
Quantifying the Loss of Coral from a Bleaching Event Using Underwater Photogrammetry and AI-Assisted Image Segmentation

 , , , ,
, , , ,  , ,
, ,  ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Reference Network Establishment, Image Acquisition, and Orthophotomosaic Generation
2.3. AI-Assisted Image Segmentation and Manual Validation and Editing
2.4. Analyses and Impact Assessment
3. Results
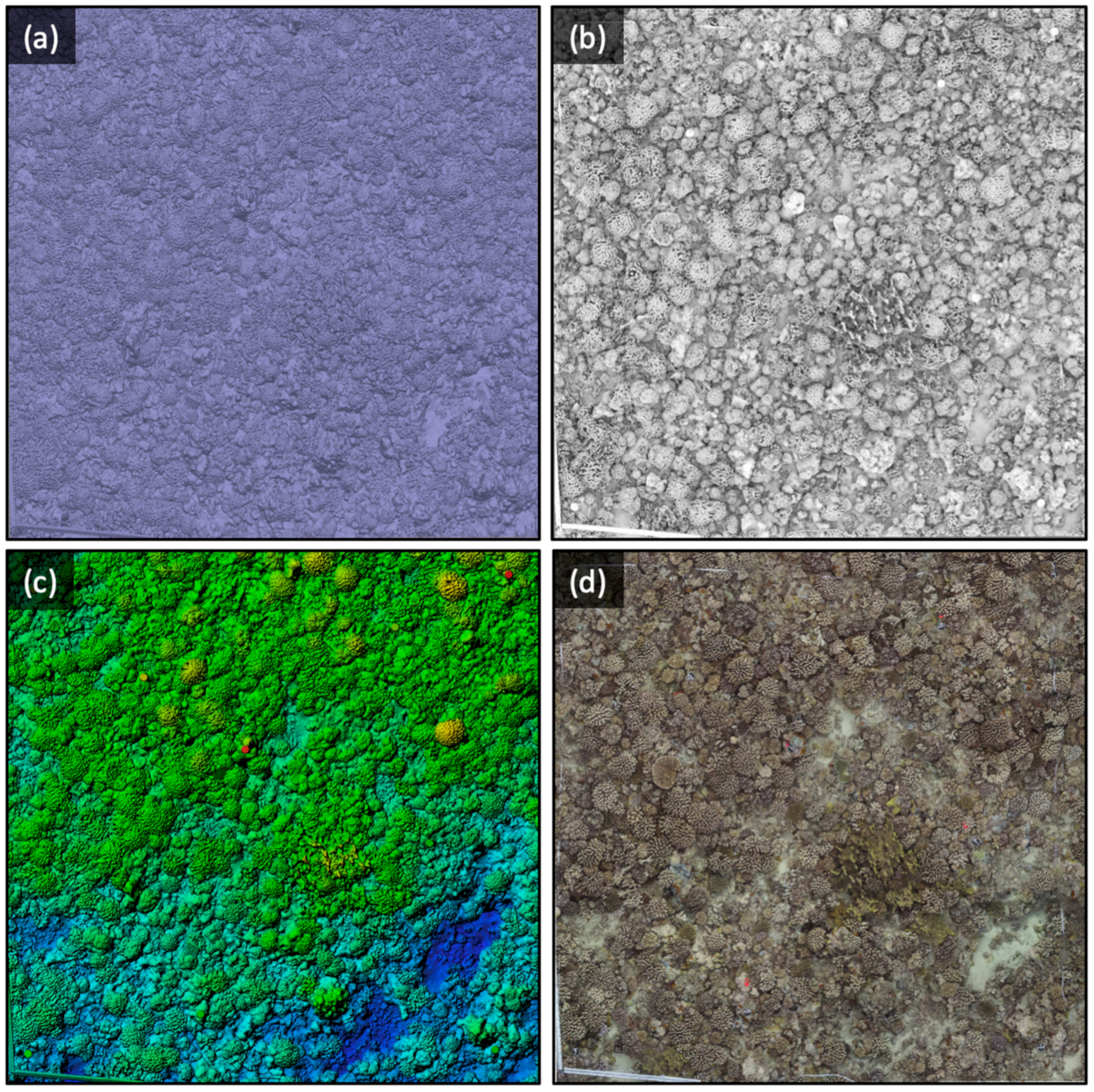
3.1. Automatic Segmentation and Manual Validation/Improvement
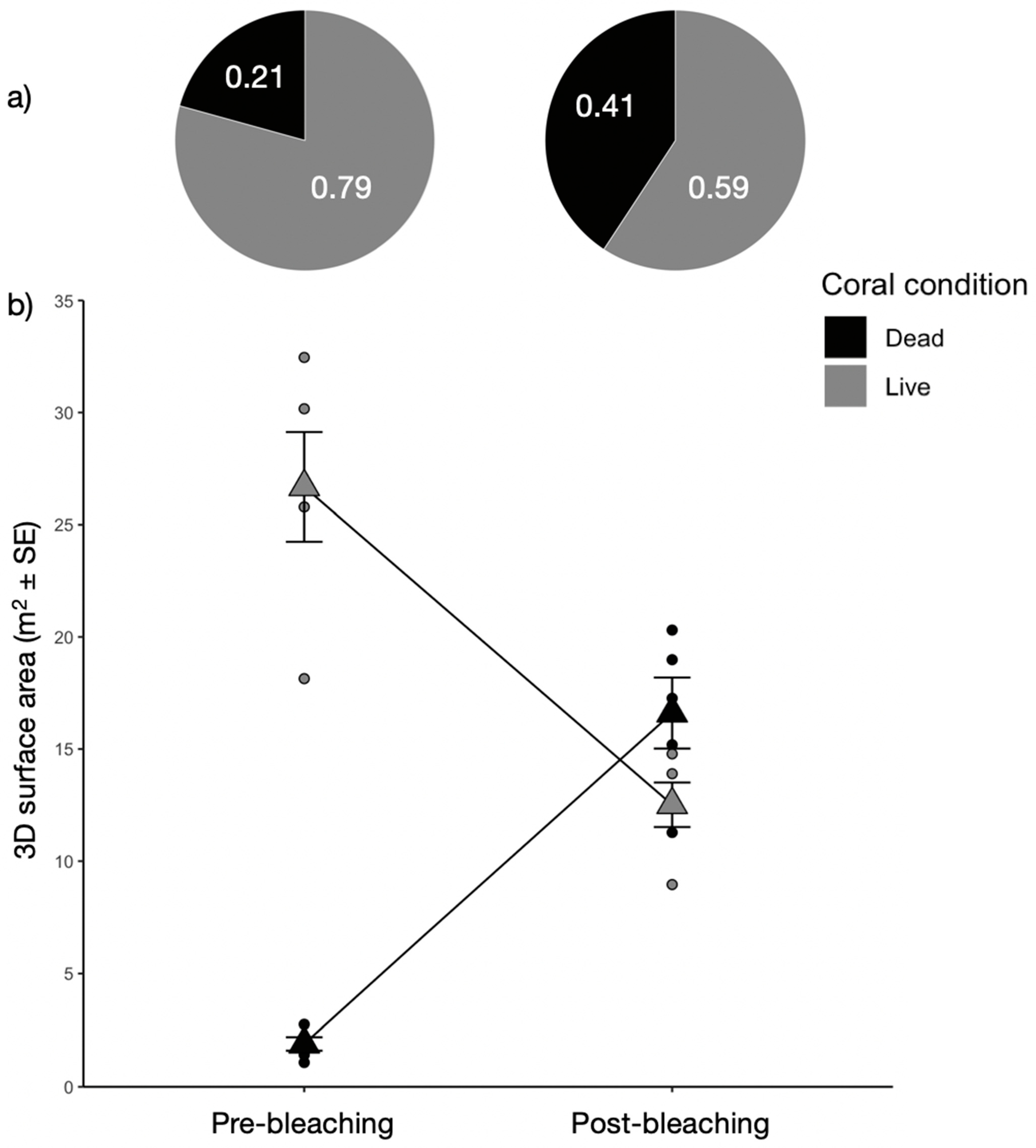
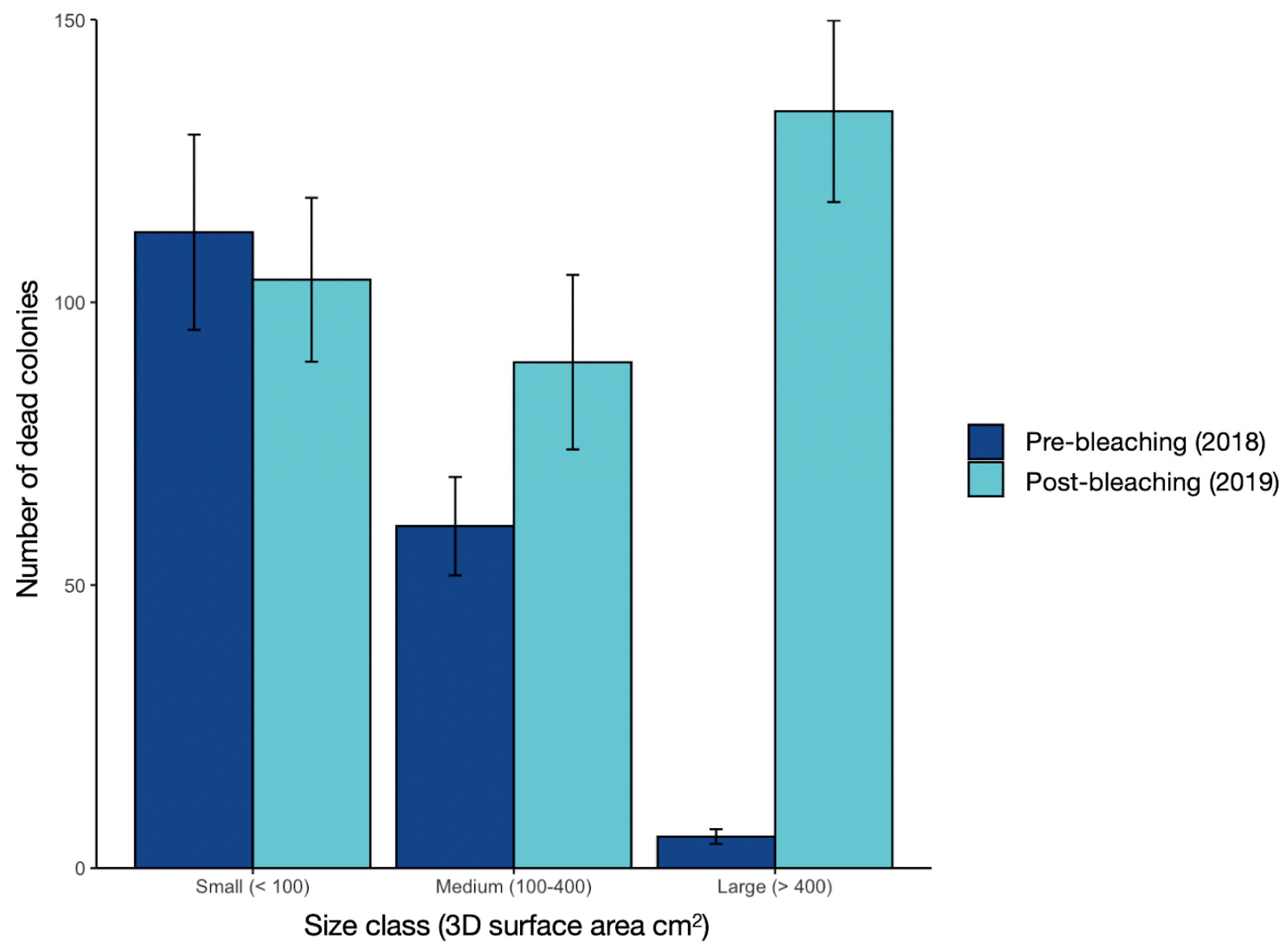
3.2. Changes in Colony Numbers, Approximated 3D Surface Areas, and Size Structure of Corals
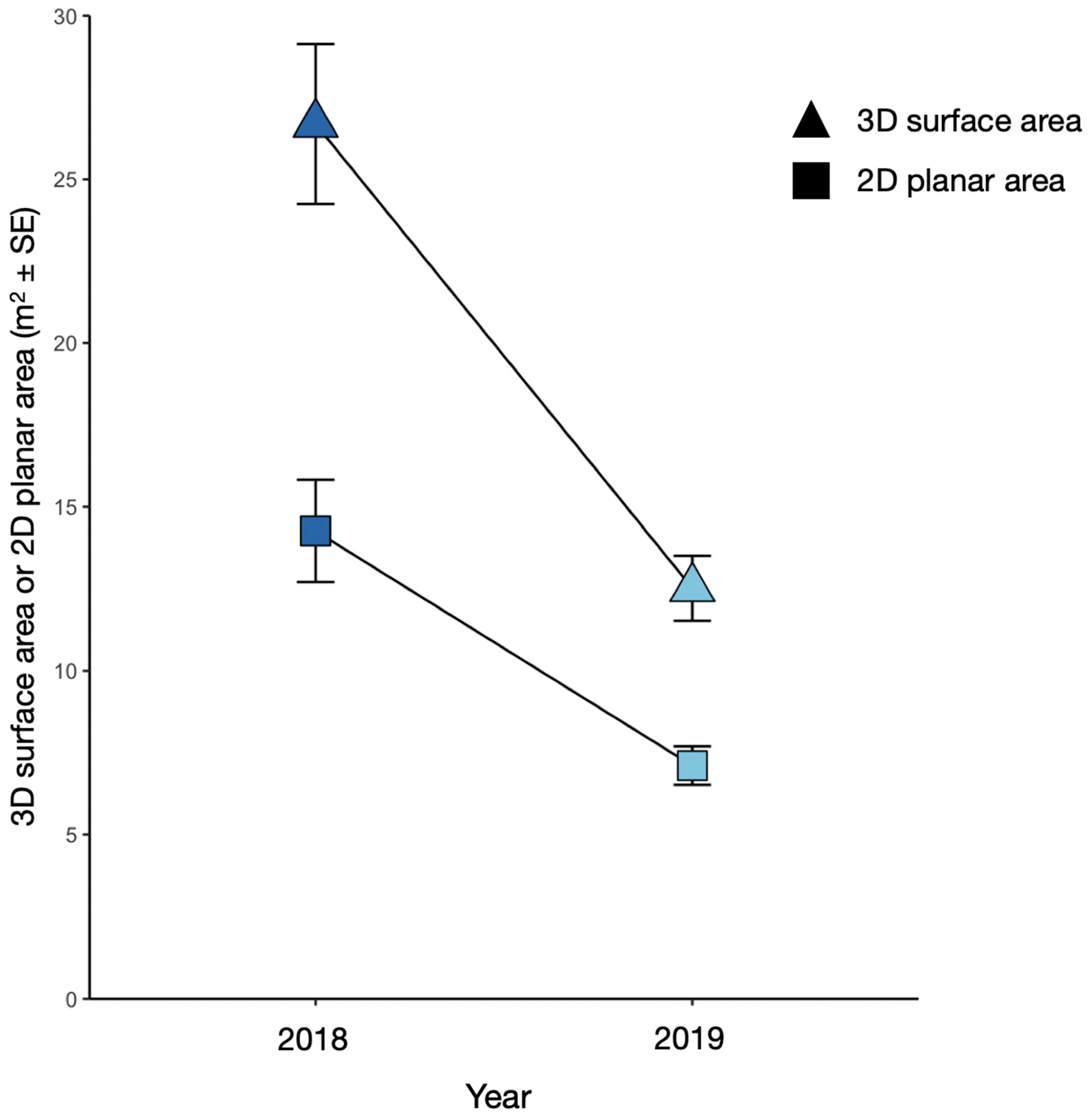
3.3. Comparison of Approximated 3D Surface Area with 2D Planar Area
4. Discussion
4.1. Quantification of the Coral Bleaching Event
4.2. Advantages to Our Approach
4.3. Caveats to Our Approach
4.4. Future Directions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | DF | Sum of Squares | Mean Square | F Value | Pr > F |
|---|---|---|---|---|---|
| Model | 3 | 1027.918483 | 342.639494 | 28.18 | <0.0001 |
| Error | 16 | 194.538179 | 12.158636 | ||
| Corrected Total | 19 | 1222.456661 | |||
| R-Square | Coeff Var | Root MSE | Area Mean | ||
| 0.840863 | 23.02680 | 3.486924 | 15.14290 | ||
| Source | DF | Type I SS | Mean Square | F Value | Pr > F |
| Area_Type | 1 | 397.6459238 | 397.6459238 | 32.70 | <0.0001 |
| Time_Point | 1 | 568.8996018 | 568.8996018 | 46.79 | <0.0001 |
| Time_Point *Area_Type | 1 | 61.3729571 | 61.3729571 | 5.05 | 0.0391 |
| Source | DF | Type III SS | Mean Square | F Value | Pr > F |
| Area_Type | 1 | 188.7322881 | 188.7322881 | 15.52 | 0.0012 |
| Time_Point | 1 | 568.8996018 | 568.8996018 | 46.79 | <0.0001 |
| Time_Point *Area_Type | 1 | 61.3729571 | 61.3729571 | 5.05 | 0.0391 |
References
- Osenberg, C.W.; Schmitt, R.J. Chapter 1—Detecting Ecological Impacts Caused by Human Activities. In Detecting Ecological Impacts; Schmitt, R.J., Osenberg, C.W., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 3–16. ISBN 978-0-12-627255-0. [Google Scholar]
- Solow, A.R. On Detecting Ecological Impacts of Extreme Climate Events and Why It Matters. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160136. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.D.; Dubel, A.K.; Adam, T.C.; Cook, D.T.; Holbrook, S.J.; Schmitt, R.J.; Rassweiler, A. Using Machine Learning to Achieve Simultaneous, Georeferenced Surveys of Fish and Benthic Communities on Shallow Coral Reefs. Limnol. Oceanogr. Methods 2023, 21, 451–466. [Google Scholar] [CrossRef]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral Reef Biodiversity. In Life in the World’s Oceans; McIntyre, A.D., Ed.; Wiley-Blackwell: Oxford, UK, 2010; pp. 65–78. ISBN 978-1-4443-2550-8. [Google Scholar]
- Urbina-Barreto, I.; Garnier, R.; Elise, S.; Pinel, R.; Dumas, P.; Mahamadaly, V.; Facon, M.; Bureau, S.; Peignon, C.; Quod, J.-P.; et al. Which Method for Which Purpose? A Comparison of Line Intercept Transect and Underwater Photogrammetry Methods for Coral Reef Surveys. Front. Mar. Sci. 2021, 8, 636902. [Google Scholar] [CrossRef]
- Couch, C.S.; Oliver, T.A.; Suka, R.; Lamirand, M.; Asbury, M.; Amir, C.; Vargas-Ángel, B.; Winston, M.; Huntington, B.; Lichowski, F.; et al. Comparing Coral Colony Surveys From In-Water Observations and Structure-From-Motion Imagery Shows Low Methodological Bias. Front. Mar. Sci. 2021, 8, 647943. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral Reefs Under Rapid Climate Change and Ocean Acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- Donovan, M.K.; Burkepile, D.E.; Kratochwill, C.; Shlesinger, T.; Sully, S.; Oliver, T.A.; Hodgson, G.; Freiwald, J.; van Woesik, R. Local Conditions Magnify Coral Loss after Marine Heatwaves. Science 2021, 372, 977–980. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Simpson, T. Large-Scale Bleaching of Corals on the Great Barrier Reef. Ecology 2018, 99, 501. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Connolly, S.R.; Baird, A.H.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; Liu, G.; et al. Ecological Memory Modifies the Cumulative Impact of Recurrent Climate Extremes. Nat. Clim. Chang. 2019, 9, 40–43. [Google Scholar] [CrossRef]
- Lough, J.M.; Anderson, K.D.; Hughes, T.P. Increasing Thermal Stress for Tropical Coral Reefs: 1871–2017. Sci. Rep. 2018, 8, 6079. [Google Scholar] [CrossRef]
- Donovan, M.K.; Adam, T.C.; Shantz, A.A.; Speare, K.E.; Munsterman, K.S.; Rice, M.M.; Schmitt, R.J.; Holbrook, S.J.; Burkepile, D.E. Nitrogen Pollution Interacts with Heat Stress to Increase Coral Bleaching across the Seascape. Proc. Natl. Acad. Sci. USA 2020, 117, 5351–5357. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Schmitt, R.J.; Messmer, V.; Brooks, A.J.; Srinivasan, M.; Munday, P.L.; Jones, G.P. Reef Fishes in Biodiversity Hotspots Are at Greatest Risk from Loss of Coral Species. PLoS ONE 2015, 10, e0124054. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Resistance and Resilience of a Coral Reef Fish Community to Changes in Coral Cover. Mar. Ecol. Prog. Ser. 2008, 371, 263–271. [Google Scholar] [CrossRef]
- Messmer, V.; Jones, G.P.; Munday, P.L.; Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Habitat Biodiversity as a Determinant of Fish Community Structure on Coral Reefs. Ecology 2011, 92, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Adam, T.C.; Schmitt, R.J.; Brooks, A.J.; Holbrook, S.J. Response of Herbivore Functional Groups to Sequential Perturbations in Moorea, French Polynesia. Coral Reefs 2016, 35, 999–1009. [Google Scholar] [CrossRef]
- Adam, T.C.; Brooks, A.J.; Holbrook, S.J.; Schmitt, R.J.; Washburn, L.; Bernardi, G. How Will Coral Reef Fish Communities Respond to Climate-Driven Disturbances? Insight from Landscape-Scale Perturbations. Oecologia 2014, 176, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kopecky, K.L.; Stier, A.C.; Schmitt, R.J.; Holbrook, S.J.; Moeller, H.V. Material Legacies Can Degrade Resilience: Structure-Retaining Disturbances Promote Regime Shifts on Coral Reefs. Ecology 2023, 104, e4006. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, R.J.; Holbrook, S.J.; Davis, S.L.; Brooks, A.J.; Adam, T.C. Experimental Support for Alternative Attractors on Coral Reefs. Proc. Natl. Acad. Sci. USA 2019, 116, 4372–4381. [Google Scholar] [CrossRef]
- Leon, J.X.; Roelfsema, C.M.; Saunders, M.I.; Phinn, S.R. Measuring Coral Reef Terrain Roughness Using ‘Structure-from-Motion’ Close-Range Photogrammetry. Geomorphology 2015, 242, 21–28. [Google Scholar] [CrossRef]
- Casella, E.; Collin, A.; Harris, D.; Ferse, S.; Bejarano, S.; Parravicini, V.; Hench, J.L.; Rovere, A. Mapping Coral Reefs Using Consumer-Grade Drones and Structure from Motion Photogrammetry Techniques. Coral Reefs 2017, 36, 269–275. [Google Scholar] [CrossRef]
- Lirman, D.; Gracias, N.R.; Gintert, B.E.; Gleason, A.C.R.; Reid, R.P.; Negahdaripour, S.; Kramer, P. Development and Application of a Video-Mosaic Survey Technology to Document the Status of Coral Reef Communities. Environ. Monit. Assess. 2007, 125, 59–73. [Google Scholar] [CrossRef]
- Kikuzawa, Y.P.; Toh, T.C.; Ng, C.S.L.; Sam, S.Q.; Taira, D.; Afiq-Rosli, L.; Chou, L.M. Quantifying Growth in Maricultured Corals Using Photogrammetry. Aquac. Res. 2018, 49, 2249–2255. [Google Scholar] [CrossRef]
- El-Khaled, Y.C.; Kler Lago, A.; Mezger, S.D.; Wild, C. Comparative Evaluation of Free Web Tools ImageJ and Photopea for the Surface Area Quantification of Planar Substrates and Organisms. Diversity 2022, 14, 272. [Google Scholar] [CrossRef]
- Rich, W.A.; Carvalho, S.; Cadiz, R.; Gil, G.; Gonzalez, K.; Berumen, M.L. Size Structure of the Coral Stylophora Pistillata across Reef Flat Zones in the Central Red Sea. Sci. Rep. 2022, 12, 13979. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.H.R.; Delparte, D.; Gates, R.D.; Takabayashi, M. Integrating Structure-from-Motion Photogrammetry with Geospatial Software as a Novel Technique for Quantifying 3D Ecological Characteristics of Coral Reefs. PeerJ 2015, 3, e1077. [Google Scholar] [CrossRef] [PubMed]
- Sandin, S.A.; Edwards, C.B.; Pedersen, N.E.; Petrovic, V.; Pavoni, G.; Alcantar, E.; Chancellor, K.S.; Fox, M.D.; Stallings, B.; Sullivan, C.J.; et al. Chapter Seven—Considering the Rates of Growth in Two Taxa of Coral across Pacific Islands. In Advances in Marine Biology; Population Dynamics of the Reef Crisis; Riegl, B.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 87, pp. 167–191. [Google Scholar]
- Zhong, J.; Li, M.; Zhang, H.; Qin, J. Combining Photogrammetric Computer Vision and Segmentation for Fine-grained Understanding of Coral Reef Growth under Climate Change. In Proceedings of the IEEE/CVF Winter Conference on Applications of Computer Vision Workshops (WACVW), Waikoloa, HI, USA, 3–7 January 2023. [Google Scholar]
- Zhang, H.; Gruen, A.; Li, M. Deep Learning for Semantic Segmentation of Coral Images in Underwater Photogrammetry. ISPRS Ann. Photogramm. Remote Sens. Spatial Inf. Sci. 2022, V-2-2022, 343–350. [Google Scholar] [CrossRef]
- Zhang, H.; Li, M.; Pan, X.; Zhang, X.; Zhong, J.; Qin, J. Novel Approaches to Enhance Coral Reefs Monitoring with Underwater Images Segmentation. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2022, XLVI-3/W1-2022, 271–277. [Google Scholar] [CrossRef]
- Nocerino, E.; Menna, F.; Gruen, A.; Troyer, M.; Capra, A.; Castagnetti, C.; Rossi, P.; Brooks, A.J.; Schmitt, R.J.; Holbrook, S.J. Coral Reef Monitoring by Scuba Divers Using Underwater Photogrammetry and Geodetic Surveying. Remote Sens. 2020, 12, 3036. [Google Scholar] [CrossRef]
- Pavoni, G.; Corsini, M.; Ponchio, F.; Muntoni, A.; Edwards, C.; Pedersen, N.; Sandin, S.; Cignoni, P. TagLab: AI-Assisted Annotation for the Fast and Accurate Semantic Segmentation of Coral Reef Orthoimages. J. Field Robot. 2022, 39, 246–262. [Google Scholar] [CrossRef]
- Peter Edmunds MCR LTER: Coral Reef: Long-Term Population and Community Dynamics: Corals, Ongoing since 2005. 2022. Available online: http://mcrlter.msi.ucsb.edu/cgi-bin/showDataset.cgi?docid=knb-lter-mcr.4 (accessed on 1 June 2023).
- Rossi, P.; Castagnetti, C.; Capra, A.; Brooks, A.J.; Mancini, F. Detecting Change in Coral Reef 3D Structure Using Underwater Photogrammetry: Critical Issues and Performance Metrics. Appl. Geomat. 2020, 12, 3–17. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; RDC Team: Vienna, Austria, 2020.
- Posit team RStudio: Integrated Development Environment for R 2022. Available online: https://www.rstudio.com/categories/integrated-development-environment/ (accessed on 1 June 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Geoffrey Thomson Manu: NZ Bird Colour Palettes 2022. Available online: https://github.com/G-Thomson/Manu (accessed on 1 June 2023).
- Speare, K.E.; Adam, T.C.; Winslow, E.M.; Lenihan, H.S.; Burkepile, D.E. Size-Dependent Mortality of Corals during Marine Heatwave Erodes Recovery Capacity of a Coral Reef. Glob. Chang. Biol. 2022, 28, 1342–1358. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.C.; Johnston, E.C.; Wyatt, A.S.J.; Leichter, J.J.; Edmunds, P.J. Hidden Differences in Bleaching Among Cryptic Coral Species. Bull. Ecol. Soc. Am. 2021, 102, e01885. [Google Scholar] [CrossRef]
- Honeycutt, R.N.; Holbrook, S.J.; Brooks, A.J.; Schmitt, R.J. Farmerfish Gardens Help Buffer Stony Corals against Marine Heat Waves. PLoS ONE 2023, 18, e0282572. [Google Scholar] [CrossRef]
- Cunning, R.; Muller, E.B.; Gates, R.D.; Nisbet, R.M. A Dynamic Bioenergetic Model for Coral-Symbiodinium Symbioses and Coral Bleaching as an Alternate Stable State. J. Theor. Biol. 2017, 431, 49–62. [Google Scholar] [CrossRef]
- Connell, J.H.; Hughes, T.P.; Wallace, C.C. A 30-Year Study of Coral Abundance, Recruitment, and Disturbance at Several Scales in Space and Time. Ecol. Monogr. 1997, 67, 461–488. [Google Scholar] [CrossRef]
- House, J.E.; Brambilla, V.; Bidaut, L.M.; Christie, A.P.; Pizarro, O.; Madin, J.S.; Dornelas, M. Moving to 3D: Relationships between Coral Planar Area, Surface Area and Volume. PeerJ 2018, 6, e4280. [Google Scholar] [CrossRef] [PubMed]
- Moberg, F.; Folke, C. Ecological Goods and Services of Coral Reef Ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Indirect Effects of Species Interactions on Habitat Provisioning. Oecologia 2011, 166, 739–749. [Google Scholar] [CrossRef]
- Menna, F.; Nocerino, E.; Chemisky, B.; Remondino, F.; Drap, P. Accurate Scaling and Leveling in Underwater Photogrammetry with a Pressure Sensor. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2021, XLIII-B2-2021, 667–672. [Google Scholar] [CrossRef]
- Nocerino, E.; Menna, F. In-Camera IMU Angular Data for Orthophoto Projection in Underwater Photogrammetry. ISPRS Open J. Photogramm. Remote Sens. 2023, 7, 100027. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopecky, K.L.; Pavoni, G.; Nocerino, E.; Brooks, A.J.; Corsini, M.; Menna, F.; Gallagher, J.P.; Capra, A.; Castagnetti, C.; Rossi, P.; et al. Quantifying the Loss of Coral from a Bleaching Event Using Underwater Photogrammetry and AI-Assisted Image Segmentation. Remote Sens. 2023, 15, 4077. https://doi.org/10.3390/rs15164077
Kopecky KL, Pavoni G, Nocerino E, Brooks AJ, Corsini M, Menna F, Gallagher JP, Capra A, Castagnetti C, Rossi P, et al. Quantifying the Loss of Coral from a Bleaching Event Using Underwater Photogrammetry and AI-Assisted Image Segmentation. Remote Sensing. 2023; 15(16):4077. https://doi.org/10.3390/rs15164077
Chicago/Turabian StyleKopecky, Kai L., Gaia Pavoni, Erica Nocerino, Andrew J. Brooks, Massimiliano Corsini, Fabio Menna, Jordan P. Gallagher, Alessandro Capra, Cristina Castagnetti, Paolo Rossi, and et al. 2023. "Quantifying the Loss of Coral from a Bleaching Event Using Underwater Photogrammetry and AI-Assisted Image Segmentation" Remote Sensing 15, no. 16: 4077. https://doi.org/10.3390/rs15164077