

Anti-Glioblastoma Potential and Phenolic Profile of Berry Juices

1
Faculty of Food Technology, University of Josip Juraj Strossmayer in Osijek, Franje Kuhača 18, 31000 Osijek, Croatia
2
Department of Cell and Molecular Biology, University of Mississippi Cancer Institute, University of Mississippi Medical Center, 2500 North State Street, Jackson, MS 39216, USA
3
Plants for Human Health Institute, Department of Food Bioprocessing and Nutrition Sciences, North Carolina Research Campus, North Carolina State University, 600 Laureate Way, Kannapolis, NC 28081, USA
4
Department of Food, Bioprocessing and Nutrition Sciences, North Carolina State University, 116C Schaub Hall, Raleigh, NC 27695, USA
*
Authors to whom correspondence should be addressed.
Processes 2024, 12(2), 242; https://doi.org/10.3390/pr12020242
Submission received: 20 December 2023
/
Revised: 20 January 2024
/
Accepted: 22 January 2024
/
Published: 23 January 2024
(This article belongs to the Special Issue Phytochemicals: Extraction, Optimization, Identification, Biological Activities, and Applications in the Food, Nutraceutical, and Pharmaceutical Industries)
Abstract
:Glioblastoma is one of the most aggressive and lethal brain tumors. Due to the failure of conventional chemotherapies and targeted drugs pursuit of natural, less toxic agents is on the rise as well as their utilization in glioblastoma treatment. Consequently, this study explores the antiproliferative potential of selected berry juices (wild blackberry (Rubus discolor), dwarf elderberry (Sambucus ebulus), and raspberry (Rubus idaeus)) on glioblastoma cells (U87-MG and GBM43) in comparison to temozolomide. The juices were assessed for total phenolic content, proanthocyanins, polyphenol profiles, and antioxidant activity. Wild blackberry and dwarf elderberry juices exhibited higher total polyphenols, proanthocyanins, and monomeric anthocyanins compared to raspberry juice. HPLC analysis revealed distinctive anthocyanins, flavonoids, and phenolic acids in each juice. With the DPPH assay, the highest antioxidant potential had wild blackberry juice, while with other assays dwarf elderberry juice had the highest potential. Antiproliferative effects were dose-dependent, with wild blackberry juice demonstrating the highest potency, surpassing temozolomide in inhibiting GBM43 cell proliferation. In U87 cells, all juices exhibited antiproliferative effects, with wild blackberry showing the strongest impact. This study highlights the potential of wild blackberry juice as a potent natural agent against glioblastoma, suggesting its superiority over the conventional treatment.
1. Introduction
Nutritional guidelines have over the years highlighted the importance of daily consumption of fruits, especially berries. Berries are well known for their high content of flavonoids, phenolic acids, tannins, and especially anthocyanins, due to which they benefit from a “health halo”. These phenolic compounds, through numerous in vitro, pre-clinical, clinical and epidemiological studies, have been linked to many beneficial health effects like improvement of the lipid balance, regulation of hyperglycemia, protection from cardiovascular disorders, possible regulation of diabetes, anticancer potential, anti-inflammation properties and antioxidant potential [1,2,3,4,5,6].
Another very important and popular aspect among the general population is the utilization of bioactive phytochemicals, among which anthocyanins have a crucial role, in order to substitute synthetic compounds used in chemotherapeutic or chemopreventive applications [7,8,9]. Anthocyanins exhibit numerous anticancer effects such as antioxidation, lipid peroxidation, antiproliferation, cytotoxicity anti-inflammation, cell cycle perturbations, epidermal growth factor receptor inhibition, and apoptosis [7].
Glioblastoma is one of the most aggressive and lethal brain tumors which is characterized by the existence of weakly differentiated anaplastic cells immersed in necrotic zones of brain tissue [10,11,12]. Despite all the efforts that were made through the years in the treatment of this tumor, it is still characterized by a low survival rate of only 13 months after diagnosis and even in the most positive situations, the majority of patients die within two years. In clinical trials, a 5-year survival rate of only 4–5% of patients has been achieved [11,12,13,14,15,16]. Conventional glioblastoma treatment comprises surgical resection followed by radiotherapy and chemotherapy including temozolomide [10,12,15]. However, the prognosis for these patients remains weak as already mentioned due to the high tumor recurrence [10,12,14,15,16]. These tumor cells are quite heterogeneous and grow rapidly, invading and infiltrating the nearby healthy brain tissues which makes the complete tumor resection challenging. Residual tumor cells cause the initiation of the occurrence of secondary glioblastoma lesions, which are even more resistant to therapy than rapidly proliferating primary tumor cells, leading to tumor recurrence [10]. In addition to the tumor recurrence and therapy resistance, additional challenges are the inability of drugs to cross the blood–brain barrier and pass through the blood–brain–tumor barrier which is formed during the later stages of tumor growth [10,17,18,19]. All those challenges and failure of conventional chemotherapies and targeted drugs govern scientist in the direction to find natural, less toxic agents and explore their utilization in glioblastoma treatment [20].
The most commonly used cell lines in cancer research, particularly in the study of glioblastoma multiforme (GBM), which is a type of malignant brain tumor, are U87-MG and GBM43. U87-MG cells are characterized by their ability to form tumors in experimental animals and are often used as a model system for studying glioblastoma. These glioblastoma cells are used to investigate various aspects of glioblastoma biology, including tumor growth, invasion, and response to therapies. They are also employed in drug testing and preclinical studies [21,22,23]. The specific characteristics of GBM43 cells may vary depending on factors such as the source and handling of the cells. They are likely to share features common to glioblastoma cells, such as rapid proliferation and invasiveness. These cells are used to gain insights into the biology of glioblastoma and to test potential therapeutic interventions [24,25,26].
The aim of this study was to evaluate the antiproliferative effect of selected berry juices (wild blackberry, dwarf elderberry and raspberry) on mentioned glioblastoma cells (U87-MG and GBM43). The effect of selected berry juices on glioblastoma cells was compared with temozolomide, the current drug of choice for treatment of glioblastoma. Additionally, berry juices were evaluated for their total phenolic content, proanthocyanidin content, individual polyphenols and antioxidant activity.
2. Materials and Methods
2.1. Chemicals
Products of Sigma-Aldrich (St. Louis, MO, USA) were 4-dimethylaminocinnamaldehyde, trolox, 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt, 2,2-diphenyl-1-picrylhydrazyl and analytical standards of chlorogenic acid, gallic acid, ellagic acid, rutin and (-)-epicatechin. Standards of hyperoside, neochlorogenic acid and anthocyanins (cyanidin-3-glucoside, cyanidin-3-galactoside, cyanidin-3-rutinoside and cyanidin-3-sophoroside) were bought from Extrasynthese (Genay, France). From T.T.T. (Sveta Nedelja, Croatia) sodium carbonate was procured, while Folin–Ciocalteu reagent and potassium persulfate were from Kemika (Zagreb, Croatia). HPLC-grade orthophosphoric acid was product of Fisher Scientific (Loughborough, UK) and HPLC-grade methanol was from J.T. Baker (Deventer, The Netherlands). From Acros Organic (Geel, Belgium), cupric chloride, neocuproine and 2,4,6-tri(2-pyridyl)-s-triazine (TPTZ) were bought. From ThermoFisher Scientific (Waltham, MA, USA), Thiazolyl Blue tetrazolium bromide, 98% (2-(3,5-diphenyltetrazol-2-ium-2-yl)-4,5-dimethyl-1,3-thiazole;bromide), Temozolomide (3-methyl-4-oxoimidazol[5,1-d][1,2,3,5]tetrazine-8-carboxamide), DMEM media w/L-Glutamine, 4.5 g/L glucose and sodium pyruvate, Penicillin Streptomycin Solution (10,000 U/mL Penicillin, 10,000 μg/mL Streptomycin in 0.85% NaCl), and 0.25% Trypsin 0.1% EDTA were obtained. U87-MB and GBM43 cells were acquired from Mayo Clinic (Rochester, MN, USA). Fetal bovine serum was bought from R&D Systems.
2.2. Preparation of Berry Juice
Wild blackberry (Rubus discolor), dwarf elderberry (Sambucus ebulus), and raspberry (Rubus idaeus) fruits were collected at location 46°18′39.7″ N 16°32′67.7″ E near Varaždin, Croatia. About 2 kg of each fruits were collected, washed and pressed. The obtained juice was filtered through cheesecloth and afterwards juice (300 mL) was thermally treated at 90 °C for two minutes in order to inactivate naturally present enzymes that could lead to degradation of polyphenols.
2.3. Spectrophotometric Analysis of Total Polyphenols, Monomeric Anthocyanins and Proanthocyanidins
The method described by Singleton and Rossi [27] was used for the determination of total polyphenols in samples. Folin–Ciocalteu reagent (7.5%) was prepared and 10 mL was mixed with a 0.2 mL of sample, 1.8 mL demineralized water and 8 mL of sodium carbonate solution (7.5%). This mixture was kept in the dark for 120 min prior to the measurement of the absorbance at 765 nm. The calibration curve was created for gallic acid. Results were expressed as g of gallic acid equivalents per L of the juice (g GAE/L). Each sample was analyzed in triplicate and all measurements were conducted using a UV/Vis spectrophotometer (Cary 60 UV-Vis, Agilent Technologies, Santa Clara, CA, USA).
Monomeric anthocyanins were determined by the pH differential method [28]. To conduct the analyses, it was necessary to prepare two different buffers (0.025 M KCl at pH 1 and 0.4 M sodium acetate at pH 4.5). Each buffer (2.8 mL) was mixed with the sample (0.2 mL). After these mixtures were kept in the dark for 15 min, absorbance was measured at 515 nm and 700 nm. The concentration of monomeric anthocyanins was expressed as mg of cyanidin-3-glucoside per L of the juice (mg cyanidin-3-glucoside/L).
To determine the concentration of proanthocyanidins, the DMAC method was applied [29]. Briefly, DMAC (4-dimethylaminocinnamaldehyde) reagent was prepared and mixed (1 mL) with sample and acidified ethanol. The absorbance of the samples was measured at 640 nm after the samples were kept in dark for 30 min. The calibration curve was set for procyanidin B2 and concentration of proanthocyanidins was expressed as mg of procyanidin B2 equivalent per L of the juice (mg B2E/L). All samples were analyzed in triplicate.
2.4. Evaluation of Antioxidant Activity—ABTS, DPPH, FRAP and CUPRAC Assays
Firstly, the ABTS assay by Arnao et al. [30] was conducted. After ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt) reagent was prepared, 3.2 mL was mixed with 0.2 mL of the diluted samples and after 95 min, and absorbance was measured at 734 nm. Secondly, the DPPH (2,2-diphenyl-1-picrylhydrazyl) solution was prepared and 3 mL was mixed with 0.2 mL of the diluted samples. Absorbance was read at 517 nm after 15 min. This assay was described in detail elsewhere [31]. Thirdly, the ferric-reducing ability was determined using the method of Benzie and Strain [32]. FRAP reagent was prepared and 3 mL was mixed with 0.2 mL of the diluted samples and absorbance was read at 593 nm after 30 min. Finally, the CUPRAC assay was applied for the determination of cupric ion-reducing antioxidant capacity. This assay was described in detail by Apak et al. [33]. Copper chloride, neocuproine and ammonium acetate buffer (pH 7) solution were mixed in a ratio of 1:1:1 and 0.2 mL of the diluted samples were addes. After 30 min absorbance was read at 450 nm. All samples were analyzed in triplicate. Calibration curves were created using Trolox, and for all assays, results are expressed as µmol of Trolox equivalents per 100 mL of juice (µmol TE/100 mL).
2.5. Sample Preparation for High Performance Liquid Chromatography
Solid phase extraction was conducted using a commercial sorbent, StrataTM-X 33 μm Polymeric Reversed Phase from Phenomenex (Torrance, CA, USA), in order to exclude impurities. After the cartridges were placed in a vacuum manifold operated at room temperture, they were preconditioned with HPLC-grade methanol. A solution of acetic acid (1% in water) was added next and then the sample was added by dripping it and allowing it to form a compact ring in the cartridges. Elution of polyphenols was performed with methanol after the cartridges were dry [34,35]. The eluents were collected and injected into the HPLC system.
2.6. Determination of Individual Polyphenols Using Reversed Phase HPLC
For the evaluation of individual polyphenols in samples, Agilent HPLC system 1260 Infinity II (Agilent Technology, Santa Clara, CA, USA) was used. This system consisted of a quaternary pump, a vial sampler, a column (Poroshell 120 EC C-18, 4.6 × 100 mm, 2.7 µm) and diode array detector that was recording in the range from 190 to 600 nm. The method used was previously published by Buljeta et al. [36]. Orthophosphoric acid (0.1%) was used as mobile phase A and HPLC-grade methanol was used as mobile phase B. Injected volume was 5 µL and the flow rate was 1 mL/min. The calibration curves for standards of cyanidin-3-glucoside, cyanidin-3-galactoside, cyanidin-3-rutinoside, cyanidin-3-sophoroside, rutin, hyperoside, ellagic acid, gallic acid, neochlorogenic acid, chlorogenic acid, and (-)-epicatechin were created and linearity was confirmed with r2 > 0.99. All measurements were carried out in duplicate and concentrations of individual polyphenols were expressed as mg of polyphenol per L of the juice (mg/L).
2.7. Antiproliferative Effects of Berry Fruit Juices on Glioblastoma Cells
Glioblastoma cells (U87-MG and GBM43) were cultured in 96-well plates, a standard format for high-throughput screening and analysis. After 24 h, cells were exposed to fruit juices at concentrations of 1%, 2%, and 3% in the culture medium (DMEM-10% fetal bovine serum-1% penicillin/streptomycin). Untreated cells served as the negative control, while temozolomide, the current standard treatment for glioblastoma, was used as the positive control for inducing cell death. After 3 days of treatment, cell survival was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay [37]. This colorimetric assay measures the metabolic activity of living cells. Viable cells convert the yellow MTT reagent into purple formazan crystals, quantified by absorbance at a specific wavelength. Higher absorbance values indicate greater cell viability, reflecting the effectiveness of the treatment. Percentage survival for each treatment group was calculated as follows:
Percentage of survival = (Absorbance of treated cells/Absorbance of untreated control cells) × 100.
2.8. Statistical Analysis
Comparisons of the obtained results for polyphenols were carried out by analysis of variance (ANOVA) and Fisher’s least significant difference (LSD), with the significance defined at p < 0.05. The software program STATISTICA 13.1 (StatSoft Inc., Tulsa, OK, USA) was applied for statistical analyses. Comparisons of the obtained results for antiproliferative effects of berry fruit juices on glioblastoma cells were graphed and statistically analyzed using one-way analysis of variance (ANOVA) to determine significant differences in survival percentages between groups. These analyses were performed using GraphPad Prism version 10.1.2.
3. Results
3.1. Phenolic Compounds and Antioxidant Activity of Berry Juices
Amounts of total polyphenols, proanthocyanidins and monomeric anthocyanins of selected berry juices are presented in Table 1. From the obtained results, it can be seen that wild blackberry and dwarf elderberry juices had approximately 3.2 g/L of total polyphenols while raspberry juice had a significantly lower amount, 1.67 g/L (p < 0.05). Investigated juices significantly differ in proanthocyanidins and monomeric anthocyanins amounts. The amount of pronanthocyanidins in juices declined in the following: order dwarf elderberry > wild blackberry > raspberry juice (71.21 mg/L > 56.21 mg/L > 32.32 mg/L) (p < 0.05). A slightly different tendency was observed for the amounts of monomeric anthocyanins, and their amounts were ranked in the following order: wild blackberry > dwarf elderberry > raspberry juice (759.99 mg/L > 433.17 mg/L > 230.39 mg/L) (p < 0.05).
HPLC profiles of individual polyphenols of investigated juices are presented in Table 2. It is evident from the results that juices differ in the type of anthocyanins inherent in their phytochemical profiles, as well as in phenolic acids and flavonoids. Dwarf elderberry juice contained cyanidin-3-galactoside (261.40 mg/L) while the other two investigated juices did not contain this anthocyanin. Wild blackberry juice contained high amounts of cyanidin-3-glucoside and cyanidin-3-rutinoside (389.60 and 269.72 mg/L, respectively). These two anthocyanins were also identified in raspberry juice but in significantly lower amounts (39.11 and 34.28 mg/L, respectively). The most abundant anthocyanin in raspberry juice was cyanidin-3-sophoroside (210.65 mg/L). Two flavonoids were detected in wild blackberry and dwarf elderberry juices, hyperoside and rutin. Dwarf elderberry and wild blackberry juice contained both of these flavonoids, but in different amounts. In dwarf elderberry juice, hyperoside was evaluated at 34.19 mg/L and rutin at 109.72 mg/L. The opposite trend was observed in wild blackberry juice, i.e., hyperoside was evaluated in the higher amount (73.27 mg/L) than rutin (3.37 mg/L). Raspberry contained only rutin in the amount of 14.70 mg/L.
Gallic, ellagic, chlorogenic and neochlorogenic acids were detected in the berry juices. Chlorogenic and neochlorogenic acids (160.02 and 27.18 mg/L, respectively) were identified only in dwarf elderberry juice in addition to gallic acid. Gallic acid was also identified in wild blackberry juice, and those two juices contained 4.57 mg/L of this phenolic acid. In raspberry and wild blackberry juices, ellagic acid was determined at 3.41 mg/L and 5.14 mg/L, respectively.
Results of the evaluation of antioxidant activities of berry juices are presented in Table 3. Antioxidant activities were evaluated using the following assays: DPPH (with DPPH radicals), ABTS (with ABTS cation radicals), FRAP (ferric reducing antioxidant power) and CUPRAC (cupric ion reducing capability) assays. Application of DPPH assay revealed that wild blackberry juice had the highest antioxidant potential, followed by the dwarf elderberry and raspberry juices (13.25, 9.86 and 6.95 µmol TE/100 mL, respectively) (p < 0.05). However, with other applied assays, a different trend was observed. Dwarf elderberry juice had the highest antioxidant potential, followed by raspberry and wild blackberry juices. For ABTS assay, these values were 26.18, 15.56 and 6.08 µmol TE/100 mL, respectively, (p < 0.05). The lowest values were obtained by FRAP (2.11, 1.38 and 0.59 µmol TE/100 mL, respectively) (p < 0.05) while the highest by CUPRAC assay (107.78, 64.00 and 27.37 µmol TE/100 mL, respectively) (p < 0.05). Comparing these results of antioxidant activity with individual polyphenols determined in berry juices, it is evident that antioxidant activity depends on the structure of polyphenols and the mechanism of action of each applied assay. Wild blackberry juice contained the highest amount of anthocyanins and had the highest antioxidant activity determined by a DPPH assay. For all other assays, dwarf elderberry juice possessed the highest antioxidant activity. It has to be noted that this juice contained, next to anthocyanins, a significant amount of flavonoids and phenolic acids in contrast to the other two berry juices. Even though raspberry juice contained a lower amount of anthocyanins and a much lower amount of flavonoids and phenolic acids than wild blackberry juice, through utilization of ABTS, FRAP and CUPRAC assay, a higher antioxidant activity was achieved than for wild blackberry juice. This can be explained by well-known phenomena of antagonistic or synergistic effects of phenolic compounds.
3.2. Antiproliferative Effects of Berry Juices
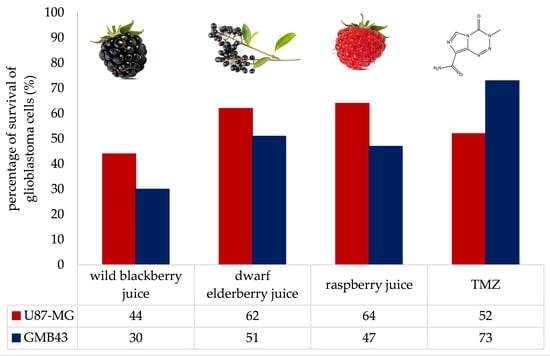
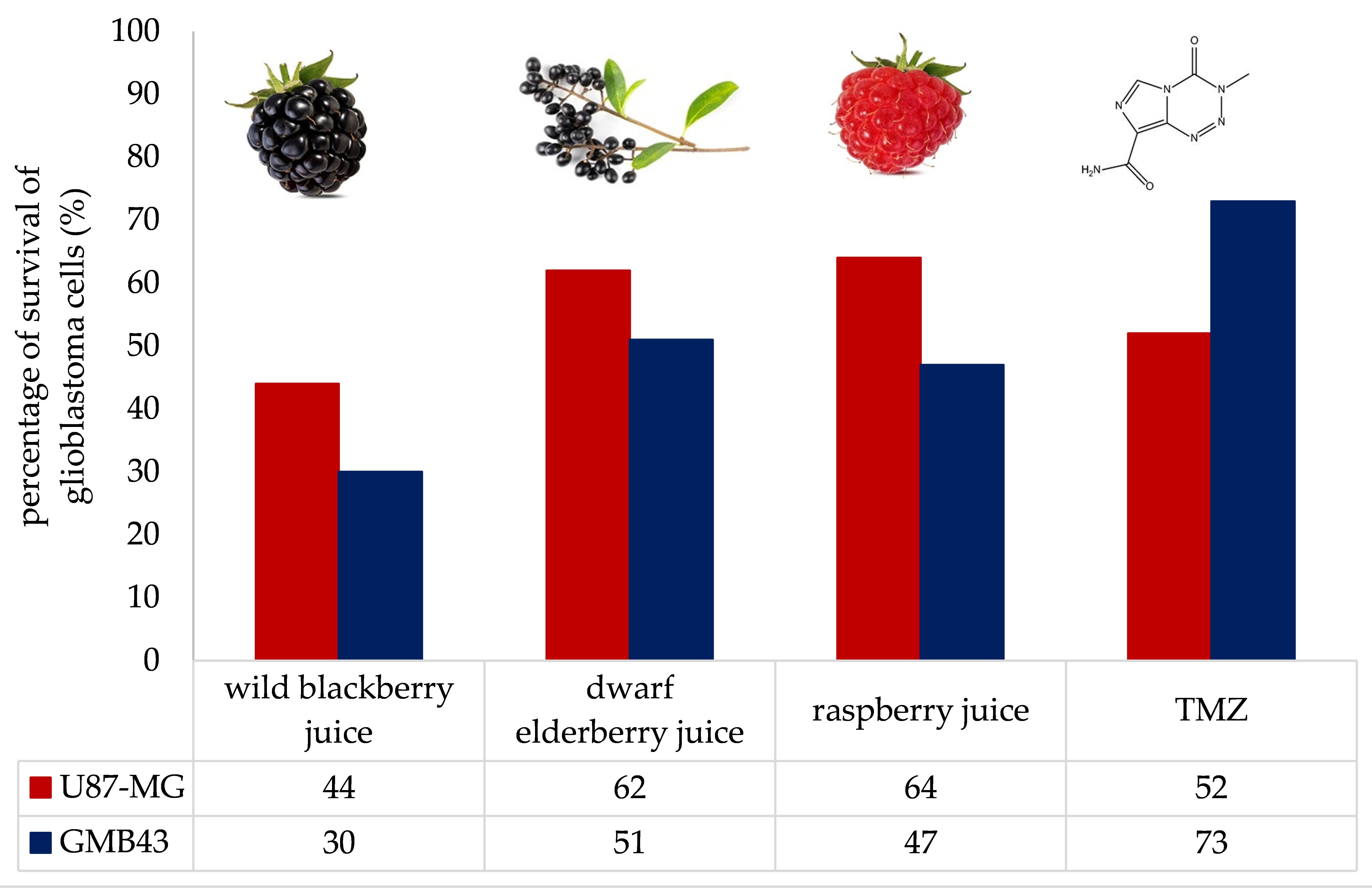
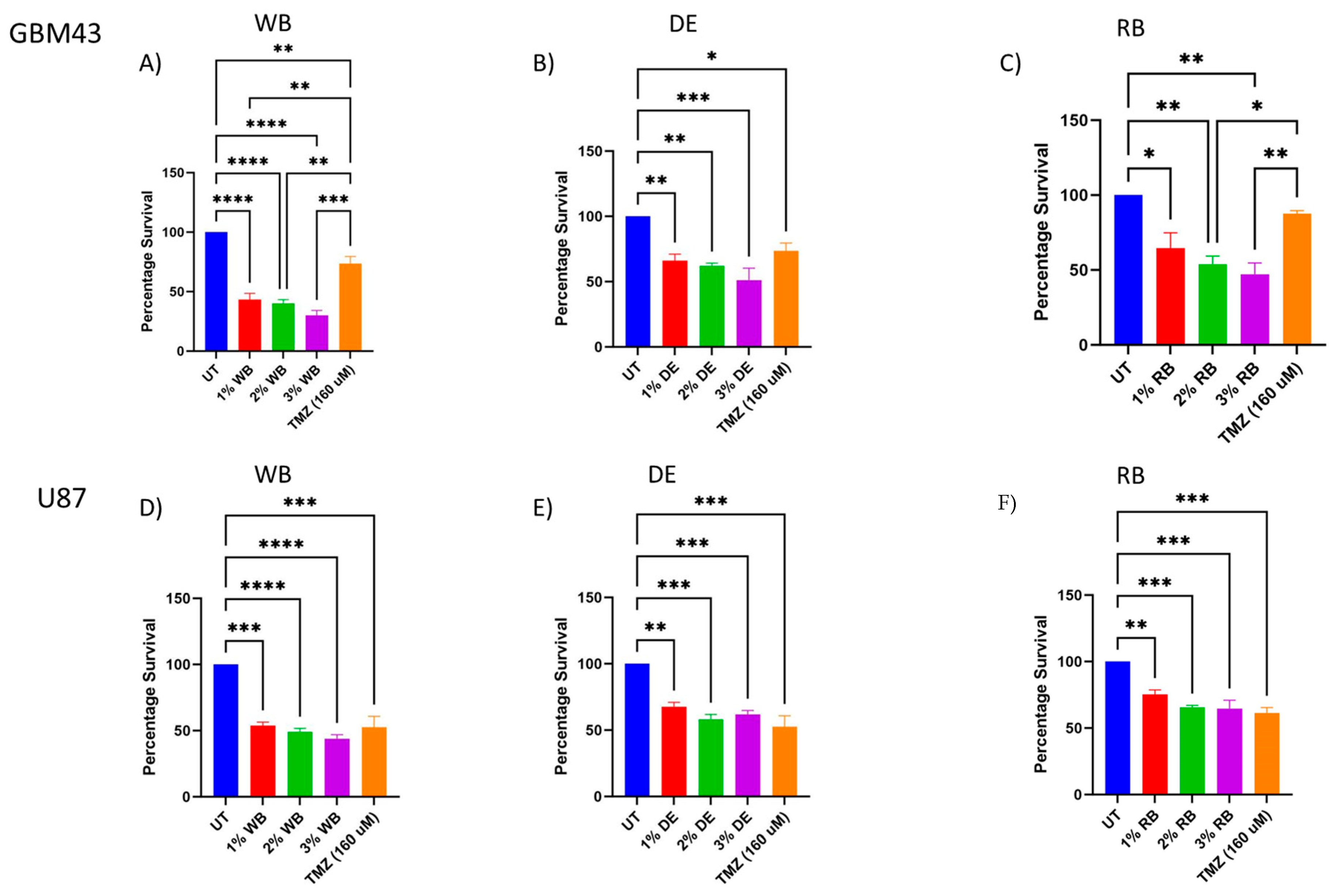
Results of the antiproliferative effect trials of berry juices on GBM43 and U87-MG cells are presented in Figure 1. Their potential for inhibition of proliferation of GBM43 and U87-MG cells was compared with temozolomide (TMZ), the drug of choice for the treatment of selected tumor cells. Juices were used in the amounts of 1%, 2% and 3%. The results demonstrated that berry juices exhibited antiproliferative effects on both GBM43 and U87-MG cells with different potency in a dose-dependent manner. The percentage of survival of U87-MG cells was 52.6% when the inhibition was conducted with TMZ. The potential for inhibition of proliferation of U87-MG cells decreased in the following order: wild blackberry > dwarf elderberry > raspberry. For the wild blackberry juice percentages of survival of U87 cells were 53.8%, 49% and 44%, respectively, when cells were treated with 1%, 2% and 3% of juice. The percentage of survival of U87 cells when treated with 1% of dwarf elderberry juice was 67.7%. 58.2% and approximately 60% when treated with higher concentrations. Raspberry juice was the least potent juice for this population of tumor cells, and percentages of survival were 75% when they were treated with 1% of juice and approximately 65% when treated with higher concentrations.
Remarkably, GBM43 cells, which are typically resistant to temozolomide, exhibited significantly higher inhibition when treated with berry juices compared to the temozolomide treatment. The percentage of survival of GBM43 cells was 73.7% when inhibition was conducted with TMZ. The potential for inhibition of proliferation of GBM43 cells decreased in the following order: wild blackberry > raspberry > dwarf elderberry. The percentages of survival of GBM43 cells were 43.3%, 40.2% and 30% when treated with 1%, 2% and 3% of wild blackberry juice. Treatment of GBM43 cells with 1%, 2% and 3% of raspberry juice resulted in survival rates of 64.5%, 54 and 47.2%, respectively. Percentages of survival rates of 66%, 62% and 51.2% were achieved when GBM43 cells were treated with 1%, 2% and 3% of dwarf elderberry juice.
These results of the in vitro study suggest that wild blackberry juice may be the most potent among the tested berry juices in combating these types of cancer. In both cell lines, wild blackberry juice demonstrated the most pronounced impact on the reduction in cell survival.
4. Discussion
In this study, we selected three berry juices, namely dwarf elderberry, raspberry and wild blackberry for the treatment of glioblastoma tumor cells (U87-MG and GBM43). Selected juices differ in their phenolic composition and antioxidant capacity. Wild blackberry and dwarf elderberry juices exhibited a higher content of total polyphenols, proanthocyanins, and monomeric anthocyanins compared to raspberry juice. HPLC analyses revealed distinctive anthocyanins, flavonoids, and phenolic acids in each juice. Comparisons of concentrations of individual phenolic compounds revealed that anthocyanins were the most prevalent flavonoid molecules in the profiles of all the juices. Wild blackberry and dwarf elderberry juice contained significant amounts of quercetin derivates, while the dwarf elderberry juice additionally contained significant amounts of phenolic acids. Wild blackberry juice had the highest potency of inhibition against both types of glioblastoma cells probably due to the highest concentration of anthocyanins, which is in agreement with other studies [7,11,16,38,39,40,41,42,43,44,45,46]. All berry juices selected for this study had cyanidin-based anthocyanins, but the base structure differed for each berry species which may account for their different potencies in inhibition of glioblastoma cells.
Other studies also highlighted anthocyanins among polyphenols as the compounds responsible for the inhibition of the proliferation of different tumor cells. Compared to other flavonoids, anthocyanins were more effective for the inhibition of direct cell growth [38]. Research on the potential for inhibition of migration of glioblastoma cells by anthocyanidins demonstrated that delphinidin, petunidin, and cyanidin could potentially inhibit these cancer cells. This property was ascribed to their structure, i.e., the number of hydroxyl groups on the B-ring. Delphinidin (with three hydroxyl groups) demonstrated the best inhibitory effect (83%) while the other two anthocyanidins (with two hydroxyl groups) demonstrated an inhibitory effect of 48% [16]. However, comparing the effectiveness of delphinidin and cyanidin in antiproliferative and apoptotic effects in MCF7 human breast cancer cells revealed that cyanidin was more effective [7]. Even though Lamy et al. [16] pointed out that aglycones are more potent in inhibition of tumor cell growth, Jing et al. [39] observed that the glycosides of anthocyanins might be more efficient since they can handicap glucose transport and cause inhibition of energy metabolism, which, in turn, may lead to mitochondrial damage and apoptosis of tumor cells.
In a study of mulberry anthocyanins (cyanidin 3-rutinoside and cyanidin 3-glucoside) on human lung cancer cells, an inhibitory effect on the migration and invasion of highly metastatic A549 cells was revealed. It was concluded that treatment of lung cancer cells with those anthocyanins could decrease the expression of matrix matallopro-tinase-2 (MMP-2) and urokinase-plasminogen activator (u-PA) on the one hand, and on the other increase the expression of tissue inhibitor of matrix matalloprotinase-2 (TIMP-2) and plasminogen activator inhibitor (PAI) [40]. Research with Mexican wild blackberries (R. liebmannii and R. palmeri) indicated that the anthocyanin fraction of extracts of these fruits mostly contained cyanidin 3-rutinoside and cyanidin 3-glucoside and it was determined that anthocyanins elicit apoptosis in C6 cell line and RG2 cell line. Both of these samples caused the arrest of the C6 cell lines in the G0/G1 phase (around 76.5% for R. liebmannii and 75.5% for R. palmeri), significantly higher compared to the control group [11]. Both of these anthocyanins were identified in our samples of wild blackberry and raspberry juices, with wild blackberry having these anthocyanins as dominant ones. Different berry ex-tracts (blackberry, black raspberry, red raspberry, blueberry, cranberry and strawberry) have been tested for the inhibition of growth of tumor cell lines like breast (MCF-7), colon (HT-29 and HCT116), prostate (LNCaP) and oral (KB and CAL27) cell lines. It was concluded that with the increase in berry extract concentration, the resulting inhibition of cell proliferation increased in all of the tested cells, with different degrees of potency between cell lines depending on the anthocyanin profiles of used berry extracts. A comparison of the effect of blackberry and raspberry extracts revealed that the blackberry extracts had a higher potency in the inhibition of tested cell lines than the raspberry extracts; however, their apoptosis effect was the same [41]. Comparison of the effects of different types of extracts (hexane, EtOAc and MeOH extracts) of blackberry and raspberry species (Jamaica-grown species: Rubus jamaicensis, Rubus rosifolius and Rubus racemosus, and of the Michigan-grown Rubus acuminatus, Rubus idaeus cv. Heritage and Rubus idaeus cv. Golden) resulted in the conclusion that their tumor cell proliferation inhibition can be attributed to anthocyanins, and the majority of species contained high amounts of cyanidin-3-glucoside. The most potent sample tested was the hexane extract of Rubus jamaicensis. It had the greatest overall capacity to inhibit the progression of tumor cell growth, inhibiting colon, breast, lung, and gastric human tumor cells by 50%, 24%, 54%, and 37%, respectively, [42].
We determined a dose-dependent behavior in the inhibition of tumor cells with the increase in applied berry juice amounts. That trend was also observed in previous studies where anthocyanins or phenolics showed a dose-dependent growth inhibition against breast, colon, stomach, central nervous system, and lung tumor cells [7,16,38,39,43,44,45].
A major mechanism of suppression of cancer is apoptosis or programmed cell death [41,46]. Generally, the growth rate of preneoplastic or neoplastic cells is higher than the growth of normal cells; thus, the initiation of apoptosis or cell cycle arrest can be a valuable mechanism for inhibition of the promotion and progression of carcinogenesis and consequently for the removal of genetically damaged, preinitiated, or neoplastic cells from the body [41,46]. Among berry phenolics, anthocyanins have been shown to possess apoptotic effects in human cancer cells [41,47,48]. In addition to anthocyanins, quercetin was also recognized for its apoptotic effects [46]. It can cause activation of caspases which are underexpressed in tumor cells leading to the apoptotic response [49].
Dwarf elderberry and wild blackberry juices contained derivatives of quercetin which could lead to induction of apoptosis in combination with anthocyanins. It has been demonstrated that quercetin and fruit extracts (strawberry and plum), in contrast to chlorogenic acid and (-)-epicatechin, caused the induction of apoptosis in HepG2 cells. Actually, quercetin and fruit extracts restrained the G1 phase in the progression of the cell cycle prior to apoptosis so they can contribute to the reduced cell viability in investigated tumor cells [47]. The anti-proliferative effect of quercetin on two breast cancer cell lines (MCF-7 and MDA-MB-231; cells that differ in hormone receptor) was also studied and it was concluded that quercetin had a significant cytotoxicity in MCF-7 cells, but not in MDA-MB-231 cells. In MCF-7 cells, quercetin also had an effect on the restriction of the G1 phase and caused effective suppression of the expression of CyclinD1, p21, Twist and phospho p38MAPK, which was not observed in the MDA-MB-231 cells [50]. Treatment of U87 and T98G cell lines with quercetin showed a significant decrease in the IL-6 mediated STAT3 activation [44]. Additionally, it caused an increase in the sensitivity of U87 and U251 cell lines to TMZ through suppression of Hsp27 known to confer drug resistance [51]. It was observed that quercetin can cause induction of mitochondria-mediated apoptosis in the resistant p53 mutant U373MG cell line [52]
Only rutin was detected in raspberry juice in significantly lower concentrations in comparison to the other two juices. The raspberry juice had a higher impact on inhibition of GBM43 glioblastoma cells than the dwarf elderberry, probably due to the already mentioned cyanidin 3-rutinoside and cyanidin 3-glucoside. Also, raspberry juice and wild blackberry juice contained ellagic acid, which is known for its anticarcinogenic properties. Both in vitro and in vivo studies have revealed that anticarcinogenic effects of ellagic acid were due to the inhibition of tumor cell proliferation, induction of apoptosis, breaking DNA binding to carcinogens, blocking virus infection, and disturbing inflammation, angiogenesis, and drug-resistance processes required for tumor growth and metastasis [53].
Asl et al. [45] also demonstrated the potential cytotoxic activities of ethyl-acetate and methanol extracts of leaf and the fruit of dwarf elderberry upon treatment of breast and stomach cancer cells (MCF-7 and AGS cells) and emphasized the importance of phenolic profile.
Ingestion of berry flavonoids, including anthocyanins, causes their degradation across the digestive system; thus, in circulation, they largely appear in the form of metabolites [1,54]. In plasma, anthocyanins can be found in their intact form, or as the corresponding phenolic acids and aldehydes, and conjugates (methyl, sulfate and glucuronyl conjugates) [54,55]. However, it was determined that flavonoids (including anthocyanins) and their metabolites can be found in the brain tissue passing through the blood–brain barrier [55,56,57], which is one of the challenges in effective glioblastoma treatment. Therefore, our results could be beneficial from this point of view. Additionally, since it has been determined that flavonoids can pass through the blood–brain barrier, our results can be beneficial from the aspect of formulation of delivery systems. Generally, drug delivery to glioblastoma cells is quite a challenging field. Significant efforts have been committed to the development of efficient delivery systems with the aim of overcoming the molecular and cellular heterogeneity of the tumor cells, its infiltrative nature, and the blood–brain barrier [17,18,19]. Since anthocyanins can pass the blood–brain barrier, they have the potential to be incorporated into delivery systems alone or, with TMZ, to boost their effect, which could be further explored.
In conclusion, our study highlights the diverse phenolic composition and antioxidant capacity of three berry juices—wild blackberry, dwarf elderberry, and raspberry—in their potential treatment of glioblastoma tumor cells (U87-MG and GBM43). The distinct anthocyanins, flavonoids, and phenolic acids present in each juice contribute to their varying inhibitory effects on glioblastoma cells. Wild blackberry juice, with its high concentration of anthocyanins, exhibited the most potent inhibition, aligning with findings from other studies.
Anthocyanins, particularly cyanidin types, emerged as the key compounds responsible for the inhibition of glioblastoma cells. The dose-dependent behavior observed in our study is consistent with previous research demonstrating the growth-inhibitory effects of anthocyanins on various tumor cell lines. Apoptosis, a major mechanism of cancer suppression, is induced by anthocyanins and other phenolic compounds, including quercetin. The combination of quercetin derivatives and anthocyanins in dwarf elderberry and wild blackberry juices suggests a potential synergistic effect in inducing apoptosis and cell cycle arrest.
Future studies should explore deeper into the specific mechanisms through which these berry juices exert their anticancer effects, investigating the interplay of individual compounds and potential synergies. Additionally, investigating the impact of different berry varieties and their specific phenolic profiles on various cancer types can provide valuable insights for the development of targeted therapies. In the realm of cancer research, the exploration of natural compounds, such as those found in berry juices, continues to offer promising venues for developing effective and potentially less toxic treatments for glioblastoma and other cancers.
5. Conclusions
Our results suggest that all three berry juices tested (wild blackberry, dwarf elderberry, and raspberry) have anticancer effects on both GBM43 and U87-MG glioblastoma cells. This is significant because GBM43 cells are resistant to temozolomide, the current drug of choice against glioblastomas. The antiproliferative effects of wild blackberry juice, specifically, against GBM43 cells are particularly interesting and warrant further investigation into its mechanism. Dwarf elderberry and raspberry also appear to significantly inhibit U87 and GBM43 cell growth, which warrants further study into the possible impacts of these juices as well. Overall, our study provides strong evidence that wild blackberry, dwarf elderberry, and raspberry juices have antiproliferative effects on glioblastoma cells. Further research is needed to investigate the mechanisms of action of these juices and to evaluate their potential as therapeutic agents for glioblastoma.
Author Contributions
Conceptualization, M.K., D.R., M.A.L. and J.Š.; methodology, M.K. and D.R.; formal analysis, M.K. and D.R.; investigation, M.K., D.R., M.A.L. and J.Š.; data curation, M.K. and D.R.; writing—original draft preparation, M.K. and D.R.; writing—review and editing, M.A.L. and J.Š.; funding acquisition, M.K., D.R., M.A.L. and J.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Croatian Science Foundation; grant number IP-2019-04-5749.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Haizhou, W.; Oliveira, G.; Lila, M.A. Protein-binding approaches for improving bioaccessibility and bioavailability of anthocyanins. Compr. Rev. Food Sci. Food Saf. 2023, 22, 333–354. [Google Scholar]
- Shen, Y.; Zhang, N.; Tian, J.; Xin, G.; Liu, L.; Sun, X.; Li, B. Advanced approaches for improving bioavailability and controlled release of anthocyanins. J. Control Release 2022, 341, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A.A.; Sarmast, E.; Fatehi, P.; Jafari, T. Impact of dietary anthocyanins on systemic and vascular inflammation: Systematic review and meta-analysis on randomised clinical trials. Food Chem. Toxicol. 2020, 135, 110922. [Google Scholar] [CrossRef] [PubMed]
- Kimble, R.; Keane, K.M.; Lodge, J.K.; Howatson, G. Dietary intake of anthocyanins and risk of cardiovascular disease: A systematic review and meta-analysis of prospective cohort studies. Crit. Rev. Food Sci. Nutr. 2019, 59, 3032–3043. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Milenkovic, D. Anthocyanins: From sources and bioavailability to cardiovascular-health benefits and molecular mechanisms of action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling anthocyanin bioavailability for human health. Ann. Rev. Food Sci. Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef]
- Tang, J.; Oroudjev, E.; Wilson, L.; Ayoub, G. Delphinidin and cyanidin exhibit antiproliferative and apoptotic effects in MCF7 human breast cancer cells. Integr. Cancer Sci. Therap. 2015, 2, 82–86. [Google Scholar]
- Hou, D.X.; Kai, K.; Li, J.J.; Lin, S.; Terahara, N.; Wakamatsu, M.; Fujii, M.; Young, M.R.; Colburn, N. Anthocyanidins inhibit activator protein 1 activity and cell transformation: Structure–activity relationship and molecular mechanisms. Carcinogenesis 2004, 25, 29–36. [Google Scholar] [CrossRef]
- Zafra-Stone, S.; Yasmin, T.; Bagchi, M.; Chatterjee, A.; Vinson, J.A.; Bagchi, D. Berry anthocyanins as novel antioxidants in human health and disease prevention. Mol. Nutr. Food Res. 2007, 51, 675–683. [Google Scholar] [CrossRef]
- Raucher, D. Tumor targeting peptides: Novel therapeutic strategies in glioblastoma. Curr. Opin. Pharmcol. 2019, 47, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Velázquez, O.A.; Cortés-Rodríguez, M.; Milán-Carrillo, J.; Montes-Ávila, J.; Robles-Bañuelos, B.; Santamaría del Ángel, A.; Cuevas-Rodríguez, E.O.; Rangel-López, E. Anti-oxidant and anti-proliferative effect of anthocyanin enriched fractions from two Mexican wild blackberries (Rubus spp.) on HepG2 and glioma cell lines. J. Berry Res. 2020, 10, 513–529. [Google Scholar] [CrossRef]
- Thakkar, J.P.; Dolecek, T.A.; Horbinski, C.; Ostrom, Q.T.; Lightner, D.D.; Barnholtz-Sloan, J.S.; Villano, J.L. Epidemiologic and molecular prognostic review of glioblastoma. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Rooprai, H.K.; Christidou, M.; Murray, S.A.; Davies, D.; Selway, R.; Gullan, R.W.; Pilkington, G.J. Inhibition of Invasion by Polyphenols from Citrus Fruit and Berries in Human Malignant Glioma Cells In Vitro. Anticancer Res. 2021, 41, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Expósito, M.J.; Martínez-Martos, J.M. The Delicate Equilibrium between Oxidants and Antioxidants in Brain Glioma. Curr. Neuropharmacol. 2019, 17, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Lamy, S.; Lafleur, S.; Bedard, V.; Moghrabi, A.; Barrete, S.; Gingras, D.; Béliveau, R. Anthocyanidins inhibit migration of glioblastoma cells: Structure-activity relationship and involvement of the plasmolytic system. J. Cell. Biochem. 2007, 100, 100–111. [Google Scholar] [CrossRef]
- Dragojevic, S.; Mackey, R.; Raucher, D. Evaluation of Elastin-Like Polypeptides for Tumor Targeted Delivery of Doxorubicin to Glioblastoma. Molecules 2019, 24, 3242. [Google Scholar] [CrossRef]
- Mathew, E.N.; Berry, B.C.; Yang, H.W.; Carroll, R.S.; Johnson, M.D. Delivering Therapeutics to Glioblastoma: Overcoming Biological Constraints. Int. J. Mol. Sci. 2022, 23, 1711. [Google Scholar] [CrossRef]
- Raucher, D.; Dragojevic, S.; Ryu, J. Macromolecular Drug Carriers for Targeted Glioblastoma Therapy: Preclinical Studies, Challenges, and Future Perspectives. Front. Oncol. 2018, 8, 624. [Google Scholar] [CrossRef]
- Vengoji, R.; Macha, M.A.; Batra, S.K.; Shonka, N.A. Natural products: A hope for glioblastoma patients. Oncotarget 2018, 9, 22194–22219. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Bjerke, M.; Edlund, H.; Nelander, S.; Westermark, B. Origin of the U87MG glioma cell line: Good news and bad news. Sci. Transl. Med. 2016, 8, 354re3. [Google Scholar] [CrossRef]
- Pevna, V.; Wagnières, G.; Huntosova, V. Autophagy and Apoptosis Induced in U87 MG Glioblastoma Cells by Hypericin-Mediated Photodynamic Therapy Can Be Photobiomodulated with 808 nm Light. Biomedicines 2021, 9, 1703. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.; Koosha, F.; Neshastehriz, A. Chemo-radiation therapy of U87-MG glioblastoma cells using SPIO@AuNP-Cisplatin-Alginate nanocomplex. Heliyon 2023, 9, e13847. [Google Scholar] [CrossRef]
- Di Cintio, F.; Dal Bo, M.; Baboci, L.; De Mattia, E.; Polano, M.; Toffoli, G. The Molecular and Microenvironmental Landscape of Glioblastomas: Implications for the Novel Treatment Choices. Front. Neurosci. 2020, 14, 603647. [Google Scholar] [CrossRef]
- Nguyen, H.-M.; Guz-Montgomery, K.; Lowe, D.B.; Saha, D. Pathogenetic Features and Current Management of Glioblastoma. Cancers 2021, 13, 856. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Ko, H.-J.; Huang, C.-Y.F.; Lin, C.-Y.; Chiou, S.-J.; Su, Y.-F.; Lieu, A.-S.; Loh, J.-K.; Kwan, A.-L.; Chuang, T.-H.; et al. Ionizing Radiation Induces Resistant Glioblastoma Stem-Like Cells by Promoting Autophagy via the Wnt/β-Catenin Pathway. Life 2021, 11, 451. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotonutric acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. In Current Protocols in Food Analytical Chemistry Current Protocols; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2001. [Google Scholar]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Ozyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Sci. Food Agric. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Pambianchi, E.; Hagenberg, Z.; Pecorelli, A.; Grace, M.; Therrien, J.-P.; Lila, M.A.; Valacchi, G. Alaskan bog blueberry (Vaccinium uliginosum) extract as an innovative topical approach to prevent uv-induced skin damage. Cosmetics 2021, 8, 112. [Google Scholar] [CrossRef]
- Može, Š.; Polak, T.; Gašperlin, L.; Koron, D.; Vanzo, A.; Poklar Ulrih, N.; Abram, V. Phenolics in Slovenian Bilberries (Vaccinium myrtillus L.) and Blueberries (Vaccinium corymbosum L.). J. Agric. Food Chem. 2011, 59, 6998–7004. [Google Scholar] [CrossRef] [PubMed]
- Buljeta, I.; Pichler, A.; Šimunović, J.; Kopjar, M. Polyphenols and antioxidant activity of citrus fiber/blackberry juice complexes. Molecules 2021, 26, 4400. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular proliferation and viability: Application to the proliferation and viability of murine macrophages. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Kamei, H.; Kojima, T.; Hasegawa, M.; Koide, T.; Umeda, T.; Yukawa, T.; Terabe, K. Suppression of tumor cell growth by anthocyanins in vitro. Cancer Invest. 1995, 13, 590–594. [Google Scholar] [CrossRef]
- Jing, N.; Song, J.; Liu, Z.; Wang, L.; Jiang, G. Glycosylation of anthocyanins enhances the apoptosis of colon cancer cells by handicapping energy metabolism. BMC Complement. Med. Ther. 2020, 20, 312. [Google Scholar] [CrossRef]
- Chen, P.-N.; Chu, S.-C.; Chiou, H.-L.; Kuo, W.-H.; Chiang, C.-L.; Hsieh, Y.-S. Mulberry anthocyanins, cyanidin 3-rutinoside and cyanidin 3-glucoside, exhibited an inhibitory effect on the migration and invasion of a human lung cancer cell line. Cancer Lett. 2006, 235, 248–259. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef]
- Bowen-Forbes, C.S.; Zhang, Y.; Nair, M.G. Anthocyanin content, antioxidant, anti-inflammatory and anticancer properties of blackberry and raspberry fruits. J. Food Compos. Anal. 2010, 23, 554–560. [Google Scholar] [CrossRef]
- Reddy, M.K.; Alexander-Lindo, R.L.; Nair, M.G. Relative inhibition of lipid peroxidation, cyclooxygenase enzymes, and human tumor cell proliferation by natural food colors. J. Agric. Food Chem. 2005, 53, 9268–9273. [Google Scholar] [CrossRef] [PubMed]
- Michaud-Levesque, J.; Bousquet-Gagnon, N.; Beliveau, R. Quercetin abrogates IL-6/STAT3 signaling and inhibits glioblastoma cell line growth and migration. Exp. Cell Res. 2012, 318, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Asl, F.R.; Barzegar, A.; Ebrahimzadeh, M.A. Evaluating Cytotoxic Potential of the Fruit and the Leaf Extracts of Sambucus ebulus (L.) on MCF7 and AGS Cell Lines. Res. Mol. Med. 2021, 9, 11–20. [Google Scholar]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Ramos, S.; Alia, M.; Bravo, L.; Goya, L. Comparative effects of food-derived polyphenols on the viability and apoptosis of a human hepatoma cell line (HepG2). J. Agric. Food Chem. 2005, 53, 1271–1280. [Google Scholar] [CrossRef]
- Heo, H.J.; Lee, C.Y. Strawberry and its anthocyanins reduce oxidative stress-induced apoptosis in PC12 cells. J. Agric. Food Chem. 2005, 53, 1984–1989. [Google Scholar] [CrossRef]
- Schnekenburger, M.; Dicato, M.; Diederich, M. Plant-derived epigenetic modulators for cancer treatment and prevention. Biotechnol. Adv. 2014, 32, 1123–1132. [Google Scholar] [CrossRef]
- Ranganathan, S.; Halagowder, D.; Sivasithambaram, N.D. Quercetin suppresses twist to induce apoptosis in MCF-7 breast cancer cells. PLoS ONE 2015, 10, e0141370. [Google Scholar] [CrossRef]
- Sang, D.P.; Li, R.J.; Lan, Q. Quercetin sensitizes human glioblastoma cells to temozolomide in vitro via inhibition of Hsp27. Acta Pharmacol. Sin. 2014, 35, 832–938. [Google Scholar] [CrossRef]
- Kim, H.; Moon, J.Y.; Ahn, K.S.; Cho, S.K. Quercetin induces mitochondrial mediated apoptosis and protective autophagy in human glioblastoma U373MG cells. Oxid. Med. Cell. Longev. 2013, 2013, 596496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-M.; Zhao, L.; Li, H.; Xu, H.; Chen, W.-W.; Tao, L. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Biol. Med. 2014, 11, 92–100. [Google Scholar] [PubMed]
- Hribar, U.; Poklar Ulrih, N. The Metabolism of Anthocyanins. Curr. Drug Metab. 2014, 15, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Janle, E.M.; Lila, M.A.; Grannan, M.; Wood, L.; Higgins, A.; Yousef, G.G.; Rogers, R.B.; Kim, H.; Jackson, G.S.; Weaver, C. Method for evaluating the potential of 14C labeled plant polyphenols to cross the blood-brain barrier using accelerator mass spectrometry. Nucl. Instrum. Methods Phys. Res. 2010, 268, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, R.; Anada, M.; Miyaguchi, A.; Nomi, Y.; Matsumoto, H. Evaluation of Blood-Brain Barrier Permeability of Polyphenols, Anthocyanins, and Their Metabolites. J. Agric. Food Chem. 2021, 69, 11676–11686. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Dobbie, M.S.; Kuhnle, G.; Proteggente, A.R.; Abbott, N.J.; Rice-Evans, C. Interaction between flavonoids and the blood–brain barrier: In vitro studies. J. Neurochem. 2003, 85, 180–192. [Google Scholar] [CrossRef]
Figure 1.
Antiproliferative effect of investigated berry juices on GBM43 and U87-MG cells. (p values * < 0.05, ** < 0.01, *** < 0.001, **** < 0.0001); DE—dwarf elderberry; WB—wild blackberry; RB—raspberry; UT—control; TMZ—Temozolomide.
Figure 1.
Antiproliferative effect of investigated berry juices on GBM43 and U87-MG cells. (p values * < 0.05, ** < 0.01, *** < 0.001, **** < 0.0001); DE—dwarf elderberry; WB—wild blackberry; RB—raspberry; UT—control; TMZ—Temozolomide.


{kind=link}
{kind=link}
Table 1.
Total polyphenols (TP), monomeric anthocyanins (MA) and proanthocyanidins (PAC) of investigated berry juices.
Table 1.
Total polyphenols (TP), monomeric anthocyanins (MA) and proanthocyanidins (PAC) of investigated berry juices.
| Juice | TP (g/L) | MA (mg/L) | PAC (mg/L) |
|---|---|---|---|
| DE | 3.16 ± 0.02 a | 433.17 ± 2.66 b | 71.21 ± 0.81 a |
| WB | 3.21 ± 0.01 a | 759.99 ± 3.53 a | 56.21 ± 1.96 b |
| RB | 1.67 ± 0.02 b | 230.39 ± 1.10 c | 32.34 ± 0.27 c |
DE—dwarf elderberry; WB—wild blackberry; RB—raspberry. Values in the same column, marked with different letters (a–c) are significantly different at p ≤ 0.05.
Table 2.
Polyphenols amount of investigated berry juices (mg/L of fresh weight).
| DE | WB | RB | |
|---|---|---|---|
| Cyanidin-3-glucoside | nd | 389.60 ± 10.98 a | 39.11 ± 0.01 b |
| Cyanidin-3-rutinoside | nd | 269.72 ± 8.52 a | 34.28 ± 0.35 b |
| Cyanidin-3-galactoside | 261.40 ± 1.41 a | nd | nd |
| Cyanidin-3-sophoroside | nd | nd | 210.65 ± 0.23 a |
| Hyperoside | 34.19 ± 0.09 b | 73.27 ± 2.39 a | nd |
| Rutin | 109.72 ± 0.05 a | 34.37 ± 0.96 b | 14.70 ± 0.41 bc |
| Gallic acid | 4.58 ± 0.04 a | 4.56 ± 0.03 a | nd |
| Ellagic acid | nd | 5.14 ± 0.03 a | 3.41 ± 0.00 b |
| Chlorogenic acid | 160.02 ± 2.87 a | nd | nd |
| Neochlorogenic acid | 27.18 ± 1.67 a | nd | nd |
DE—dwarf elderberry; WB—wild blackberry; RB—raspberry; nd—not detected. Values in the same row, marked with different letters (a–c) are significantly different at p ≤ 0.05.
Table 3.
Antioxidant activity (µmol TE/100 mL of fresh weight) of investigated berry juices.
| Juice | DPPH | ABTS | FRAP | CUPRAC |
|---|---|---|---|---|
| DE | 9.86 ± 0.01 b | 26.18 ± 0.57 a | 2.11 ± 0.01 a | 107.78 ± 0.56 a |
| WB | 13.25 ± 0.06 a | 6.08 ± 0.01 c | 0.59 ± 0.01 c | 27.37 ± 0.37 c |
| RB | 6.95 ± 0.03 c | 12.59 ± 0.12 b | 1.38 ± 0.00 b | 64.00 ± 0.11 b |
DE—dwarf elderberry; WB—wild blackberry; RB—raspberry; DPPH (2,2-diphenyl-1-picrylhydrazyl radicals assay); ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt cation radicals assay); FRAP (ferric reducing antioxidant power assay); CUPRAC (cupric ion reducing capability assay). Values in the same column, marked with different letters (a–c) are significantly different at p ≤ 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kopjar, M.; Raucher, D.; Lila, M.A.; Šimunović, J. Anti-Glioblastoma Potential and Phenolic Profile of Berry Juices. Processes 2024, 12, 242. https://doi.org/10.3390/pr12020242
AMA Style
Kopjar M, Raucher D, Lila MA, Šimunović J. Anti-Glioblastoma Potential and Phenolic Profile of Berry Juices. Processes. 2024; 12(2):242. https://doi.org/10.3390/pr12020242
Chicago/Turabian StyleKopjar, Mirela, Drazen Raucher, Mary Ann Lila, and Josip Šimunović. 2024. "Anti-Glioblastoma Potential and Phenolic Profile of Berry Juices" Processes 12, no. 2: 242. https://doi.org/10.3390/pr12020242
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.