

The Crucial Impact of Microbial Growth and Bioenergy Conversion on Treating Livestock Manure and Antibiotics Using Chlorella sorokiniana


Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculum and Culture Conditions
2.2. Livestock Excreta Characteristics and Antibiotics
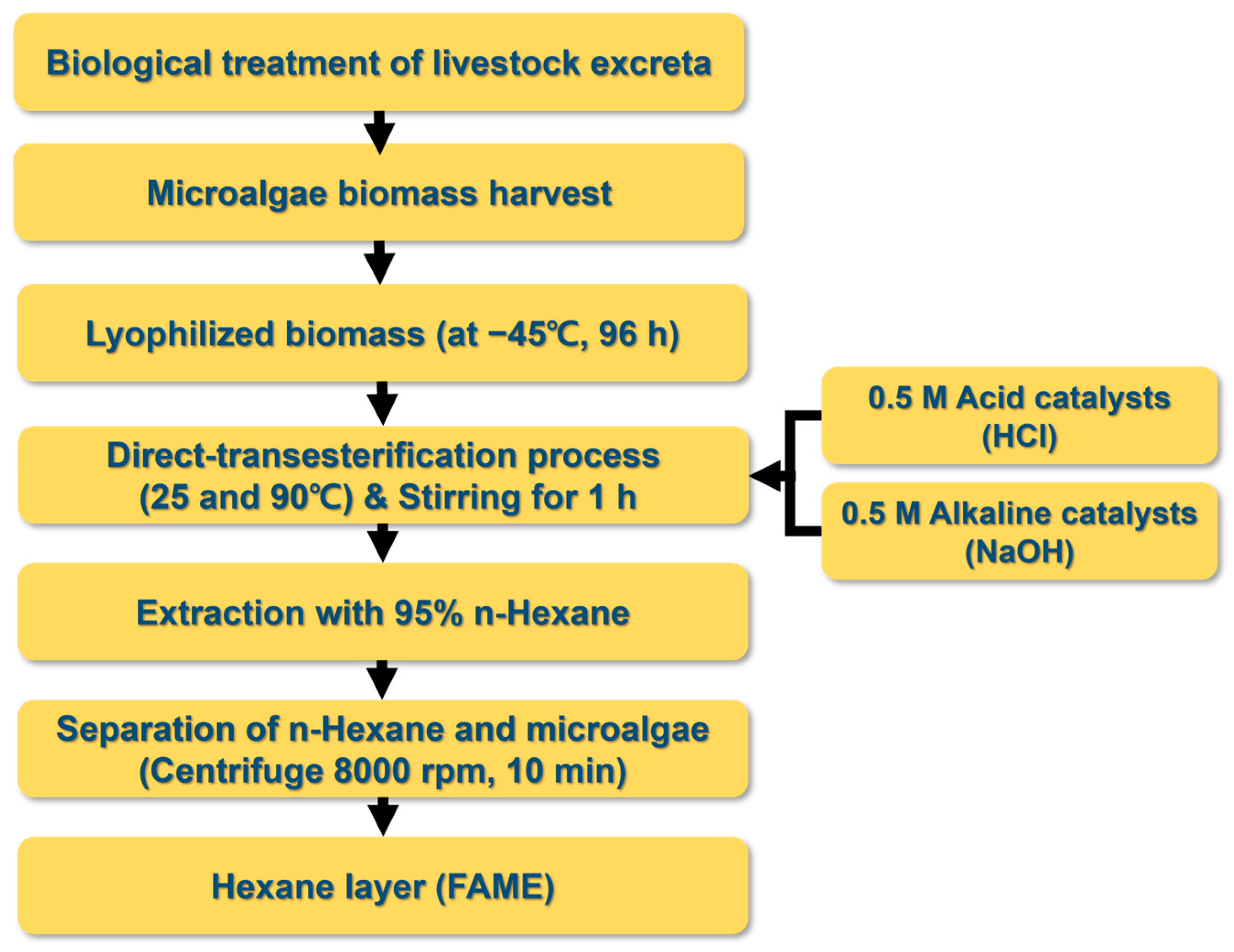
2.3. Biodiesel Production from the Microalgae Biomass
2.4. Analytical Methods for Water Quality and Fatty Acid Methyl Ester (FAME)
3. Results and Discussion
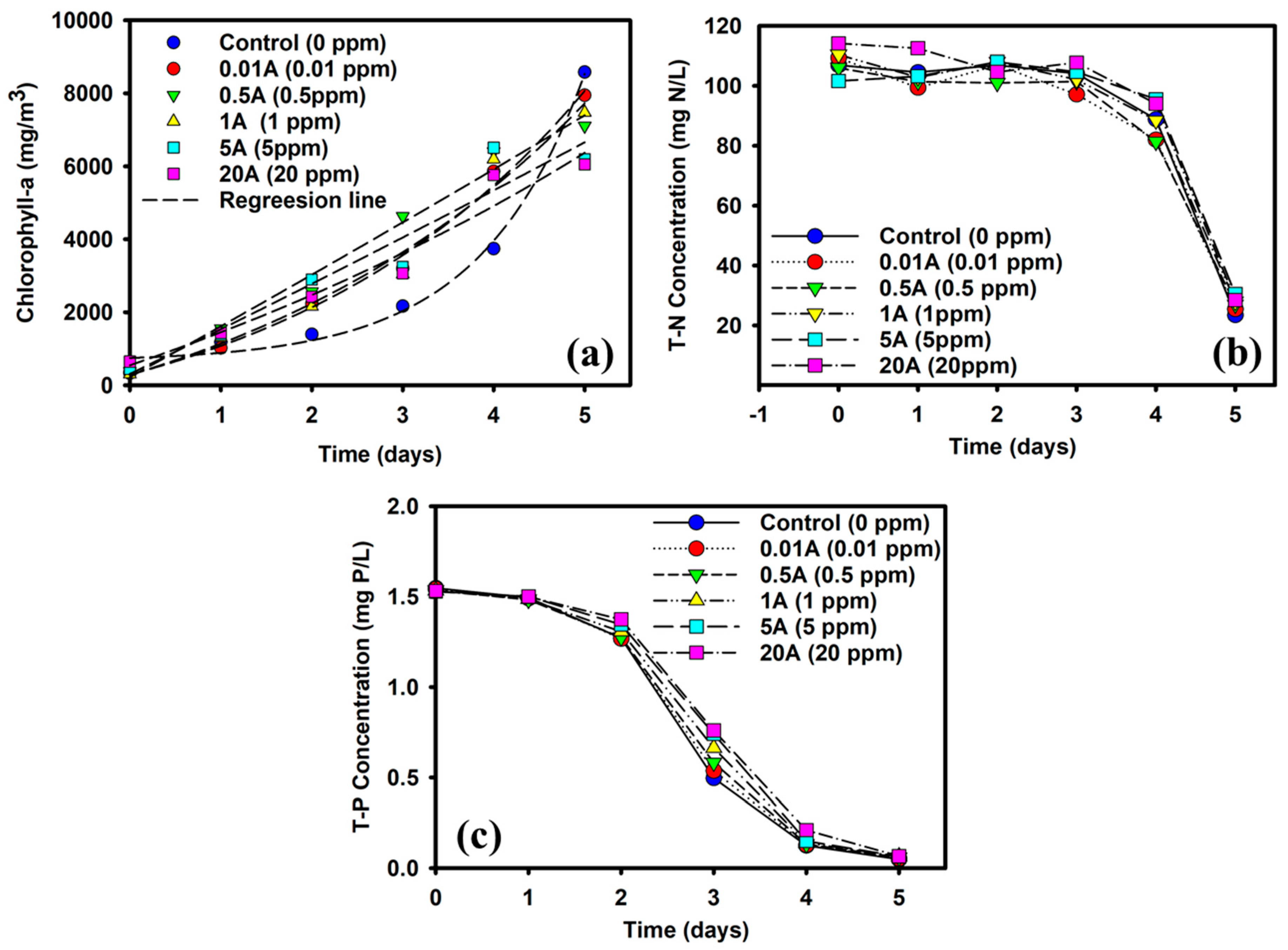
3.1. Negative Effect on the Microalgae Growth of AMX
3.2. Nutrient and AMX Removal in Microalgae Treatment
3.2.1. Effect of AMX on T-N Removal

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgae | Antibiotic Type | Antibiotic Strength (mg/L) | Initial T-N (mg N/L) | Effluent T-N (mg N/L) | Removal Efficiency (%) | Operation Period | Reference |
|---|---|---|---|---|---|---|---|
| Chlorella sorokiniana | AMX | 0 | 106.9 | 23.5 | 78.0 | 5 | This study |
| 0.01 | 109.4 | 25.5 | 77.0 | ||||
| 0.5 | 106.0 | 27.0 | 76.0 | ||||
| 1 | 110.4 | 28.0 | 75.2 | ||||
| 5 | 101.7 | 30.5 | 74.6 | ||||
| 20 | 114.2 | 28.5 | 74.5 | ||||
| Chlorella vulgaris and Scenedesmus dimorphus | - | - | - | - | 70.4 | 10 | [51] |
| Chlorella vulgaris and Scenedesmus dimorphus | - | - | - | - | 64.5 | 10 | [52] |
| Chlorella sorokiniana | - | - | 214.9 | 150.3 | 29.4 | 7 | [50] |
| Chlorella vulgaris | - | - | 113.3 | 51.8 | 54.3 | 7 | [53] |
| Chlorella vulgaris | SMX | 0.5 | 55.4 | 0.84 | 98.5 | 7 | [49] |
| Chlorella regularis | AMX | 3 | 45 | 7.92 | 82.4 | 18 | [43] |
3.2.2. Effect of AMX on T-P Removal
| Microalgae | Antibiotic Type | Antibiotic Strength (mg/L) | Initial T-P (mg P/L) | Effluent T-P (mg P/L) | Highest Removal Rate (mg P/L/day) | Removal Efficiency (%) | Operation Periods | Reference |
|---|---|---|---|---|---|---|---|---|
| Chlorella sorokiniana | AMX | 0 | 1.55 | 0.04 | 0.77 | 96.8 | 5 | This study |
| 0.01 | 1.54 | 0.05 | 0.73 | 96.5 | ||||
| 0.5 | 1.54 | 0.06 | 0.69 | 96.1 | ||||
| 1 | 1.53 | 0.06 | 0.64 | 95.8 | ||||
| 5 | 1.53 | 0.06 | 0.60 | 95.8 | ||||
| 20 | 1.53 | 0.07 | 0.61 | 95.6 | ||||
| Chlorella sp. | - | - | 57.3 | 18.1 | 0.89 | 68.4 | 45 | [56] |
| Asterarcys quadricellulare | - | - | 0.40 | 0.20 | - | 50.0 | 8 | [55] |
| Neochloris aquatica | - | - | 0.40 | 0.25 | - | 37.0 | ||
| Chlorella vulgaris and Scenedesmus dimorphus | - | - | - | - | - | 79.7 | 10 | [51] |
| Chlorella vulgaris and Ganoderma lucidum | - | - | - | - | - | 70.3 | 10 | [57] |
| Chlorella vulgaris | SMX | 0.5 | 27.2 | 0.41 | 3.82 | 98.5 | 7 | [49] |
| Chlorella regularis | AMX | 3 | 9.1 | 0.12 | - | 98.7 | 18 | [43] |
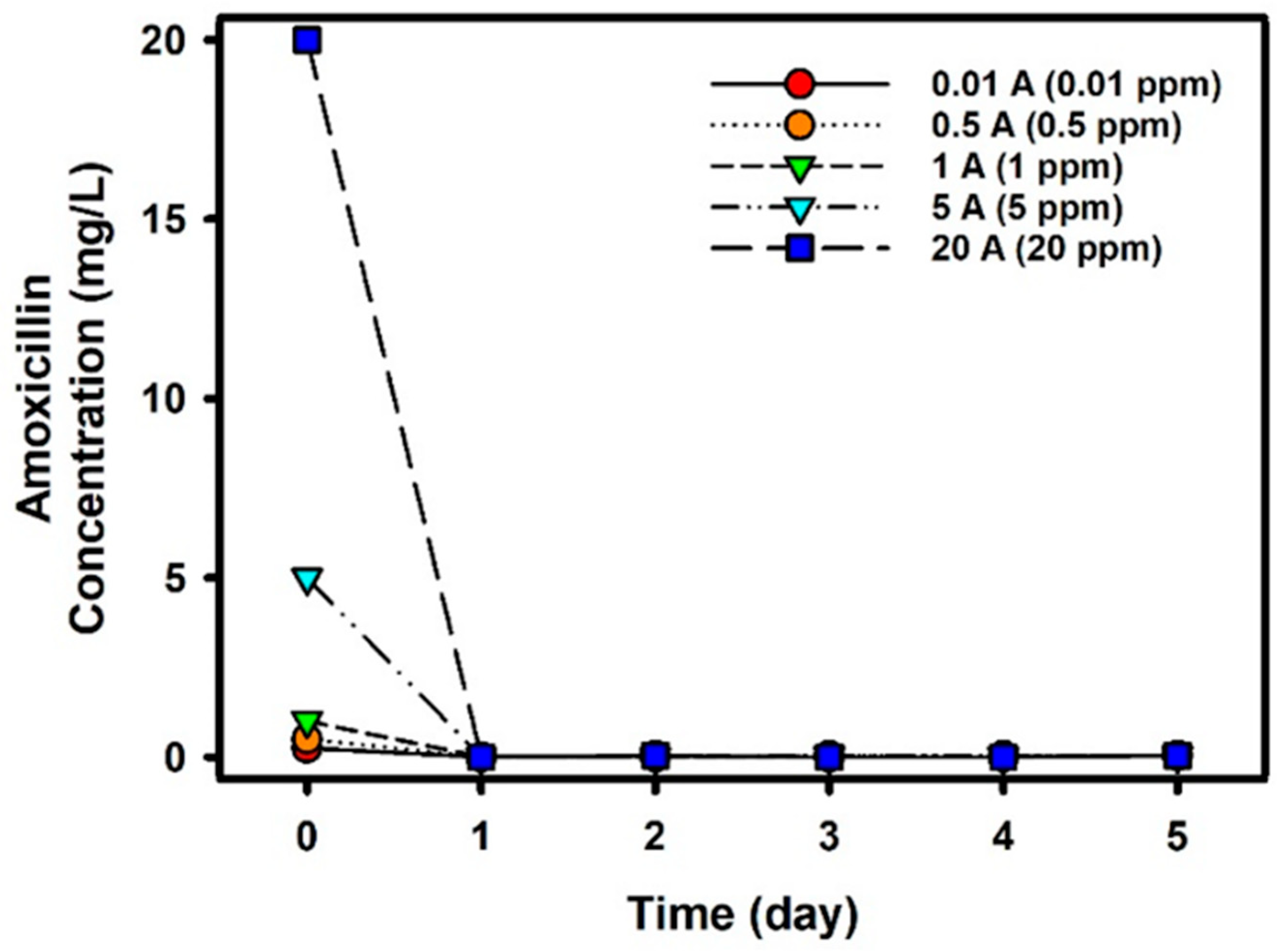
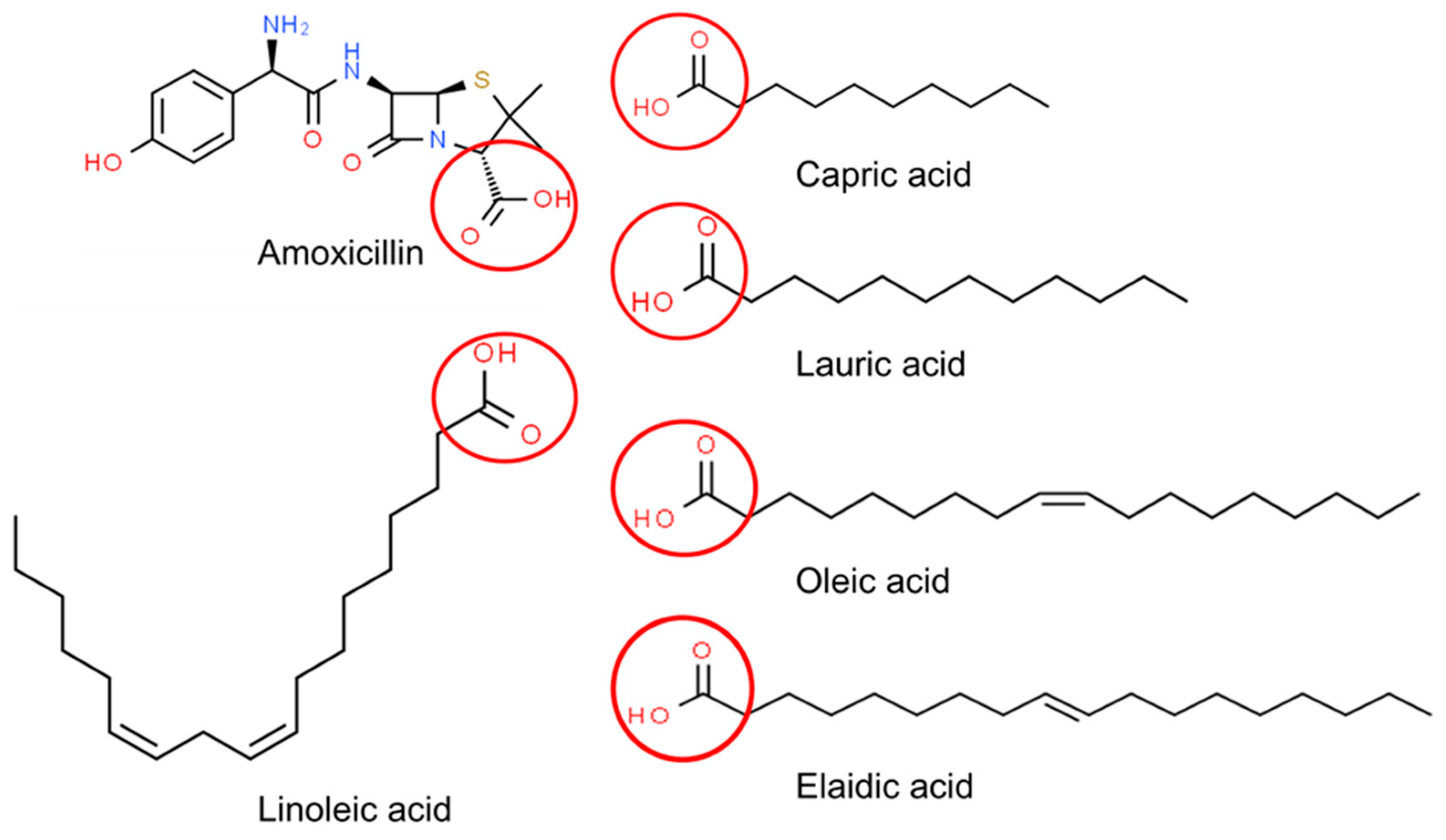
3.2.3. Reduction of Amoxicillin
3.3. Inhibitory Effect of AMX on Biodiesel Production
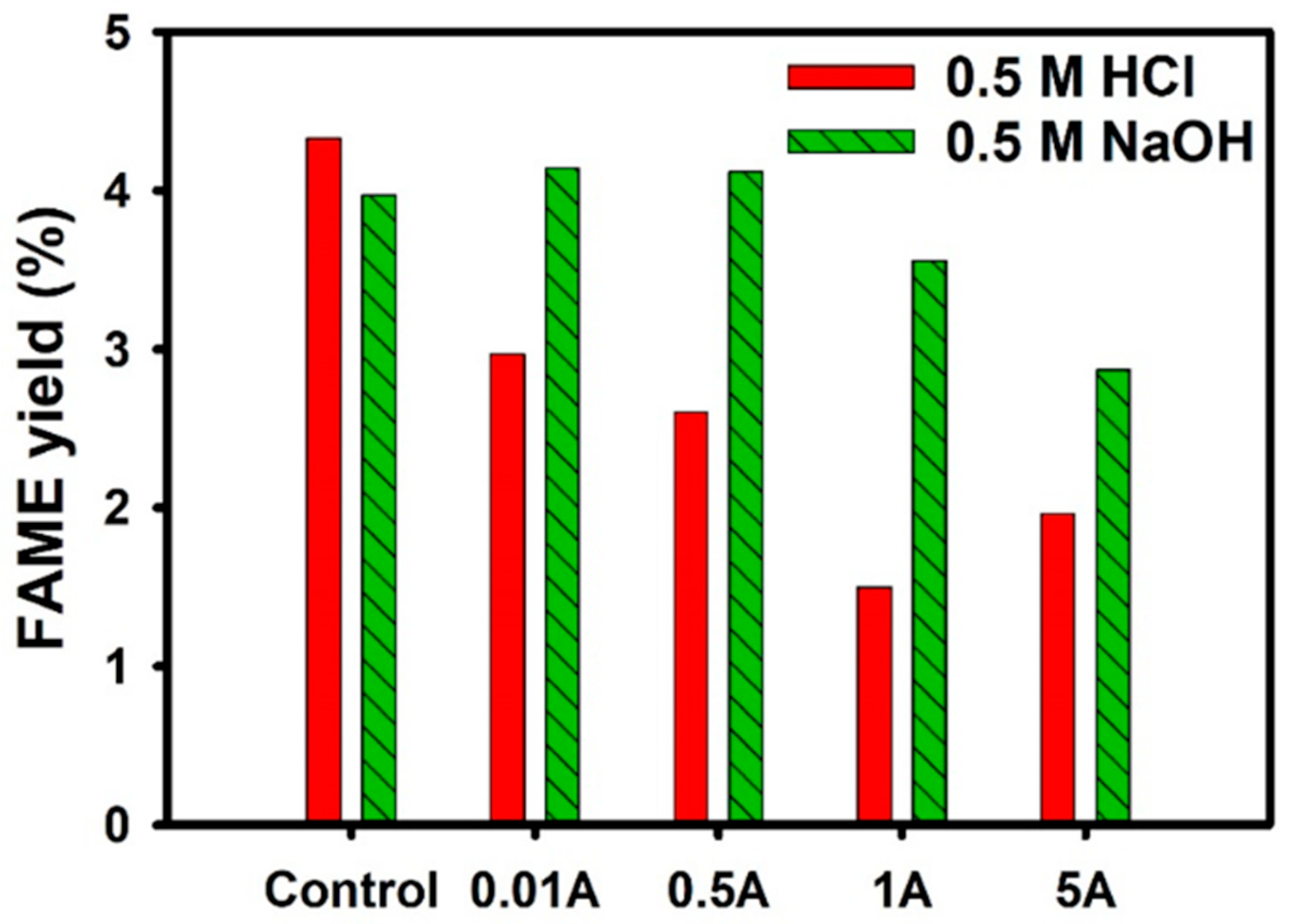
3.3.1. Negative Effect of AMX Inhibition in Transesterification
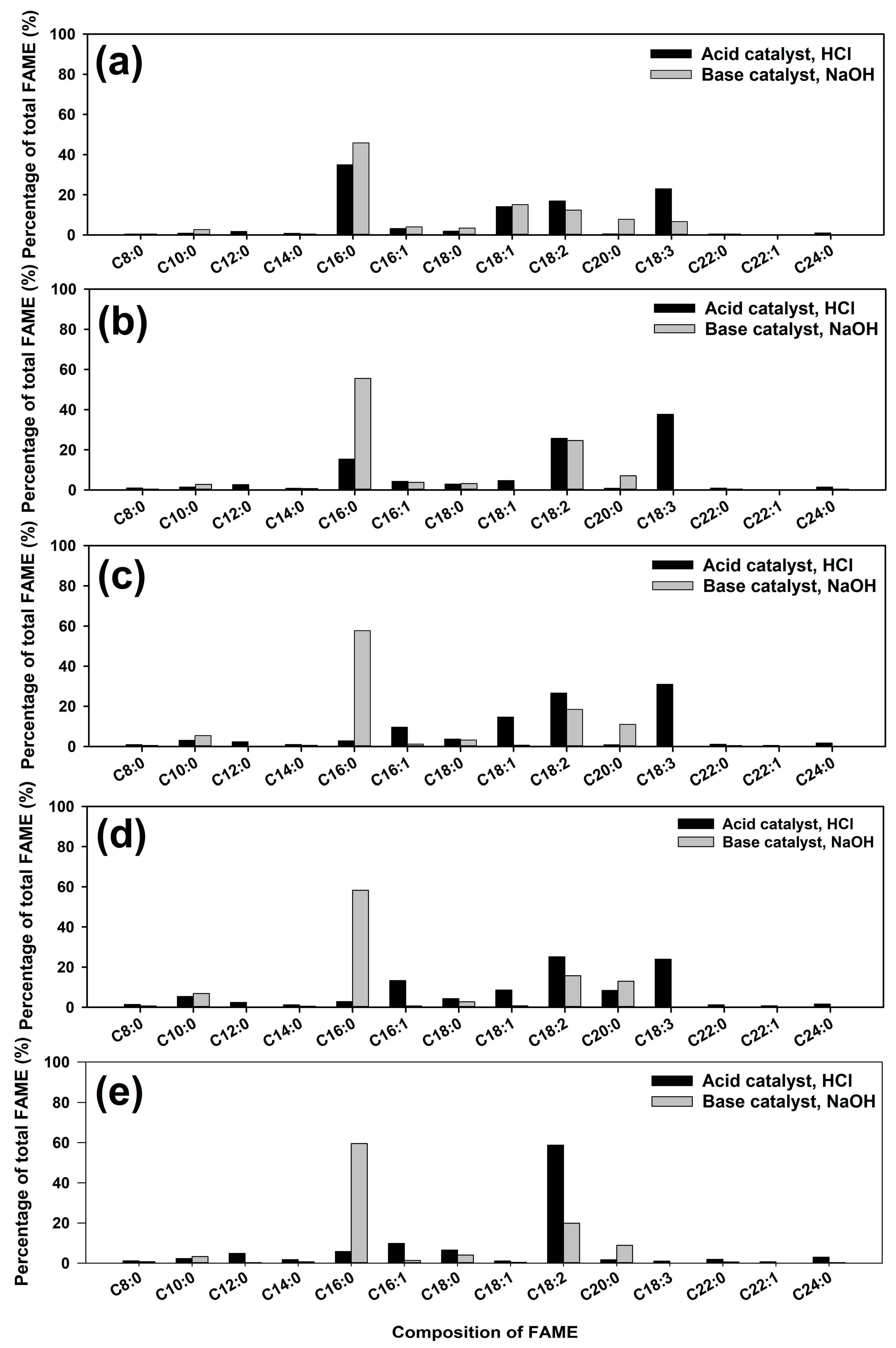
3.3.2. Changes in FAME Composition of Biodiesel Due to AMX
3.4. Limitations and Implications
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| LE | Livestock excreta |
| FAME | Fatty acid methyl ester |
| ppb | Parts per billion |
| AMX | Amoxicillin |
| COD | Chemical oxygen demand |
| T-N | Total nitrogen |
| T-P | Total phosphorus |
| DT | Direct transesterification |
| Chl-a | Chlorophyll-a |
References
- Wei, Z.; Chen, X.; Huang, Z.; Jiao, H.; Xiao, X. Insights into the removal of gaseous oxytetracycline by combined ozone and membrane biofilm reactor. Environ. Eng. Res. 2022, 27, 210469. [Google Scholar] [CrossRef]
- He, L.; Wang, D.; Wu, Z.; Li, S.; Lv, Y. Co-pyrolysis of pig manure and magnesium-containing waste residue and phosphorus recovery for planting feed corn. J. Water Process Eng. 2022, 49, 103146. [Google Scholar] [CrossRef]
- Ossowski, M.; Wlazło, Ł.; Nowakowicz-Dębek, B.; Florek, M. Effect of natural sorbents in the diet of fattening pigs on meat quality and suitability for processing. Animals 2021, 11, 2930. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, S.; Dajnowska, A.; Arciszewski, M.B.; Rudyk, H.; Śliwa, J.; Krakowiak, D.; Piech, M.; Nowakowicz-Dębek, B.; Czech, A. Effect of Fermented Rapeseed Meal in Feeds for Growing Piglets on Bone Morphological Traits, Mechanical Properties, and Bone Metabolism. Animals 2023, 13, 1080. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Li, X.; Ma, X.; Ru, S.; Qiu, T.; Lu, A. Veterinary antibiotics and estrogen hormones in manures from concentrated animal feedlots and their potential ecological risks. Environ. Res. 2021, 198, 110463. [Google Scholar] [CrossRef] [PubMed]
- Seibert, D.; Zorzo, C.F.; Borba, F.H.; de Souza, R.M.; Quesada, H.B.; Bergamasco, R.; Baptista, A.T.; Inticher, J.J. Occurrence, statutory guideline values and removal of contaminants of emerging concern by Electrochemical Advanced Oxidation Processes: A review. Sci. Total Environ. 2020, 748, 141527. [Google Scholar] [CrossRef] [PubMed]
- Egea-Corbacho, A.; Ruiz, S.G.; Alonso, J.M.Q. Removal of emerging contaminants from wastewater using nanofiltration for its subsequent reuse: Full-scale pilot plant. J. Clean. Prod. 2019, 214, 514–523. [Google Scholar] [CrossRef]
- Zubair, M.; Wang, S.; Zhang, P.; Ye, J.; Liang, J.; Nabi, M.; Zhou, Z.; Tao, X.; Chen, N.; Sun, K. Biological nutrient removal and recovery from solid and liquid livestock manure: Recent advance and perspective. Bioresour. Technol. 2020, 301, 122823. [Google Scholar] [CrossRef]
- Lou, E.G.; Harb, M.; Smith, A.L.; Stadler, L.B. Livestock manure improved antibiotic resistance gene removal during co-treatment of domestic wastewater in an anaerobic membrane bioreactor. Environ. Sci. Water Res. Technol. 2020, 6, 2832–2842. [Google Scholar] [CrossRef]
- Xiao, G.; Chen, J.; Show, P.L.; Yang, Q.; Ke, J.; Zhao, Q.; Guo, R.; Liu, Y. Evaluating the application of antibiotic treatment using algae-algae/activated sludge system. Chemosphere 2021, 282, 130966. [Google Scholar] [CrossRef]
- Ma, Z.; Li, M.; Wang, X.; Wang, Q.; Li, Q.; Wang, Y.; Zhang, Z.; Gao, J.; Gao, X.; Yuan, H. Selective and high-efficient removal of tetracycline from antibiotic-containing aqueous solution via combining adsorption with membrane pre-concentration. J. Water Process Eng. 2022, 50, 103281. [Google Scholar] [CrossRef]
- Chan, R.; Chiemchaisri, C.; Chiemchaisri, W. Effect of sludge recirculation on removal of antibiotics in two-stage membrane bioreactor (MBR) treating livestock wastewater. J. Environ. Health Sci. Eng. 2020, 18, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.K.; Kumar, M.; Singh, D.K. Insight into the amoxicillin resistance, ecotoxicity, and remediation strategies. J. Water Process Eng. 2021, 39, 101858. [Google Scholar] [CrossRef]
- Khan, W.; Nam, J.-Y.; Byun, S.; Kim, S.; Han, C.; Kim, H.-C. Emerging investigator series: Quaternary treatment with algae-assisted oxidation for antibiotics removal and refractory organics degradation in livestock wastewater effluent. Environ. Sci. Water Res. Technol. 2020, 6, 3262–3275. [Google Scholar] [CrossRef]
- Waddington, J.C.; Meng, X.; Naisbitt, D.J.; Park, B.K. Immune drug-induced liver disease and drugs. Curr. Opin. Toxicol. 2018, 10, 46–53. [Google Scholar] [CrossRef]
- Nam, J.-H.; Shin, J.-H.; Kim, T.-H.; Yu, S.; Lee, D.-H. Comparison of biological and chemical assays for measuring the concentration of residual antibiotics after treatment with gamma irradiation. Environ. Eng. Res. 2020, 25, 614–621. [Google Scholar] [CrossRef]
- Aryee, A.A.; Han, R.; Qu, L. Occurrence, detection and removal of amoxicillin in wastewater: A review. J. Clean. Prod. 2022, 368, 133140. [Google Scholar] [CrossRef]
- Saldarriaga, J.F.; Montoya, N.A.; Estiati, I.; Aguayo, A.T.; Aguado, R.; Olazar, M. Unburned material from biomass combustion as low-cost adsorbent for amoxicillin removal from wastewater. J. Clean. Prod. 2021, 284, 124732. [Google Scholar] [CrossRef]
- Hazra, M.; Durso, L.M. Performance efficiency of conventional treatment plants and constructed wetlands towards reduction of antibiotic resistance. Antibiotics 2022, 11, 114. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.-C.; Nam, J.-Y.; Kim, H.-W. Degradation of sulfonamide antibiotics and their intermediates toxicity in an aeration-assisted non-thermal plasma while treating strong wastewater. Chemosphere 2018, 209, 901–907. [Google Scholar] [CrossRef]
- Park, J.-G.; Lee, B.; Lee, U.-J.; Jun, H.-B. An anaerobic digester with microbial electrolysis cell enhances relative abundance of methylotrophic methanogens in bulk solution. Environ. Eng. Res. 2022, 27, 210666. [Google Scholar] [CrossRef]
- Jung, S.; Shetti, N.P.; Reddy, K.R.; Nadagouda, M.N.; Park, Y.-K.; Aminabhavi, T.M.; Kwon, E.E. Synthesis of different biofuels from livestock waste materials and their potential as sustainable feedstocks—A review. Energy Convers. Manag. 2021, 236, 114038. [Google Scholar] [CrossRef]
- Simsek, S.; Uslu, S. Comparative evaluation of the influence of waste vegetable oil and waste animal oil-based biodiesel on diesel engine performance and emissions. Fuel 2020, 280, 118613. [Google Scholar] [CrossRef]
- Vishwakarma, R.; Dhaka, V.; Ariyadasa, T.U.; Malik, A. Exploring algal technologies for a circular bio-based economy in rural sector. J. Clean. Prod. 2022, 354, 131653. [Google Scholar] [CrossRef]
- Daneshvar, E.; Zarrinmehr, M.J.; Koutra, E.; Kornaros, M.; Farhadian, O.; Bhatnagar, A. Sequential cultivation of microalgae in raw and recycled dairy wastewater: Microalgal growth, wastewater treatment and biochemical composition. Bioresour. Technol. 2019, 273, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, S.B.; Ahmad, A.; Imron, M.F.; Abdullah, S.R.S.; Othman, A.R.; Hasan, H.A. Potential of microalgae cultivation using nutrient-rich wastewater and harvesting performance by biocoagulants/bioflocculants: Mechanism, multi-conversion of biomass into valuable products, and future challenges. J. Clean. Prod. 2022, 365, 132806. [Google Scholar] [CrossRef]
- da Silva Rodrigues, D.A.; da Cunha, C.C.R.F.; Freitas, M.G.; de Barros, A.L.C.; e Castro, P.B.N.; Pereira, A.R.; de Queiroz Silva, S.; da Fonseca Santiago, A.; Afonso, R.J.d.C.F. Biodegradation of sulfamethoxazole by microalgae-bacteria consortium in wastewater treatment plant effluents. Sci. Total Environ. 2020, 749, 141441. [Google Scholar] [CrossRef]
- Leng, L.; Wei, L.; Xiong, Q.; Xu, S.; Li, W.; Lv, S.; Lu, Q.; Wan, L.; Wen, Z.; Zhou, W. Use of microalgae based technology for the removal of antibiotics from wastewater: A review. Chemosphere 2020, 238, 124680. [Google Scholar] [CrossRef]
- Deng, X.; Li, D.; Xue, C.; Chen, B.; Dong, J.; Tetteh, P.A.; Gao, K. Cultivation of Chlorella sorokiniana using wastewaters from different processing units of the silk industry for enhancing biomass production and nutrient removal. J. Chem. Technol. Biotechnol. 2020, 95, 264–273. [Google Scholar] [CrossRef]
- Gatidou, G.; Anastopoulou, P.; Aloupi, M.; Stasinakis, A.S. Growth inhibition and fate of benzotriazoles in Chlorella sorokiniana cultures. Sci. Total Environ. 2019, 663, 580–586. [Google Scholar] [CrossRef]
- Kim, H.W.; Vannela, R.; Zhou, C.; Harto, C.; Rittmann, B.E. Photoautotrophic nutrient utilization and limitation during semi-continuous growth of Synechocystis sp. PCC6803. Biotechnol. Bioeng. 2010, 106, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, L.; Qiu, S.; Ge, S. Determination of microalgal lipid content and fatty acid for biofuel production. BioMed. Res. Int. 2018, 2018, 1503126. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-C.; Kim, H.-W. Convergence of direct-transesterification and anaerobic digestion for improved bioenergy potentials of microalgae. J. Clean. Prod. 2018, 178, 749–756. [Google Scholar] [CrossRef]
- Guo, J.; Sun, S.; Liu, J. Conversion of waste frying palm oil into biodiesel using free lipase A from Candida antarctica as a novel catalyst. Fuel 2020, 267, 117323. [Google Scholar] [CrossRef]
- Rice, E.W.; Bridgewater, L. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012; Volume 10. [Google Scholar]
- Anjali, R.; Shanthakumar, S. Insights on the current status of occurrence and removal of antibiotics in wastewater by advanced oxidation processes. J. Environ. Manag. 2019, 246, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhou, Y.; Wang, Z.; Torres, O.L.; Guo, R.; Chen, J. Investigation of the removal mechanism of antibiotic ceftazidime by green algae and subsequent microbic impact assessment. Sci. Rep. 2017, 7, 4168. [Google Scholar] [CrossRef] [PubMed]
- Hom-Diaz, A.; Jaén-Gil, A.; Rodríguez-Mozaz, S.; Barceló, D.; Vicent, T.; Blánquez, P. Insights into removal of antibiotics by selected microalgae (Chlamydomonas reinhardtii, Chlorella sorokiniana, Dunaliella tertiolecta and Pseudokirchneriella subcapitata). Algal Res. 2022, 61, 102560. [Google Scholar] [CrossRef]
- Ricky, R.; Chiampo, F.; Shanthakumar, S. Efficacy of Ciprofloxacin and Amoxicillin Removal and the Effect on the Biochemical Composition of Chlorella vulgaris. Bioengineering 2022, 9, 134. [Google Scholar] [CrossRef]
- Zhong, X.; Zhu, Y.; Wang, Y.; Zhao, Q.; Huang, H. Effects of three antibiotics on growth and antioxidant response of Chlorella pyrenoidosa and Anabaena cylindrica. Ecotoxicol. Environ. Saf. 2021, 211, 111954. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Dong, S.; Zhou, D. Could co-substrate sodium acetate simultaneously promote Chlorella to degrade amoxicillin and produce bioresources? J. Hazard. Mater. 2021, 417, 126147. [Google Scholar] [CrossRef]
- Chai, W.S.; Tan, W.G.; Munawaroh, H.S.H.; Gupta, V.K.; Ho, S.-H.; Show, P.L. Multifaceted roles of microalgae in the application of wastewater biotreatment: A review. Environ. Pollut. 2020, 269, 116236. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xue, R.; Fu, L.; Chen, C.; Ndayisenga, F.; Zhou, D. Carbon dots enhance the recovery of microalgae bioresources from wastewater containing amoxicillin. Bioresour. Technol. 2021, 335, 125258. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, Q.; Li, J.; Chen, X.; Lang, Q.; Kuang, S. Combined effects of erythromycin and enrofloxacin on antioxidant enzymes and photosynthesis-related gene transcription in Chlorella vulgaris. Aquat. Toxicol. 2019, 212, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Mickiewicz, K.; Errington, J. Lysozyme counteracts β-lactam antibiotics by promoting the emergence of L-form bacteria. Cell 2018, 172, 1038–1049.e10. [Google Scholar] [CrossRef]
- Du, Y.; Wang, J.; Li, H.; Mao, S.; Wang, D.; Xiang, Z.; Guo, R.; Chen, J. The dual function of the algal treatment: Antibiotic elimination combined with CO2 fixation. Chemosphere 2018, 211, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Mofijur, M.; Parisa, T.A.; Islam, N.; Kusumo, F.; Inayat, A.; Badruddin, I.A.; Khan, T.Y.; Ong, H.C. Progress and challenges of contaminate removal from wastewater using microalgae biomass. Chemosphere 2021, 286, 131656. [Google Scholar] [CrossRef]
- Xiong, Q.; Hu, L.-X.; Liu, Y.-S.; Zhao, J.-L.; He, L.-Y.; Ying, G.-G. Microalgae-based technology for antibiotics removal: From mechanisms to application of innovational hybrid systems. Environ. Int. 2021, 155, 106594. [Google Scholar] [CrossRef]
- Xie, B.; Tang, X.; Ng, H.Y.; Deng, S.; Shi, X.; Song, W.; Huang, S.; Li, G.; Liang, H. Biological sulfamethoxazole degradation along with anaerobically digested centrate treatment by immobilized microalgal-bacterial consortium: Performance, mechanism and shifts in bacterial and microalgal communities. Chem. Eng. J. 2020, 388, 124217. [Google Scholar] [CrossRef]
- Li, D.; Amoah, P.K.; Chen, B.; Xue, C.; Hu, X.; Gao, K.; Deng, X. Feasibility of growing Chlorella sorokiniana on cooking cocoon wastewater for biomass production and nutrient removal. Appl. Biochem. Biotechnol. 2019, 188, 663–676. [Google Scholar] [CrossRef]
- Zhu, L.; Li, S.; Hu, T.; Nugroho, Y.K.; Yin, Z.; Hu, D.; Chu, R.; Mo, F.; Liu, C.; Hiltunen, E. Effects of nitrogen source heterogeneity on nutrient removal and biodiesel production of mono-and mix-cultured microalgae. Energy Convers. Manag. 2019, 201, 112144. [Google Scholar] [CrossRef]
- Hu, D.; Zhang, J.; Chu, R.; Yin, Z.; Hu, J.; Nugroho, Y.K.; Li, Z.; Zhu, L. Microalgae Chlorella vulgaris and Scenedesmus dimorphus co-cultivation with landfill leachate for pollutant removal and lipid production. Bioresour. Technol. 2021, 342, 126003. [Google Scholar] [CrossRef]
- Deng, X.-Y.; Gao, K.; Zhang, R.-C.; Addy, M.; Lu, Q.; Ren, H.-Y.; Chen, P.; Liu, Y.-H.; Ruan, R. Growing Chlorella vulgaris on thermophilic anaerobic digestion swine manure for nutrient removal and biomass production. Bioresour. Technol. 2017, 243, 417–425. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, W.; Chen, H.; Zhan, J.; He, C.; Wang, Q. Ammonium Nitrogen Tolerant Chlorella Strain Screening and Its Damaging Effects on Photosynthesis. Front. Microbiol. 2019, 9, 3250. [Google Scholar] [CrossRef]
- Odjadjare, E.C.; Mutanda, T.; Chen, Y.-F.; Olaniran, A.O. Evaluation of pre-chlorinated wastewater effluent for microalgal cultivation and biodiesel production. Water 2018, 10, 977. [Google Scholar] [CrossRef]
- Gao, F.; Peng, Y.-Y.; Li, C.; Yang, G.-J.; Deng, Y.-B.; Xue, B.; Guo, Y.-M. Simultaneous nutrient removal and biomass/lipid production by Chlorella sp. in seafood processing wastewater. Sci. Total Environ. 2018, 640, 943–953. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, C.; Liu, J.; Sun, S.; Zhao, Y.; Wei, J. Effects of exogenous GR24 on biogas upgrading and nutrient removal by co-culturing microalgae with fungi under mixed LED light wavelengths. Chemosphere 2021, 281, 130791. [Google Scholar] [CrossRef]
- Daneshvar, E.; Zarrinmehr, M.J.; Hashtjin, A.M.; Farhadian, O.; Bhatnagar, A. Versatile applications of freshwater and marine water microalgae in dairy wastewater treatment, lipid extraction and tetracycline biosorption. Bioresour. Technol. 2018, 268, 523–530. [Google Scholar] [CrossRef]
- Bai, X.; Acharya, K. Algae-mediated removal of selected pharmaceutical and personal care products (PPCPs) from Lake Mead water. Sci. Total Environ. 2017, 581, 734–740. [Google Scholar] [CrossRef]
- Chen, J.; Xie, S. Overview of sulfonamide biodegradation and the relevant pathways and microorganisms. Sci. Total Environ. 2018, 640, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Norvill, Z.N.; Toledo-Cervantes, A.; Blanco, S.; Shilton, A.; Guieysse, B.; Muñoz, R. Photodegradation and sorption govern tetracycline removal during wastewater treatment in algal ponds. Bioresour. Technol. 2017, 232, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-Q.; Zheng, H.-S.; Li, S.; Du, J.-S.; Feng, X.-C.; Yin, R.-L.; Wu, Q.-L.; Ren, N.-Q.; Chang, J.-S. Removal of cephalosporin antibiotics 7-ACA from wastewater during the cultivation of lipid-accumulating microalgae. Bioresour. Technol. 2016, 221, 284–290. [Google Scholar] [CrossRef]
- Braun, J.V.; dos Santos, V.O.; Fontoura, L.A.; Pereira, E.; Napp, A.; Seferin, M.; Lima, J.; Ligabue, R.; Vainstein, M.H. GC-FID methodology validation for the fatty esters content determination in biodiesel with hexadecyl acetate as the internal standard. Quim. Nova 2017, 40, 1111–1116. [Google Scholar] [CrossRef]
- Chaudhary, N.K.; Mishra, P. Spectral Investigation and In Vitro Antibacterial Evaluation of NiII and CuI Complexes of Schiff Base Derived from Amoxicillin and α-Formylthiophene (αft). J. Chem. 2015, 2015, 136285. [Google Scholar] [CrossRef]
- Cheng, J.; Qiu, Y.; Huang, R.; Yang, W.; Zhou, J.; Cen, K. Biodiesel production from wet microalgae by using graphene oxide as solid acid catalyst. Bioresour. Technol. 2016, 221, 344–349. [Google Scholar] [CrossRef]
- Lee, J.-C.; Jang, J.K.; Kim, H.-W. Sulfonamide degradation and metabolite characterization in submerged membrane photobioreactors for livestock excreta treatment. Chemosphere 2020, 261, 127604. [Google Scholar] [CrossRef]
- Gao, J.; Cui, Y.; Tao, Y.; Huang, L.; Peng, D.; Xie, S.; Wang, X.; Liu, Z.; Chen, D.; Yuan, Z. Multiclass method for the quantification of 92 veterinary antimicrobial drugs in livestock excreta, wastewater, and surface water by liquid chromatography with tandem mass spectrometry. J. Sep. Sci. 2016, 39, 4086–4095. [Google Scholar] [CrossRef]
- Lee, H.-J.; Ryu, H.-D.; Chung, E.G.; Kim, K.; Lee, J.K. Characteristics of veterinary antibiotics in intensive livestock farming watersheds with different liquid manure application programs using UHPLC-q-orbitrap HRMS combined with on-line SPE. Sci. Total Environ. 2020, 749, 142375. [Google Scholar] [CrossRef]






| Characteristics | Unit | Value |
|---|---|---|
| pH | - | 7.5 |
| CODcr (Chemical Oxygen Demand) | mg COD/L | 280 |
| T-N (Total Nitrogen) | mg N/L | 120 |
| T-P (Total Phosphorus) | mg P/L | 1.5 |
| AMX Strength (mg/L) | Initial Chl-a (mg/m3) | Final Chl-a (mg/m3) | Average Daily Microalgae Growth (mg/m3/Day) | Specific Growth Rate (Days−1) |
|---|---|---|---|---|
| 0 | 328 1 | 8583 | 1651 | 0.87 |
| 0.01 | 7947 | 1519 | 0.46 | |
| 0.5 | 7108 | 1345 | 0.35 | |
| 1 | 7475 | 1435 | 0.43 | |
| 5 | 6194 | 1150 | 0.34 | |
| 20 | 6052 | 1080 | 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Jeong, S.; Lee, Y.; Lee, J.-C.; Kim, H.-W. The Crucial Impact of Microbial Growth and Bioenergy Conversion on Treating Livestock Manure and Antibiotics Using Chlorella sorokiniana. Processes 2024, 12, 252. https://doi.org/10.3390/pr12020252
Kim H-J, Jeong S, Lee Y, Lee J-C, Kim H-W. The Crucial Impact of Microbial Growth and Bioenergy Conversion on Treating Livestock Manure and Antibiotics Using Chlorella sorokiniana. Processes. 2024; 12(2):252. https://doi.org/10.3390/pr12020252
Chicago/Turabian StyleKim, Hee-Jun, Sangjun Jeong, YeonA Lee, Jae-Cheol Lee, and Hyun-Woo Kim. 2024. "The Crucial Impact of Microbial Growth and Bioenergy Conversion on Treating Livestock Manure and Antibiotics Using Chlorella sorokiniana" Processes 12, no. 2: 252. https://doi.org/10.3390/pr12020252