
Influence of Hanseniasporauvarum AS27 on Chemical and Sensorial Characteristics of Aglianico Wine

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeasts and Growth Conditions
2.2. Winemaking Design
2.3. Microbiological Analysis
2.4. Chemical Analytical Methods
2.5. Sensory Profile of Wines
2.6. Statistical Analysis
3. Results and Discussion
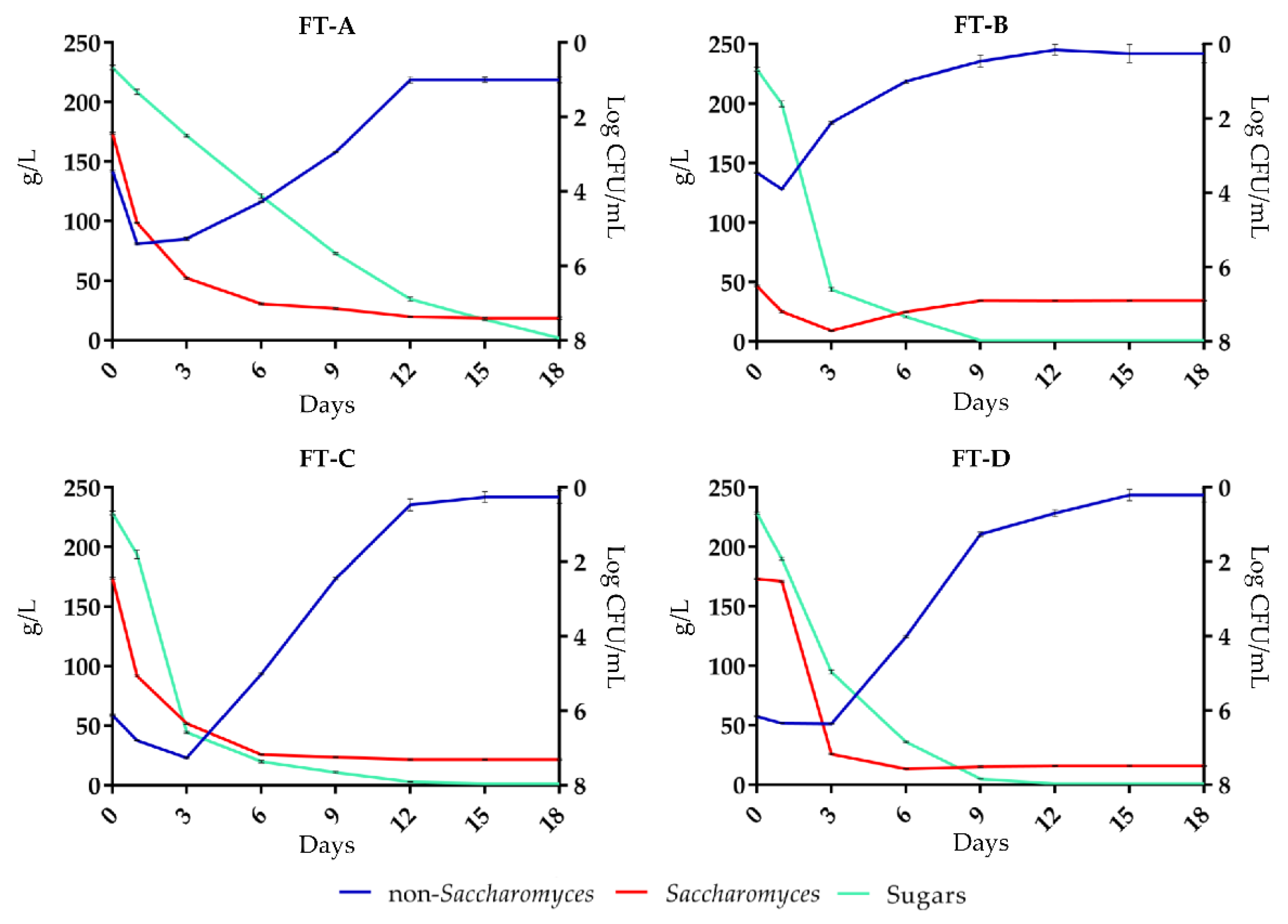
3.1. Fermentation Kinetics
3.2. Wine Chemistry

{kind=link}
{kind=link}
| Compounds | FT-A | FT-B | FT-C | FT-D | Threshold (µg/L) | Odor Descriptor | Ref. |
|---|---|---|---|---|---|---|---|
| Acetaldehyde ** | 11.5 ± 0.1 b | 9.61 ± 0.12 a | 9.52 ± 0.31 a | 9.27 ± 0.12 a | 500 | Green leaves, fruity | [52] |
| Ethyl acetate ** | 118 ± 1 d | 44.8 ± 1.1 a | 68.5 ± 1.8 c | 55.5 ± 1.9 b | 18,000 | Solvent, fruity, sweetish | [53] |
| 1-Heptanol | 186 ± 11 c | 111 ± 7 b | 82.4 ± 5.1 a | 109 ± 12 b | 200–300 | Lemon, orange, copper | [54] |
| 2-Methyl propanol | 24.4 ± 0.1 a | 27.8 ± 0.1 c | 26.6 ± 0.2 b | 24.5 ± 0.4 a | 40,000 | Wine, solvent, bitter | [55] |
| 1-Octanol | 237 ± 2 c | 207 ± 2 b | 198 ± 2 a | 212 ± 3 b | 820 | Coconut, walnut, oily | [52] |
| 1-Pentanol | 63.1 ± 1.1 c | 35.1 ± 0.9 a | 47.8 ± 1.2 b | 58.9 ± 2.6 c | 64,430 | Alcohol, medicinal | [52] |
| 1-Propanol | 18.9 ± 1.3 b | 31.8 ± 1.2 d | 13.7 ± 1.5 a | 22.8 ± 1.6 c | 830 | Pungent, harsh, ripe fruit | [55] |
| 2,3-Butanediol | 412 ± 12 a | 598 ± 14 c | 551 ± 12 b | 618 ± 17 c | 120,000 | Butter, creamy | [54] |
| 2-Methyl-1-butanol | 156 ± 12 a | 173. ± 12 a | 164 ± 14 a | 203 ± 13 b | 32,000 | Alcohol, banana | [52] |
| 2-Pentanol | 46.4 ± 1.1 a | 55.4 ± 1.1 b | 82.4 ± 1.6 c | 81.4 ± 1.6 c | - | Green | [56] |
| 2-Phenylethanol | 89.8 ± 11.3 a | 69.1 ± 4.1 a | 78.7 ± 14.2 a | 112 ± 12 b | 14,000 | Roses | [57] |
| 3-Methyl-1-butanol | 86.8 ± 1.2 a | 108 ± 11 a | 101 ± 12 a | 119 ± 12 b | 30,000 | Cheese | [57] |
| Hexanoic acid | 64.8 ± 1.6 a | 83.2 ± 1.6 b | 66.4 ± 1.9 a | 78.6 ± 2.7 b | 420 | Cheese, rancid | [54] |
| Methanol | 119 ± 1 | 112 ± 13 | 107 ± 4 | 105 ± 6 | 100,000 | Sweet | [58] |
| Methionol | 163 ± 2 b | 104 ± 2 a | 99.2 ± 3.6 a | 98.4 ± 6.1 a | 500 | Boiled potato, rubber | [59] |
| Linalool | 32.2 ± 0.3 b | 23.2 ± 1.4 a | 69.3 ± 0.7 c | 68.6 ± 0.8 c | 25.2 | Muscat, flowery, fruit | [54] |
| 4-Terpineol | 19.7 ± 0.1 a | 28.5 ± 0.3 b | 45.8 ± 0.6 d | 43.5 ± 0.5 c | 110–400 | Light aroma, wood, soil | [54] |
| α-Terpineol | 8.4 ± 0.7 a | 14.9 ± 0.8 b | 37.5 ± 0.4 d | 32.8 ± 0.8 c | 250 | Anise | [57] |
| Nerol | 34.5 ± 0.7 a | 42.6 ± 1.1 b | 77.8 ± 0.5 c | 98.6 ± 1.0 d | 400 | Rose-like aromas | [60] |
| Nonanol | 12.5 ± 0.9 a | 29.53 ± 1.4 b | 68.2 ± 1.5 c | 66.5 ± 0.9 c | 310 | Coconut, walnut, oily | [52] |
| Geraniol | 10.4 ± 0.2 b | 5.62 ± 0.13 a | 10.9 ± 0.2 b | 12.5±0.2 c | 130 | Floral | [60] |
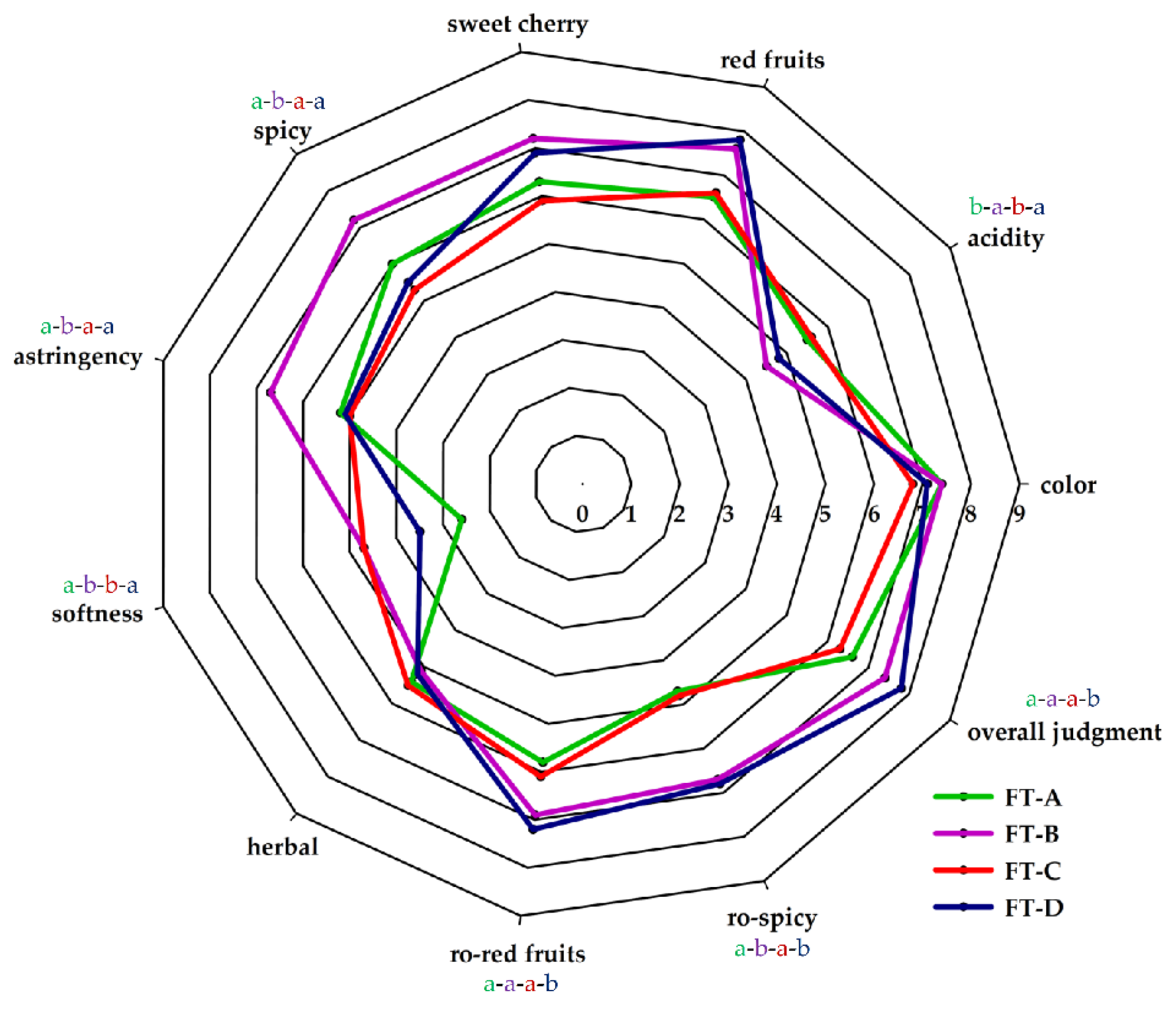
3.3. Sensory Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wolfe, B.E.; Dutton, R.J. Fermented Foods as Experimentally Tractable Microbial Ecosystems. Cell 2015, 161, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of Pure and Mixed Cultures of the Main Wine Yeast Species on Grape Must Fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Zulian, L.; Ferreres, À.; Pastor, R.; Esteve-Zarzoso, B.; Beltran, G.; Mas, A. Sequential Inoculation of Native Non-Saccharomyces and Saccharomyces Cerevisiae Strains for Wine Making. Front. Microbiol. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile Compounds Contribution of Hanseniaspora Guilliermondii and Hanseniaspora Uvarum during Red Wine Vinifications. Food Control 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled Mixed Culture Fermentation: A New Perspective on the Use of Non-Saccharomyces Yeasts in Winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected Non-Saccharomyces Wine Yeasts in Controlled Multistarter Fermentations with Saccharomyces Cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of Non-Saccharomyces Yeasts with Enzymatic Activities of Interest in Wine-Making. Int. J. Food Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Mendes Ferreira, A.; Clímaco, M.C.; Mendes Faia, A. The Role of Non-Saccharomyces Species in Releasing Glycosidic Bound Fraction of Grape Aroma Components—A Preliminary Study. J. Appl. Microbiol. 2001, 91, 67–71. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Mateo, J.J.; Maicas, S. Screening of Hanseniaspora Strains for the Production of Enzymes with Potential Interest for Winemaking. Fermentation 2016, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Haight, K.G.; Gump, B.H. The Use of Macerating Enzymes in Grape Juice Processing. Am. J. Enol. Vitic. 1994, 45, 113. [Google Scholar]
- Winterhalter, P.; Skouroumounis, G.K. Glycoconjugated aroma compounds: Occurrence, role and biotechnological transformation. In Biotechnology of Aroma Compounds; Berger, R.G., Babel, W., Blanch, H.W., Cooney, C.L., Enfors, S.-O., Eriksson, K.-E.L., Fiechter, A., Klibanov, A.M., Mattiasson, B., Primrose, S.B., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 73–105. ISBN 978-3-540-68602-6. [Google Scholar]
- Barbagallo, R.N.; Spagna, G.; Palmeri, R.; Restuccia, C.; Giudici, P. Selection, Characterization and Comparison of β-Glucosidase from Mould and Yeasts Employable for Enological Applications. Enzym. Microb. Technol. 2004, 35, 58–66. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Effects of Non-Saccharomyces Yeasts on Color, Anthocyanin, and Anthocyanin-Derived Pigments of Tannat Grapes during Fermentation. Am. J. Enol. Vitic. 2018, 69, 148. [Google Scholar] [CrossRef]
- Mendoza, L.; Farias, M.E. Improvement of Wine Organoleptic Characteristics by Non-Saccharomyces Yeasts. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Formatex Research Center: Badajoz, Spain, 2010; Volume 2, pp. 908–919. [Google Scholar]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of Non-Saccharomyces Yeasts and Oenological Tannin in Red Winemaking: Influence on Colour, Aroma and Sensorial Properties of Young Wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Mendes, F.; Guedes de Pinho, P.; Hogg, T.; Vasconcelos, I. Heavy Sulphur Compounds, Higher Alcohols and Esters Production Profile of Hanseniaspora Uvarum and Hanseniaspora Guilliermondii Grown as Pure and Mixed Cultures in Grape Must. Int. J. Food Microbiol. 2008, 124, 231–238. [Google Scholar] [CrossRef]
- Lombardi, S.J.; Pannella, G.; Iorizzo, M.; Moreno-Arribas, M.V.; Tremonte, P.; Succi, M.; Sorrentino, E.; Macciola, V.; Di Renzo, M.; Coppola, R. Sequential Inoculum of Hanseniaspora Guilliermondii and Saccharomyces Cerevisiae for Winemaking Campanino on an Industrial Scale. World J. Microbiol. Biotechnol. 2018, 34, 161. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dou, G.; Guo, H.; Zhang, Q.; Qin, X.; Yu, W.; Jiang, C.; Xiao, H. Volatile Organic Compounds of Hanseniaspora Uvarum Increase Strawberry Fruit Flavor and Defense during Cold Storage. Food Sci. Nutr. 2019, 7, 2625–2635. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Mateo, J.; Maicas, S. Characterisation of Hanseniaspora Isolates with Potential Aroma-Enhancing Properties in Muscat Wines. S. Afr. J. Enol. Vitic. 2014, 35, 292–303. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Berbegal, C.; Tufariello, M.; Grieco, F.; Spano, G. Impact of Co-Inoculation of Saccharomyces Cerevisiae, Hanseniaspora Uvarum and Oenococcus Oeni Autochthonous Strains in Controlled Multi Starter Grape Must Fermentations. LWT 2019, 109, 241–249. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased Flavour Diversity of Chardonnay Wines by Spontaneous Fermentation and Co-Fermentation with Hanseniaspora Vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jin, G.-J.; Xu, Y.-H.; Tao, Y.-S. Wine Aroma Response to Different Participation of Selected Hanseniaspora Uvarum in Mixed Fermentation with Saccharomyces Cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Imazio, S.; Biagini, B.; Failla, O.; Scienza, A. Pedigree Reconstruction of the Italian Grapevine Aglianico (Vitisvinifera L.) from Campania. Mol. Biotechnol. 2013, 54, 634–642. [Google Scholar] [CrossRef]
- Gambuti, A.; Strollo, D.; Erbaggio, A.; Lecce, L.; Moio, L. Effect of Winemaking Practices on Color Indexes and Selected Bioactive Phenolics of Aglianico Wine. J. Food Sci. 2007, 72, S623–S628. [Google Scholar] [CrossRef]
- Bonfante, A.; Alfieri, S.M.; Albrizio, R.; Basile, A.; De Mascellis, R.; Gambuti, A.; Giorio, P.; Langella, G.; Manna, P.; Monaco, E.; et al. Evaluation of the Effects of Future Climate Change on Grape Quality through a Physically Based Model Application: A Case Study for the Aglianico Grapevine in Campania Region, Italy. Agric. Syst. 2017, 152, 100–109. [Google Scholar] [CrossRef]
- Picariello, L.; Rinaldi, A.; Forino, M.; Errichiello, F.; Moio, L.; Gambuti, A. Effect of Different Enological Tannins on Oxygen Consumption, Phenolic Compounds, Color and Astringency Evolution of Aglianico Wine. Molecules 2020, 25, 4607. [Google Scholar] [CrossRef]
- Testa, B.; Lombardi, S.J.; Iorizzo, M.; Letizia, F.; Di Martino, C.; Di Renzo, M.; Strollo, D.; Tremonte, P.; Pannella, G.; Ianiro, M.; et al. Use of Strain Hanseniaspora Guilliermondii BF1 for Winemaking Process of White Grapes Vitis Vinifera Cv Fiano. Eur. Food Res. Technol. 2020, 246, 549–561. [Google Scholar] [CrossRef]
- Regulation, H.A.T. Commission Regulation (EEC) No. 2676/90 Determining Community Methods for the Analysis of Wines. Off. J. L 1990, 272, 1–192. [Google Scholar]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora Fermentations. Am. J. Enol. Vitic. 2001, 52, 198. [Google Scholar]
- Sugiyama, J. Yeasts: Characteristics and Identification, Third Edition, by J.A. Barnett, R.W. Payne, and D. Yarrow. Mycopathologia 2001, 149, 159–160. [Google Scholar] [CrossRef]
- Urso, R.; Rantsiou, K.; Dolci, P.; Rolle, L.; Comi, G.; Cocolin, L. Yeast Biodiversity and Dynamics during Sweet Wine Production as Determined by Molecular Methods. FEMS Yeast Res. 2008, 8, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Testa, B.; Lombardi, S.J.; Tremonte, P.; Succi, M.; Tipaldi, L.; Pannella, G.; Sorrentino, E.; Iorizzo, M.; Coppola, R. Biodiversity of Lactobacillus Plantarum from Traditional Italian Wines. World J. Microbiol. Biotechnol. 2014, 30, 2299–2305. [Google Scholar] [CrossRef] [Green Version]
- OIV. Compendium of International Methods of Analysis of Spirituous Beverages of Vitivinicultural Origin; OIV: Paris, France, 2014. [Google Scholar]
- Iorizzo, M.; Macciola, V.; Testa, B.; Lombardi, S.J.; De Leonardis, A. Physicochemical and Sensory Characteristics of Red Wines from the Rediscovered Autochthonous Tintilia Grapevine Grown in the Molise Region (Italy). Eur. Food Res. Technol. 2014, 238, 1037–1048. [Google Scholar] [CrossRef]
- Arcari, S.G.; Caliari, V.; Sganzerla, M.; Godoy, H.T. Volatile Composition of Merlot Red Wine and Its Contribution to the Aroma: Optimization and Validation of Analytical Method. Talanta 2017, 174, 752–766. [Google Scholar] [CrossRef]
- Lombardi, S.J.; Pannella, G.; Iorizzo, M.; Testa, B.; Succi, M.; Tremonte, P.; Sorrentino, E.; Di Renzo, M.; Strollo, D.; Coppola, R. Inoculum Strategies and Performances of Malolactic Starter Lactobacillus Plantarum M10: Impact on Chemical and Sensorial Characteristics of Fiano Wine. Microorganisms 2020, 8, 516. [Google Scholar] [CrossRef] [Green Version]
- Presidential decree DPR 563 8 Luglio 1981-Riconoscimento Della Personalità Giuridica dell’O.N.A.V; G.U., 1981; Volume 280.
- Ministerial decree DM 25 Luglio 2003-Disciplina Degli Esami Chimico-Fisici ed Organolettici e Dell’attività Delle Commissioni di Degustazione dei vini D.O.C.G. e D.O.C.; G.U., 2003; Volume 210.
- Lisanti, M.T.; Gambuti, A.; Genovese, A.; Piombino, P.; Moio, L. Partial Dealcoholization of Red Wines by Membrane Contactor Technique: Effect on Sensory Characteristics and Volatile Composition. Food Bioprocess Technol. 2013, 6, 2289–2305. [Google Scholar] [CrossRef]
- Ciani, M.; Maccarelli, F. Oenological Properties of Non-Saccharomyces Yeasts Associated with Wine-Making. World J. Microbiol. Biotechnol. 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Carlton, W.K.; Gump, B.; Fugelsang, K.; Hasson, A.S. Monitoring Acetaldehyde Concentrations during Micro-Oxygenation of Red Wine by Headspace Solid-Phase Microextraction with On-Fiber Derivatization. J. Agric. Food Chem. 2007, 55, 5620–5625. [Google Scholar] [CrossRef]
- Picariello, L.; Gambuti, A.; Picariello, B.; Moio, L. Evolution of Pigments, Tannins and Acetaldehyde during Forced Oxidation of Red Wine: Effect of Tannins Addition. LWT 2017, 77, 370–375. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of Ethyl Acetate and Isoamyl Acetate by Various Species of Wine Yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I. Yeast and Its Importance to Wine Aroma-a Review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Pretorius, I.S. Yeast Modulation of Wine Flavor. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2005; pp. 131–175. Volume 57, ISBN 0065-2164. [Google Scholar]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative Determination of the Odorants of Young Red Wines from Different Grape Varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Genovese, A.; Dimaggio, R.; Lisanti, M.T.; Piombino, P.; Moio, L. Aroma Composition of Red Wines by Different Extraction Methods and Gas Chromatography-SIM/Mass Spectrometry Analysis. Ann. Chim. 2005, 95, 383–394. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine Flavor and Aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145. [Google Scholar] [CrossRef]
- Miller, G.H. Back Matter. In Whisky Science; Springer: Berlin/Heidelberg, Germany, 2019; pp. 421–467. [Google Scholar]
- Selfridge, T.B.; Amerine, M.A. Odor Thresholds and Interactions of Ethyl Acetate and Diacetyl in an Artificial Wine Medium. Am. J. Enol. Vitic. 1978, 29, 1. [Google Scholar]
- Tao, Y.-S.; Li, H. Active Volatiles of Cabernet Sauvignon Wine from Changli County. Health 2009, 1, 176. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Diéguez, S.; Rodriguez-Solana, R.; Domínguez, J.M.; Díaz, E. Impact Odorants and Sensory Profile of Young Red Wines from Four Galician (NW of Spain) Traditional Cultivars. J. Inst. Brew. 2015, 121, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Kong, J.; Xiao, Z.; Chen, F.; Ma, N.; Zhu, J. Characterization of Odor-Active Compounds of Various Chinese “Wuliangye” Liquors by Gas Chromatography–Olfactometry, Gas Chromatography–Mass Spectrometry and Sensory Evaluation. Int. J. Food Prop. 2017, 20, S735–S745. [Google Scholar] [CrossRef] [Green Version]
- Culleré, L.; Escudero, A.; Cacho, J.; Ferreira, V. Gas Chromatography−Olfactometry and Chemical Quantitative Study of the Aroma of Six Premium Quality Spanish Aged Red Wines. J. Agric. Food Chem. 2004, 52, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Leonardos, G.; Kendall, D.; Barnard, N. Odor Threshold Determinations of 53 Odorant Chemicals. J. Air Pollut. Control Assoc. 1969, 19, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Zhang, Z.-W. A Preliminary Study of Aroma Composition and Impact Odorants of Cabernet Franc Wines under Different Terrain Conditions of the Loess Plateau Region (China). Molecules 2018, 23, 1096. [Google Scholar] [CrossRef] [Green Version]
- Čuš, F.; Jenko, M. The Influence of Yeast Strains on the Composition and Sensory Quality of Gewürztraminer Wine. Food Technol. Biotechnol. 2013, 51, 547–553. [Google Scholar]
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Fariña, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and Transcriptomic Basis of Hanseniaspora Vineae’s Impact on Flavor Diversity and Wine Quality. Appl. Environ. Microbiol. 2019, 85, e01959-18. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Beco, L.; Comitini, F. Fermentation Behaviour and Metabolic Interactions of Multistarter Wine Yeast Fermentations. Int. J. Food Microbiol. 2006, 108, 239–245. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast Interactions and Wine Flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- De Mets, G.; Goos, P.; Hertog, M.; Peeters, C.; Lammertyn, J.; Nicolaï, B.M. Sensory Quality of Wine: Quality Assessment by Merging Ranks of an Expert-Consumer Panel. Aust. J. Grape Wine Res. 2017, 23, 318–328. [Google Scholar] [CrossRef]
- Genovese, A.; Lisanti, M.T.; Gambuti, A.; Piombino, P.; Moio, L. Relationship between Sensory Perception and Aroma Compounds of Monovarietal Red Wines. Acta Hortic. 2007, 754, 549–556. [Google Scholar] [CrossRef]
- García-Estévez, I.; Ramos-Pineda, A.M.; Escribano-Bailón, M.T. Interactions between Wine Phenolic Compounds and Human Saliva in Astringency Perception. Food Funct. 2018, 9, 1294–1309. [Google Scholar] [CrossRef]
- Diako, C.; McMahon, K.; Mattinson, S.; Evans, M.; Ross, C. Alcohol, Tannins, and Mannoprotein and Their Interactions Influence the Sensory Properties of Selected Commercial Merlot Wines: A Preliminary Study. J. Food Sci. 2016, 81, S2039–S2048. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.; Mateus, N.; Plet, B.; Pianet, I.; Dufourc, E.; De Freitas, V. Influence of Wine Pectic Polysaccharides on the Interactions between Condensed Tannins and Salivary Proteins. J. Agric. Food Chem. 2006, 54, 8936–8944. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.; Gambuti, A.; Moio, L. Precipitation of Salivary Proteins After the Interaction with Wine: The Effect of Ethanol, PH, Fructose, and Mannoproteins. J. Food Sci. 2012, 77, C485–C490. [Google Scholar] [CrossRef]
- Gawel, R.; Godden, P.; Williamson, P.; Francis, L.; Smith, P.; Waters, L.; Herderich, M.; Johnson, D. Influence of Phenolics on White Wine Quality and Style. Wine Vitic. J. 2014, V29N3. [Google Scholar]
- Vidal, S.; Francis, L.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E. The Mouth-Feel Properties of Polysaccharides and Anthocyanins in a Wine like Medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Troszyńska, A.; Narolewska, O.; Robredo, S.; Estrella, I.; Hernández, T.; Lamparski, G.; Amarowicz, R. The Effect of Polysaccharides on the Astringency Induced by Phenolic Compounds. Food Qual. Prefer. 2010, 21, 463–469. [Google Scholar] [CrossRef]
- Brandão, E.; Silva, M.S.; García-Estévez, I.; Williams, P.; Mateus, N.; Doco, T.; de Freitas, V.; Soares, S. The Role of Wine Polysaccharides on Salivary Protein-Tannin Interaction: A Molecular Approach. Carbohydr. Polym. 2017, 177, 77–85. [Google Scholar] [CrossRef]
- Lombardi, S.J.; De Leonardis, A.; Lustrato, G.; Testa, B.; Iorizzo, M. Yeast Autolysis in Sparkling Wine Aging: Use of Killer and Sensitive Saccharomyces Cerevisiae Strains in Co-Culture. Recent Pat. Biotechnol. 2015, 9, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of Non-Saccharomyces Wine Yeasts as Novel Sources of Mannoproteins in Wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef]
- Giovani, G.; Rosi, I.; Bertuccioli, M. Quantification and Characterization of Cell Wall Polysaccharides Released by Non-Saccharomyces Yeast Strains during Alcoholic Fermentation. Int. J. Food Microbiol. 2012, 160, 113–118. [Google Scholar] [CrossRef]
- Gawel, R.; Smith, P.A.; Waters, E.J. Influence of Polysaccharides on the Taste and Mouthfeel of White Wine. Aust. J. Grape Wine Res. 2016, 22, 350–357. [Google Scholar] [CrossRef]
- Lubbers, S.; Verret, C.; Voilley, A. The Effect of Glycerol on the Perceived Aroma of a Model Wine and a White Wine. LWT-Food Sci. Technol. 2001, 34, 262–265. [Google Scholar] [CrossRef]
- Gambuti, A.; Genovese, A.; Lamorte, S.; Capuano, R.; Lisanti, M.T.; Piombino, P.; Moio, L. Study of the Influence of Grape Ripeness Degree on Aroma Characteristics of Aglianico Wines by Instrumental and Sensory Analysis. Acta Hortic. 2007, 754, 533–540. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Parameters | FT-A | FT-B | FT-C | FT-D |
|---|---|---|---|---|
| pH | 3.18 ± 0.07 | 3.23 ± 0.15 | 3.21 ± 0.10 | 3.21 ± 0.10 |
| Titratable acidity * (g/L) | 9.32 ± 0.16 | 9.10 ± 0.30 | 9.35 ± 0.15 | 9.26 ± 0.15 |
| Reducing sugars (g/L) | 1.90 ± 0.10 | 1.20 ± 0.20 | 1.47 ± 0.60 | 1.18 ± 0.60 |
| Alcohol (% v/v) | 13.0 ± 0.1 | 12.9 ± 0.1 | 12.9 ± 0.2 | 13.0 ± 0.1 |
| Glycerol (g/L) | 7.00 ± 0.18 a | 8.76 ± 0.35 b | 7.23 ± 0.25 a | 8.36 ± 0.25 b |
| Volatile acidity ** (g/L) | 0.97 ± 0.06 d | 0.29 ± 0.02 a | 0.62 ± 0.03 c | 0.46 ± 0.02 b |
| Malic acid (g/L) | 3.21 ± 0.20 | 3.13 ± 0.02 | 3.25 ± 0.10 | 3.11 ± 0.12 |
| Lactic acid (g/L) | 0.02 ± 0.00 a | 0.04 ± 0.01 a | 0.02 ± 0.00 a | 0.03 ± 0.00 b |
| Catechins (mg/L) | 384 ± 4 a | 512.03 ± 7 c | 460 ± 6 b | 543 ± 5 d |
| Total polyphenols *** (mg/L) | 2267 ± 2 a | 2451 ± 3 c | 2371 ± 6 b | 2550 ± 7 d |
| Anthocyanins **** (mg/L) | 463 ± 1 a | 522 ± 1 b | 450 ± 9 a | 552 ± 9 c |
| Color intensity (IC) | 13.7 ± 0.7 a | 15.6 ± 0.3 b | 13.8 ± 0.4 a | 16.6 ± 0.3 b |
| Color tonality (CT) | 0.49 ± 0.04 | 0.47 ± 0.04 | 0.42 ± 0.04 | 0.46 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Testa, B.; Coppola, F.; Lombardi, S.J.; Iorizzo, M.; Letizia, F.; Di Renzo, M.; Succi, M.; Tremonte, P. Influence of Hanseniasporauvarum AS27 on Chemical and Sensorial Characteristics of Aglianico Wine. Processes 2021, 9, 326. https://doi.org/10.3390/pr9020326
Testa B, Coppola F, Lombardi SJ, Iorizzo M, Letizia F, Di Renzo M, Succi M, Tremonte P. Influence of Hanseniasporauvarum AS27 on Chemical and Sensorial Characteristics of Aglianico Wine. Processes. 2021; 9(2):326. https://doi.org/10.3390/pr9020326
Chicago/Turabian StyleTesta, Bruno, Francesca Coppola, Silvia Jane Lombardi, Massimo Iorizzo, Francesco Letizia, Massimo Di Renzo, Mariantonietta Succi, and Patrizio Tremonte. 2021. "Influence of Hanseniasporauvarum AS27 on Chemical and Sensorial Characteristics of Aglianico Wine" Processes 9, no. 2: 326. https://doi.org/10.3390/pr9020326