
Enzyme-Assisted Method for Phycobiliproteins Extraction from Porphyra and Evaluation of Their Bioactivity

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Production of Crude Enzyme Solutions from Marine Bacterial Strains
2.3. Determination of pH and Enzyme Activity of Crude Enzyme Solutions
2.4. Process of Phycobiliprotein Extraction from Porphyra
2.5. Purification of Phycobiliproteins by Fast Protein Liquid Chromatography (FPLC)
2.6. Analysis of PE and PC within Fractions
2.7. Antioxidation and Angiotensin-Converting-Enzyme (ACE) Inhibition Assays
2.8. Statistical Analysis
3. Results and Discussion
3.1. Proximate Compositions of Porphyra Powder
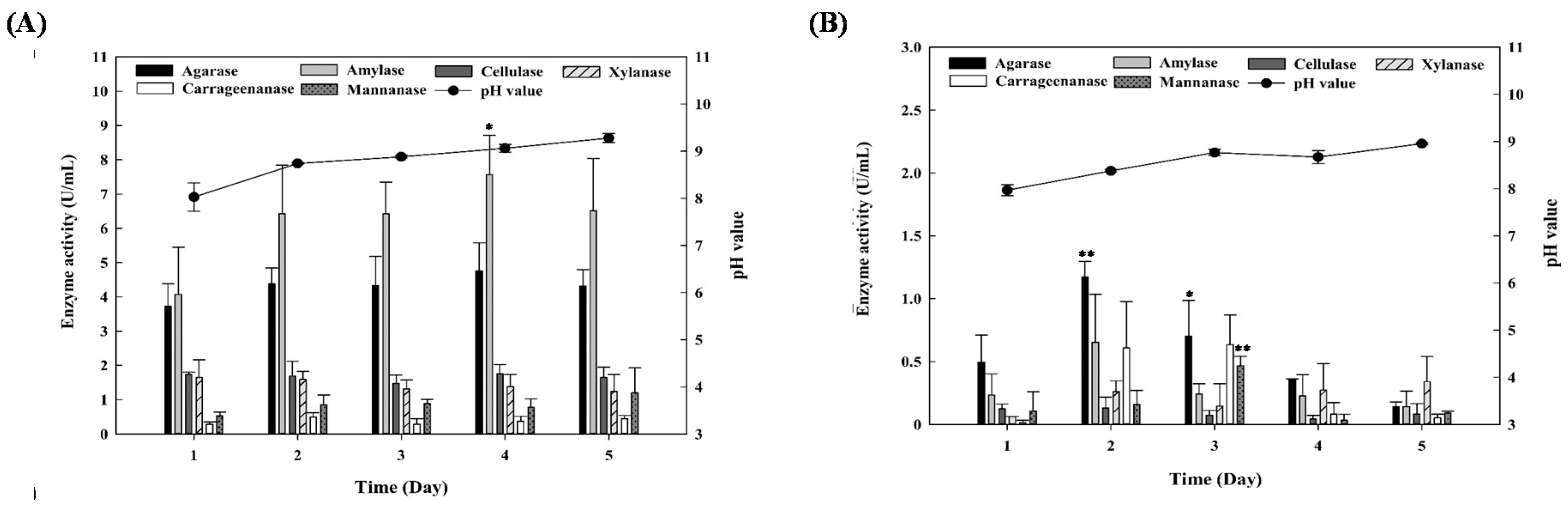
3.2. Enzyme Activity of Crude Enzyme Solutions Produced by Marine Bacterial Strains
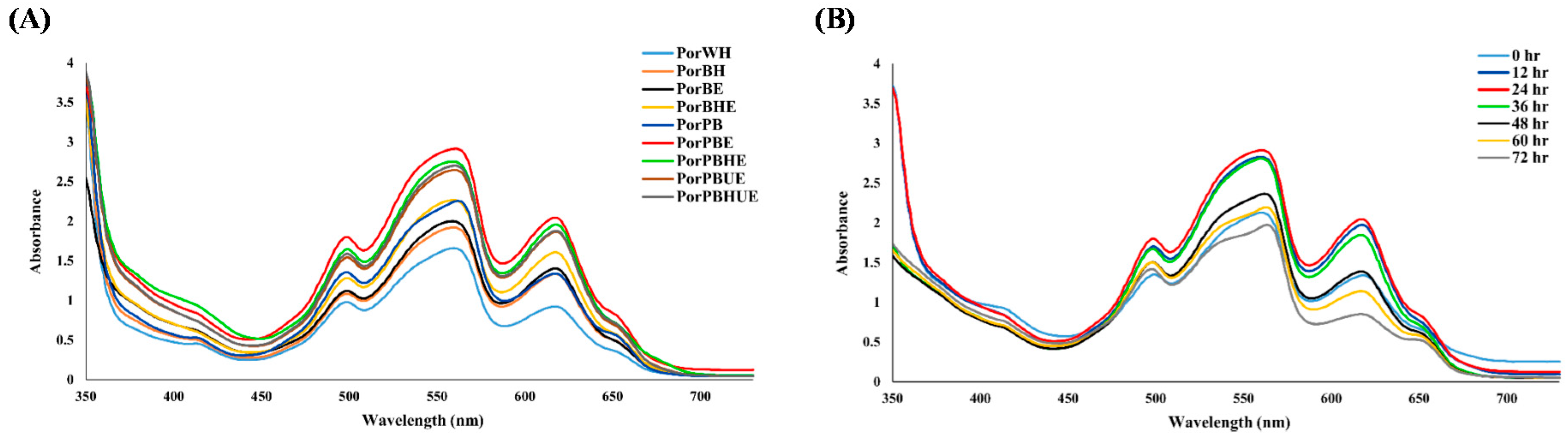
3.3. Impact of Solvent, Homogenization and Ultrasonication on Enzyme-Assisted Extraction of Phycobiliproteins from Dry Porphyra and Porphyra Powder
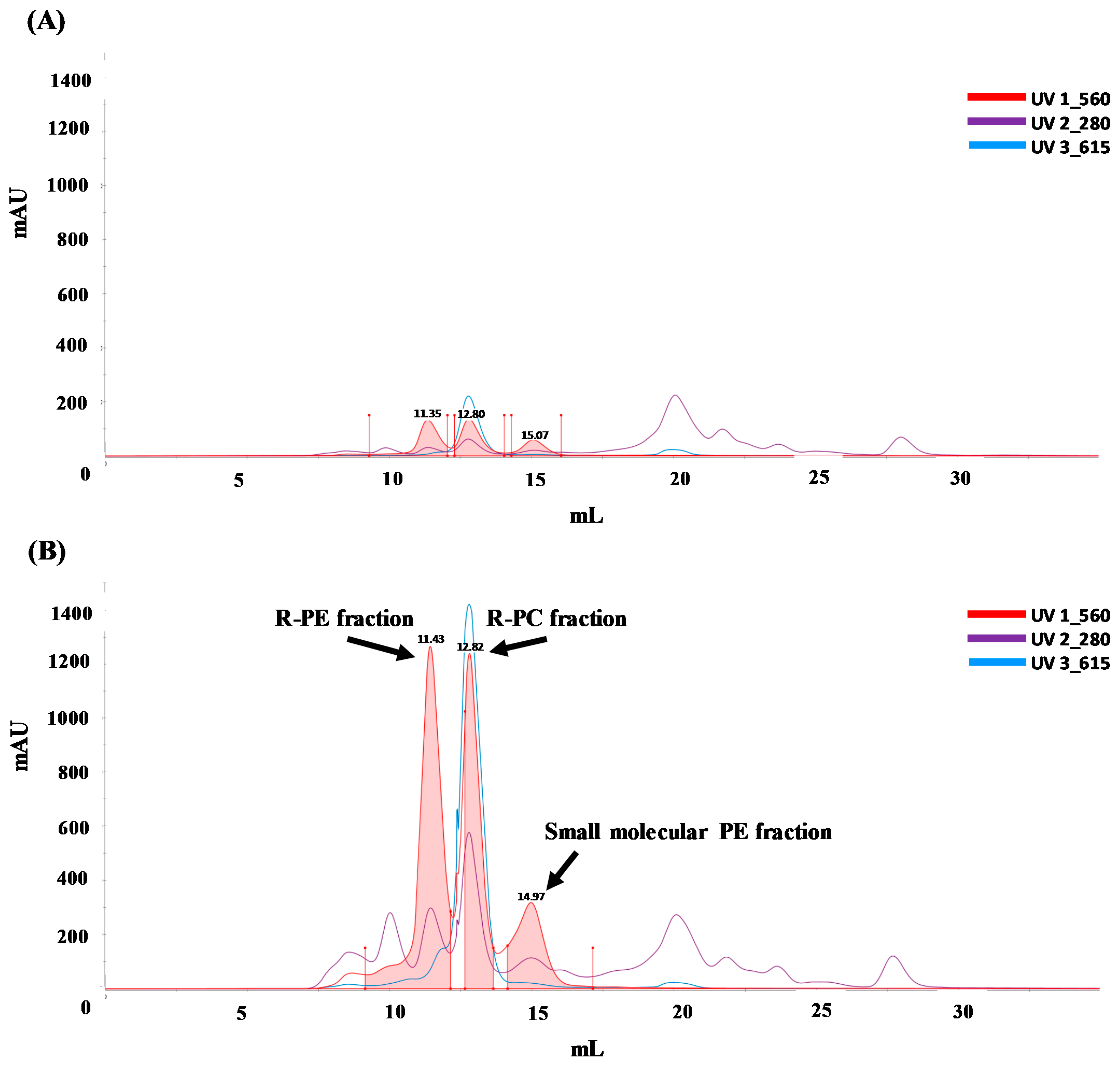
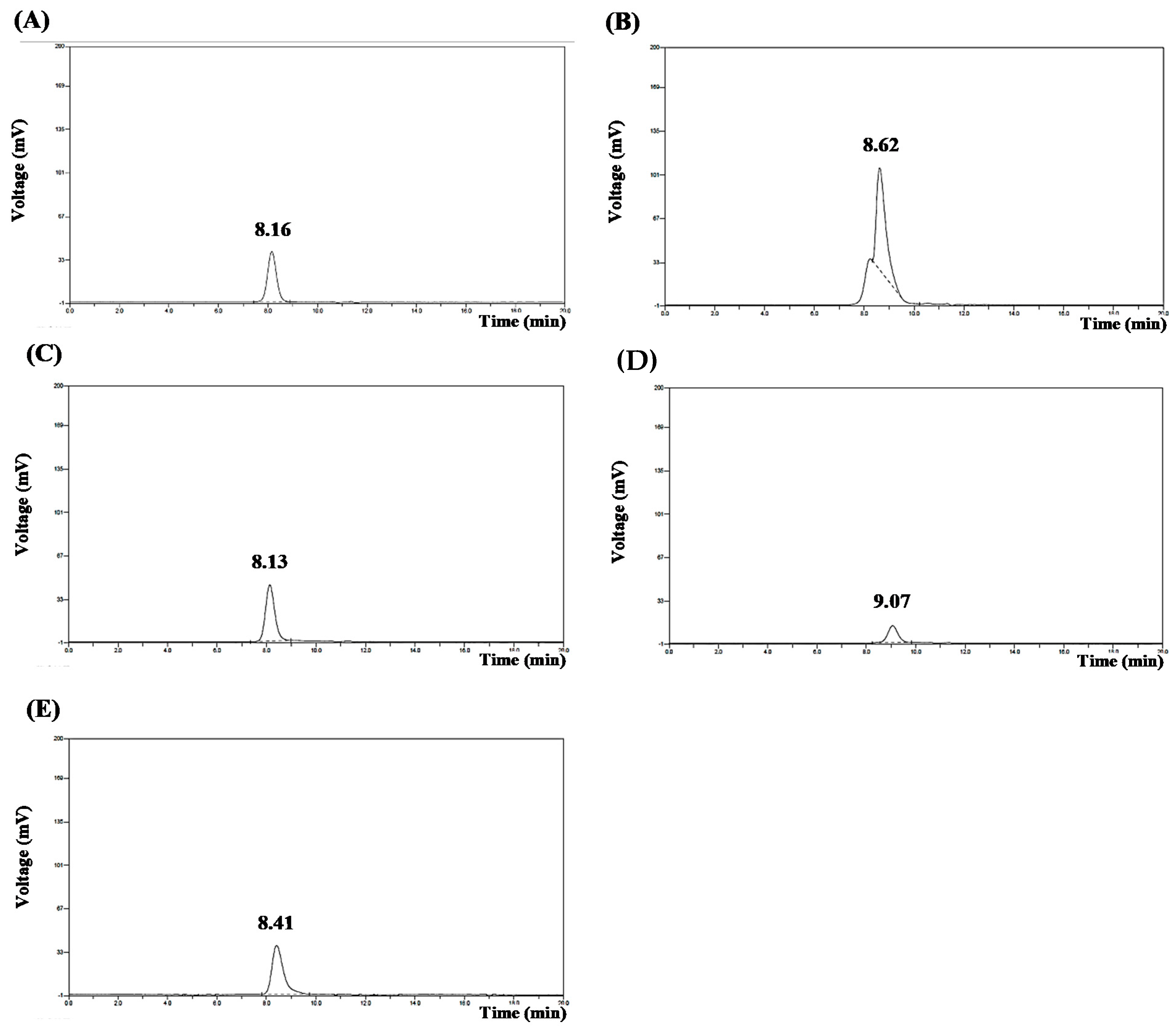
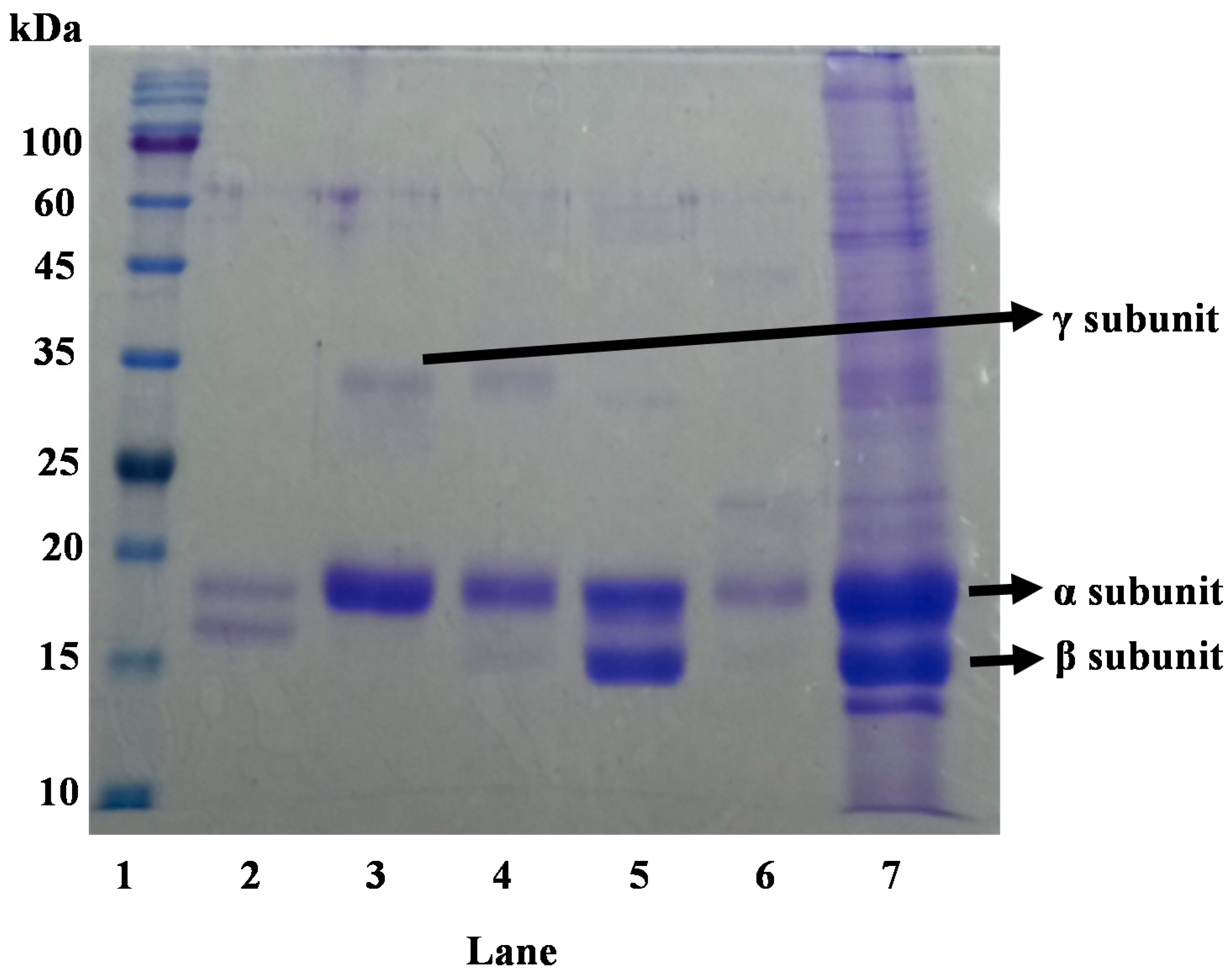
3.4. Purity of PE and PC
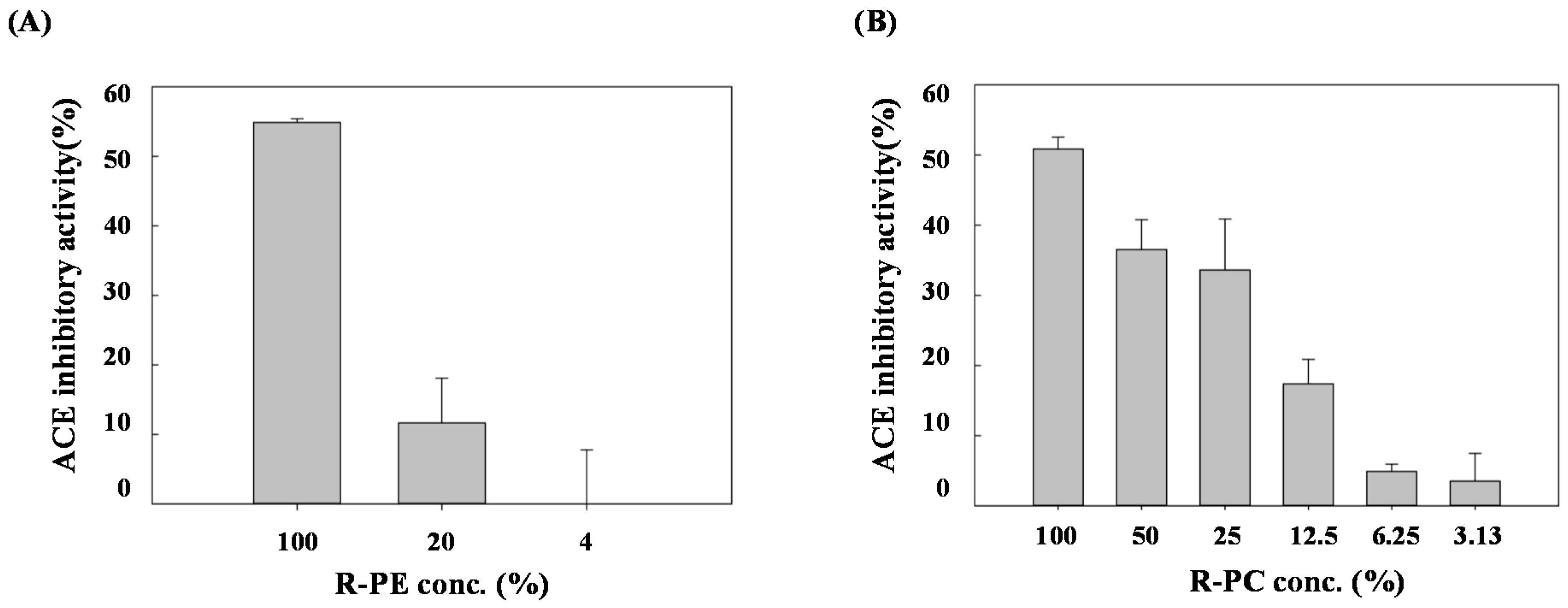
3.5. Antioxidant and ACE Inhibitory Activities of Purified PE and PC
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tiwari, B.K.; Troy, D.J. Seaweed Sustainability: Food and Nonfood Applications; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–6. [Google Scholar]
- Punampalam, R.; Khoo, K.S.; Sit, N. Evaluation of antioxidant properties of phycobiliproteins and phenolic compounds extracted from Bangia atropurpurea. Malays. J. Fundam. Appl. Sci. 2018, 14, 289–297. [Google Scholar] [CrossRef]
- Domínguez, G.D.; Marsán Suárez, V.; del Valle Pérez, L.O. Main immunomodulatory and anti-inflamatory properties of phycobiliproteins C-phycocyanin. Rev. Cuba. Hematol. Inmunol. Hemoter. 2016, 32, 447–454. [Google Scholar]
- Afreen, S.; Fatma, T. Extraction, purification and characterization of phycoerythrin from Michrochaete and its biological activities. Biocatal. Agric. Biotechnol. 2018, 13, 84–89. [Google Scholar]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J.; Levine, I. Seaweed in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Bryant, D.A. Phycoerythrocyanin and phycoerythrin: Properties and occurrence in cyanobacteria. Microbiology 1982, 128, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Zhongzheng, P.; Baicheng, Z.; Chengkui, Z.; Tseng, C. The effect of pH on both spectral types of R-phycoerythrin. Chin. J. Oceanol. Limnol. 1987, 5, 73–79. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M. Algal proteins: Extraction, application, and challenges concerning production. Foods 2017, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Extraction and characterization of protein from Irish brown seaweed Ascophyllum nodosum. Int. Food Res. J. 2017, 99, 1021–1027. [Google Scholar] [CrossRef]
- Fleurence, J. The enzymatic degradation of algal cell walls: A useful approach for improving protein accessibility? J. Appl. Phycol. 1999, 11, 313–314. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Extraction of protein from the macroalga Palmaria palmata. LWT Food Sci. Technol. 2013, 51, 375–382. [Google Scholar] [CrossRef]
- Ganeva, V.; Galutzov, B.; Teissié, J. High yield electroextraction of proteins from yeast by a flow process. Anal. Biochem. 2003, 315, 77–84. [Google Scholar] [CrossRef]
- AOAC. Offical Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Washington, DC, USA, 1998. [Google Scholar]
- Liang, W.-S.; Fang, T.-Y.; Lin, H.-T.; Liu, T.C.; Lu, W.-J.; Tzou, W.-S.; Tang, S.-J.; Lin, F.-P.; Liu, S.-M.; Pan, C.-L. Cloning, expression, and characterization of Pseudomonas vesicularis MA103 β-1, 3-xylanase in Escherichia coli ClearColi BL21 (DE3). Fish. Sci. 2015, 81, 1135–1143. [Google Scholar] [CrossRef]
- Hassairi, I.; Amar, R.B.; Nonus, M.; Gupta, B. Production and separation of α-agarase from Altermonas agarlyticus strain GJ1B. Bioresour. Technol. 2001, 79, 47–51. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, alpha and beta. Methods Enzymol. 1955, 1, 149–158. [Google Scholar]
- Baghel, R.S.; Trivedi, N.; Reddy, C. A simple process for recovery of a stream of products from marine macroalgal biomass. Bioresour. Technol. 2016, 203, 160–165. [Google Scholar] [CrossRef]
- Cian, R.E.; Salgado, P.R.; Drago, S.R.; González, R.J.; Mauri, A.N. Development of naturally activated edible films with antioxidant properties prepared from red seaweed Porphyra columbina biopolymers. Food Chem. 2014, 146, 6–14. [Google Scholar] [CrossRef]
- Munier, M.; Jubeau, S.; Wijaya, A.; Morancais, M.; Dumay, J.; Marchal, L.; Jaouen, P.; Fleurence, J. Physicochemical factors affecting the stability of two pigments: R-phycoerythrin of Grateloupia turuturu and B-phycoerythrin of Porphyridium cruentum. Food Chem. 2014, 150, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, R.; Acién, F.G.; Ibáñez, M.J.; Fernández, J.M.; Molina, E.; Alvarez-Pez, J.M. Preparative purification of B-phycoerythrin from the microalga Porphyridium cruentum by expanded-bed adsorption chromatography. J. Chromatogr. B 2003, 790, 317–325. [Google Scholar] [CrossRef]
- Ma, S.-Y.; Wang, G.-C.; Sun, H.-B.; Zeng, C.-K. Characterization of the artificially covalent conjugate of B-phycoerythrin and R-phycocyanin and the phycobilisome from Porphyridium cruentum. Plant Sci. 2003, 164, 253–257. [Google Scholar] [CrossRef]
- Kazłowski, B.; Pan, C.L.; Ko, Y.T. Monitoring and preparation of neoagaro-and agaro-oligosaccharide products by high performance anion exchange chromatography systems. Carbohydr. Polym. 2015, 122, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Fu, X.; Sun, L. Characteristics of an R-Phycoerythrin with two γ subunits prepared from red macroalga Polysiphonia urceolata. PLoS ONE 2015, 10, e0120333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arndt, C.; Koristka, S.; Bartsch, H.; Bachmann, M. Native polyacrylamide gels. In Protein Electrophoresis; Springer: Berlin, Germany, 2012; pp. 49–53. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Dinis, T.C.; Madeira, V.M.; Almeida, L.M. Action of phenolic derivatives (acetaminophen, salicylate, and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Lin, H.-T.V.; Wu, G.-J.; Hsieh, M.-C.; Chang, S.-H.; Tsai, G.-J. Purification and characterization of Nattokinase from cultural filtrate of red alga Porphyra dentata fermented by Bacillus subtilis N1. J. Mar. Sci. Technol. 2015, 23, 240–248. [Google Scholar]
- Nair, D.; Krishna, J.G.; Panikkar, M.V.N.; Nair, B.G.; Pai, J.G.; Nair, S.S. Identification, purification, biochemical and mass spectrometric characterization of novel phycobiliproteins from a marine red alga, Centroceras clavulatum. Int. J. Biol. Macromol. 2018, 114, 679–691. [Google Scholar] [CrossRef]
- Malairaj, S.; Muthu, S.; Gopal, V.B.; Perumal, P.; Ramasamy, R. Qualitative and quantitative determination of R-phycoerythrin from Halymenia floresia (Clemente) C. Agardh by polyacrylamide gel using electrophoretic elution technique. J. Chromatogr. A 2016, 1454, 120–126. [Google Scholar] [CrossRef]
- Chethana, S.; Nayak, C.A.; Madhusudhan, M.; Raghavarao, K. Single step aqueous two-phase extraction for downstream processing of C-phycocyanin from Spirulina platensis. J. Food Sci. Technol. 2015, 52, 2415–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Content (%) |
|---|---|
| Moisture | 10.93 ± 0.41 |
| Ash | 8.66 ± 1.47 |
| Crude protein | 33.94 ± 1.59 |
| Crude fat | 1.10 ± 0.45 |
| Carbohydrate | 45.37 ± 0.21 |
| Sample | R-PE (mg/mL) | R-PC (mg/mL) | Fluorescence Intensity | Purity Indexes | ||
|---|---|---|---|---|---|---|
| R-PE | R-PE | A560/A280 | A615/A280 | |||
| without UF | 0.17 ± 0.1 b | 0.22 ± 0.1 b | 18,505 ± 1144 a | 2476 ± 106 a | 0.33 | 0.23 |
| UF < 100 kDa | 0.04 ± 0.1 c | 0.04 ± 0.1 c | 8653 ± 719 b | 782 ± 71 b | 0.05 | 0.01 |
| UF > 100 kDa | 1.15 ± 0.1 a | 0.69 ± 0.1 a | 18560 ± 381 a | 2492 ± 26 a | 0.99 | 0.70 |
| PE (mg/mL) | PC (mg/mL) | Fluorescence Intensity | Purity Indexes | |||
|---|---|---|---|---|---|---|
| PE | PC | A560/A280 | A615/A280 | |||
| R-PE | 0.36 ± 0.01 | 0.03 ± 0.01 | 45,438 ± 2164 | 1628 ± 356 | 3.18 | 0.32 |
| R-PC | 0.06 ± 0.01 | 0.86 ± 0.05 | 2684 ± 235 | 3916 ± 377 | 1.75 | 3.37 |
| Small molecule PE | 0.13 ± 0.02 | 0.03 ± 0.01 | 39,558 ± 573 | 950 ± 357 | 2.32 | 0.73 |
| Fractions (mg/mL) | DPPH Scavenging Effect (%) | Equal to μg/mL Trolox |
|---|---|---|
| R-PE | ||
| 5 | 30.17 ± 0.95 b | 7.14 ± 0.33 |
| 10 | 63.06 ± 1.08 a | 15.14 ± 0.37 |
| R-PC | ||
| 5 | 23.87 ± 1.12 c | 5.61 ± 0.39 |
| 10 | 59.46 ± 1.23 a | 14.26 ± 0.42 |
| Small molecule PE | ||
| 5 | 28.23 ± 0.88 b | 6.67 ± 0.30 |
| 10 | 63.96 ± 1.37 a | 15.36 ± 0.47 |
| Fractions (mg/mL) | OD700nm | Equal to μg/mL Trolox |
|---|---|---|
| R-PE | ||
| 5 | 0.046 ± 0.005 e | 5.7493 ± 0.5274 |
| 10 | 0.087 ± 0.007 c | 11.958 ± 0.9880 |
| R-PC | ||
| 5 | 0.138 ± 0.006 b | 19.5375 ± 0.6028 |
| 10 | 0.250 ± 0.028 a | 36.4900 ± 4.2910 |
| Small molecule PE | ||
| 5 | 0.029 ± 0.006 f | 3.1123 ± 0.6781 |
| 10 | 0.058 ± 0.005 d | 7.5580 ± 0.6910 |
| Fractions (μg/mL) | Fe2+ Chelating Effect (%) | Equal to μg/mL EDTA |
|---|---|---|
| R-PE | ||
| 31.25 | 60.59 ± 0.74 e | 26.4 ± 0.9 |
| 62.50 | 71.61 ± 0.52 c | 39.6 ± 0.6 |
| 125 | 87.63 ± 0.23 b | 58.9 ± 0.3 |
| 250 | 91.21 ± 0.11 a | 63.2 ± 0.1 |
| R-PC | ||
| 31.25 | 67.10 ± 0.45 d | 34.2 ± 0.5 |
| 62.5 | 73.61 ± 0.31 c | 42. ± 0.4 |
| 125 | 86.18 ± 0.30 b | 57.1 ± 0.4 |
| 250 | 90.31 ± 0.11 a | 62.1 ± 0.1 |
| Small molecule PE | ||
| 31.25 | 63.09 ± 1.21 e | 29.4 ± 1.5 |
| 62.5 | 73.96 ± 0.23 c | 42.5 ± 0.3 |
| 125 | 89.18 ± 0.15 b | 60.7 ± 0.2 |
| 250 | 92.11 ± 0.11 a | 64.2 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-H.; Chen, W.-C.; Gao, Y.-H.; Chen, G.-W.; Lin, H.-T.V.; Pan, C.-L. Enzyme-Assisted Method for Phycobiliproteins Extraction from Porphyra and Evaluation of Their Bioactivity. Processes 2021, 9, 560. https://doi.org/10.3390/pr9030560
Huang C-H, Chen W-C, Gao Y-H, Chen G-W, Lin H-TV, Pan C-L. Enzyme-Assisted Method for Phycobiliproteins Extraction from Porphyra and Evaluation of Their Bioactivity. Processes. 2021; 9(3):560. https://doi.org/10.3390/pr9030560
Chicago/Turabian StyleHuang, Chung-Hsiung, Wei-Chen Chen, Yu-Huei Gao, Guan-Wen Chen, Hong-Ting Victor Lin, and Chorng-Liang Pan. 2021. "Enzyme-Assisted Method for Phycobiliproteins Extraction from Porphyra and Evaluation of Their Bioactivity" Processes 9, no. 3: 560. https://doi.org/10.3390/pr9030560