
Generation of Fluorescent Bacteria with the Genes Coding for Lumazine Protein and Riboflavin Biosynthesis


Department of Biochemistry, Chungnam National University, Daejeon 34134, Korea
*
Author to whom correspondence should be addressed.
Sensors 2021, 21(13), 4506; https://doi.org/10.3390/s21134506
Submission received: 3 June 2021
/
Revised: 24 June 2021
/
Accepted: 28 June 2021
/
Published: 30 June 2021
(This article belongs to the Special Issue Application of Smart Fluorescent Proteins for Biosensing)
Abstract
:Lumazine protein is a member of the riboflavin synthase superfamily and the intense fluorescence is caused by non-covalently bound to 6,7-dimethyl 8-ribityllumazine. The pRFN4 plasmid, which contains the riboflavin synthesis genes from Bacillus subtilis, was originally designed for overproduction of the fluorescent ligand of 6,7-dimethyl 8-ribityllumazine. To provide the basis for a biosensor based on the lux gene from bioluminescent bacteria of Photobacterium leiognathi, the gene coding for N-terminal domain half of the lumazine protein extending to amino acid 112 (N-LumP) and the gene for whole lumazine protein (W-LumP) from P. leiognathi were introduced by polymerase chain reaction (PCR) and ligated into pRFN4 vector, to construct the recombinant plasmids of N-lumP-pRFN4 and W-lumP-pRFN4 as well as their modified plasmids by insertion of the lux promoter. The expression of the genes in the recombinant plasmids was checked in various Escherichia coli strains, and the fluorescence intensity in Escherichia coli 43R can even be observed in a single cell. These results concerning the co-expression of the genes coding for lumazine protein and for riboflavin synthesis raise the possibility to generate fluorescent bacteria which can be used in the field of bio-imaging.
1. Introduction
Bioluminescence and chemiluminescence refer to the process of visible light emission that occurs via exergonic chemical reactions, whereas fluorescence and phosphorescence involve remission of light from the singlet and triplet excited states, respectively [1]. Bioluminescence has long been used to image biological process in vivo. It can be detected in tissue and living organisms, providing sensitive and invasive data regarding physiological conditions [2].
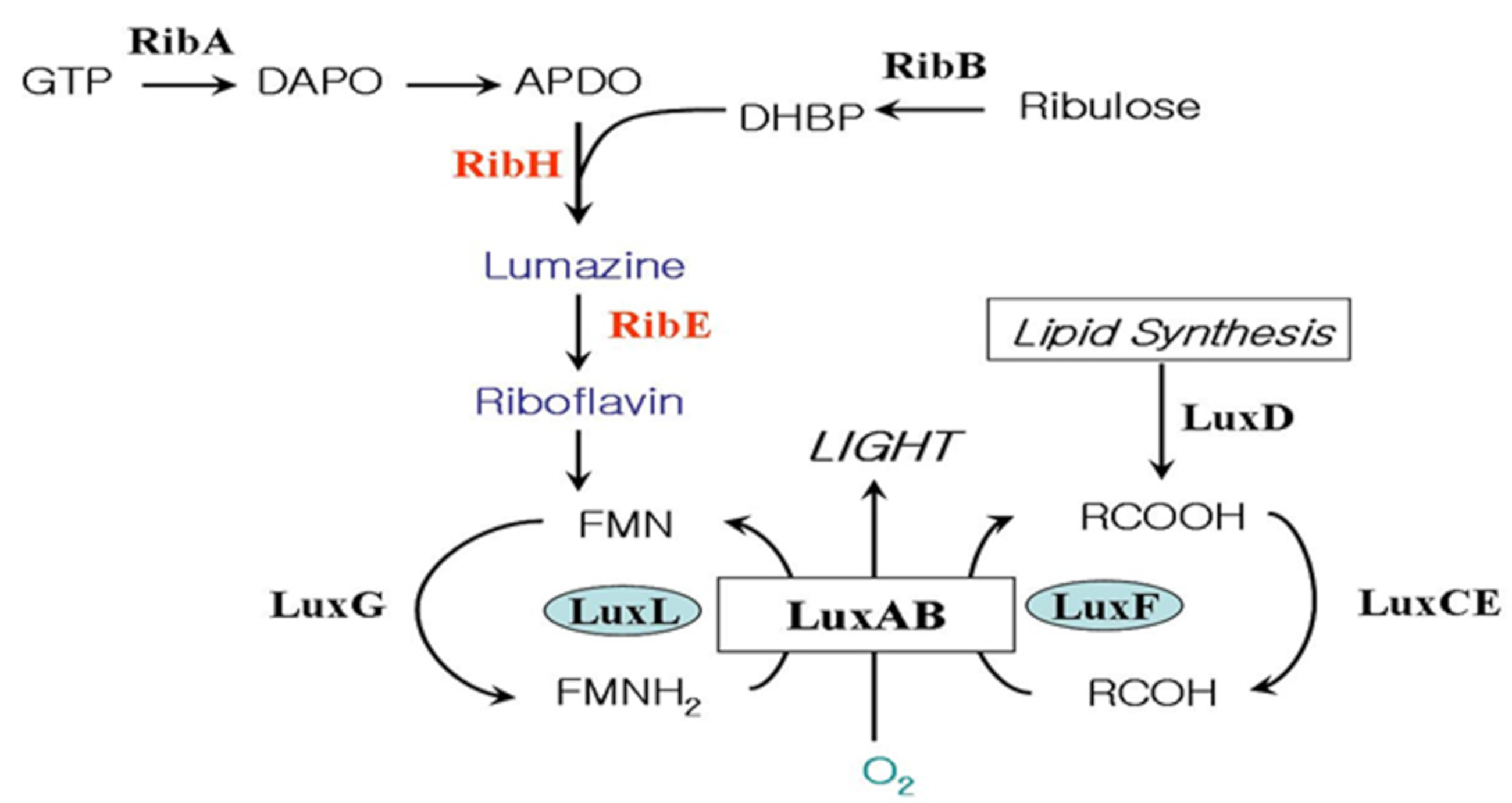
Bacterial bioluminescence reaction involves the oxidation of long chain fatty aldehyde and reduced flavin mononucleotide (FMNH2) with the emission of greenish-blue light [3] (Figure 1).
FMNH2 + RCHO + O2 → FMN + H2O + RCOOH + light
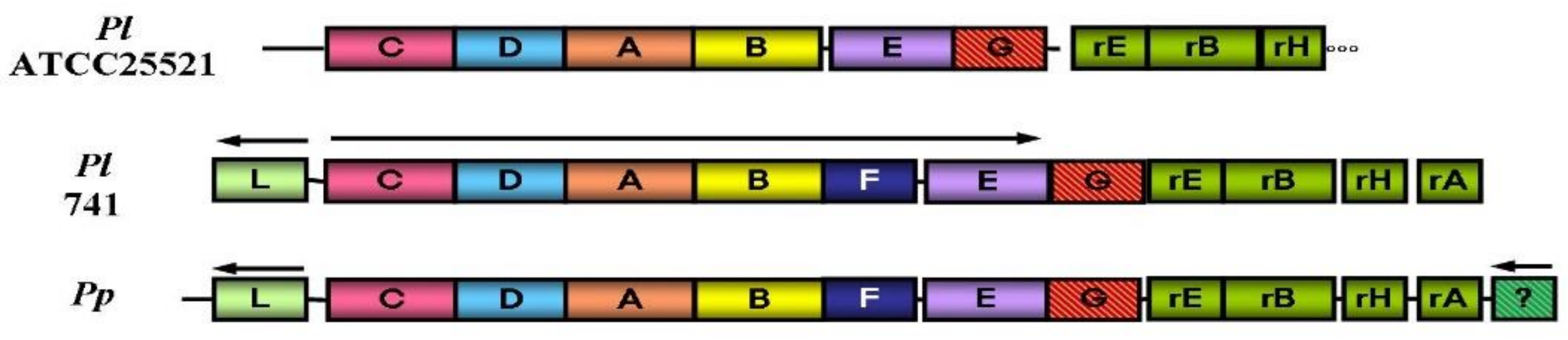
The genes coding for the enzymes and proteins responsible for the light- emitting reaction in the bioluminescent bacteria exist in a cluster forming an operon [4]. The genes related to riboflavin (vitamin B2) biosynthesis on the downstream of the lux operon of Photobacterium species were reported [5,6,7] (Figure 2). The biosynthesis of riboflavin is essential in the bioluminescent bacteria, as it is a precursor of FMNH2 (riboflavin 5′-phosphate), which is the substrate of the bioluminescence reaction as shown in Figure 1. The detection of the riboflavin genes just downstream of luxG is particularly relevant to luminescence in Photobacterium species as this genus produces the highest level of light intensities of any luminescent bacteria, with luciferase levels reaching up to 20% of the soluble proteins [8].
The fact that the first gene (ribE) next to luxG codes for riboflavin synthase in Photobacterium species is interested in the aspect of molecular genetics. This protein is thought to be created through gene duplication, as it shows about 30% amino acid identity with riboflavin synthase and is under the superfamily of riboflavin synthase [6,9]. The lumazine protein, found in most Photobacterium species in 1970 [10], was the first antenna protein that shortens the wavelength and amplifies the maximum bioluminescence intensity in Photobacterium species [11]. The lumazine protein found in Photobacterium species are a paralog of riboflavin synthase which are devoid of enzymatic activity but bind the riboflavin synthase substrate, 6,7-dimethyl 8-ribityllumazine with high affinity, and serve as an optical transponder for bioluminescence emission [11,12].
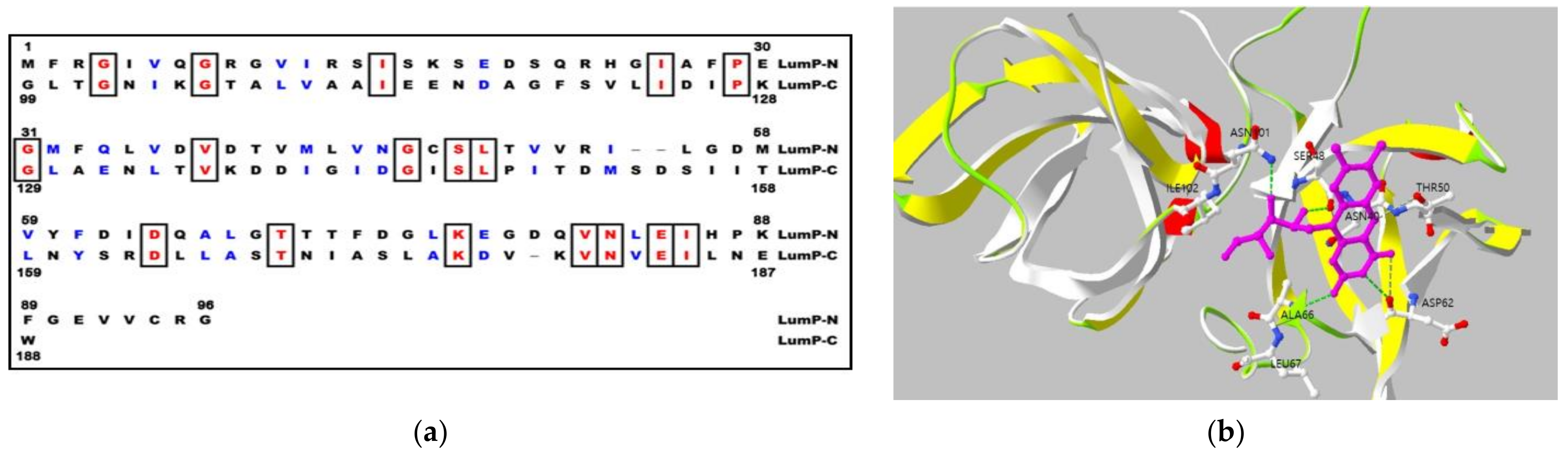
They show differences in the quaternary structure of protein, the lumazine protein has a monomeric structure whereas the riboflavin synthase forms trimeric structure [6,13]. The peculiar characteristics of lumazine protein is the intramolecular sequence similarity between N-terminal and C-terminal domain half (Figure 3a), and it was reported that the protein binds to one molecule of lumazine at N-terminal domain half [14,15] (Figure 3b).
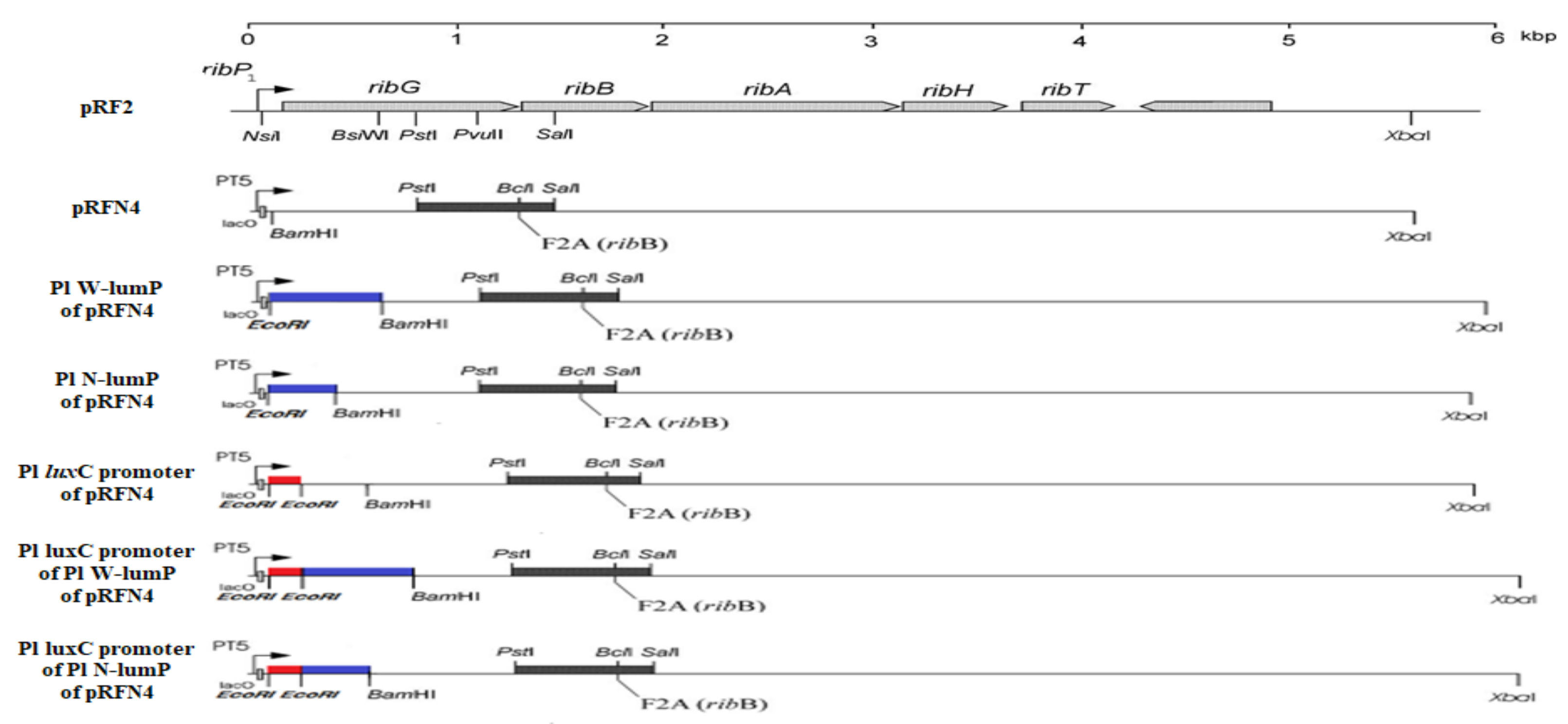
In this research, we inserted a gene coding for the lumazine protein into the pRFN4 plasmid, which contains the riboflavin biosynthesis genes of Bacillus subtilis, in order to increase the fluorescence intensity since the lumazine chromophore binds to the lumazine protein with a high affinity. The pRFN4 recombinant plasmid containing whole riboflavin operon of Bacillus subtilis were generated to produce 6,7-dimethyl 8-ribityllumazine by the missense mutation of F2A in ribE gene for riboflavin synthase [16] (Figure 4).
Internal sequence similarity (Figure 3a) as well as the comparison with the paralogous of riboflavin synthase (RS), whose three-dimensional structure of N-terminal domain half has been determined by NMR and X-ray crystallization [17,18] (Figure 3b), suggest that lumazine protein folds into two domains with closely similar folding topology. Therefore, a truncated gene specifying the N-terminal domain half of the lumazine protein (N-LumP) was constructed and the cognate protein was expressed. Using the ligand of 6,7-dimethyl-8-ribityllumazine, the fluorescence characteristics of N-LumP was investigated and reveal that the N-terminal domain half itself can bind lumazine and emit fluorescence [19]. In the paper, the gene coding for half of the N-terminal domain of lumazine protein (N-LumP), as the minimal version of fluorescent lumazine protein, as well as the whole gene coding for the whole lumazine protein (W-LumP) were amplified by polymerase chain reaction (PCR) and inserted to pRFN4 to generate recombinant plasmids for producing of fluorescent bacteria (Figure 4).
In addition, we also inserted the DNA of the lux promoter region from P. leiognathi into the above recombinant plasmids to increase the expression of the lumazine protein gene. The lux promoter DNA from bioluminescent bacteria of Photobacterium species was amplified by PCR and ligated into pRFN4, Pl W-lumP of pRFN4, and Pl N-lumP of pRFN4 plasmids with the gene coding for whole lumazine protein (W-lumP) or N-terminal domain half of lumazine protein (N-lumP) from P. leiognathi. Therefore, the recombinant plasmids were constructed into Pl luxC promoter of pRFN4, Pl luxC promoter of Pl W-lumP of pRFN4, Pl luxC promoter of Pl N-lumP of pRFN4, respectively (Figure 4). We transformed into the several E. coli strains with the pRFN4 plasmid and the various recombined plasmids. Then, we conducted a spectroscopic study on the transformed E. coli, evaluating the fluorescence intensity and examining the fluorescence of a single cell by confocal microscopy.
2. Materials and Methods
2.1. Cloning Plasmids
2.2. Strains and Cell Culture
2.3. Restriction Enzymes and Chemicals
EcoRI and BamHI were purchased from New England Biolabs, PCR premix from Genet-Bio, and PCR primers from Bioneer. To amplify the gene coding for amino-terminal domain of lumazine protein, PCR was performed with Taq polymerase (New England Biolabs).
2.4. Generation of Recombinant Plasmids
The PCR conditions were in five steps: pre-denaturation 95 °C 5 min, denaturation 9 °C 30 s, annealing 53 °C 30 s, extension 72 °C 80 sec, post-extension 72 °C 7 min. From the steps, denaturation, annealing, and extension were performed in 25 cycles. After the PCR and through the gel extraction, DNA and pRFN4 vector were cleaved with EcoRI and BamHI restriction enzymes (Figure 4). After the restriction, ligation and transformation were performed with E. coli XL-1 Blue competent cell.
The genes coding for the N-terminal lumazine protein (N-LumP) extending the gene coding for the 112th position of amino acids in lumazine protein and for whole lumazine protein (W-LumP) from P. leiognathi 741 (Figure 3) were amplified by PCR, digested with EcoRI and BamH1, and ligated into pRFN4 (Figure 4) to generate Pl N-lumP in pRFN4 and Pl W-lumP in pRFN4 recombinant plasmids. The forward primers for PCR were 5’ GGAGACCACAACGAATTCCCTCTAG3’ (Pl-lumP forward), 5’CAGAAAACCTGGGATCCTTCTATTCTATTGC3’ (Pl-N-lumP reverse), 5′GCTTGGGCTGGATCCGTTAATCACTAC3′(Pl-W-lumP-reverse). The restriction sites are underlined. The pPHl36 plasmid [6] containing the gene for lumazine protein from P. leiognathi was used as a template for PCR.
To amplify the DNA of the luxC promoter region from P. leiognathi, we used the forward primer 5′-CATGAAAAATGAATTCTAAAAAAAT CAG- 3′ (Pl luxC promoter forward) and the reverse primer 5′ -CTTAATCATGAATTCTCCTTTG GTA- 3′ (Pl luxC promoter reverse). The site of restriction enzyme EcoRI are marked by underline in the DNA sequence of the primers. The 201 bp DNA was amplified by polymerase chain reaction (PCR) using the PlXba.pT7-3 plasmid [6] as the template.
After these PCR processes, the amplified DNA was purified by agarose-gel electrophoresis, then the purified product digested with EcoRI, was inserted into the recombinant plasmids of Pl N-lumP of pRFN4 and of Pl W-lumP of pRFN4 cut by the same restriction enzyme EcoRI by ligation to generate additional recombinant plasmid of pRFN4, generating Pl luxC promoter of pRFN4, Pl luxC promoter of Pl W-lumP of pRFN4, Pl luxC promoter of Pl N-lumP of pRFN4, respectively (Figure 4). The various recombinant plasmids of pRFN4 DNA were transferred to the competent cells of E. coli XL-1. We incubated the colony and extracted a small amount of DNA. Finally, we confirmed the insertion of the right DNA sequence by the automated di-deoxynucleotide DNA sequencing analysis.
2.5. Fluorescence Measurement of Transformed E. coli
As a representative strain of transformation of recombinant DNA, the recombinant plasmids derived from pRFN4 were transferred into E. coli XL-1 Blue. In addition, the plasmids were also transformed into E. coli 43R which is known to the strain of overproduction of bioluminescence genes (Table 1). Then, to measure fluorescence, they were incubated in the conditions described as following. E. coli XL-1 Blue and E. coli 43R were incubated in LB medium without ampicillin, whereas E. coli cells harboring recombinant plasmids pRFN4, Pl-N-lumP in pRFN4, Pl-W-lumP pRFN4, as well as the recombinant plasmids of containing the promoter (Pl luxC promoter of pRFN4, Pl luxC promoter of Pl N-lumP of pRFN4, and Pl luxC promoter of Pl W-lumP of pRFN4) were grown LB media at 30 °C supplemented with 100 μg/mL ampicillin.
The cells were inoculated onto solid medium and after 24 h at 30 °C. A single bright colony was sterilely selected, inoculated in 100 mL liquid medium, and grown with shaking at 30 °C in a dark room to an absorbance of 2.5 at 600 nm. The constant amount growing cells (A600 × volume (ml) = 30) were harvested by centrifugation. The cell pellet was resuspended by buffer A (Tris-HCl 50 mM, EDTA 0.5 mM, DTT 0.5 mM, pH 7.2), sonicated (20 s, three times) and centrifuged to remove cellular debris.
2.6. Fluorescence Spectroscopic Analysis
Using an LS 45 fluorescence spectrometer (PerkinElmer), we scanned the emission spectrum of each supernatant at the fixed excitation wavelengths of 410 nm and 450 nm, respectively. The molar absorptivity of 6,7-dimethyl-8-ribityllumazine and riboflavin are 10,300/M·cm and 13,153/M·cm, respectively [16,19].
2.7. Imaging of Fluorescent E. coli under Confocal Microscopy
The E. coli cells transformed with recombinant plasmids were incubated on poly-L-lysine coated slide glasses for 1 h and mounted sample using Antifade Mounting Medium with DAPI and FITC (Thermo Fisher Scientific). We examined the single cell image of the fluorescent E. coli with a Zeiss LSM 880 confocal laser-scanning microscope (Carl Zeiss, Oberkochen, Germany) and Zen® Blue edition software (Zeiss).
3. Results
In addition to the amino acids such as Ser 48, Thr 50, and Ala 66 at the binding sites in N-terminal half of lumazine protein, several studies have shown that Asn 101 and Ile 102 located beyond N-terminal region are involved in binding of the lumazine ligand [14] (Figure 3a,b). Therefore, the genes coding for N-terminal domain half of lumazine protein extending to the amino acid 112 (N-LumP) and for the whole lumazine protein (W-LumP) from P. leiognathi were synthesized by PCR, ligated into pRFN4 vector to construct the recombinant plasmids of N-lumP-pRFN4 and W-lumP-pRFN4. We also conducted the PCR process inserting the luxC promoter domain, whose template is the PlXba.pT7-3 plasmid containing the lux genes P. leiognathi [20]. From these experiments, we generated that the recombinant plasmids, such as the Pl luxC promoter of pRFN4, the Pl luxC promoter of Pl W-lumP of pRFN4, and the Pl luxC promoter of Pl N-lumP of pRFN4, had sizes of around 9.0 kbp, 9.5 kbp, and 9.3 kbp, respectively (Figure 4).
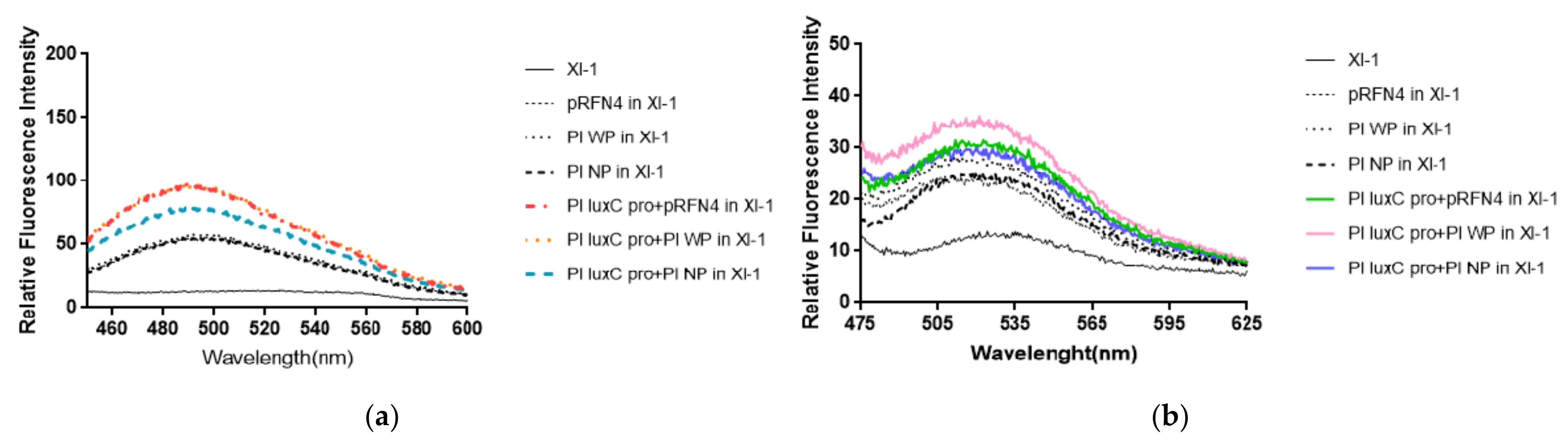
Fluorescent intensities from E. coli cells harboring the recombinant plasmid containing the genes coding for the N-terminal domain half (N-LumP) and the whole lumazine protein (W-LumP) from P. leiognathi were tested. The fluorescence intensities of supernatants of the cells in buffer A was measured with spectrofluorimeter. It was observed that the supernatant before sonication also show fluorescent, indicating that the fluorophore was present in the liquid media. Fluorescence was detected by checking for the presence of 6,7-dimethyl-8-ribityllumazine (excitation at 410 nm, molar absorptivity 10,300/M·cm) and of riboflavin (excitation at 450 nm, molar absorptivity 13,153/M·cm). We fixed the excitation wavelength of lumazine at 410 nm and measured the emission spectrum. The fluorescence intensity of E. coli XL-1 Blue that was transformed with Pl W-lumP of pRFN4 was about 1.2∼2 times stronger than that of E. coli XL-1 Blue that was transformed with pRFN4 plasmid. The fluorescence intensity of E. coli XL-1 Blue that was transformed with pRFN4 plasmid containing the DNA of the luxC promoter region of P. leiognathi ATCC25521 was about 1.5∼2 times stronger than that of E. coli XL-1 Blue that was transformed with pRFN4 plasmid. Among all transformed E. coli, the fluorescence intensity was the highest in the E. coli XL-1 Blue that was transformed with the Pl W-lumP of pRFN4 plasmid containing the DNA of the luxC promoter region (Figure 5a).
Compared to the E. coli XL-1 Blue that was transformed with pRFN4 plasmid, the fluorescence intensity was stronger in the E. coli XL-1 Blue that was transformed with Pl W-lumP of pRFN4. This result is due to the non-covalent bonding between the lumazine protein and its chromophore lumazine. In addition, the fluorescence intensity can be stimulated due to the increase of expression of the genes coding for the lumazine protein by the insertion of DNA containing the region of the lux promoter region from P. leiognathi.
Based on a previous study that reported that riboflavin competes with lumazine to bind with the lumazine protein [13], we fixed the excitation wavelength of riboflavin at 450 nm and examined the emission spectrum. As you can see in Figure 5a, we found that the emission spectrum of riboflavin excited at 450 nm was similar to that of lumazine shown in Figure 5b. Compared to the E. coli XL-1 Blue that was transformed with pRFN4 plasmid, the fluorescence intensities from riboflavin was stronger in the E. coli XL-1 Blue that was transformed with Pl luxC promoter of Pl W-lumP of pRFN4 or Pl luxC promoter of Pl N-lumP of pRFN4.
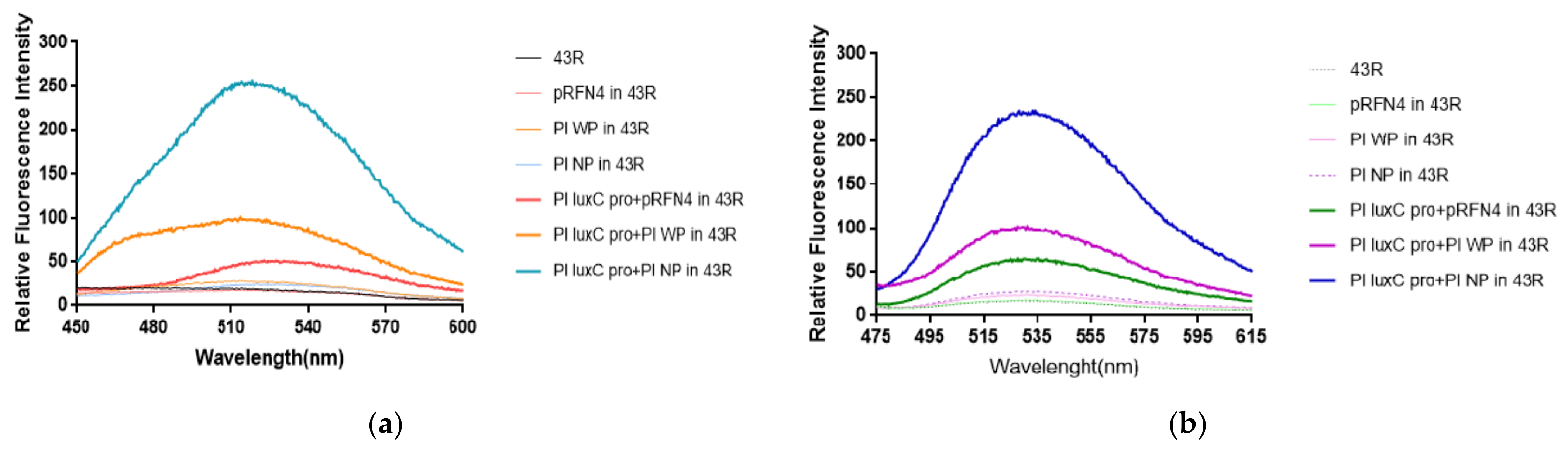
The author’s previous studies [20,21] show that luminescence intensity was significantly enhanced over 1000 times in E. coli 43R when transformed with the PlXba.pT7-3 plasmid among different E. coli strains. According to the result, we expected a larger increase of fluorescence intensity in E. coli 43R by the transformation of the pRFN4 recombinant plasmids compared to the transformation in E. coli XL-1, whereby E. coli transformed with pRFN4 plasmids show high fluorescence intensity (Figure 6a,b).
We examined the emission spectrum in the transformed E. coli 43R with the excitation wavelength fixed at 410 nm as we did in E. coli XL-1. As a result, the E. coli 43R transformed with Pl N-lumP of the pRFN4 plasmid showed about 1.5∼2 times stronger fluorescence intensity than the E. coli 43R transformed with the pRFN4 plasmid. Furthermore, the fluorescence intensity of the E. coli 43R transformed with the Pl luxC promoter of Pl N-lumP of pRFN4 was about 10 times stronger than that of the E. coli 43R transformed with the pRFN4 plasmid, and it was the strongest of all the transformed E. coli 43R (Figure 6a).
Accordingly, we suggest that this result is due to the increase of the binding of the lumazine ligand with the lumazine protein, as the lux promoter from P. leiognathi stimulates the expression of the gene coding the N-LumP. In addition, we fixed the excitation wavelength to that of riboflavin at 450 nm and examined the emission spectrum. In line with the emission spectrum of the excitation wavelength fixed to that of lumazine at 410 nm, the intensity of fluorescence was the strongest in the E. coli 43R transformed with the Pl luxC promoter of Pl N-lumP of the pRFN4 plasmid when the excitation wavelength was fixed at 450 nm (Figure 6b).
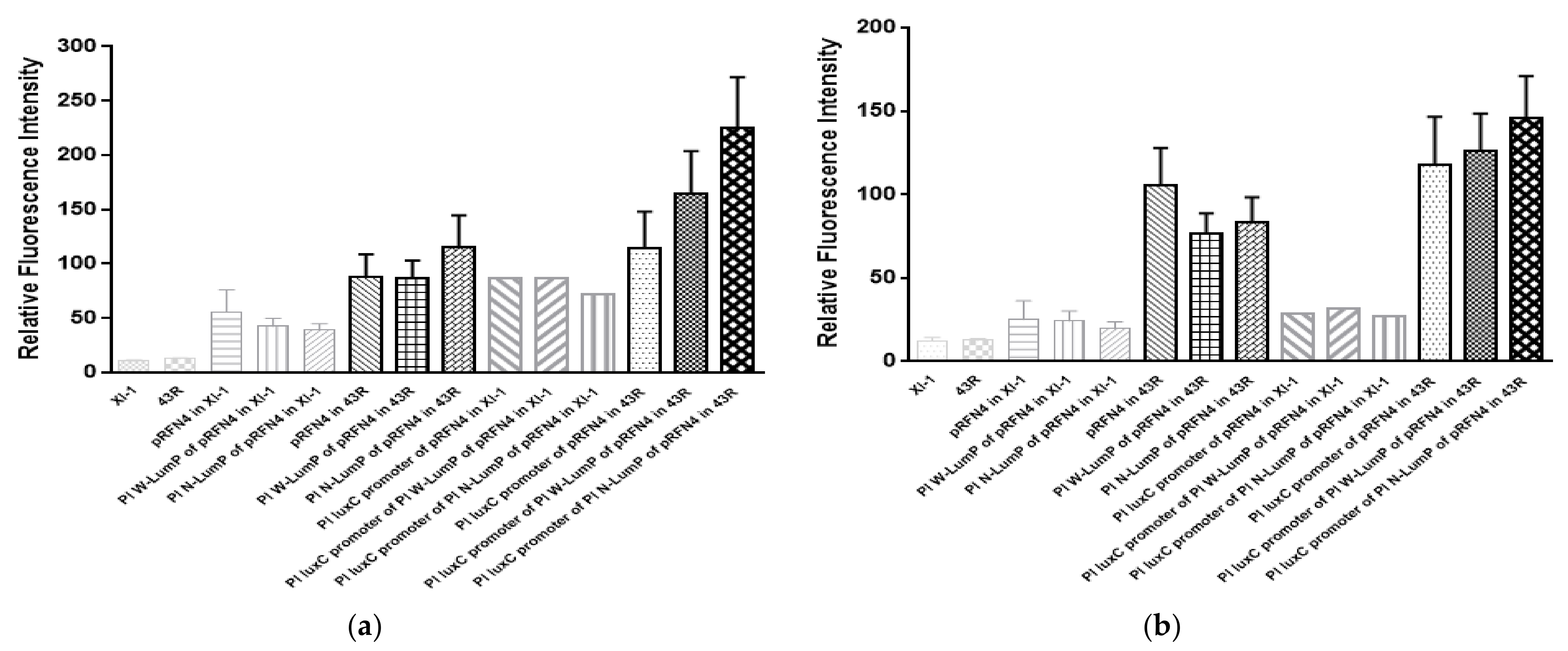
Taken together, we analyzed the average value of the emission spectrum of lumazine and riboflavin (Figure 7a,b). The increase of the fluorescence intensity at 410 nm from the cell extract of E. coli XL-1 Blue to E. coli 43R was gradually increased by about 2∼6 times when transformed with the same plasmid (Figure 7a). Moreover, in the analysis of the mean value of the emission spectrum of riboflavin excited at 450 nm, the increase of the fluorescence intensity from E. coli XL-1 Blue to E. coli 43R was sharp and varied by about 5∼6 times when transformed with the same plasmid (Figure 7b). Therefore, the highest fluorescence intensity of the cell containing the recombinant plasmid of Pl luxC promoter of Pl N-lumP of pRFN4 has a value 250 times higher compared to the intensity from E. coli XL or E. coli 43R itself.
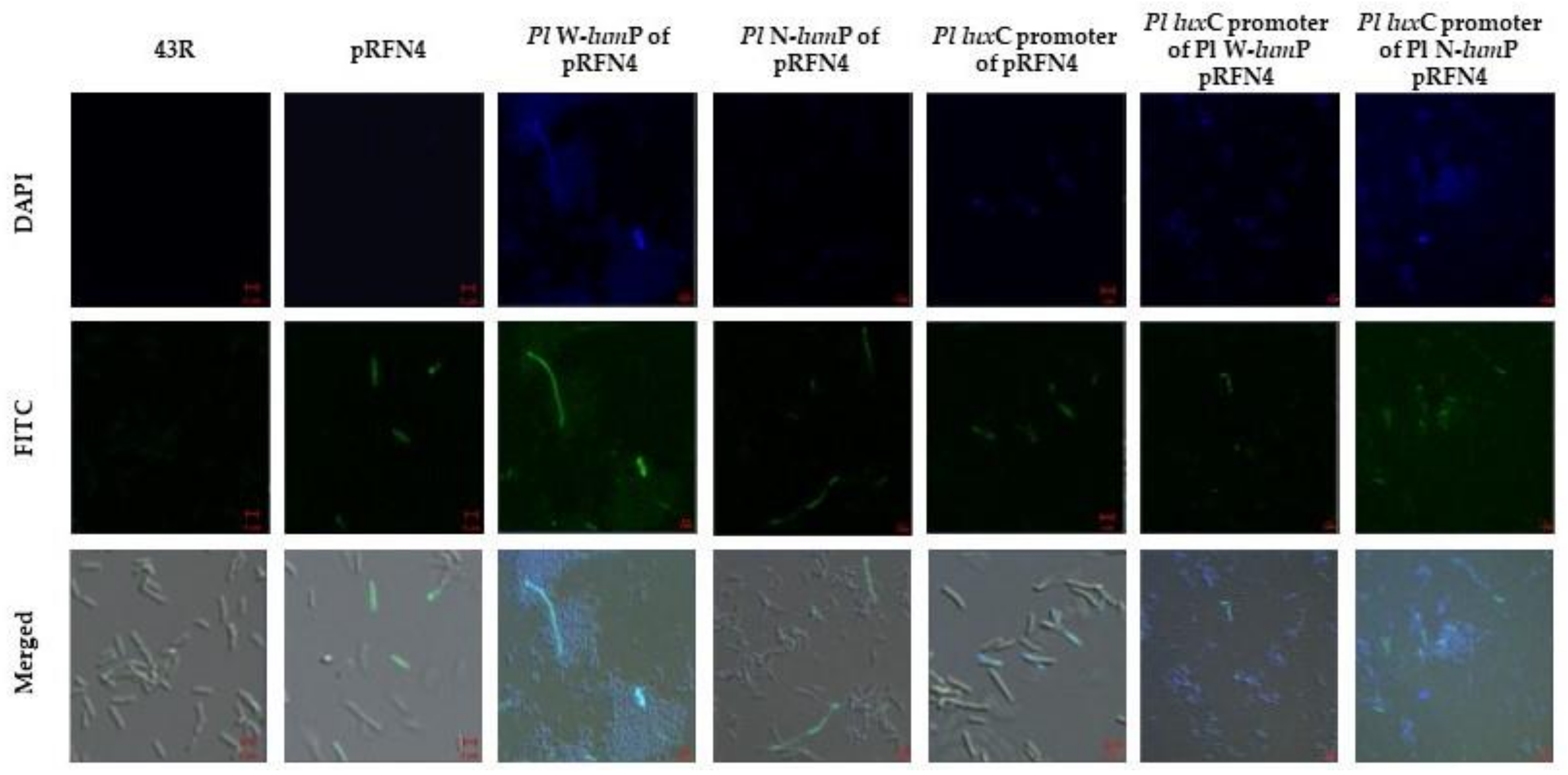
Finally, we checked the imaging of fluorescent E. coli with confocal microscopy. The E. coli 43R that was transformed using the recombination plasmid with the lux gene of Photobacterium species was incubated in a small amount of LB medium. We examined the single cell image of fluorescent E. coli using DAPI (excitation 405 nm) and FITC (excitation 458 nm) filter sets in the super-resolution confocal laser scanning microscope LSM880. The E. coli 43R that was not transformed did not show any fluorescence. On the other hand, some of the E. coli 43R that was transformed with the recombinant plasmids of pRFN4 showed fluorescence (Figure 8). We observed fluorescence in the single cell of E. coli 43R transformed with the recombinant plasmids of pRFN4 that was inserted with the genes for W-LumP and N-LumP.
The fluorescence was stronger and observed in more cells compared to the E. coli 43R transformed with only the pRFN4 plasmid. Furthermore, in the E. coli 43R transformed with the recombinant plasmid that was additionally inserted with the DNA of the P. leiognathi luxC promoter domain, the fluorescence was more intense and most of the cells expressed the fluorescence as shown in the image (Figure 8). The fluorescence intensities and frequencies in a single cell of E. coli transformed with the recombinant plasmids containing the gene coding for the lumazine protein from P. leiognathi correlated with the result with the fluorescence intensity shown in Figure 6.
4. Discussion
Bioluminescence imaging of a single cell is often complicated by the requirements of exogenous luciferin that can be poorly cell permeable or produce a high background signal [3]. Gregor et al. have engineered an improve operon ilux, which enable long term visualization of single bacterial cells while simultaneously providing information about cellular viability [22]. Bacterial bioluminescent system and light emission is being applied sensitive and safe assay not only for prokaryote gene expression but single mammalian cells [23,24]. Gregor et al. reported high luminescence levels that support the autonomous bioluminescence microscopy of mammalian cells [23].
We inserted the genes for the lumazine protein and the DNA of the lux promoter region of Photobacterium species into the pRFN4 plasmid that overproduces the chromophore lumazine, generating to various recombinant plasmids such as Pl W-lumP of pRFN4 and Pl N-lumP of pRFN4 plasmids that contain the lumazine protein gene of P. leiognathi. Then, these recombinant plasmids were transformed into the E. coli. We evaluated the fluorescence intensity using a fluorescence spectrometer and performed an imaging study of a single cell through confocal microscopy in each type of transformed E. coli. The E. coli cells that were transformed with the pRFN4 plasmids more clearly expressed the fluorescence by the overproduced lumazine ligand compared to the E. coli that was not transformed. The fluorescence was significantly stronger when E. coli was transformed with the recombinant plasmid that was also inserted with the lumazine protein gene of P. leiognathi with the luxC promoter. This gives spectroscopic evidence that the fluorescence intensity is increased when the lumazine protein is expressed along with the chromophore lumazine or riboflavin.
The recombinant plasmid DNA was transformed into E. coli strain 43R and the fluorescence intensity of the bacteria was measured. As a result, the recombinant plasmid in which the gene coding for the lumazine protein and the DNA containing lux promoter, the expression of this gene was so high to show fluorescence in single colony of E. coli. The increasing variety of fluorescent proteins and FRET (fluorescence resonance energy transfer) has significantly impacted many molecular and cellular investigations [25]. Therefore, the fluorescent system using bacterial bioluminescence genes can be utilized as a biosensor with high sensitivity and short analysis time.
In this study, we constructed series of pRFN4 recombinant plasmid derivatives, including Pl W-lumP of pRFN4, and Pl N-lumP of pRFN4 plasmids, with the gene coding for whole lumazine protein (W-LumP) or N-terminal domain half of lumazine protein (N-LumP) from P. leiognathi as well as Pl luxC promoter of pRFN4, Pl luxC promoter of Pl W-lumP of pRFN4, and Pl luxC promoter of Pl N-lumP of pRFN4 inserted the lux promoter DNA from bioluminescent bacteria of Photobacterium species, respectively. By inserting of the luxC promoter region into the plasmids, the fluorescent ligand largely bound the lumazine protein because of the high expression of the gene for lumazine protein by lux promoter in the transformed the E. coli. The fluorescent intensity of these transformed the Escherichia coli were measured by fluorescence spectrometer, and a single cell of the transformed E. coli was observed the appearance of fluorescence by using confocal microscopy. Notably, the expression of recombinant plasmids in E. coli 43R was so strong as to allow observation of the fluorescence in a single cell, raising the possibility for the generation of fluorescent bacteria which can provide the basis for a microbial biosensor.
There has been drastic interest in fluorescence for cellular and molecular imaging [26,27], and fluorescence is now a dominant methodology applied to biotechnology, flow cytometry, DNA sequencing, and genetic analysis. Fluorescence imaging can provide the information on the localization and measurement of intracellular molecules [26]. Many genetically encoded proteins that possess fluorescence and FRET have been developed to better understand the spatiotemporal regulation of various cellular process [26,27]. Whole cell sensing systems based on bioluminescence have the benefit of detecting a variety of environmental pollutants [2,28]. Therefore, the results presented in this paper raise the possibility that the Escherichia coli 43R harboring the recombinant plasmid inserted the genes coding for lumazine protein and riboflavin biosynthesis can be used as fluorescent bacteria for application in the field of bio-imaging.
Author Contributions
C.-Y.L. designed experiments. S.L. and E.O. conducted most of cell biology experiments. M.C. and E.L. performed molecular biological work with help from E.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a research grant from Chungnam National University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, J.; Müller, F.; Visser, A.J. The Sensitized Bioluminescence Mechanism of Bacterial Luciferase. Photochem. Photobiol. 2019, 95, 679–704. [Google Scholar] [CrossRef] [Green Version]
- Love, A.C.; Prescher, J.A. Seeing (and Using) the Light: Recent Developments in Bioluminescence Technology. Cell Chem. Biol. 2020, 27, 904–920. [Google Scholar] [CrossRef] [PubMed]
- Brodl, E.; Winkler, A.; Macheroux, P. Molecular Mechanisms of Bacterial Bioluminescence. Comput. Struct. Biotechnol. J. 2018, 16, 551–564. [Google Scholar] [CrossRef]
- Meighen, E.A. Genetics of bacterial bioluminescence. Annu. Rev. Genet. 1994, 28, 117–139. [Google Scholar] [CrossRef]
- Lee, C.Y.; Meighen, E.A. The Lux Genes in Photobacteriumleiognathi are Closely Linked with Genes Corresponding in Sequence to Riboflavin Synthesis Genes. Biochem. Biophys. Res. Commun. 1992, 186, 690–697. [Google Scholar] [CrossRef]
- Illarionov, B.; Illarionova, V.; Lee, J.; Vandongen, W.; Vervoort, J. Expression and Properties of the Recombinant Lumazine (Riboflavin) Protein from Photobacterium Leiognathi. Biochim. et Biophys. Acta BBA Gen. Subj. 1994, 1201, 251–258. [Google Scholar] [CrossRef]
- Lee, C.Y.; O’Kane, D.J.; Meighen, E.A. Riboflavin Synthesis Genes are Linked with the Lux Operon of Photobacterium Phosphoreum. J. Bacteriol. 1994, 176, 2100–2104. [Google Scholar] [CrossRef] [Green Version]
- Wall, L.; Rodriquez, A.; Meighen, E. Differential Acylation in vitro with Tetradecanoyl Coenzyme A and Tetradecanoic Acid (+ATP) of Three Polypeptides Shown to have Induced Synthesis in Photobacterium Phosphoreum. J. Biol. Chem. 1984, 259, 1409–1414. [Google Scholar] [CrossRef]
- O’Kane, D.J.; Woodward, B.; Lee, J.; Prasher, D.C. Borrowed Proteins in Bacterial Bioluminescence. Proc. Natl. Acad. Sci. USA 1991, 88, 1100–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gast, R.; Lee, J. Isolation of the in vivo Emitter in Bacterial Bioluminescence. Proc. Natl. Acad. Sci. USA 1978, 75, 833–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Kane, D.J.; Lee, J. Chemical Characterization of Lumazine Protein from Photobacterium Leiognathi: Comparison with Lumazine Protein from Photobacterium Phosphoreum. Biochemistry 1985, 24, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- O’Kane, D.J.; Karle, V.A.; Lee, J. Purification of Lumazine Proteins from Photobacterium Leiognathi and Photobacterium Phosphoreum: Bioluminescence Properties. Biochemistry 1985, 24, 1461–1467. [Google Scholar] [CrossRef]
- Paulus, B.; Illarionov, B.; Nohr, D.; Roellinger, G.; Kacprzak, S.; Fischer, M.; Weber, S.; Bacher, A.; Schleicher, E. One Protein, Two Chromophores: Comparative Spectroscopic Characterization of 6,7-Dimethyl-8-ribityllumazine and Riboflavin Bound to Lumazine Protein. J. Phys. Chem. B 2014, 118, 13092–13105. [Google Scholar] [CrossRef] [PubMed]
- Chatwell, L.; Illarionova, V.; Illarionov, B.; Eisenreich, W.; Huber, R.; Skerra, A.; Bacher, A.; Fischer, M. Structure of Lumazine Protein, an Optical Transponder of Luminescent Bacteria. J. Mol. Biol. 2008, 382, 44–55. [Google Scholar] [CrossRef]
- Illarionov, B.; Eisenreich, W.; Wirth, M.; Lee, C.Y.; Woo, Y.E.; Bacher, A.; Fischer, M. Lumazine Proteins from Photobacteria: Localization of the Single Ligand Binding Site to the N-Terminal Domain. Biol. Chem. 2007, 388, 1313–1323. [Google Scholar] [CrossRef]
- Illarionov, B.; Markus, F.; Chan Yong, L.; Adelbert, B.; Wolfgang, E. Rapid Preparation of Isotopolog Libraries by in Vivo Transformation of 13c-Glucose. Studies on 6, 7-Dimethyl-8-Ribityllumazine, a Biosynthetic Precursor of Vitamin B2. J. Org. Chem. 2004, 69, 5588–5594. [Google Scholar] [CrossRef] [PubMed]
- Meining, W.; Eberhardt, S.; Bacher, A.; Ladenstein, R. The Structure of the N-terminal Domain of Riboflavin Synthase in Complex with Riboflavin at 2.6 Å Resolution. J. Mol. Biol. 2003, 331, 1053–1063. [Google Scholar] [CrossRef]
- Truffaulta, V.; Colesa, M.; Diercksa, T.; Abelmanna, K.; Eberhardta, S.; Lüttgena, H.; Bachera, A.; Kesslera, H. The Solution Structure of the N-Terminal Domain of Riboflavin Synthase. J. Mol. Biol. 2001, 309, 949–960. [Google Scholar] [CrossRef]
- Kang, K.-S.; Kim, S.-Y.; Lee, J.-H.; Nam, K.-S.; Lee, E.H.; Lee, C.Y. Spectrofluorometric Properties of N-Terminal Domain of Lumazine Protein from Photobacterium leiognathi. Bull. Korean Chem. Soc. 2013, 34, 1673–1678. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, C.; Byers, D.; Graham, A.F.; Meighen, E.A. Expression of Bioluminescence by Escherichia Coli Containing Recombinant Vibrio Harveyi DNA. J. Bacteriol. 1987, 169, 247–253. [Google Scholar] [CrossRef] [Green Version]
- LEE, C.Y.; Szittner, R.B.; Meighen, E.A. The Lux Gene of the Luminous Bacterial Symbiont, Photo-bacterium Leiognathi, of the Ponyfish: Nucleotide Sequence, Difference in Gene Organization, and High Expression in Mu-tant Escherichia Coli. Eur. J. Biochem. 1991, 201, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Gregor, C.; Gwosch, K.; Sahl, S.J.; Hell, S.W. Strongly Enhanced Bacterial Bioluminescence with the Ilux Operon for Single-Cell Imaging. Proc. Natl. Acad. Sci. USA 2018, 115, 962–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregor, C.; Pape, J.K.; Gwosch, K.C.; Gilat, T.; Sahl, S.J.; Hell, S.W. Autonomous Bioluminescence Imaging of Single Mammalian Cells with the Bacterial Bioluminescence System. Proc. Natl. Acad. Sci. USA 2019, 116, 26491–26496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klose, A.D.; Paragas, N. Automated Quantification of Bioluminescence Images. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Boxer, S.G. Mechanism of Color and Photoacidity Tuning for the protonated Green Fluorescent Protein Chromophore. J. Am. Chem. Soc. 2020, 142, 11032–11041. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A. Development of Probes for Cellular Functions Using Fluorescent Proteins and Fluorescence Resonance Energy Transfer. Annu. Rev. Biochem. 2011, 80, 357–373. [Google Scholar] [CrossRef]
- Rizzo, M.A.; Springer, G.H.; Granada, B.; Piston, D.W. An Improved Cyan Fluorescent Protein Variant Useful for FRET. Nat. Biotechnol. 2004, 22, 445–449. [Google Scholar] [CrossRef]
- Rowe, L.; Dikici, E.; Daunert, S. Engineering Bioluminescent Proteins: Expanding their Analytical Potential. Anal. Chem. 2009, 81, 8662–8668. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The genes involved in bacterial bioluminescence reaction of Photobacterium species. The functions of the gene products are as follow: luciferase (LuxAB), fatty acid reductase complex (LuxCDE), non-fluorescent flavoprotein (LuxF), flavin reductase (LuxG), lumazine protein (LuxL), GTP cyclohydrolase Ⅱ (RibA), dihydroxy-butanone 4-phosphate synthase (RibB), lumazine synthase (RibH), and riboflavin synthase (RibE).
Figure 1.
The genes involved in bacterial bioluminescence reaction of Photobacterium species. The functions of the gene products are as follow: luciferase (LuxAB), fatty acid reductase complex (LuxCDE), non-fluorescent flavoprotein (LuxF), flavin reductase (LuxG), lumazine protein (LuxL), GTP cyclohydrolase Ⅱ (RibA), dihydroxy-butanone 4-phosphate synthase (RibB), lumazine synthase (RibH), and riboflavin synthase (RibE).

Figure 2.
The gene organization of lux operon region in bioluminescence bacteria of Photobacterium species. Photobacterium leiognathi ATCC 25,521 (Pl), Photobacterium leiognathi 741 (Pl), Photobacterium phosphoreum NCMB 844 (Pp). Arrow indicates the direction of transcription. ‘r’ represent for riboflavin and the functions of genes are shown in Figure 1.
Figure 2.
The gene organization of lux operon region in bioluminescence bacteria of Photobacterium species. Photobacterium leiognathi ATCC 25,521 (Pl), Photobacterium leiognathi 741 (Pl), Photobacterium phosphoreum NCMB 844 (Pp). Arrow indicates the direction of transcription. ‘r’ represent for riboflavin and the functions of genes are shown in Figure 1.

Figure 3.
(a) Internal amino acid sequences between N-terminal domain half (LumP-N) and C-terminal domain half (LumP-C) of lumazine protein from P. leiognathi 741. Identical amino acids are marked in box and similar amino acids shown in blue letters; (b) Model of binding site topology of lumazine protein.
Figure 3.
(a) Internal amino acid sequences between N-terminal domain half (LumP-N) and C-terminal domain half (LumP-C) of lumazine protein from P. leiognathi 741. Identical amino acids are marked in box and similar amino acids shown in blue letters; (b) Model of binding site topology of lumazine protein.

Figure 4.
The riboflavin genes from Bacillus subtilis in the plasmids and the gene map of recombinant plasmids with inserting Photobacterium leiognathi lux genes. Cloning plasmid of pRF2, contains the entire riboflavin operon of B. subtilis inserted into the pNCO113 plasmid reprinted from reference [16], was designed for efficient expression in E. coli. The nomenclature of the gene coding for riboflavin synthase from B. subtilis (ribB) differs from that used for that in Photobacterium species (ribE). Therefore, ribB from B. subtilis and ribE from Photobacterium species genes have the same function of coding for riboflavin synthase. The lists of recombinant plasmids of pRFN4 using this study; pRFN4, Pl W-lumP of pRFN4, Pl N-lumP of pRFN4, Pl luxC promoter of pRFN4, Pl luxC promoter of Pl W-lumP of pRFN4, Pl luxC promoter of Pl N-lumP of pRFN4 plasmids.
Figure 4.
The riboflavin genes from Bacillus subtilis in the plasmids and the gene map of recombinant plasmids with inserting Photobacterium leiognathi lux genes. Cloning plasmid of pRF2, contains the entire riboflavin operon of B. subtilis inserted into the pNCO113 plasmid reprinted from reference [16], was designed for efficient expression in E. coli. The nomenclature of the gene coding for riboflavin synthase from B. subtilis (ribB) differs from that used for that in Photobacterium species (ribE). Therefore, ribB from B. subtilis and ribE from Photobacterium species genes have the same function of coding for riboflavin synthase. The lists of recombinant plasmids of pRFN4 using this study; pRFN4, Pl W-lumP of pRFN4, Pl N-lumP of pRFN4, Pl luxC promoter of pRFN4, Pl luxC promoter of Pl W-lumP of pRFN4, Pl luxC promoter of Pl N-lumP of pRFN4 plasmids.

Figure 5.
The fluorescence properties of E. coli XL-1 Blue transformed with recombinant plasmids of pRFN4 comtainin the lux gene from P. leiognathi. (a) Emission spectrum by lumazine in fixation at excitation wavelength of 410 nm. (b) Emission spectrum by riboflavin excited at excitation wavelength of 450 nm. NP and WP denote N-lumP and W-lumP, respectively.
Figure 5.
The fluorescence properties of E. coli XL-1 Blue transformed with recombinant plasmids of pRFN4 comtainin the lux gene from P. leiognathi. (a) Emission spectrum by lumazine in fixation at excitation wavelength of 410 nm. (b) Emission spectrum by riboflavin excited at excitation wavelength of 450 nm. NP and WP denote N-lumP and W-lumP, respectively.

Figure 6.
The fluorescence properties of E. coli 43R transformed with recombinant plasmids of pRFN4 containing the lux from P. leiognathi (a) Emission spectrum by lumazine excited at excitation wavelength of 410 nm. (b) Emission spectrum by riboflavin excited at wavelength of 450 nm. NP and WP denote N-lumP and W-lumP, respectively.
Figure 6.
The fluorescence properties of E. coli 43R transformed with recombinant plasmids of pRFN4 containing the lux from P. leiognathi (a) Emission spectrum by lumazine excited at excitation wavelength of 410 nm. (b) Emission spectrum by riboflavin excited at wavelength of 450 nm. NP and WP denote N-lumP and W-lumP, respectively.

Figure 7.
(a) Analysis of average fluorescence emission intensity from the different transformants. The x axis of bar graph means relative fluorescence intensity by lumazine in fixation at excitation wavelength of 410 nm from the E. coli transformed with recombinant plasmids from P. leiognathi lux gene. (b) Average emission value by riboflavin were obtained in fixation at excitation wavelength of 450 nm. The x axis of bar graph means relative fluorescence intensity from the E. coli transformed with recombinant plasmids from P. leiognathi lux gene.
Figure 7.
(a) Analysis of average fluorescence emission intensity from the different transformants. The x axis of bar graph means relative fluorescence intensity by lumazine in fixation at excitation wavelength of 410 nm from the E. coli transformed with recombinant plasmids from P. leiognathi lux gene. (b) Average emission value by riboflavin were obtained in fixation at excitation wavelength of 450 nm. The x axis of bar graph means relative fluorescence intensity from the E. coli transformed with recombinant plasmids from P. leiognathi lux gene.

Figure 8.
The microscopic images of E. coli 43R containing the recombinant plasmids of pRFN4 inserted the lux genes from P. leiognathi.
Figure 8.
The microscopic images of E. coli 43R containing the recombinant plasmids of pRFN4 inserted the lux genes from P. leiognathi.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains and plasmids used in this study.
| Strains | Characteristics | Source |
| E. coli XL-1 Blue | Cloning strain | Real Biotech corporation |
| E. coli 43R | Mutant of E. coli RR1 strain | C. Miyamoto et al., [20] |
| Plasmids | Characteristics | Source |
| pRFN4 | Recombinant plasmid containing the riboflavin genes from Bacillus subtilis | Illarionov, B., et al., [16] |
| pPhl36 | pT7-7 containing the gene for the wild type of lumazine protein from Photobacterium leiognathi 741 | Illarinov, B., et al., [6] |
| PlXba in pT7 | The recombinant pT7-3 plasmids containing the cloned P.leiognathi ATCC 25,521 lu0078 DNA | Lee et al., [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lim, S.; Oh, E.; Choi, M.; Lee, E.; Lee, C.-Y. Generation of Fluorescent Bacteria with the Genes Coding for Lumazine Protein and Riboflavin Biosynthesis. Sensors 2021, 21, 4506. https://doi.org/10.3390/s21134506
AMA Style
Lim S, Oh E, Choi M, Lee E, Lee C-Y. Generation of Fluorescent Bacteria with the Genes Coding for Lumazine Protein and Riboflavin Biosynthesis. Sensors. 2021; 21(13):4506. https://doi.org/10.3390/s21134506
Chicago/Turabian StyleLim, Sunjoo, Eugeney Oh, Miae Choi, Euiho Lee, and Chan-Yong Lee. 2021. "Generation of Fluorescent Bacteria with the Genes Coding for Lumazine Protein and Riboflavin Biosynthesis" Sensors 21, no. 13: 4506. https://doi.org/10.3390/s21134506
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.