

Assessing Tree Water Balance after Forest Thinning Treatments Using Thermal and Multispectral Imaging
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
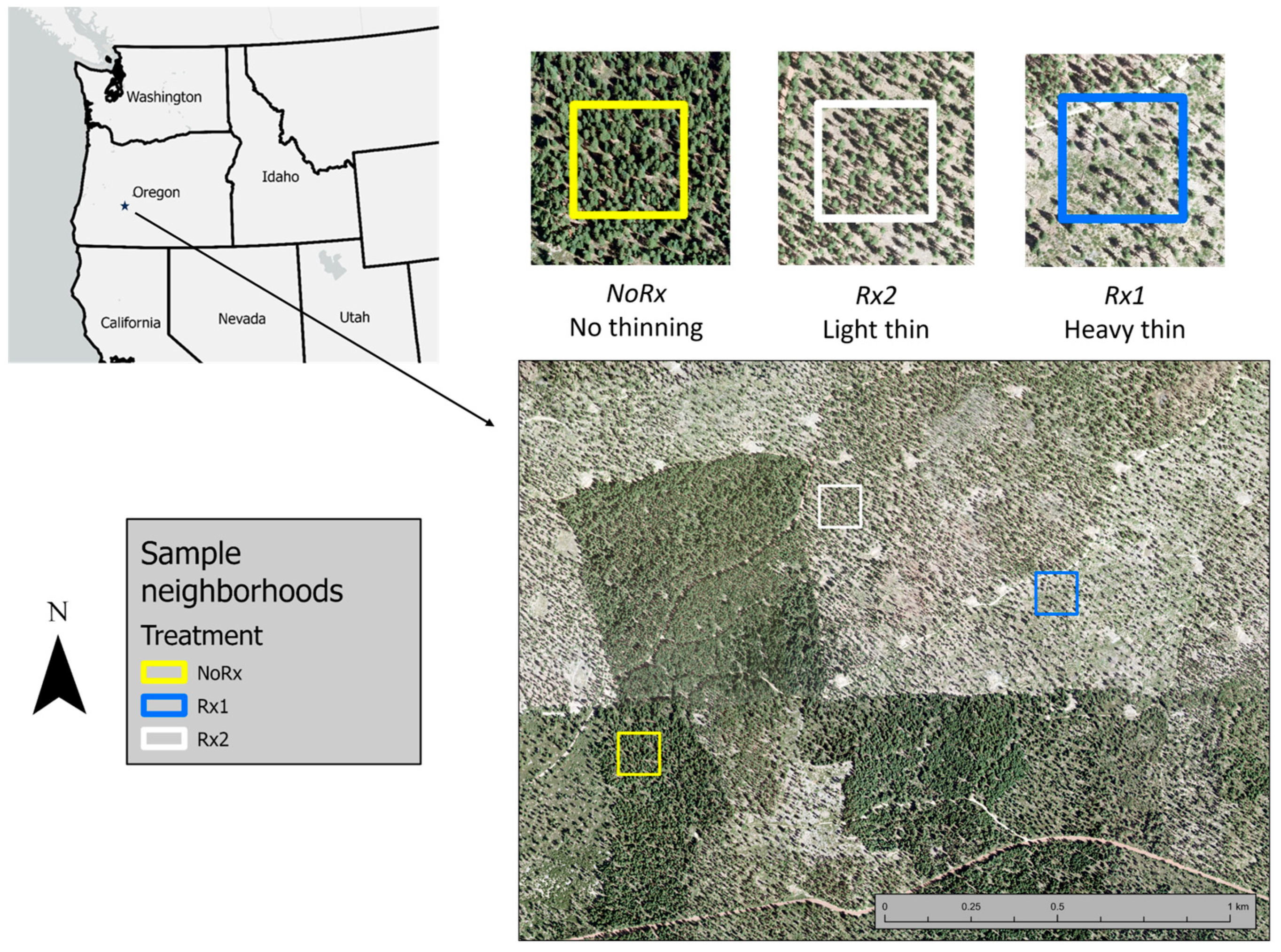
2.1. Site Location
2.2. Stand Characteristics
2.3. Whole Tree and Crown Morphological Attributes
2.4. Tree-Level Transpiration
2.5. On-Site Temperature, Vapor Pressure Deficit, and Humidity
2.6. Remote Sensing Data Collection
2.7. Data Compilation and Analysis
3. Results
3.1. Tree Morphological Attributes and Transpiration by Treatment
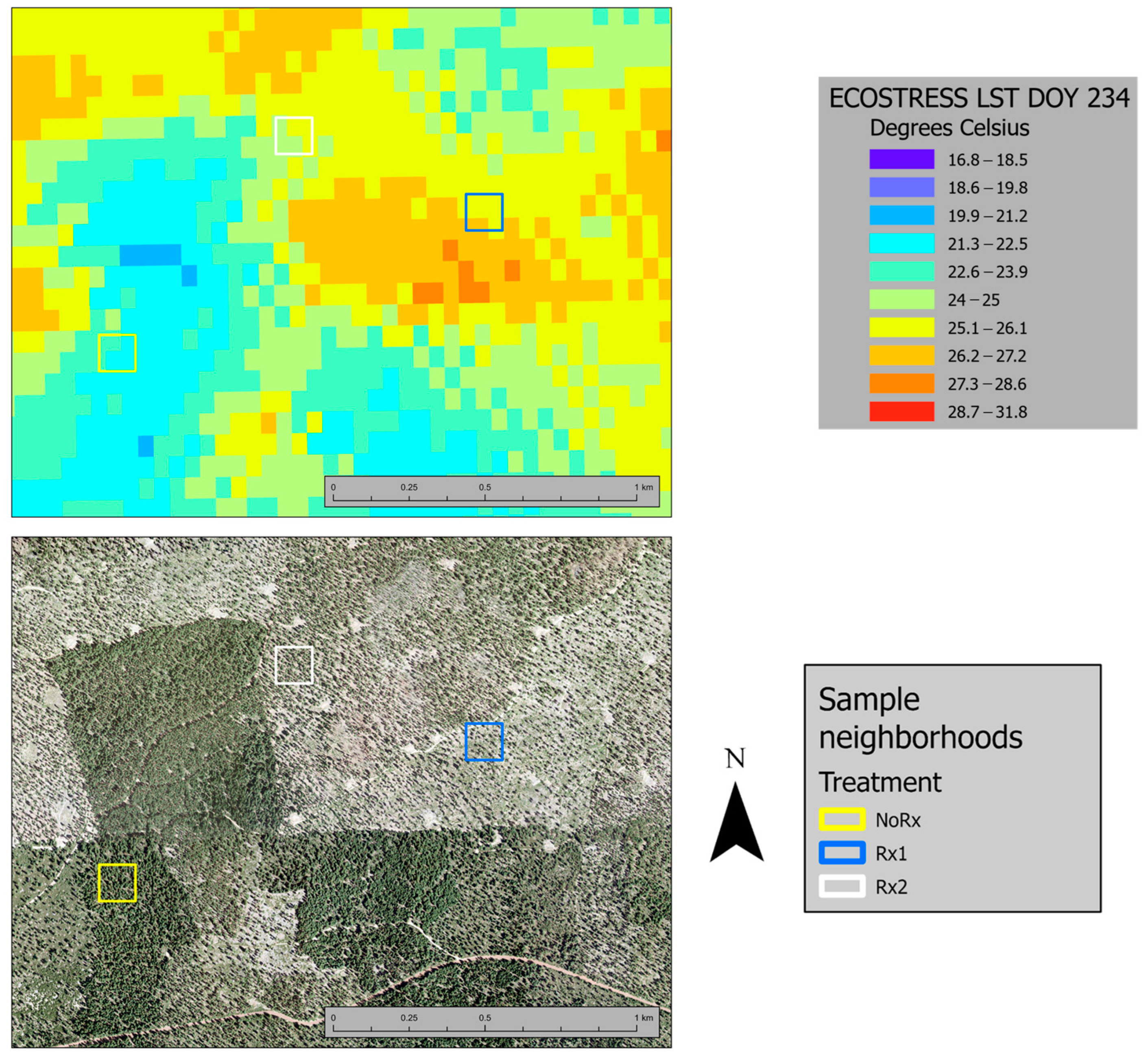
3.2. Remote Detection of Crown Temperature
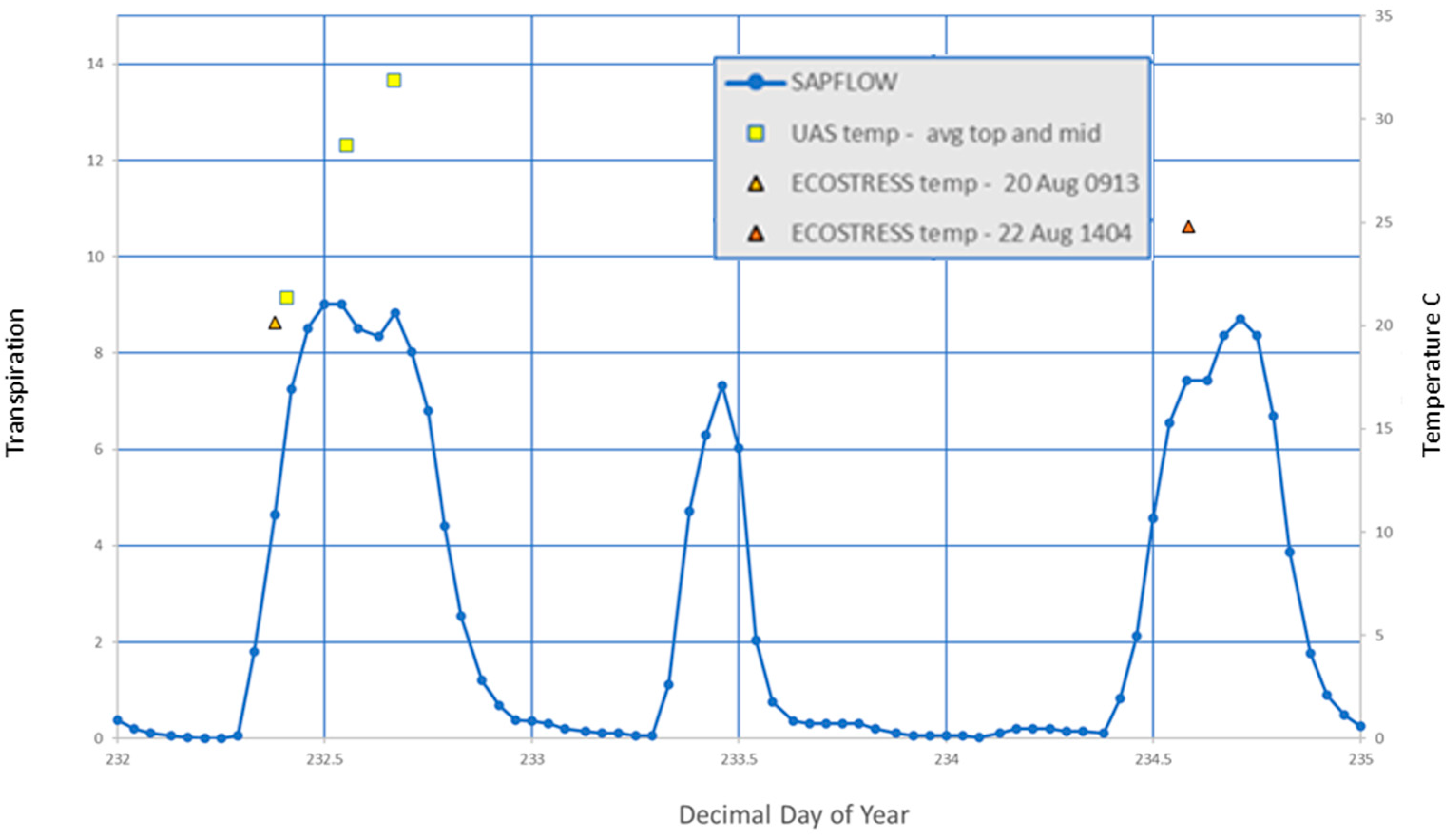
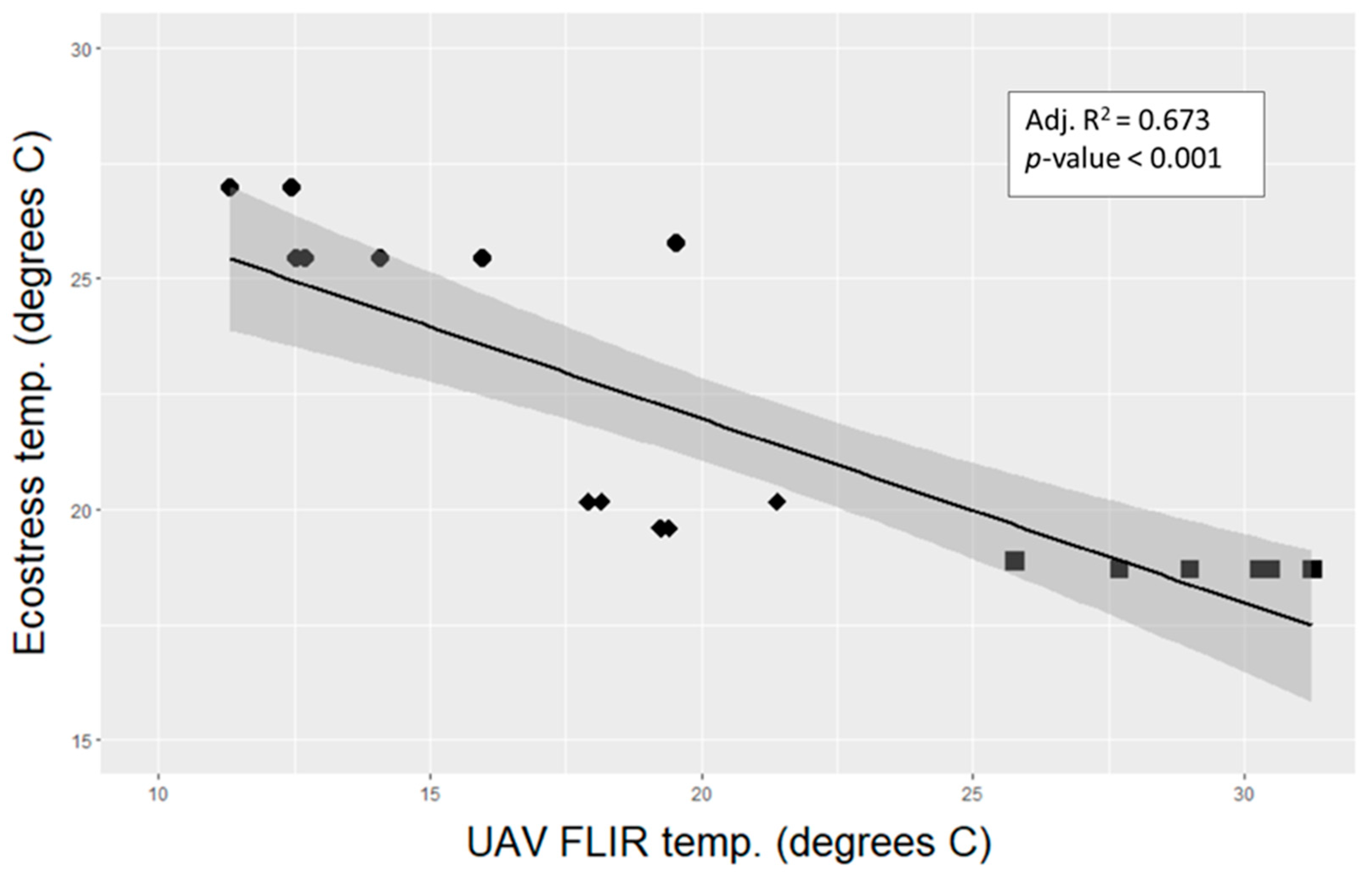
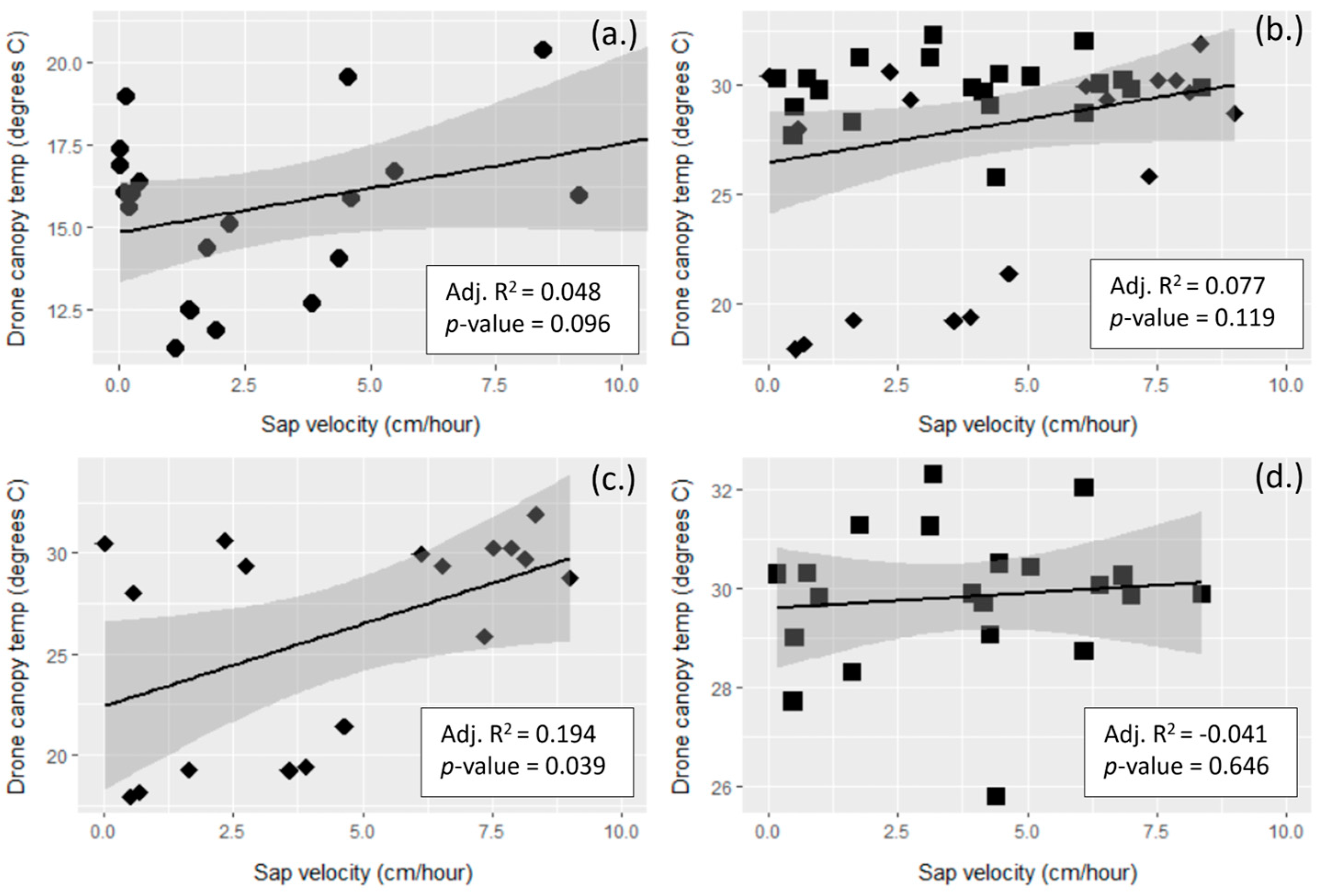
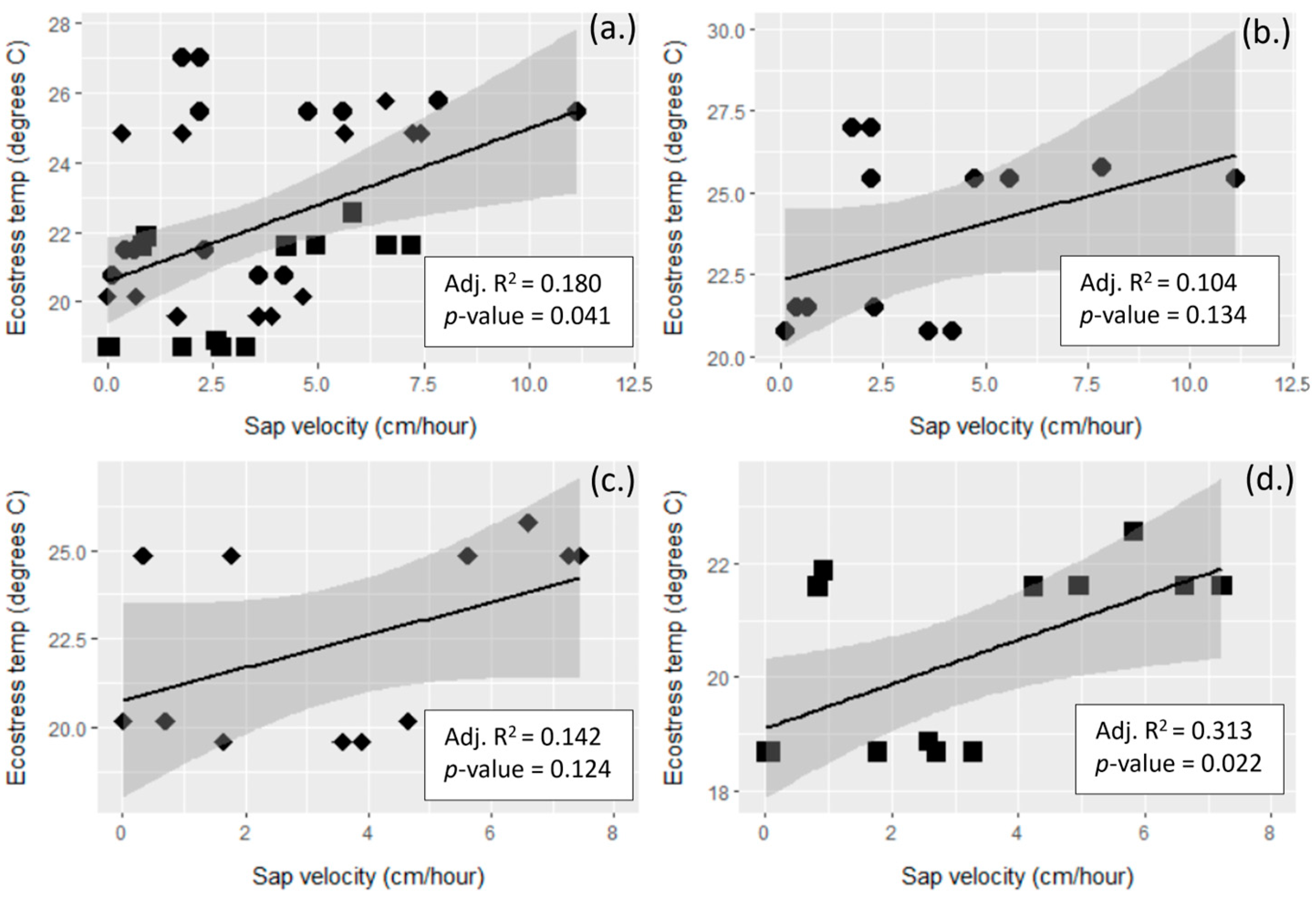
3.3. Remotely Sensed Temperature vs. Tree Transpiration
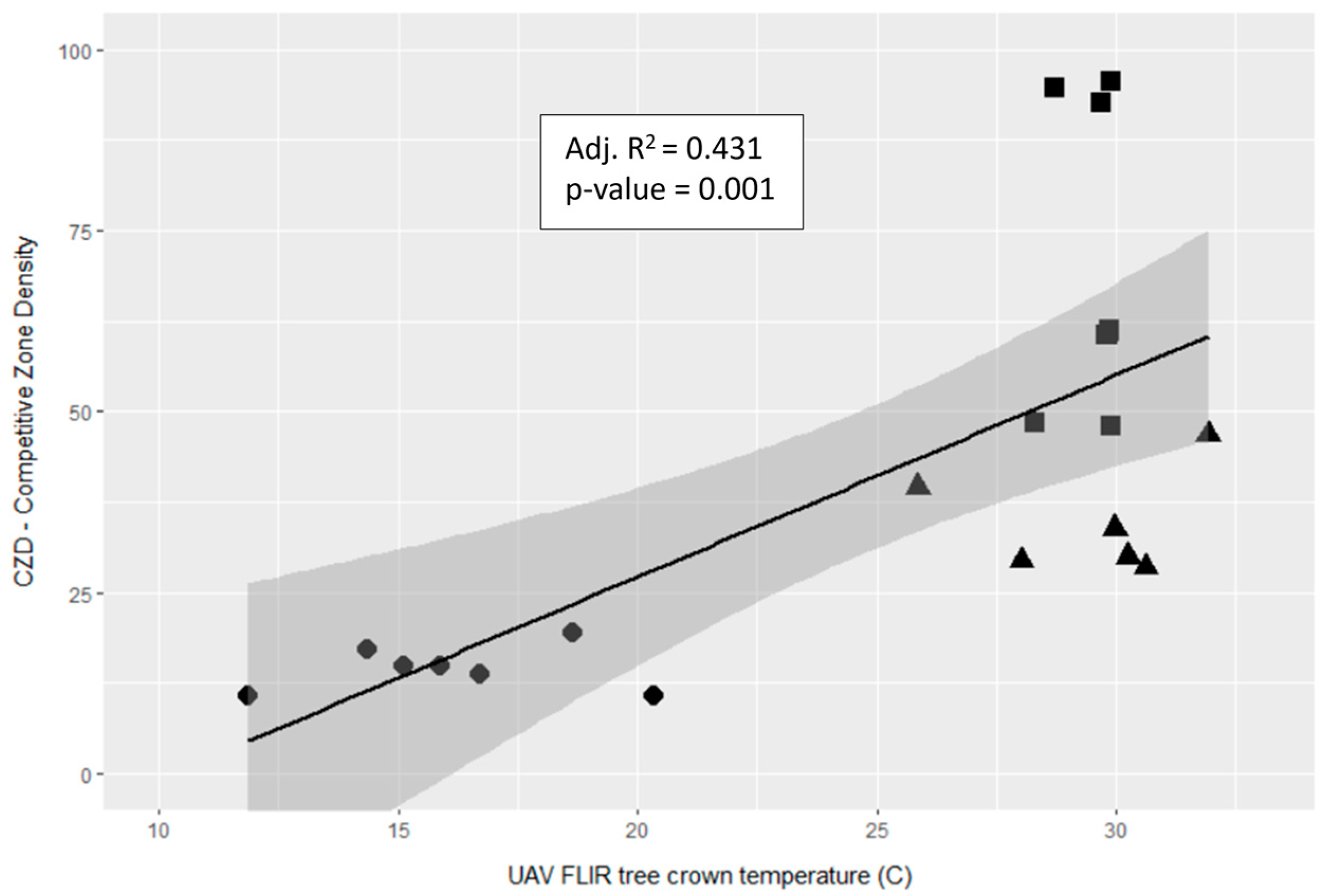
3.4. Contributing Predictors of UAV FLIR Crown Temperature (Td)
3.5. Contributing Predictors of Tree Level Transpiration (Etrans)
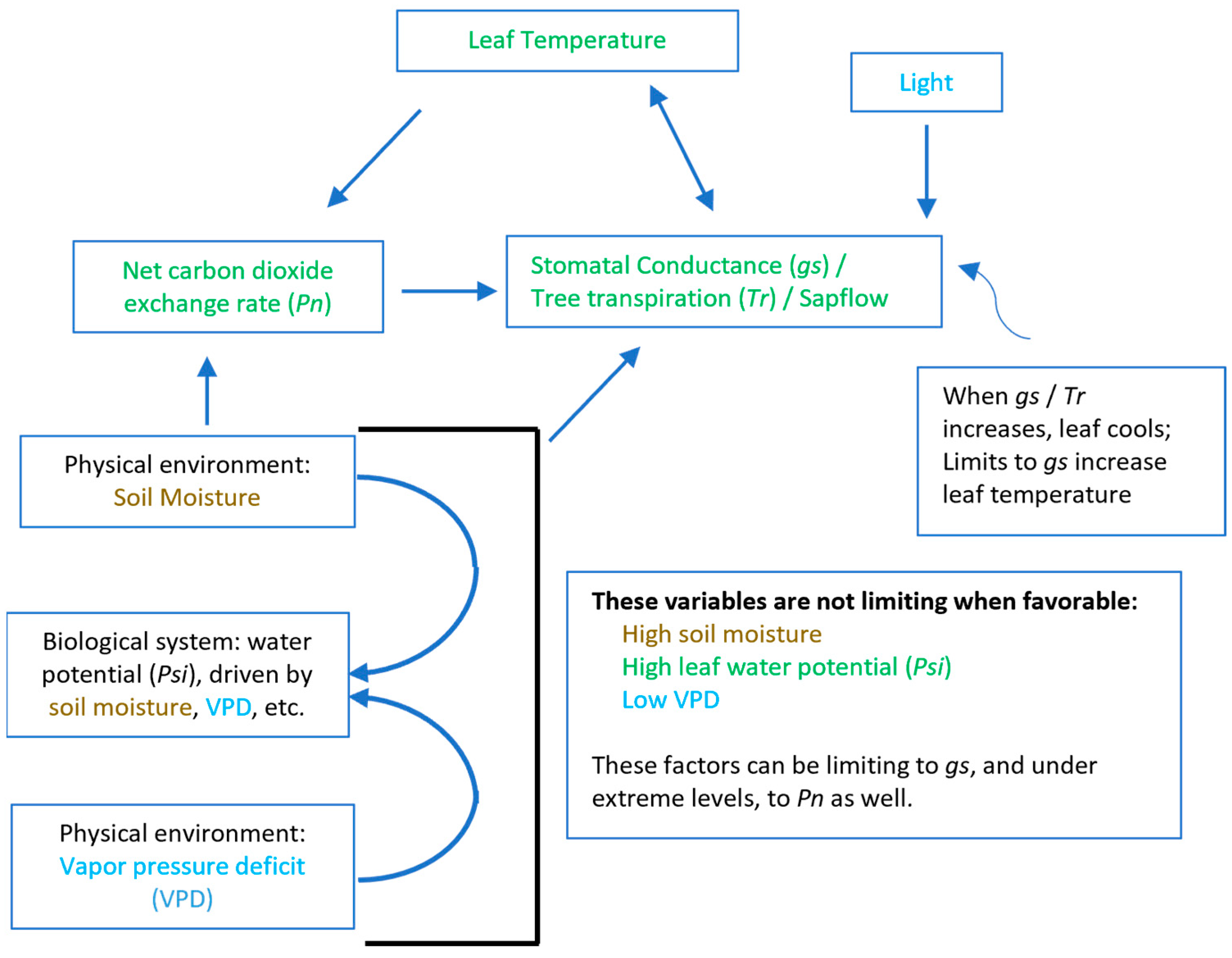
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Branchlet diameter (BRDIA)—Diameter measured in mm (±0.01 mm) at the base of the previous year branchlet.
- Branchlet elongation (BRN)—Measured length of the current year branchlet (±2.0 mm) (BRN1) or the previous year’s branchlet (BRN2). BRN1% is the percentage of the current year’s branchlet length relative to the maximum branchlet length of all annual branchlet length segments present on the branch.
- Chlorosis (CHL)—Chlorosis level of needles expressed as a percent of healthy, green needles in the same age class by ocular estimation. Age class is indicated by integer, e.g., CHL2 is the chlorosis level for the previous year’s needle age class.
- Insect and disease (DISEASE)—a sum of the frequency of abiotic and biotic vectors on sampled branches in each crown [14,48,49]. Abiotic vectors included needle tip dieback, whole needle dieback, and early needle senescence (in August instead of October), all likely driven by drought stress. Biotic vectors included presence or absence of pine needle weevil, phloem feeder, armored scale, black pineleaf scale, and needle blight (Latin authorities given in [14]).
- Needle whorls (#WHL)—The number of needle ages retained on the branchlet. Ponderosa pine needles are in distinct groups on branchlets; these groups are established annually. See [14] for examples.
- Needle elongation (NLN)—Measured length of the current year needle length (+2.0 mm) (NLN1). NLN2% is the percentage of the current year needle length relative to the maximum needle length retained on the branchlet.
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Fettig, C.J.; Mortenson, L.A.; Bulaon, B.M.; Foulk, P.B. Tree Mortality Following Drought in the Central and Southern Sierra Nevada, California, U.S. For. Ecol. Manag. 2019, 432, 164–178. [Google Scholar] [CrossRef]
- Ganey, J.L.; Vojta, S.C. Tree Mortality in Drought-Stressed Mixed-Conifer and Ponderosa Pine Forests, Arizona, USA. For. Ecol. Manag. 2011, 261, 162–168. [Google Scholar] [CrossRef]
- USDA Forest Service. 2019 Ecosystem Services; USDA Forest Service: Washington, DC, USA, 2019. [Google Scholar]
- Simpson, M. Forested Plant Associations of the Oregon East Cascades; USDA, Forest Service, Pacific Northwest Region: Portland, OR, USA, 2007. [Google Scholar]
- Arno, S.F.; Smith, H.Y.; Krebs, M.A. Old Growth Ponderosa Pine and Western Larch Stand Structures: Influences of Pre-1900 Fires and Fire Exclusion; Forest Service Research Paper; Intermountain Research Station, Forest Service, US Department of Agriculture: Ogden, UT, USA, 1997. [Google Scholar]
- Richardson, D.M.; Rundel, P.W.; Jackson, S.T.; Teskey, R.O.; Aronson, J.; Bytnerowicz, A.; Wingfield, M.J.; Procheş, Ş. Human Impacts in Pine Forests: Past, Present, and Future. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 275–297. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Prichard, S.J.; Hagmann, R.K.; Povak, N.A.; Lake, F.K. Wildfire and Climate Change Adaptation of Western North American Forests: A Case for Intentional Management. Ecol. Appl. 2021, 31, e02432. [Google Scholar] [CrossRef] [PubMed]
- Sherman, L.M.; Anderson, P.D.; Fettig, C.J. Forest Dynamics after Thinning and Fuel Reduction in the Pringle Falls Experimental Forest—Establishment and Early Observations of the Lookout Mountain Thinning and Fuels Reduction Study; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2023. [Google Scholar]
- Graham, R.; Jain, T. Ponderosa Pine Ecosystems. In Proceedings of the Symposium on Ponderosa Pine: Issues, Trends, and Management, Klamath Falls, OR, USA, 18–21 October 2004; Ritchie, M.W., Maguire, D.A., Youngblood, A., tech. coordinators, Eds.; Gen. Tech. Rep PSW-GTR-198. USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2005; pp. 1–32. [Google Scholar]
- Agee, J. Fire Ecology of Pacific Northwest Forests. In The Bark Beetles, Fuels, and Fire Bibliography; Island Press: Washington, DC, USA, 1996. [Google Scholar]
- Busse, M.D.; Cochran, P.H.; Hopkins, W.E.; Johnson, W.H.; Riegel, G.M.; Fiddler, G.O.; Ratcliff, A.W.; Shestak, C.J. Developing Resilient Ponderosa Pine Forests with Mechanical Thinning and Prescribed Fire in Central Oregon’s Pumice Region. Can. J. For. Res. 2009, 39, 1171–1185. [Google Scholar] [CrossRef]
- Zavadilova, I.; Szatniewska, J.; Stojanović, M.; Fleischer, P., Jr.; Vágner, L.; Pavelka, M.; Petrík, P. The Effect of Thinning Intensity on Sap Flow and Growth of Norway Spruce. J. For. Sci. 2023, 69, 205–216. [Google Scholar] [CrossRef]
- Grulke, N.; Bienz, C.; Hrinkevich, K.; Maxfield, J.; Uyeda, K. Quantitative and Qualitative Approaches to Assess Tree Vigor and Stand Health in Dry Pine Forests. For. Ecol. Manag. 2020, 465, 118085. [Google Scholar] [CrossRef]
- Klein, T.; Hoch, G.; Yakir, D.; Körner, C. Drought Stress, Growth and Nonstructural Carbohydrate Dynamics of Pine Trees in a Semi-Arid Forest. Tree Physiol. 2014, 34, 981–992. [Google Scholar] [CrossRef]
- Schrader-Patton, C.; Grulke, N.; Bienz, C. Assessment of Ponderosa Pine Vigor Using Four-Band Aerial Imagery in South Central Oregon: Crown Objects to Landscapes. Forests 2021, 12, 612. [Google Scholar] [CrossRef]
- Javadian, M.; Smith, W.K.; Lee, K.; Knowles, J.F.; Scott, R.L.; Fisher, J.B.; Moore, D.J.P.; van Leeuwen, W.J.D.; Barron-Gafford, G.; Behrangi, A. Canopy Temperature Is Regulated by Ecosystem Structural Traits and Captures the Ecohydrologic Dynamics of a Semiarid Mixed Conifer Forest Site. J. Geophys. Res. Biogeosci. 2022, 127, e2021JG006617. [Google Scholar] [CrossRef]
- Grulke, N.; Maxfield, J.; Riggan, P.; Schrader-Patton, C. Pre-Emptive Detection of Mature Pine Drought Stress Using Multispectral Aerial Imagery. Remote Sens. 2020, 12, 2338. [Google Scholar] [CrossRef]
- Reid, A.M.; Chapman, W.K.; Prescott, C.E.; Nijland, W. Using Excess Greenness and Green Chromatic Coordinate Colour Indices from Aerial Images to Assess Lodgepole Pine Vigour, Mortality and Disease Occurrence. For. Ecol. Manag. 2016, 374, 146–153. [Google Scholar] [CrossRef]
- Brown, H.T.; Escombe, F. Researches on Some of the Physiological Processes of Green Leaves, with Special Reference to the Interchange of Energy between the Leaf and Its Surroundings. Proc. R. Soc. Lond. Ser. B Contain. Pap. Biol. Character 1997, 76, 29–111. [Google Scholar] [CrossRef]
- Keen, F.P. Ponderosa Pine Tree Classes Redefined. J. For. 1943, 41, 249–253. [Google Scholar]
- Weber, F.P.; Polcyn, F.C. Remote Sensuin to Detect Stress in Forests. Photogramm. Eng. Remote Sens. 1972, 38, 163–175. [Google Scholar]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A. A Review of Vegetation Indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Yang, B.; Knyazikhin, Y.; Lin, Y.; Yan, K.; Chen, C.; Park, T.; Choi, S.; Mõttus, M.; Rautiainen, M.; Myneni, R.B.; et al. Analyses of Impact of Needle Surface Properties on Estimation of Needle Absorption Spectrum: Case Study with Coniferous Needle and Shoot Samples. Remote Sens. 2016, 8, 563. [Google Scholar] [CrossRef]
- Williams, D.L. A Comparison of Spectral Reflectance Properties at the Needle, Branch, and Canopy Level for Selected Conifer Species. Remote Sens. Environ. 1991, 35, 79–93. [Google Scholar] [CrossRef]
- Leuzinger, S.; Körner, C.; Leuzinger, S.; Korner, C. Tree Species Diversity Affects Canopy Leaf Temperatures in a Mature Temperate Forest. Agric. For. Meteorol. 2007, 146, 29–37. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Cihlar, J.; Ly, H.; Li, Z.; Chen, J.; Pokrant, H.; Huang, F. Multitemporal, Multichannel AVHRR Data Sets for Land Biosphere Studies—Artifacts and Corrections. Remote Sens. Environ. 1997, 60, 35–57. [Google Scholar] [CrossRef]
- Goward, S.N.; Markham, B.; Dye, D.G.; Dulaney, W.; Yang, J. Normalized Difference Vegetation Index Measurements from the Advanced Very High Resolution Radiometer. Remote Sens. Environ. 1991, 35, 257–277. [Google Scholar] [CrossRef]
- Sellers, P.J. Canopy Reflectance, Photosynthesis and Transpiration. Int. J. Remote Sens. 1985, 6, 1335–1372. [Google Scholar] [CrossRef]
- Wong, C.; D’Odorico, P.; Bhathena, Y.; Arain, M.; Ensminger, I. Carotenoid Based Vegetation Indices for Accurate Monitoring of the Phenology of Photosynthesis at the Leaf-Scale in Deciduous and Evergreen Trees. Remote Sens. Environ. 2019, 233, 111407. [Google Scholar] [CrossRef]
- Garbulsky, M.F.; Peñuelas, J.; Ogaya, R.; Filella, I. Leaf and Stand-Level Carbon Uptake of a Mediterranean Forest Estimated Using the Satellite-Derived Reflectance Indices EVI and PRI. Int. J. Remote Sens. 2013, 34, 1282–1296. [Google Scholar] [CrossRef]
- Gamon, J.A.; Kovalchuck, O.; Wong, C.Y.S.; Harris, A.; Garrity, S.R. Monitoring Seasonal and Diurnal Changes in Photosynthetic Pigments with Automated PRI and NDVI Sensors. Biogeosciences 2015, 12, 4149–4159. [Google Scholar] [CrossRef]
- Brodrick, P.G.; Asner, G.P. Remotely Sensed Predictors of Conifer Tree Mortality during Severe Drought. Environ. Res. Lett. 2017, 12, 115013. [Google Scholar] [CrossRef]
- Eitel, J.U.H.; Vierling, L.A.; Litvak, M.E.; Long, D.S.; Schulthess, U.; Ager, A.A.; Krofcheck, D.J.; Stoscheck, L. Broadband, Red-Edge Information from Satellites Improves Early Stress Detection in a New Mexico Conifer Woodland. Remote Sens. Environ. 2011, 115, 3640–3646. [Google Scholar] [CrossRef]
- Jones, H.G. Application of Thermal Imaging and Infrared Sensing in Plant Physiology and Ecophysiology. In Incorporating Advances in Plant Pathology; Advances in Botanical Research; Academic Press: New York, NY, USA, 2004; Volume 41, pp. 107–163. [Google Scholar]
- Fuchs, M.; Tanner, C.B. Infrared Thermometry of Vegetation1. Agron. J. 1966, 58, 597–601. [Google Scholar] [CrossRef]
- Jones, H.G. Use of Infrared Thermometry for Estimation of Stomatal Conductance as a Possible Aid to Irrigation Scheduling. Agric. For. Meteorol. 1999, 95, 139–149. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Ino, T.; Kramer, P.J.; Naylor, A.W.; Strain, B.R. Dynamic Analysis of Water Stress of Sunflower Leaves by Means of a Thermal Image Processing System 1. Plant Physiol. 1984, 76, 266–269. [Google Scholar] [CrossRef]
- Still, C.; Powell, R.; Aubrecht, D.; Kim, Y.; Helliker, B.; Roberts, D.; Richardson, A.D.; Goulden, M. Thermal Imaging in Plant and Ecosystem Ecology: Applications and Challenges. Ecosphere 2019, 10, e02768. [Google Scholar] [CrossRef]
- Kim, Y.; Still, C.J.; Roberts, D.A.; Goulden, M.L. Thermal Infrared Imaging of Conifer Leaf Temperatures: Comparison to Thermocouple Measurements and Assessment of Environmental Influences. Agric. For. Meteorol. 2018, 248, 361–371. [Google Scholar] [CrossRef]
- Sankey, T.; Tatum, J. Thinning Increases Forest Resiliency during Unprecedented Drought. Sci. Rep. 2022, 12, 9041. [Google Scholar] [CrossRef]
- Kuenzer, C.; Guo, H.; Ottinger, M.; Zhang, J.; Dech, S. Spaceborne Thermal Infrared Observation—An Overview of Most Frequently Used Sensors for Applied Research. In Thermal Infrared Remote Sensing: Sensors, Methods, Applications; Springer: Berlin/Heidelberg, Germany, 2013; pp. 131–148. [Google Scholar]
- Hook, S.J. ECOSTRESS, SBG, and HyTES: Status and Results. In Proceedings of the International Workshop on High Resolution Thermal EO, Frascati, Italy, 10–12 May 2023. [Google Scholar]
- Volland, L.A. Plant Associations of the Central Oregon Pumice Zone; USDA Forest Service, Pacific Northwest Region: Portland, OR, USA, 1988. [Google Scholar]
- Cochran, P.; Geist, J.; Clemens, D.; Clausnitzer, R.; Powell, D. Suggested Stocking Levels for Forest Stands in Northeastern Oregon and Southeastern Washington; Forest Service Research Note; USDA, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1994. [Google Scholar]
- Shaw, R.C. Tree Vigor Response and Competitive Zone Density in Mature Ponderosa Pine; Oregon State University: Corvallis, OR, USA, 2016. [Google Scholar]
- Grulke, N.E.; Lee, E.H. Assessing Visible Ozone-Induced Foliar Injury in Ponderosa Pine. Can. J. For. Res. 1997, 27, 1658–1668. [Google Scholar] [CrossRef]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Granier, A. Evaluation of Transpiration in a Douglas-Fir Stand by Means of Sap Flow Measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef]
- PRISM Climate Group. Parameter-Elevation Regressions on Independent Slopes Model; Oregon State University: Corvallis, OR, USA, 2023. [Google Scholar]
- ESRI ArcGIS Pro, v 3.1.2; Environmental Systems Research Institute: Redlands, CA, USA, 2023.
- Pix4D Pix4Dmapper, v 4.6.3; Pix4d SA: Prilly, Switzerland, 2020.
- Teledyne FLIR FLIR Thermal Studio Suite, v 1.9.10; Teledyne FLIR LLC: Wilsonville, OR, USA, 2020.
- Agisoft Agisoft Metashape Professional Edition, Version 2.1; Agisoft LLC: St. Petersburg, Russia, 2020.
- Stimson, H.C.; Breshears, D.D.; Ustin, S.L.; Kefauver, S.C. Spectral Sensing of Foliar Water Conditions in Two Co-Occurring Conifer Species: Pinus Edulis and Juniperus Monosperma. Remote Sens. Environ. 2005, 96, 108–118. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Wong, C.Y.S.; Ensminger, I.; Garrity, S.; Hollinger, D.Y.; Noormets, A.; Peñuelas, J. A Remotely Sensed Pigment Index Reveals Photosynthetic Phenology in Evergreen Conifers. Proc. Natl. Acad. Sci. USA 2016, 113, 13087–13092. [Google Scholar] [CrossRef]
- Eitel, J.U.H.; Keefe, R.F.; Long, D.S.; Davis, A.S.; Vierling, L.A. Active Ground Optical Remote Sensing for Improved Monitoring of Seedling Stress in Nurseries. Sensors 2010, 10, 2843–2850. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Berlyn, G.P. Changes in Foliar Spectral Reflectance and Chlorophyll Fluorescence of Four Temperate Species Following Branch Cutting. Tree Physiol. 2002, 22, 499–506. [Google Scholar] [CrossRef]
- Wong, C.Y.S.; Gamon, J.A. Three Causes of Variation in the Photochemical Reflectance Index (PRI) in Evergreen Conifers. New Phytol. 2015, 206, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Microsoft Microsoft Excel for Microsoft 365 MSO, Version 2311 Build 16.0.17029.20140. 64-Bit. Microsoft Corporation: Redmond, WA, USA, 2023.
- Fisher, J.B.; Baldocchi, D.D.; Misson, L.; Dawson, T.E.; Goldstein, A.H. What the Towers Don’t See at Night: Nocturnal Sap Flow in Trees and Shrubs at Two AmeriFlux Sites in California. Tree Physiol. 2007, 27, 597–610. [Google Scholar] [CrossRef]
- RStudio: Integrated Development for R, RStudio version 2022.12.1+402 software; RStudio Team: Boston, MA USA, 2022.
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Restaino, C.; Young, D.J.N.; Estes, B.; Gross, S.; Wuenschel, A.; Meyer, M.; Safford, H. Forest Structure and Climate Mediate Drought-Induced Tree Mortality in Forests of the Sierra Nevada, USA. Ecol. Appl. 2019, 29, e01902. [Google Scholar] [CrossRef] [PubMed]
- Wilder, B.A.; Kinoshita, A.M. Incorporating ECOSTRESS Evapotranspiration in a Paired Catchment Water Balance Analysis after the 2018 Holy Fire in California. Catena 2022, 215, 106300. [Google Scholar] [CrossRef]
- Li, K.; Guan, K.; Jiang, C.; Wang, S.; Peng, B.; Cai, Y. Evaluation of Four New Land Surface Temperature (LST) Products in the U.S. Corn Belt: ECOSTRESS, GOES-R, Landsat, and Sentinel-3. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 9931–9945. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Scaling-Up Gas Exchange and Energy Balance from the Leaf to the Canopy Level. In Plant Physiological Ecology; Lambers, H., Chapin, F.S., Pons, T.L., Eds.; Springer: New York, NY, USA, 1998; pp. 230–238. ISBN 978-1-4757-2855-2. [Google Scholar]
- Jarvis, P.G.; McNaughton, K.G. Stomatal Control of Transpiration: Scaling Up from Leaf to Region. In Advances in Ecological Research; MacFadyen, A., Ford, E.D., Eds.; Academic Press: New York, NY, USA, 1986; Volume 15, pp. 1–49. ISBN 0065-2504. [Google Scholar]
- De Kauwe, M.G.; Medlyn, B.E.; Knauer, J.; Williams, C.A. Ideas and Perspectives: How Coupled Is the Vegetation\hack\newline to the Boundary Layer? Biogeosciences 2017, 14, 4435–4453. [Google Scholar] [CrossRef]
- Callaway, R.M.; DeLucia, E.H.; Moore, D.; Nowak, R.; Schlesinger, W.H. Competition and Facilitation: Contrasting Effects of Artemisia Tridentata on Desert vs. Montane Pines. Ecology 1996, 77, 2130–2141. [Google Scholar] [CrossRef]
- Steckel, M.; Moser, W.K.; del Río, M.; Pretzsch, H. Implications of Reduced Stand Density on Tree Growth and Drought Susceptibility: A Study of Three Species under Varying Climate. Forests 2020, 11, 627. [Google Scholar] [CrossRef]
- Wong, C.Y.S.; Young, D.J.N.; Latimer, A.M.; Buckley, T.N.; Magney, T.S. Importance of the Legacy Effect for Assessing Spatiotemporal Correspondence between Interannual Tree-Ring Width and Remote Sensing Products in the Sierra Nevada. Remote Sens. Environ. 2021, 265, 112635. [Google Scholar] [CrossRef]
- Colwell, R.N. Determining the prevalence of certain cereal crop diseases by means of aerial photography. Hilgardia 1956, 26, 223–286. [Google Scholar] [CrossRef]
- Carter, G.A.; Knapp, A.K. Leaf Optical Properties in Higher Plants: Linking Spectral Characteristics to Stress and Chlorophyll Concentration. Am. J. Bot. 2001, 88, 677–684. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and Physiological Basis for the Reflectance of Visible and Near-Infrared Radiation from Vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | UMZ | Unit | Pre-BA | Post-BA | Pre-SDI | Post-SDI | Pre-TPH | Post-TPH |
|---|---|---|---|---|---|---|---|---|
| Rx1 | 50 | 41 | 38 | 11 | 270 | 38 | 3884 | 831 |
| Rx2 | 100 | 24 | 43 | 24 | 300 | 140 | 2252 | 473 |
| NoRx | n/a | 25 | 43 | 49 | 282 | 316 | 2252 | 2252 |
| DOY | 1000 h | 1300 h | 1700 h | 24 h Avg. | Avg. Max | Avg. Min | Day Avg | Night Avg |
|---|---|---|---|---|---|---|---|---|
| 232 | 24.09 | 29.31 | 24.38 | 19.71 | 30.98 | 13.42 | 24.71 | 14.70 |
| 234 | 16.43 | 20.89 | 21.26 | 14.42 | 23.48 | 9.11 | 18.53 | 10.32 |
| DOY | VPD Min (hPa) | VPD Max (hPa) | RH Min | RH Max |
|---|---|---|---|---|
| 232 | 2.97 | 32.84 | 23.22 | 92.56 |
| 234 | 0.14 | 14.34 | 50.27 | 107.74 |
| Band Name | Band Center (nm) |
|---|---|
| Blue (B) | 475 ± 20 |
| Green (G) | 560 ± 20 |
| Red (R) | 668 ± 10 |
| Red edge (Re) | 717 ± 10 |
| Near infrared (NIR) | 840 ± 40 |
| Site | DOY | Inspire Micasense | Phantom FLIR | ECOSTRESS |
|---|---|---|---|---|
| Rx1 | 234 | N/A | 11:22, 14:21, 17:01 | 14:04 |
| Rx2 | 232 | 10:14, 12:39, 15:40 | 10:47, 14:17, 16:58 | 09:13 |
| NoRx | 232 | 10:01, 12:22, 15:12 | 10:08, 13:01, 16:03 | 09:13 |
| Index | Formula | Reference |
|---|---|---|
| Normalized Difference Vegetation Index (NDVI) | (NIR − R)/(NIR + R) | [34,56] |
| Chlorophyll/Carotenoid Index (CCI) | (G − R)/(G + R) | [31,57] |
| Normalized Difference Red Edge Index (NDRE) | (NIR − Re)/(NIR + Re) | [35,58] |
| Site | #WHL | BRDIA | BRN% | NL% | CHL1 | CHL2 | CHL4 | DISEASE |
|---|---|---|---|---|---|---|---|---|
| Rx1 | 6.5 (0.2) | 10.3 (0.6) | 59.7 (4.3) | 67.5 (3.8) | 26.8 (7.5) | 21.4 (5.7) | 27.9 (7.2) | 0.4 (0.0) |
| Rx2 | 6.5 (0.1) | 10.0 (0.4) | 67.3 (2.9) | 74.7 (1.5) | 20.1 (3.8) | 13.2 (2.2) | 18.4 (1.7) | 0.3 (0.0) |
| NoRx | 6.6 (0.2) | 9.8 (0.5) | 66.3 (5.4) | 83.4 (3.3) | 20.4 (2.5) | 6.6 (1.9) | 17.7 (6.6) | 0.3 (0.1) |
| All Rx | 6.5 (0.1) | 10.1 (0.3) | 64.2 (2.4) | 74.0 (231) | 22.7 (3.2) | 14.7 (2.6) | 21.9 (3.2) | 0.3 (0.0) |
| Site (I) | Mean | Max | Min | SE | Stand (J) | Mean Difference | p-Value |
|---|---|---|---|---|---|---|---|
| Rx1 | 1.92 | 13.3 | 0 | 0.108 | Rx2 | 0.10 | 0.779 |
| NoRx | 0.42 | 0.009 | |||||
| Rx2 | 1.82 | 9.41 | 0 | 0.115 | Rx1 | −0.10 | 0.779 |
| NoRx | 0.32 | 0.076 | |||||
| NoRx | 1.50 | 8.94 | 0 | 0.086 | Rx1 | −0.42 | 0.009 |
| Rx2 | −0.32 | 0.076 |
| Site (I) | DOY | Mean °C | SE | Stand (J) | Mean Difference | p-Value |
|---|---|---|---|---|---|---|
| Rx1 | 234 | 14.1 | 1.1 | Rx2 | −5.14 | <0.0001 |
| NoRx | −15.2 | <0.0001 | ||||
| Rx2 | 232 | 19.2 | 0.50 | Rx1 | −5.14 | <0.0001 |
| NoRx | −10.0 | 0.003 | ||||
| NoRx | 232 | 29.3 | 0.73 | Rx1 | 15.2 | <0.0001 |
| Rx2 | 10.0 | 0.003 |
| Site (I) | DOY | Mean °C | SE | Stand (J) | Mean Difference | p-Value |
|---|---|---|---|---|---|---|
| Rx1 | 232 | 25.9 | 0.28 | Rx2 | 6.03 | <0.0001 |
| NoRx | 7.20 | <0.0001 | ||||
| Rx2 | 232 | 19.9 | 0.13 | Rx1 | −6.03 | <0.0001 |
| NoRx | 1.16 | 0.0009 | ||||
| NoRx | 232 | 18.7 | 0.03 | Rx1 | −7.20 | <0.0001 |
| Rx2 | −1.16 | 0.0009 |
| Coefficient | Estimate | SE | t | p-Value |
|---|---|---|---|---|
| (Intercept) | 14.4 | 0.984 | <0.001 | 0.982 |
| Etrans | 1093 | 516 | 2.12 | 0.039 |
| CZD | −0.01 | 0.031 | −0.375 | 0.709 |
| Site Rx2 | 10.1 | 1.27 | 7.96 | <0.001 |
| Site NoRx | 15.3 | 2.20 | 6.93 | <0.001 |
| Residual SE: 3.302 Degrees of freedom: 55 F-Statistic: 54.12 AIC: 146.3 | Multiple R2: 0.797 Adjusted R2: 0.783 p-value: <0.001 | |||
| Coefficient | Estimate | SE | t | p-Value |
|---|---|---|---|---|
| (Intercept) | 19.2 | 5.09 | 3.78 | <0.001 |
| NDVI | −14.5 | 15.3 | −0.95 | 0.350 |
| NDRE | 41.9 | 16.0 | 2.61 | 0.013 |
| CCI | 63.3 | 18.5 | 3.41 | 0.001 |
| B | 0.002 | 0.002 | 1.07 | 0.293 |
| NIR | −0.003 | 0.002 | −1.85 | 0.072 |
| Residual SE: 2.2 Degrees of freedom: 40 F-Statistic: 5.26 AIC: 211.00 | Multiple R2: 0.397 Adjusted R2: 0.321 p-value: 0.0008 | |||
| Coefficient | Estimate | SE | t | p-Value |
|---|---|---|---|---|
| (Intercept) | −18.15 | 4.860 | −3.734 | <0.001 |
| UAV FLIR (Td) | 0.432 | 0.158 | 3.856 | 0.010 |
| ECOSTRESS (Tes) | 0.636 | 0.165 | 3.856 | <0.001 |
| CZD | 0.040 | 0.029 | 1.415 | 0.167 |
| Site Rx2 | −2.55 | 1.458 | −1.750 | 0.0891 |
| Site NoRx | −7.396 | 3.178 | −2.327 | 0.026 |
| Residual SE: 2.279 Degrees of freedom: 34 F-Statistic: 4.298 AIC: 186.91 | Multiple R2: 0.387 Adjusted R2: 0.297 p-value: 0.004 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrader-Patton, C.; Grulke, N.E.; Anderson, P.D.; Chaitman, J.; Webb, J. Assessing Tree Water Balance after Forest Thinning Treatments Using Thermal and Multispectral Imaging. Remote Sens. 2024, 16, 1005. https://doi.org/10.3390/rs16061005
Schrader-Patton C, Grulke NE, Anderson PD, Chaitman J, Webb J. Assessing Tree Water Balance after Forest Thinning Treatments Using Thermal and Multispectral Imaging. Remote Sensing. 2024; 16(6):1005. https://doi.org/10.3390/rs16061005
Chicago/Turabian StyleSchrader-Patton, Charlie, Nancy E. Grulke, Paul D. Anderson, Jamieson Chaitman, and Jeremy Webb. 2024. "Assessing Tree Water Balance after Forest Thinning Treatments Using Thermal and Multispectral Imaging" Remote Sensing 16, no. 6: 1005. https://doi.org/10.3390/rs16061005