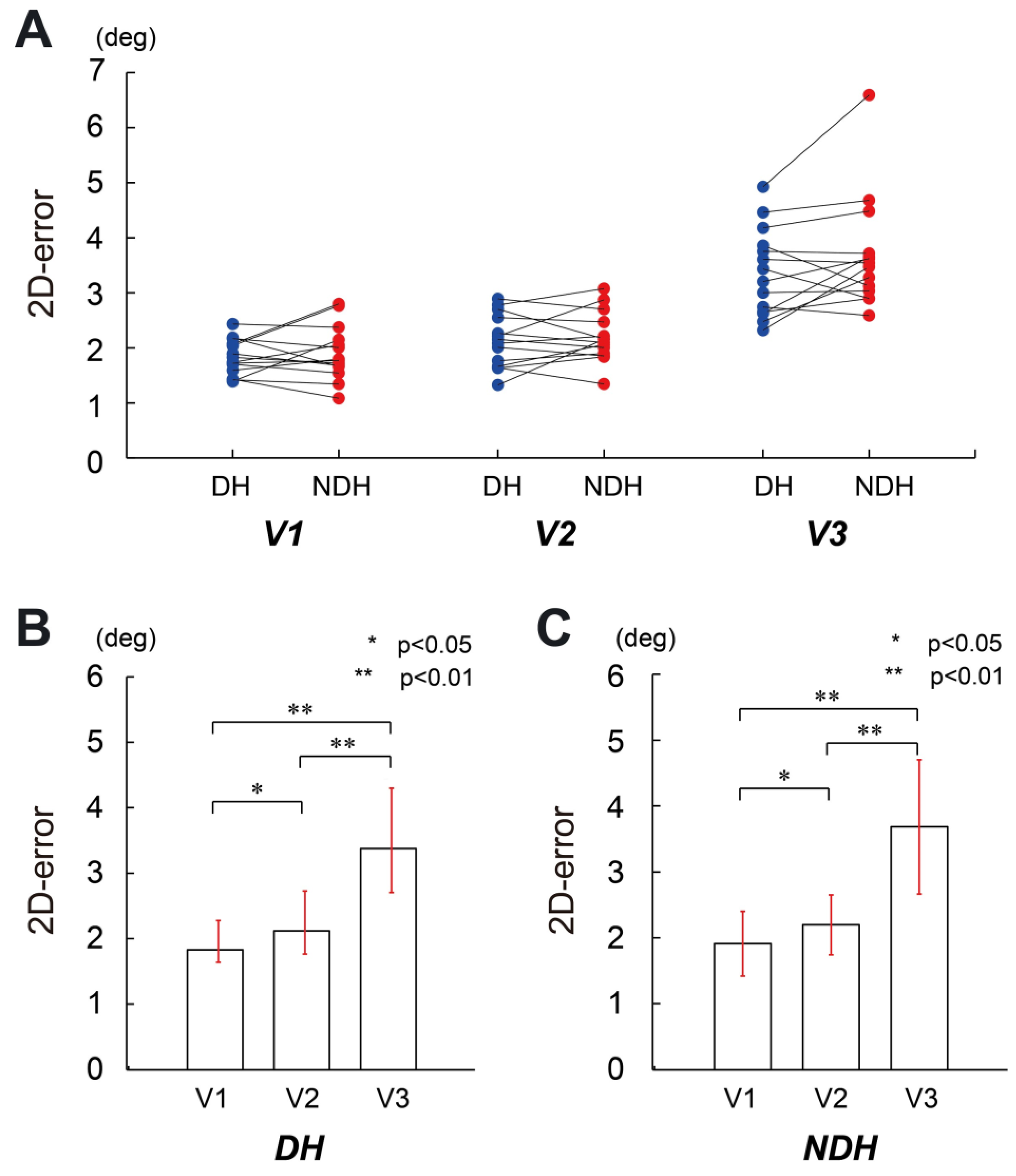
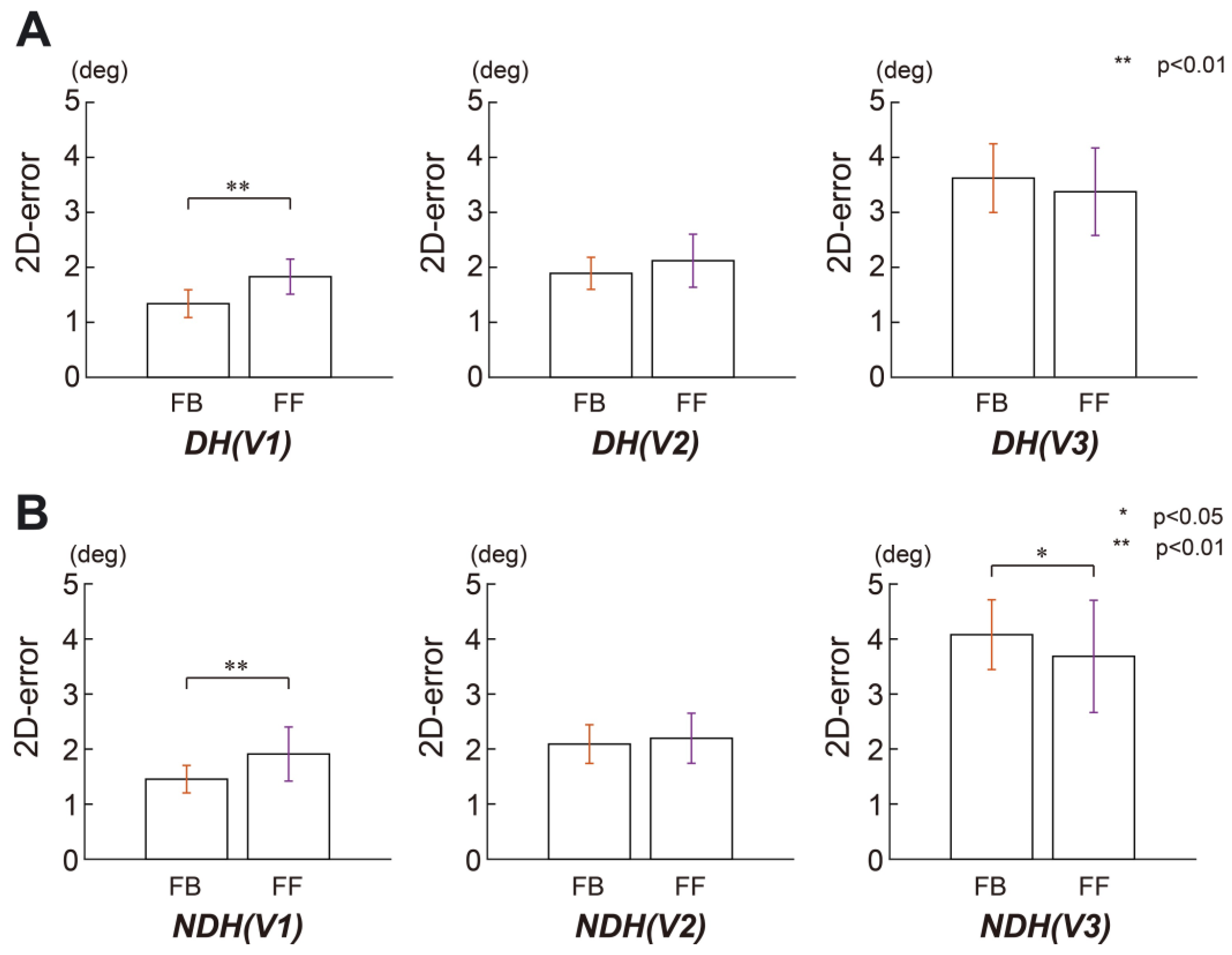
3. Results
4. Discussion
4.1. Motor Control in Single-Joint Movements
4.2. Difference in the Transference of Motor Control for Both Arms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kousaka, H.; Mizoguchi, H.; Yoshikawa, M.; Tanaka, H.; Matsumoto, Y. Role Analysis of Dominant and Non-dominant Hand in Daily Life. In Proceedings of the 2013 IEEE International Conference on Systems, Man, and Cybernetics, Manchester, UK, 13–16 October 2013; IEEE: Piscataway, NJ, USA, 2013; pp. 3972–3977. [Google Scholar]
- Mcsp, I.C.B.; Dipcot, J.A. A Comparison of Dominant and Non-dominant Hand Function in both Right- and Left-Handed Individuals using the Southampton Hand Assessment Procedure (SHAP). Br. J. Hand Ther. 2003, 8, 4–10. [Google Scholar] [CrossRef]
- Li, X.; He, W.; Li, C.; Wang, Y.-C.; Slavens, B.A.; Zhou, P. Motor unit number index examination in dominant and non-dominant hand muscles. Laterality 2015, 20, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Sharma, S.; Bilodeau, M. Neuromuscular fatigue of elbow flexor muscles of dominant and non-dominant arms in healthy humans. J. Electromyogr. Kinesiol. 2002, 12, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Troy, K.L.; Mancuso, M.E.; Johnson, J.E.; Butler, T.A.; Ngo, B.H.; Schnitzer, T.J. Dominant and nondominant distal radius microstructure: Predictors of asymmetry and effects of a unilateral mechanical loading intervention. Bone Rep. 2021, 14, 101012. [Google Scholar] [CrossRef]
- Tanaka, M.; McDonagh, M.J.; Davies, C.T. A comparison of the mechanical properties of the first dorsal interosseous in the dominant and non-dominant hand. Eur. J. Appl. Physiol. Occup. Physiol. 1984, 53, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.A.; Oldham, J.A. A comparison of dominant and non-dominant hand strengths. J. Hand Surg. Br. 1999, 24, 421–425. [Google Scholar] [CrossRef]
- Adam, A.; De Luca, C.J.; Erim, Z. Hand dominance and motor unit firing behavior. J. Neurophysiol. 1998, 80, 1373–1382. [Google Scholar] [CrossRef]
- Brouwer, B.; Sale, M.V.; Nordstrom, M.A. Asymmetry of motor cortex excitability during a simple motor task: Relationships with handedness and manual performance. Exp. Brain Res. 2001, 138, 467–476. [Google Scholar] [CrossRef]
- Goble, D.J.; Brown, S.H. Upper limb asymmetries in the matching of proprioceptive versus visual targets. J. Neurophysiol. 2008, 99, 3063–3074. [Google Scholar] [CrossRef]
- Sainburg, R.L.; Kalakanis, D. Differences in control of limb dynamics during dominant and nondominant arm reaching. J. Neurophysiol. 2000, 83, 2661–2675. [Google Scholar] [CrossRef]
- Sainburg, R.L. Evidence for a dynamic-dominance hypothesis of handedness. Exp. Brain Res. 2002, 142, 241–258. [Google Scholar] [CrossRef]
- Sainburg, R.L.; Schaefer, S.Y. Interlimb differences in control of movement extent. J. Neurophysiol. 2004, 92, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H. Control of the dominant and nondominant hand: Exploitation and taming of nonmuscular forces. Exp. Brain Res. 2007, 178, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, B.; Przybyla, A.; Murphy, T.E.; Akpinar, S.; Sainburg, R. Reaction time asymmetries provide insight into mechanisms underlying dominant and non-dominant hand selection. Exp. Brain Res. 2022, 240, 2791–2802. [Google Scholar] [CrossRef] [PubMed]
- Callaert, D.V.; Vercauteren, K.; Peeters, R.; Tam, F.; Graham, S.; Swinnen, S.P.; Sunaert, S.; Wenderoth, N. Hemispheric asymmetries of motor versus nonmotor processes during (visuo)motor control. Hum. Brain Mapp. 2011, 32, 1311–1329. [Google Scholar] [CrossRef]
- Jo, H.; Choi, W.; Lee, G.; Park, W.; Kim, J. Analysis of Visuo Motor Control between Dominant Hand and Non-Dominant Hand for Effective Human-Robot Collaboration. Sensors 2020, 20, 6368. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T. Manual asymmetry of time-delay effect on visual feedback in arm movements. In Proceedings of the 6th International Conference on Soft Computing and Intelligent Systems, and the 13th International Symposium on Advanced Intelligence Systems, Kobe, Japan, 20–24 November 2012; IEEE: Piscataway, NJ, USA, 2012; pp. 2015–2020. [Google Scholar]
- Bagesteiro, L.B.; Sainburg, R.L. Nondominant arm advantages in load compensation during rapid elbow joint movements. J. Neurophysiol. 2003, 90, 1503–1513. [Google Scholar] [CrossRef]
- Lenhard, A.; Hoffmann, J. Constant error in aiming movements without visual feedback is higher in the preferred hand. Laterality 2007, 12, 227–238. [Google Scholar] [CrossRef]
- Hu, W.; Wei, N.; Li, K.; Yue, S. Asymmetrical Mass Object Manipulations during Dominant and Non-dominant Precision Grip. In Proceedings of the 2018 14th IEEE International Conference on Signal Processing (ICSP), Beijing, China, 12–16 August 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 1010–1014. [Google Scholar]
- Gordon, J.; Ghez, C. EMG patterns in antagonist muscles during isometric contraction in man: Relations to response dynamics. Exp. Brain Res. 1984, 55, 167–171. [Google Scholar] [CrossRef]
- Gribble, P.L.; Ostry, D.J. Compensation for interaction torques during single- and multijoint limb movement. J. Neurophysiol. 1999, 82, 2310–2326. [Google Scholar] [CrossRef]
- Lee, J.; Kagamihara, Y.; Tomatsu, S.; Kakei, S. The functional role of the cerebellum in visually guided tracking movement. Cerebellum 2012, 11, 426–433. [Google Scholar] [CrossRef]
- Lee, J.; Kagamihara, Y.; Kakei, S. A New Method for Functional Evaluation of Motor Commands in Patients with Cerebellar Ataxia. PLoS ONE 2015, 10, e0132983. [Google Scholar] [CrossRef]
- Kakei, S.; Lee, J.; Mitoma, H.; Tanaka, H.; Manto, M.; Hampe, C.S. Contribution of the cerebellum to predictive motor control and its evaluation in ataxic patients. Front. Hum. Neurosci. 2019, 13, 216. [Google Scholar] [CrossRef] [PubMed]
- Kawato, M.; Gomi, H. A computational model of four regions of the cerebellum based on feedback-error learning. Biol. Cybern. 1992, 68, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Imamizu, H.; Miyauchi, S.; Tamada, T.; Sasaki, Y.; Takino, R.; Pütz, B.; Yoshioka, T.; Kawato, M. Human cerebellar activity reflecting an acquired internal model of a new tool. Nature 2000, 403, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Miall, R.C.; Weir, D.J.; Stein, J.F. Manual tracking of visual targets by trained monkeys. Behav. Brain Res. 1986, 20, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Miall, R.C.; Weir, D.J.; Stein, J.F. Intermittency in human manual tracking tasks. J. Mot. Behav. 1993, 25, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Kakei, S.; Kim, J. Motor control characteristics for circular tracking movements of human wrist. Adv. Robot. 2017, 31, 29–39. [Google Scholar] [CrossRef]
- Fine, J.M.; Ward, K.L.; Amazeen, E.L. Manual coordination with intermittent targets: Velocity information for prospective control. Acta Psychol. 2014, 149, 24–31. [Google Scholar] [CrossRef]
- Sabes, P.N. The planning and control of reaching movements. Curr. Opin. Neurobiol. 2000, 10, 740–746. [Google Scholar] [CrossRef]
- Nagaoka, M.; Tanaka, R. Contribution of kinesthesia on human visuomotor elbow tracking movements. Neurosci. Lett. 1981, 26, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Roitman, A.V.; Massaquoi, S.G.; Takahashi, K.; Ebner, T.J. Kinematic analysis of manual tracking in monkeys: Characterization of movement intermittencies during a circular tracking task. J. Neurophysiol. 2004, 91, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Beppu, H.; Suda, M.; Tanaka, R. Analysis of cerebellar motor disorders by visually guided elbow tracking movement. Brain 1984, 107 Pt 3, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Yanagihara, N.; Li, L.; Kim, J.; Lee, J. Visuomotor control of intermittent circular tracking movements with visually guided orbits in 3D VR environment. PLoS ONE 2021, 16, e0251371. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Ghahramani, Z.; Jordan, M.I. An internal model for sensorimotor integration. Science 1995, 269, 1880–1882. [Google Scholar] [CrossRef] [PubMed]
- Bagesteiro, L.B.; Sainburg, R.L. Handedness: Dominant arm advantages in control of limb dynamics. J. Neurophysiol. 2002, 88, 2408–2421. [Google Scholar] [CrossRef]
- Crevecoeur, F.; Thonnard, J.-L.; Lefèvre, P.; Scott, S.H. Long-Latency Feedback Coordinates Upper-Limb and Hand Muscles during Object Manipulation Tasks. eNeuro 2016, 3, 1–12. [Google Scholar] [CrossRef]
- Pruszynski, J.A.; Kurtzer, I.; Nashed, J.Y.; Omrani, M.; Brouwer, B.; Scott, S.H. Primary motor cortex underlies multi-joint integration for fast feedback control. Nature 2011, 478, 387–390. [Google Scholar] [CrossRef]
- Roy, E.A.; Kalbfleisch, L.; Elliott, D. Kinematic analyses of manual asymmetries in visual aiming movements. Brain Cogn. 1994, 24, 289–295. [Google Scholar] [CrossRef]
- Walker, E.H.E.; Perreault, E.J. Arm dominance affects feedforward strategy more than feedback sensitivity during a postural task. Exp. Brain Res. 2015, 233, 2001–2011. [Google Scholar] [CrossRef]
- Park, W.; Choi, W.; Jo, H.; Lee, G.; Kim, J. Analysis of Control Characteristics between Dominant and Non-Dominant Hands by Transient Responses of Circular Tracking Movements in 3D Virtual Reality Space. Sensors 2020, 20, 3477. [Google Scholar] [CrossRef] [PubMed]
- Córdova Bulens, D.; Cluff, T.; Blondeau, L.; Moore, R.T.; Lefèvre, P.; Crevecoeur, F. Different Control Strategies Drive Interlimb Differences in Performance and Adaptation during Reaching Movements in Novel Dynamics. eNeuro 2023, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Todorov, E.; Jordan, M.I. Optimal feedback control as a theory of motor coordination. Nat. Neurosci. 2002, 5, 1226–1235. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Age | Sex | Handedness |
|---|---|---|---|
| 1 | 23 | M | R |
| 2 | 22 | M | R |
| 3 | 30 | M | R |
| 4 | 21 | F | R |
| 5 | 21 | M | R |
| 6 | 22 | M | R |
| 7 | 22 | M | R |
| 8 | 22 | M | R |
| 9 | 22 | M | R |
| 10 | 22 | F | R |
| 11 | 22 | F | R |
| 12 | 22 | F | R |
| 13 | 22 | F | R |
| 14 | 22 | F | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Min, K.; Kim, Y.; Igarashi, S.; Kim, D.; Kim, H.; Lee, J. Analysis of Differences in Single-Joint Movement of Dominant and Non-Dominant Hands for Human-like Robotic Control. Sensors 2023, 23, 9443. https://doi.org/10.3390/s23239443
Kim S, Min K, Kim Y, Igarashi S, Kim D, Kim H, Lee J. Analysis of Differences in Single-Joint Movement of Dominant and Non-Dominant Hands for Human-like Robotic Control. Sensors. 2023; 23(23):9443. https://doi.org/10.3390/s23239443
Chicago/Turabian StyleKim, Samyoung, Kyuengbo Min, Yeongdae Kim, Shigeyuki Igarashi, Daeyoung Kim, Hyeonseok Kim, and Jongho Lee. 2023. "Analysis of Differences in Single-Joint Movement of Dominant and Non-Dominant Hands for Human-like Robotic Control" Sensors 23, no. 23: 9443. https://doi.org/10.3390/s23239443