
Progress on the Effect of Nitrogen on Transformation of Soil Organic Carbon
by
Danyang Meng
1,2, 
Hangxin Cheng
3,
Yang Shao
1,
Min Luo
1,
Diandou Xu
1,
Zhiming Liu
2,* and
Lingling Ma
1,* 1
Beijing Ray Imaging Technology and Equipment Engineering Technology Research Center, Institute of High Energy Physics, Chinese Academy of Sciences, Beijing 100049, China
2
State Key Laboratory of Chemical Resource Engineering, Beijing University of Chemical Technology, Beijing 100029, China
3
Institute of Geophysical and Geochemical Exploration, Chinese Academy of Geological Sciences, Langfang 065000, China
*
Authors to whom correspondence should be addressed.
Processes 2022, 10(11), 2425; https://doi.org/10.3390/pr10112425
Submission received: 17 October 2022
/
Revised: 8 November 2022
/
Accepted: 15 November 2022
/
Published: 16 November 2022
(This article belongs to the Special Issue Trends in Carbon Capture, Storage and Utilisation)
Abstract
:Carbon and nitrogen are the essential elements constituting living organisms and are closely coupled during biogeochemical cycles. Due to the atmospheric nitrogen deposition and increased agricultural nitrogen fertilizer input, the effect of nitrogen on the sequestration of soil organic carbon (SOC) is controversial. To facilitate a comprehensive understanding of this issue, the progress of recent studies on the different SOC stabilization mechanisms is reviewed. Based on the differences in the stability and fate mechanisms of particulate organic carbon (POC) and mineral-associated organic carbon (MAOC), nitrogen input can increase POC input and inhibit microbial decomposition of POC by increasing terrestrial biomass, changing the quality of litter and promoting the formation of aggregates. N input reduces the chemical stability of MAOC by altering the chemical bonding of mineral–organic complexes. This study has promising implications for understanding the effect of N on SOC transformation by different stabilization mechanisms to promote soil carbon sequestration.
1. Introduction
Soil organic carbon (SOC) is a significant indicator of soil fertility, and its sequestration is a vital strategy to mitigate greenhouse gases. The soil carbon pool is the largest in terrestrial ecosystems, accounting for 81.2% of the carbon in the ecosystem and containing about 250 billion tons of soil organic carbon [1]. For the global soil carbon cycle, even a minor disturbance will cause a great response to the climate [2]. In addition, organic carbon also has a variety of functions. For example, chemically organic carbon is a store of soil nutrients; physically, it improves soil aggregation and water-holding capacity (WHC), and biologically it increases the energy supply of microorganisms [3].
The fixed potential of soil organic carbon is related to the quantity and quality of input organic matter, soil microbial activity, and physical and chemical properties [4,5,6]. Soil carbon and nitrogen are closely coupled, and nitrogen is involved in the biogeochemical cycle of carbon as an energy supply element for microbial and plant growth [7]. Since the 1960s, nitrogen fertilizer use has increased dramatically [8]. Nitrogen input can gradually affect soil organic carbon stocks by altering the importation of fresh particulate organic carbon to the soil [9] and the output of microbially decomposed organic carbon [10]. Previous studies have reported that organic carbon sequestration under N addition may change in three different directions of change: increasing, constant, and decreasing [11,12,13].
Nitrogen bioavailability is a critical factor affecting organic carbon cycling and sequestration [14]. For example, plant litter with low C/N has a faster decomposition rate than high C/N [15]. The rate of SOM decomposition is positively related to soil nitrogen effectiveness; therefore, increasing nitrogen effectiveness accelerates SOM decomposition and is detrimental to SOC sequestration. However, nitrogen increases SOC input by promoting plant growth and plant litter formation [16]. At the same time, nitrogen input also inhibits the decomposition of SOC by reducing microbial activity and promoting the formation of aggregates [17]. The main effect of inorganic nitrogen input on soil’s physical and chemical properties is to cause soil acidification. The conversion of ammonium nitrogen applied to agricultural soils to nitrate nitrogen through nitrification, the uptake of ammonium nitrogen by plants, and the loss of nitrogen by leaching all lead to soil acidification [18,19], by the following reactions [20]:
Previous studies have suggested that a decrease in PH under N addition drives Al toxicity [21], reducing fungal abundance [17,22] and inhibiting microbial catabolic activity. Chang [23] et al. conducted a two-year ammonium nitrate application experiment on alpine ecosystems. They found that MWD values of soil aggregates were significantly higher, MWD positively correlated with SOC content, and MWD negatively correlated with CO2 release. Thus, increased nitrogen effectiveness can enhance soil organic carbon input and slow down soil organic carbon decomposition, promoting soil SOC sequestration. Overall, the effect of N input on soil organic carbon turnover is attributed to the interaction between plants, soil physicochemical properties, and microorganisms.
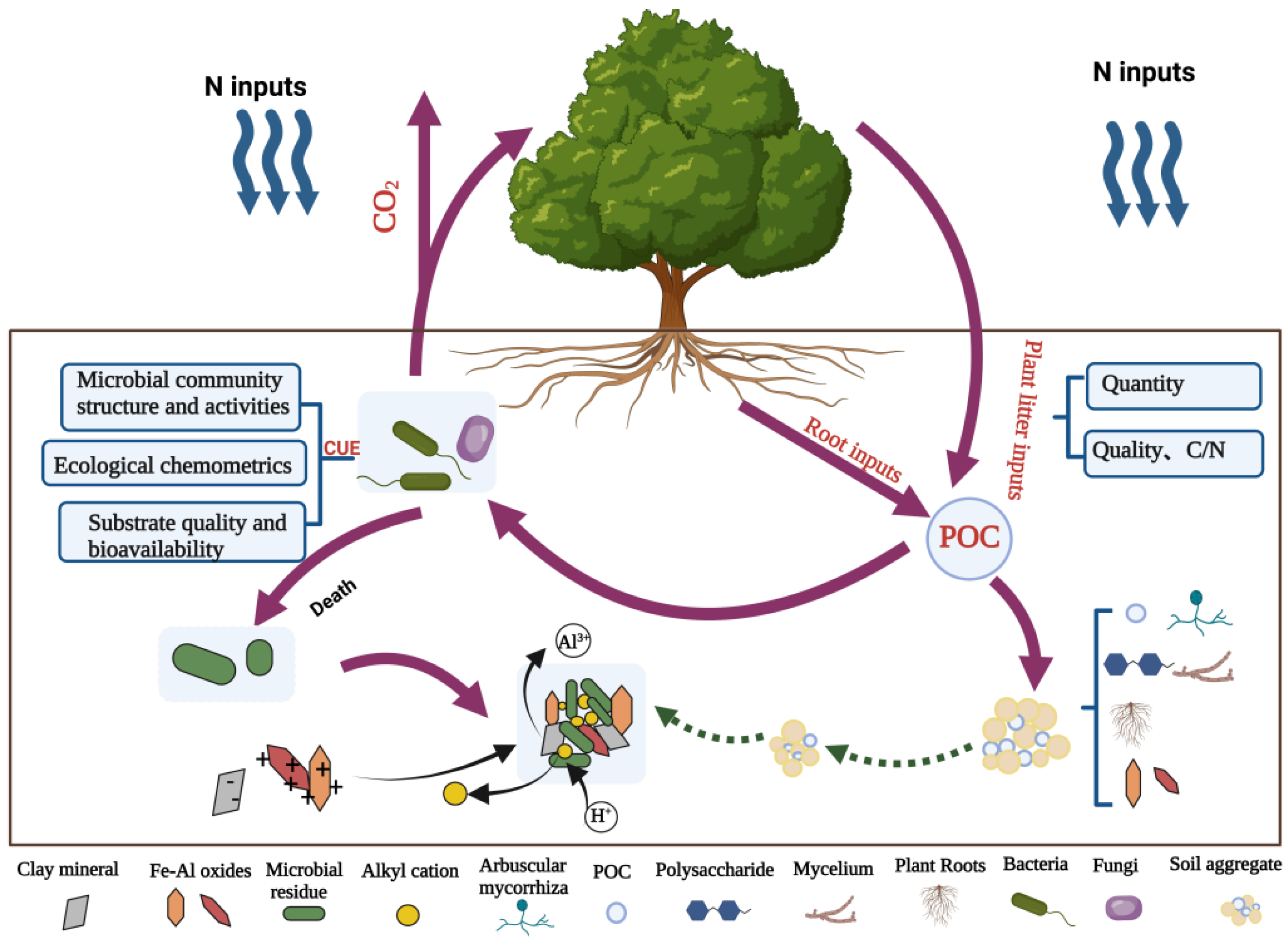
Previous studies focused on microorganisms’ organic carbon cycle [23,24,25]. In comparison, there were few studies on the transformation and fixation of organic carbon of different components in soil under nitrogen input. The division of organic carbon into particulate organic carbon (POC) and mineral-associated organic carbon (MAOC) is fundamental to understanding organic carbon turnover processes. Numerous previous studies have shown that the MAOC fraction, mainly derived from microbial residues [26,27], has a slow turnover, is difficult to decompose, requires large amounts of nitrogen for its formation, and has a limited storage capacity. In contrast, the POC fraction, an essential precursor to MAOC, is mainly derived from plant and animal residues and root secretions [28], is susceptible to external disturbances, has poor stability and rapid turnover rates [29,30], has a low demand for nitrogen, and may accumulate indefinitely. Given that the two carbon pools have different turnover rates and stability, there are also different conversion directions and stabilization mechanisms for the effects of nitrogen addition (Table 1, Figure 1). Nitrogen addition promotes aboveground biomass in nitrogen-limited terrestrial ecosystems [31], but there is significant uncertainty about belowground organic carbon cycling and fixation. Therefore, this paper reviews the research progress in recent years on how nitrogen input changes the transformation of organic carbon produced by POC and MAOC to comprehensively understand the impact of nitrogen on the occurrence and transformation of organic carbon in four aspects: the quantity and quality of plant litter, the physical protection of aggregates, microbial carbon utilization efficiency (CUE), and the formation of organic–mineral complexes.
2. Particulate Organic Carbon (POC)
Particulate organic carbon (POC) is usually composed of undissolved or partially decomposed animal, plant, and root residues [26]. It refers to the part of organic carbon combined with soil sand, which has a fast turnover rate and is vulnerable to external interference. It is a component and quantitative indicator of soil active organic matter [32]. The mineralization of POC is mainly restricted by the chemical resistance of imported soil organic molecules and the physical protection of large aggregates.
2.1. Quantity and Quality of Plant Litter
Nitrogen addition reduces competition for bioavailable N between microorganisms and plants, increases net primary plant productivity, and promotes carbon input from soil root secretions and plant residues [33]. Global net primary production of vegetation (NPP) is limited by N, with NPP increasing by up to 30% under N deposition [34]. Although this implies a higher POC input, there are differences in the direction of SOC sequestration in soils. Previous studies have shown that the net rate of soil organic carbon fixation is positively correlated with root mass. Fornara et al. [35] found that a potential mechanism for SOC accumulation in grassland soils is an N-induced increase in root mass through long-term NH4NO3 addition experiments on grassland soils. However, Lajtha et al. [36] did not find a significant effect of the amount of annual litter on SOC in a model formulation of soil carbon based on first-order decomposition kinetics, most of which may be lost through leaching and microbial respiration. Root secretions facilitate the co-metabolic process of decomposition of the original organic carbon of the soil by providing a readily bioavailable energy supply [37]. Soil acidification from excess N inputs can also lead to increased levels of toxic elements such as Al and Mn [38]. In acidic environments, heavy metal ions are more likely to bind to water in an ionic state. The more acidic the environment, the higher the ionic solubility and the easier it is for plants to take up or migrate, leading to lower/net primary productivity and reduced POC input.
Reasonable input of N increases the net primary productivity of vegetation, but the direction of change in SOC stocks may depend on changes in litter mass and C/N [39]. Nitrogen addition may lead to elevated levels of structurally complex organic matter such as lignin, which favors POC formation by resisting microbial degradation but is detrimental to the long-term sequestration of soil SOC [40]. It has also been found that N addition had no significant effect on the ratio of lignin, glucose, and cellulose in plant tissues and significantly contributed to SOC accumulation mainly by reducing C/N in foliage and roots [35,41]. The results of several studies have shown that low-C/N, easily decomposable litter, where microorganisms are more likely to obtain high-quality inputs and improve litter turnover, is not conducive to POC sequestration but can effectively form MAOM, which is beneficial to SOC accumulation in the long run [42,43,44]. In contrast, high C/N recalcitrant litter resists microbial decomposition in the short term and contributes to the accumulation of short-term carbon pool POC, but less of the initial material is converted to microbial products that can be stably present. [45]. Nitrogen addition can also influence the conversion of SOC in soils by altering plant community composition [35]. High N addition drives the conversion of terrestrial vegetation from C4 to C3 plants, which have lower C/N and higher root mass than C4 plants [46], contributing to a more efficient transformation of POC to MAOC.
2.2. Physical Protection of Aggregates
Soil aggregates are the basic structural unit of soil and the leading site of organic carbon fixation. About 90% of organic carbon in the surface soil of terrestrial ecosystems is located in aggregates [47]. The spatial segregation of microorganisms and organic carbon generated during the formation of aggregates is the primary stabilization mechanism of soil organic carbon [48]. Soil aggregates’ stability was significantly and positively correlated with SOC content [49], especially the POC fraction [50,51]. Chen et al. [52] found that POC could explain 87.1% of the variation in MWD and macroaggregates by redundancy analysis (RDA) between soil aggregates and basic physicochemical properties in saline-alkali soil, indicating that POC was a suitable index for evaluating the formation of macroaggregates in saline-alkali soil. The relationship between POC and soil aggregates is interactive: on the one hand, POC promotes the formation of microaggregates and macroaggregates; on the other hand, the highly complex spatial arrangement and composition of solid particles make the soil structure more inhomogeneous, affecting the distribution and biological effectiveness of POC and creating a physical protection effect on POC.
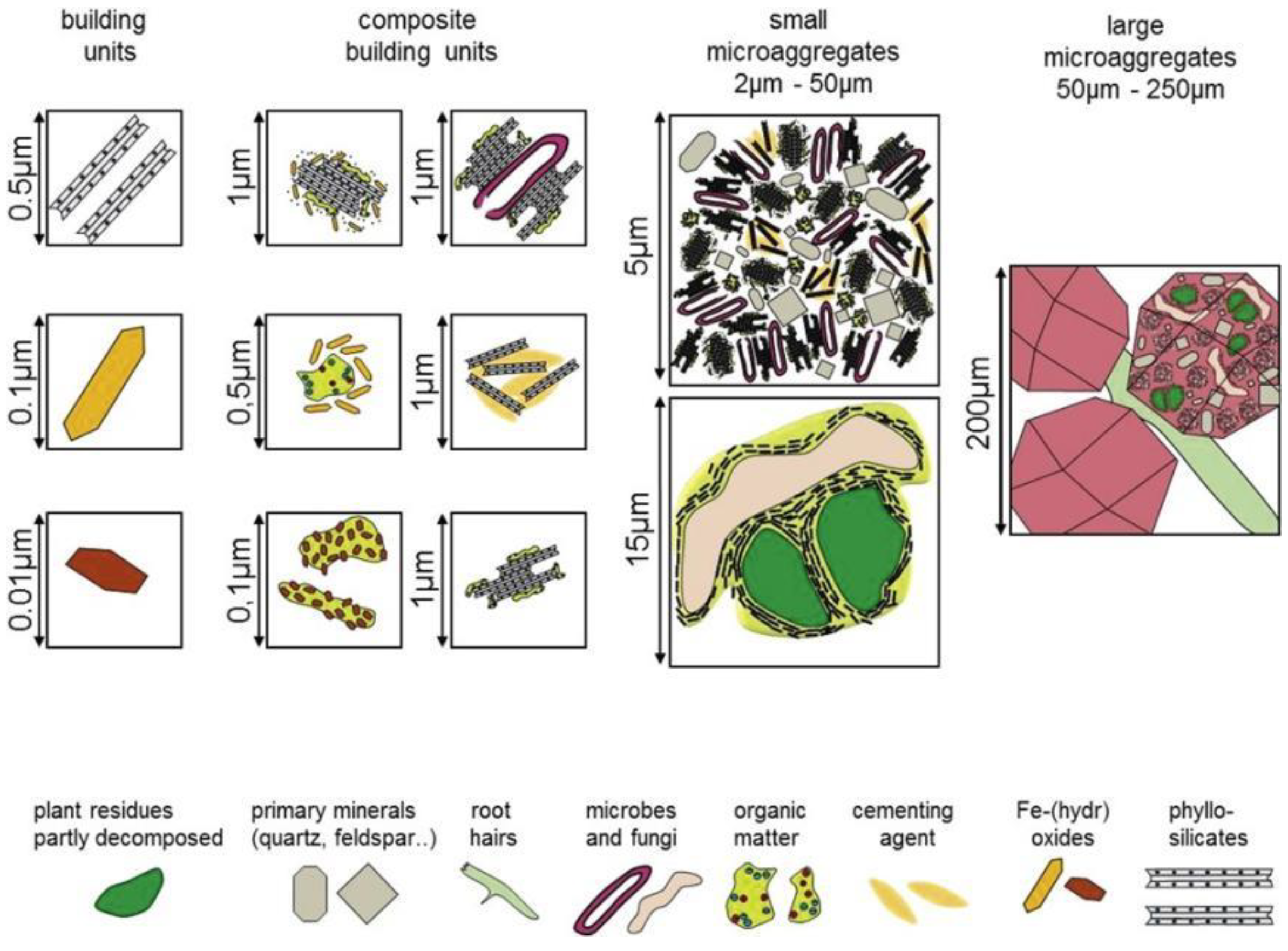
N deposition can help stabilize soil structure and reduce POC decomposition by promoting aggregate formation and increasing MWD [23,53], as shown in Figure 2 [54]. According to the soil aggregates hierarchical development model [55], the formation of aggregates is influenced by various types of caking agents. Primarily, persistent cementing substances (highly humified organic matter, multivalent metal cations, and adsorbed complexes) and powder-mucilage particles polymerize to form microaggregates. Then, the microaggregates combine to form macroaggregates through temporary (fungal mycelium and roots) and transient (polysaccharides secreted by microorganisms and roots) cementing substances [56], as shown in Figure 3 [57].
On the one hand, nitrogen deposition can affect the formation of an organic cementing agent. Nitrogen addition increases microbial activity, promotes the mycelial growth of fungi [58], and releases polysaccharides, lipids, mucus, etc., from microbial secretions [59]. Fungus mycelium will provide resistance against mechanical breakdown and exfoliation. The linear molecular structure of polysaccharides and the formation of contacts, acid groups, and hydroxy–hydrogen bonds in the molecules promote the formation of soil aggregates. The application of organic nitrogen fertilizer activates clumped mycorrhizae [60], enhances the production of insoluble glycoproteins [61], and facilitates the production of aggregates. N enrichment significantly contributed to aboveground biomass, but there was a threshold effect on belowground plant roots [62]: belowground biomass increased under low-dose N additions and slightly decreased under high-dose N additions. Thus, appropriate N addition promotes the entanglement of plant roots on aggregate formation [56], inhibits microbial respiration, and protects the POC produced by plant litter to some extent. Organic N promotes soil macroaggregate formation better than inorganic N. This is because organic N alleviates soil acidification [63,64] and significantly increases aboveground biomass, leading to more plant residues entering the soil and forming POC; in addition, organic N deposition can be considered as increasing unstable organic matter in the soil [65], acting as an organic cementing agent for aggregate formation. On the other hand, nitrogen addition affects the cementation of soil inorganic cementing agents by changing the soil PH. Soil inorganic cementing agents include oxides and carbonates. Fe-Al oxides are amphoteric, showing a positive charge at low pH and a negative charge at high pH [66]. Soil acidification due to nitrogen enrichment results in positively charged Fe-Al oxides, which cover the mucilage surface as a gel film [67] and act as a “bond bridge” in the soil aggregation process. In acidic soils with low organic matter and Fe-Al oxide content, the cementation of Fe-Al oxides plays a key role in the formation of aggregates [68]. Carbonates in semi-arid calcareous soils can contribute significantly to the formation of stable aggregates [69]. The increase in H+ content under nitrogen deposition leads to an extensive depletion of soil inorganic carbon in arid and semi-arid regions and karsts, which affects the cementation of inorganic carbonates [70]. However, it has also been found that carbon accumulation in surface soil aggregates under short-term nitrogen deposition is related to the inhibition of soil respiration under microbial reduction without a direct relationship with soil acidification [23].
3. Mineral-Associated Organic Carbon (MAOC)
Mineral-associated organic carbon (MAOC) is the most stable and longest turnover organic carbon. The sorption of organic matter by soil minerals prolongs the ecosystem’s organic carbon cycle. It reduces its bioavailability [26,71]. Microbial growth rate and microbial residues control the amount of newly formed MAOC, since microbial residues are an essential source of MAOC formation [72,73]. Clay minerals and iron aluminum oxide hydrate in soil minerals play a significant role in organic carbon stabilization [74]. Ligand exchange adsorption of soil minerals on organic matter, multivalent cation bonding bridges, complexation of free Ca2+, Al3+, and Fe3+ and their mineral forms on organic ligands, hydrogen bonding, condensation, and van der Waals forces are the main chemoprotective mechanisms for MAOC persistence [75,76]. Thus, N addition may affect MAOC sequestration through two pathways. First, N addition alters microbial CUE and thus microbial mass carbon by affecting the microbial activity and community structure; second, soil acidification due to N addition triggers changes in the chemical bonding of mineral–organic complexes.
3.1. Microbial Carbon Utilization Efficiency
Microorganisms mediate the transfer from plant inputs to mineral-related organic carbon pools and are the main drivers of the soil carbon cycle [77]. Microbially driven soil C dynamics may be closely related to microbial catabolic and anabolic processes [78]. In the process of decomposing organic matter, one part of organic matter is lost through respiration and extracellular enzymes, and the other part is converted into microbial biomass carbon and microbial biomass nitrogen through assimilation [79]. Liang et al. [72] proposed the concept of a “soil microbial carbon pump,” a theoretical system that defined for the first time the “entombing effect,” i.e., the long-term microbial assimilation process leads to the continuous accumulation of iterations of microbial residues, which promotes the formation of a series of organic substances including microbial residues. Microbial carbon utilization efficiency (CUE) [80] is the efficiency of converting total organic carbon absorbed by microorganisms into microbial biomass carbon. Under thermodynamic constraints, the maximum theoretical value of CUE is 0.8 [81], which indicates that up to 80% of organic carbon can be converted to MBC, and at least 20% of organic carbon is used to sustain microbial respiration and secretion of extracellular enzymes.
Nitrogen availability strongly influences microbial CUE because microorganisms require energy to balance catabolic and anabolic reactions. N input drives changes in microbial CUE by altering soil microbial activity and community structure, substrate quality and bioavailability, and ecological stoichiometry [82,83].
3.1.1. Microbial Community Structure and Activities
Microbial community structure and activity are the strongest drivers of CUE because different microbial communities metabolize organic carbon in different pathways [84]. Fungi exhibit high CUE because they have a more comprehensive range of C/N variability than bacteria and have a higher demand for C [77]. Hawkes et al. [85] concluded that the dominance of fungi in soil implies higher carbon stocks.
Both increased nitrogen effectiveness and changes in soil pH under N addition can induce a rapid shift in microbial communities [86,87], but the two are contradictory. Higher nitrogen effectiveness drives a change in microbial community structure toward bacterial dominance [22,88], which negatively affects CUE because bacteria have a higher demand for soil nutrients than fungi [22]. Soil acidification due to N addition drives a shift in microbial community structure towards fungal dominance. Bacterial/fungal community structure was positively related to soil pH [89] due to different microbial dominant communities for soil organic matter decomposition activities at different pH levels. Bacteria are most active under neutral conditions, fungi grow best under neutral or slightly acidic conditions, and fungi have better resistance to acidification [90]. The input of mineral N, especially nitrate N, leads to soil acidification and shifts the microbial community towards fungal dominance with high CUE. In contrast, organic N fertilization and organic–inorganic pairing will alleviate soil acidification and move the microbial community towards bacterial authority [91].
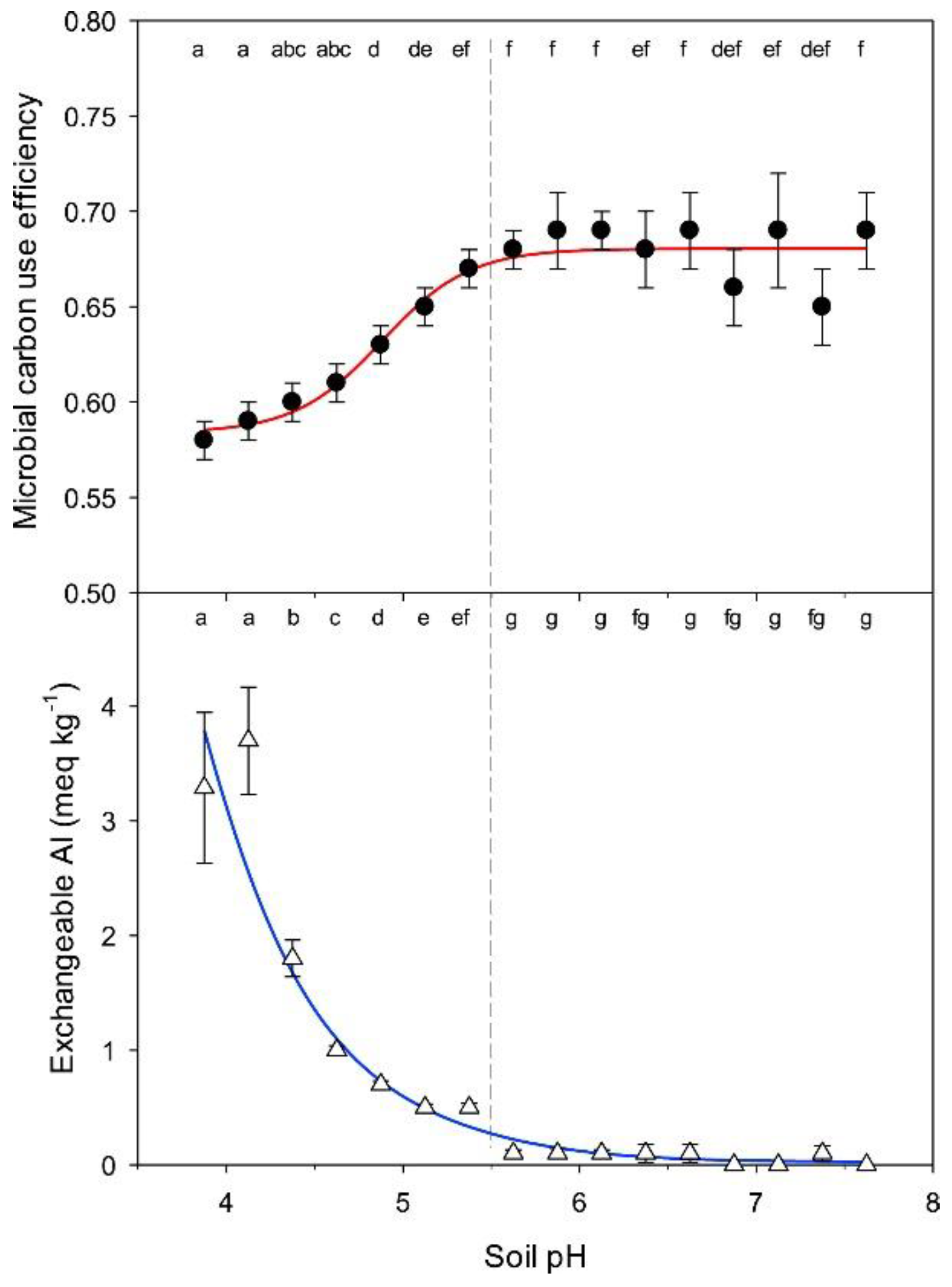
Nitrogen additions can also affect microbial activity by altering soil pH [88]. Soil acidification caused by excessive mineral N input forces microorganisms to allocate large amounts of SOC for energy production [92], which inhibits microbial growth and reduces microbial CUE. Organic N input alleviates soil acidification and enhances microbial CUE [93]. First, the decrease in pH leads to the leaching of Ca2+ and Mg2+ [94], and microorganisms may become Ca and Mg nutrient-limited, directly inhibiting microbial growth and reducing microbial CUE. Second, the decrease in pH and the increase in Al3+ migration lead to elevated microbial heavy metal resistance genes [95]. Microorganisms resist heavy metals through efflux pump mechanisms, enzymatic transport mechanisms, and chelation of heavy metals in vivo [96,97]. This process usually requires ATP, so that more C must be transferred to respiration and microbial CUE reduced. Davey et al. [98] measured microbial CUE using 14C-labeled glucose in 970 soil samples from the southwestern agricultural region of Australia. They found that the relationship between microbial CUE and pH showed a clear turning point at pH = 5.5, with a gradual decrease in CUE as the soil became progressively more acidic, and that at the pH = 5.5 turning point, soil exchange complexes with Al3 + began to dominate in the soil exchange complex (Figure 4). There is a strong negative correlation between microbial CUE and exchangeable Al3+, as microbial resistance to toxic metal elements is energy intensive [99]. Under environmental conditions of low pH and high Al concentration, the metabolic pathways of microbiology shift [100]. First, microorganisms produce and secrete compounds (e.g., extracellular polymers and organic acids) to the external environment to combine and detoxify the harmful Al3+ [96,101]. Second, microorganisms secrete energy to overcome acidic stress and maintain cellular integrity, indirectly inhibiting microbial growth and reducing microbial CUE [102].
3.1.2. Ecological Chemometrics
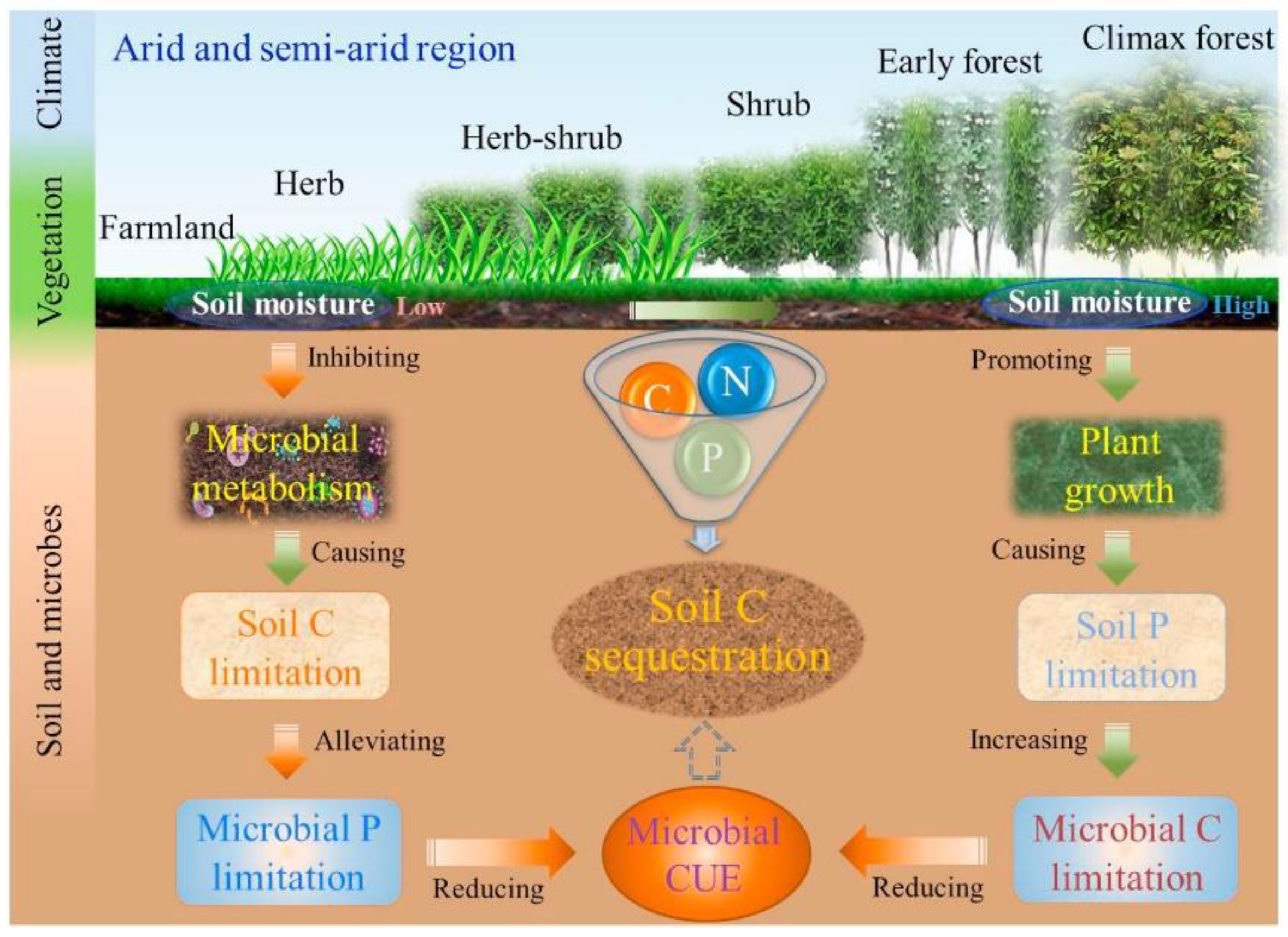
Ecological stoichiometry is a key control driver of microbial metabolic activity [103,104]. Microbial growth and metabolism are optimal if the stoichiometry of microbial demand matches that of the soil matrix [105]; conversely, if in a stoichiometric imbalance, microbial activity will be limited by specific nutrients. Thus, microbial growth and training are often singularly or jointly limited by resources (e.g., C, N, and P) in terrestrial ecosystems [106], as shown in Figure 5 [104].
MBC and MBN are usually positively correlated [107], and high microbial biomass increases demand for N during microbial life activities [108]. Moderate N input increases microbial activity by supplying energy to microorganisms, which respond to changes in soil nutrient effectiveness by altering their chemoenzymatic-regulated carbon assimilation processes [109]. According to steady-state stoichiometric theory, microorganisms increase CUE to adapt to high N environments [79], and soil microbial CUE negatively correlates with soil matrix C/N [7,110]. When N is limiting, catabolic and anabolic processes of microorganisms in soil uncouple [79], and microbial metabolites such as extracellular enzymes and polysaccharides increase for energy [111], producing more significant C loss; at the same time, microorganisms release excess carbon from the substrate as CO2 (a respiratory metabolite) and CUE decrease. As N availability increases, microbes allocate more carbon to maintain a stable stoichiometric ratio of microbes (C/N = 7–8.6) [112]. At the same time, microbes also reduce the secretion of extracellular enzymes used to mine N [113], resulting in higher CUE [104]. N addition mainly promotes the efficiency of microbial utilization of active C. However, it has also been found that an increase in N alone has no effect on the decomposition of organic matter in soil [114]. The simultaneous addition of N and P affects the conversion of organic matter because N enrichment also exacerbates ecosystem dependence on phosphorus. The chemometrics suggest that the optimal C:N:P ratio to satisfy microbial activity is 60:7:1 [115].
During progressive N enrichment, microbial growth will gradually shift from nutrient limitation (e.g., N and P) to C limitation [116,117]. Microbial C metabolism, especially microbial growth, is inherently more C-limited than nutrient limitation. This is because the reduced power and energy required to sustain substrate uptake and metabolism come from the oxidation of other substrates. Carbon taken up by microbial cells can only be used for their growth after the different basic energy maintenance needs are met, producing positive CUE [118]. Previous studies have proposed several potential mechanisms to explain the increased C limitation during N enrichment. Soil acidification due to N enrichment significantly increased the fungal: bacterial and MBC: MBN ratios [119], thus reducing the C/N gap between microbial biomass and its resources. Furthermore, this gap decreased linearly with decreasing soil pH, resulting in a shift from N limitation to C limitation. Nitrogen-induced changes in biological C accessibility have an essential role in the growth of microbial C metabolism. N addition decreases C availability by increasing the N content in plant litter [120], decreasing C/N in the soil matrix, and reducing C availability. Second, N addition decreases the abundance of bacterial and fungal communities [121], which may be attributed to a decrease in subsurface carbon allocation. Plants provide carbon to soil microbes in exchange for other soil nutrients such as N [122]; however, due to the increased effectiveness of N under N deposition, plants have a reduced need to acquire N and thus release less carbon to soil microbes [123]; third, soil acidification caused by N addition reduces plant productivity and root exudation, thus reducing effective soil C [124]. Fourth, the mass of organic matter decreases under N addition, and condensation reactions between mineral N and organic matter increase recalcitrant organic matter [125]. However, it has also been suggested that increased plant carbon input and reduced mineral protection [126] under N addition can alleviate microbial C limitation by increasing soil C availability, promoting microbial growth, and increasing microbial CUE.
3.1.3. Substrate Quality and Bioavailability
Microbial CUE may be influenced by the substrate quality and the metabolic pathways used for substrate utilization, as microbial decomposition uses soil organic carbon and plant residues to sustain its life activities [127,128]. Microbial decomposition of low-quality substrates (e.g., lignin) requires more enzymatic processes [129], increasing microbial extracellular enzyme secretion and respiratory metabolism and resulting in lower microbial CUE. As Allison et al. [130] found, difficult-to-use polymers such as lignin and cellulose must depolymerize to be taken up by microbial cells, which means that microorganisms reallocate more C elements from cell growth to extracellular enzyme secretion.
N addition inhibits microbial CUE by reducing substrate quality in four main ways. First, N addition usually promotes the production of recalcitrant substrates (e.g., lignin) of plant origin [40]. Second, increased N effectiveness leads to decreased activity of lignin-degrading enzymes secreted by fungi [116,131], reducing substrate effectiveness, as lignin can bind to or physically prevent access to other compounds (e.g., cellulose) in plant tissues. Third, nitrogenous compounds may react chemically with phenolics to promote the production of recalcitrant organic matter [132], and Gillespie et al. [133] found that NH4+ forms anti-biodegradable complexes with lignin. Fourth, nitrogen-containing compounds can also coagulate with carbohydrates, producing melanin [134]. Melanin is a highly aromatic biopolymer [135] resistant to decomposition.
3.2. Chemical Bonding of Mineral–Organic Complexes
Soil mineral–organic complexes are connected in various ways, but mainly through multivalent cationic bond bridges (Ca2+, Mg2+, Fe3+, and Al3+) linking the organic matter to the soil minerals [136] or through functional groups (e.g., -COOH and -OH) on the surface of the organic matter in the form of hydrogen bonds to the soil minerals. Soil minerals that play a significant role in stabilizing organic carbon are clay minerals and Fe-Al oxides, the content of which determines the potential of the soil for organic carbon sorption. The main factor affecting soil minerals’ surface charge and sorption sites is pH [17]. Nitrogen addition can affect the persistent sequestration of MAOC by lowering the soil PH and changing the mineral–organic complex interaction.
3.2.1. Soil Minerals
Fe-Al oxides have a variable charge, and clay minerals have a permanent negative charge [137]. Kaiser et al. [138] found a positive correlation between soil organic carbon content and iron oxide content. Fe-Al oxides and their hydrates are positively charged under acidic conditions [137]. They can promote the chemical bonding of mineral–organic complexes through several effects: first, Fe-Al oxides and their hydrates immobilize negatively charged organic matter in soils through positive and negative charge adsorption [139]; second, Fe-Al oxides and their hydrates act as a bond bridge for clay mineral–organic matter binding to form a clay mineral–Fe-Al oxides–organic matter tri-complex [57]; third, the positively charged Fe-Al oxides also neutralize the negative charge on the surface of clay minerals and promote the adsorption of organic matter [140]. Therefore, the decrease in pH caused by nitrogen addition can promote organic carbon adsorption by soil minerals.
However, oxalic acid in root secretions under N addition increases microbial access to MAOC by releasing organic compounds from protective association with minerals [126]. Furthermore, most of the H+ also reacts with soil clay minerals, releasing aluminum from the soil aluminosilicate mineral structure and producing exchangeable Al3+, leading to a decrease in soil buffering capacity [141], which is detrimental to the long-term fixation of soil organic carbon.
3.2.2. Bond Bridge
MAOC content depends on the balance between the adsorption and desorption of charged compounds. Bond bridges formed by Ca2+ and Mg2+ are the primary stabilization mechanism of organic carbon bound to minerals in neutral and alkaline soils [142]. H+ competes for adsorption sites with exchangeable salt-based cations such as Na+, K+, Ca2+, and Mg2+ during soil acidification [20], leading to extensive leaching of salt-based ions, which affects the metal bond bridges of mineral–organic complexes and reduces the chemical stability of organic matter. Ye et al. [17] found that the Fe-bound organic carbon content increased and the total mineral–organic bound carbon content decreased with decreasing pH in alkaline soils. They suggest that acidification partially shifts the mineral-organic complexes from calcium ion bond bridges to iron ion bond bridges.
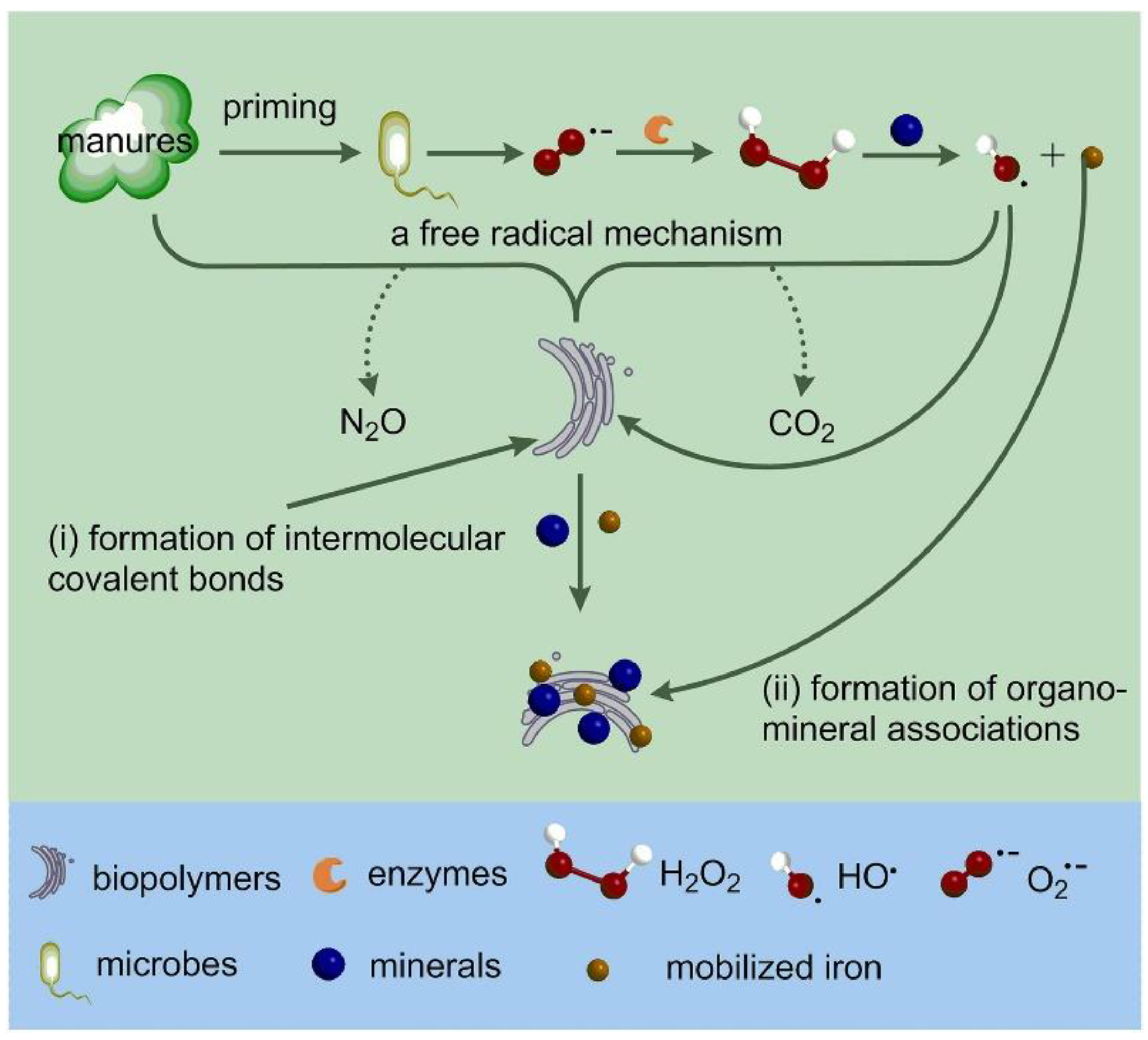
Microorganisms generally have the ability to superoxide (O2−). Long-term use of organic nitrogen fertilizer significantly increased soil bacterial diversity, generated easily mobile Fe oxides [143,144], stimulated Fenton/Fenton-like reactions catalyzed by Fe minerals in the soil, and promoted the formation of hydroxyl radicals (HO·) (Equations (4) and (5)) [145]. Hydroxyl radicals (HO·) increase the appearance of intermolecular covalent bonds and mineral-organic complexes in SOC (Figure 6) [146], which is essential for long-term organic carbon sequestration in Fe-rich soils (e.g., red dirt and red brick soil).
4. Conclusions
This review discusses the transformation direction of particulate organic carbon (POC) and mineral-associated organic carbon (MAOC) with different sources and assignment mechanisms under nitrogen input to facilitate a more comprehensive understanding of soil organic carbon sequestration and transformation mechanisms.
Moderate N input promotes POC input by increasing net primary productivity. However, soil SOC sequestration may depend on the quality rather than the quantity of POC input. Increased intake of recalcitrant organic matter under N addition generates microbial resistance, which contributes to POC formation but is detrimental to long-term organic carbon sequestration. POC is mainly found in soil aggregates, and reasonable N input helps stabilize soil structure by promoting organic cement production and entanglement of plant roots, and the decomposition of POC is reduced.
N input can affect MAOC sequestration by altering the chemical bonding of microbial CUE and mineral–organic complexes. N inputs drive changes in microbial CUE through changes in soil microbial activity and community structure, substrate quality and bioavailability, and ecological stoichiometry. However, there is uncertainty about the direction of change. High inorganic N input decreases pH, which makes Fe-Al oxides positively charged and promotes the formation of MAOC. In neutral and alkaline soils, H+ competes with exchangeable salt-based cations for adsorption sites, affecting cationic bond bridges and reducing organic carbon stability.
Briefly, the effects of nitrogen on the transformation of POC and MAOC are influenced by the interaction of soil physicochemical properties, the level and type of N input, microorganisms, etc. These cross-influences lead to contradictory directions in the transformation of SOC under N input, as it currently appears. These conflicting responses will further increase the uncertainty in predicting the change in POC and MAOC under N addition. Despite the significant limitations of the current study, given the strong coupling of carbon and nitrogen in soils, these results can provide a theoretical basis for maximizing the soil carbon pool to some extent. We believe that future work should further explore the share of positive and negative feedback mechanisms generated by these influencing factors on the POC and MAOC fugacity processes to reach the maximum soil sequestration capacity.
Author Contributions
Conceptualization, D.M., L.M., and Z.L.; validation, H.C. and Y.S.; writing—original draft, D.M.; writing—review and editing, M.L. and D.X.; funding acquisition, L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science and Technology of China (Grant Number: 2022xjkk0300); the National Natural Science Foundation of China (Grant Number: 11275301; 12075261); and the Innovation Fund of the Institute of High Energy Physics, Chinese Academy of Sciences (2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimann, M.; Reichstein, M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 2008, 451, 289–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, O.; Petersen, G.W.; Needelman, B.A. Environmental Indicators of Agroecosystems. Adv. Agron. 1999, 69, 75–97. [Google Scholar]
- Miner, G.L.; Delgado, J.A.; Ippolito, J.A.; Barbarick, K.A.; Stewart, C.E.; Manter, D.K.; Del Grosso, S.J.; Halvorson, A.D.; Floyd, B.A.; D’Adamo, R.E. Influence of long-term nitrogen fertilization on crop and soil micronutrients in a no-till maize cropping system. Field Crops Res. 2018, 228, 170–182. [Google Scholar] [CrossRef]
- Huang, J.; Liu, W.; Yang, S.; Yang, L.; Peng, Z.; Deng, M.; Xu, S.; Zhang, B.; Ahirwal, J.; Liu, L. Plant carbon inputs through shoot, root, and mycorrhizal pathways affect soil organic carbon turnover differently. Soil Biol. Biochem. 2021, 160, 108322. [Google Scholar] [CrossRef]
- Soares, M.; Rousk, J. Microbial growth and carbon use efficiency in soil: Links to fungal-bacterial dominance, SOC-quality and stoichiometry. Soil Biol. Biochem. 2019, 131, 195–205. [Google Scholar] [CrossRef]
- Wei, X.; Zhu, Z.; Liu, Y.; Luo, Y.; Deng, Y.; Xu, X.; Liu, S.; Richter, A.; Shibistova, O.; Guggenberger, G.; et al. C:N:P stoichiometry regulates soil organic carbon mineralization and concomitant shifts in microbial community composition in paddy soil. Biol. Fertil. Soils 2020, 56, 1093–1107. [Google Scholar] [CrossRef]
- Barton, L.; Butterbach-Bahl, K.; Kiese, R.; Murphy, D.V. Nitrous oxide fluxes from a grain-legume crop (narrow-leafed lupin) grown in a semiarid climate. Glob. Change Biol. 2011, 17, 1153–1166. [Google Scholar] [CrossRef]
- Frey, S.D.; Ollinger, S.; Nadelhoffer, K.; Bowden, R.; Brzostek, E.; Burton, A.; Caldwell, B.A.; Crow, S.; Goodale, C.L.; Grandy, A.S.; et al. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests. Biogeochemistry 2014, 121, 305–316. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Sithole, N.J.; Magwaza, L.S.; Thibaud, G.R. Long-term impact of no-till conservation agriculture and N-fertilizer on soil aggregate stability, infiltration and distribution of C in different size fractions. Soil Tillage Res. 2019, 190, 147–156. [Google Scholar] [CrossRef]
- Romanenkov, V.; Belichenko, M.; Petrova, A.; Raskatova, T.; Jahn, G.; Krasilnikov, P. Soil organic carbon dynamics in long-term experiments with mineral and organic fertilizers in Russia. Geoderma Reg. 2019, 17, e00221. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Wang, M.Y.; Hu, S.J.; Zhang, X.D.; Ouyang, Z.; Zhang, G.L.; Huang, B.A.; Zhao, S.W.; Wu, J.S.; Xie, D.T.; et al. Economics- and policy-driven organic carbon input enhancement dominates soil organic carbon accumulation in Chinese croplands. Proc. Natl. Acad. Sci. USA 2018, 115, 4045–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Tang, M.; Zhu, B. Soil priming effect and its responses to nutrient addition along a tropical forest elevation gradient. Glob. Change Biol. 2021, 27, 2793–2806. [Google Scholar] [CrossRef]
- Zhang, X.-M.; Wang, Y.-D.; Zhao, Y.; Xu, X.-W.; Lei, J.-Q.; Hill, R.L. Litter decomposition and nutrient dynamics of three woody halophytes in the Taklimakan Desert Highway Shelterbelt. Arid Land Res. Manag. 2017, 31, 335–351. [Google Scholar] [CrossRef]
- Du, E.; de Vries, W. Nitrogen-induced new net primary production and carbon sequestration in global forests. Environ. Pollut. 2018, 242, 1476–1487. [Google Scholar] [CrossRef]
- Ye, C.L.; Chen, D.M.; Hall, S.J.; Pan, S.; Yan, X.B.; Bai, T.S.; Guo, H.; Zhang, Y.; Bai, Y.F.; Hu, S.J. Reconciling multiple impacts of nitrogen enrichment on soil carbon: Plant, microbial and geochemical controls. Ecol. Lett. 2018, 21, 1162–1173. [Google Scholar] [CrossRef] [Green Version]
- Boxman, A.W.; Peters, R.C.J.H.; Roelofs, J.G.M. Long term changes in atmospheric N and S throughfall deposition and effects on soil solution chemistry in a Scots pine forest in The Netherlands. Environ. Pollut. 2008, 156, 1252–1259. [Google Scholar] [CrossRef]
- Skeffington, R.A. Accelerated nitrogen inputs—A new problem or a new perspective? Plant Soil 1990, 128, 1–11. [Google Scholar] [CrossRef]
- Barak, P.; Jobe, B.O.; Krueger, A.R.; Peterson, L.A.; Laird, D.A. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 1997, 197, 61–69. [Google Scholar] [CrossRef]
- Drabek, O.; Boruvka, L.; Mladkova, L.; Kocarek, M. Possible method of aluminium speciation in forest soils. J. Inorg. Biochem. 2003, 97, 8–15. [Google Scholar] [CrossRef]
- Wang, J.Q.; Shi, X.Z.; Zheng, C.Y.; Suter, H.; Huang, Z.Q. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total Environ. 2021, 755, 142449. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.; Zhou, W.J.; Fang, Y.T.; Bing, H.J.; Sun, X.Y.; Wang, G.X. Anthropogenic Nitrogen Deposition Increases Soil Carbon by Enhancing New Carbon of the Soil Aggregate Formation. J. Geophys. Res.-Biogeo 2019, 124, 572–584. [Google Scholar] [CrossRef]
- Tao, B.X.; Wang, Y.P.; Yu, Y.; Li, Q.Z.; Luo, C.Y.; Zhang, B.H. Interactive effects of nitrogen forms and temperature on soil organic carbon decomposition in the coastal wetland of the Yellow River Delta, China. Catena 2018, 165, 408–413. [Google Scholar] [CrossRef]
- Weidhuner, A.; Hanauer, A.; Krausz, R.; Crittenden, S.J.; Gage, K.; Sadeghpour, A. Tillage impacts on soil aggregation and aggregate-associated carbon and nitrogen after 49 years. Soil Tillage Res. 2021, 208, 104878. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Change Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Miltner, A.; Bombach, P.; Schmidt-Brucken, B.; Kastner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Chen, J.G.; Xiao, W.; Zheng, C.Y.; Zhu, B. Nitrogen addition has contrasting effects on particulate and mineral-associated soil organic carbon in a subtropical forest. Soil Biol. Biochem. 2020, 142, 107708. [Google Scholar] [CrossRef]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Turner, B.L.; Zhou, G.Y.; Mo, J.M. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. Proc. Natl. Acad. Sci. USA 2021, 118, e2020790118. [Google Scholar] [CrossRef]
- Mosier, S.; Apfelbaum, S.; Byck, P.; Calderon, F.; Teague, R.; Thompson, R.; Cotrufo, M.F. Adaptive multi-paddock grazing enhances soil carbon and nitrogen stocks and stabilization through mineral association in southeastern U.S. grazing lands. J. Environ. Manag. 2021, 288, 112409. [Google Scholar] [CrossRef]
- Cheng, S.; Fang, H.; Xu, M.; Geng, J.; He, S.; Yu, G.; Chao, Z. Regulation of plant -soil -microbe interactions to soil organic carbon in natural ecosystems under elevated nitrogen deposition: A review. Acta Ecol. Sin. 2018, 38, 8285–8295. [Google Scholar]
- Johnston, A.E.; Poulton, P.R.; Coleman, K. Chapter 1 Soil Organic Matter: Its Importance in Sustainable Agriculture and Carbon Dioxide Fluxes. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2009; Volume 101, pp. 1–57. [Google Scholar]
- Schulte-Uebbing, L.; de Vries, W. Global-scale impacts of nitrogen deposition on tree carbon sequestration in tropical, temperate, and boreal forests: A meta-analysis. Glob. Change Biol. 2018, 24, E416–E431. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Fornara, D.A.; Tilman, D. Soil carbon sequestration in prairie grasslands increased by chronic nitrogen addition. Ecology 2012, 93, 2030–2036. [Google Scholar] [CrossRef] [Green Version]
- Lajtha, K.; Townsend, K.L.; Kramer, M.G.; Swanston, C.; Bowden, R.D.; Nadelhoffer, K. Changes to particulate versus mineral-associated soil carbon after 50 years of litter manipulation in forest and prairie experimental ecosystems. Biogeochemistry 2014, 119, 341–360. [Google Scholar] [CrossRef]
- Wei, H.; Chen, X.; He, J.; Huang, L.; Shen, W. Warming but Not Nitrogen Addition Alters the Linear Relationship between Microbial Respiration and Biomass. Front. Microbiol. 2019, 10, 1055. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.L.; Hättenschwiler, S.; Yang, J.J.; Sistla, S.; Wei, H.W.; Zhang, Z.W.; Hu, Y.Y.; Wang, R.Z.; Cui, S.Y.; Lü, X.T.; et al. Increasing rates of long-term nitrogen deposition consistently increased litter decomposition in a semi-arid grassland. New Phytol. 2021, 229, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xu, S.; Ciais, P.; Manzoni, S.; Fang, J.; Yu, G.; Tang, X.; Zhou, P.; Wang, W.; Yan, J.; et al. Climate and litter C/N ratio constrain soil organic carbon accumulation. Natl. Sci. Rev. 2019, 6, 746–757. [Google Scholar] [CrossRef] [Green Version]
- Marschner, B.; Brodowski, S.; Dreves, A.; Gleixner, G.; Gude, A.; Grootes, P.M.; Hamer, U.; Heim, A.; Jandl, G.; Ji, R.; et al. How relevant is recalcitrance for the stabilization of organic matter in soils. J. Plant Nutr. Soil Sci. 2008, 171, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Ziter, C.; MacDougall, A.S. Nutrients and defoliation increase soil carbon inputs in grassland. Ecology 2013, 94, 106–116. [Google Scholar] [CrossRef]
- Fulton-Smith, S.; Cotrufo, M.F. Pathways of soil organic matter formation from above and belowground inputs in a Sorghum bicolor bioenergy crop. GCB Bioenergy 2019, 11, 971–987. [Google Scholar] [CrossRef] [Green Version]
- Sokol, N.W.; Bradford, M.A. Microbial formation of stable soil carbon is more efficient from belowground than aboveground input. Nat. Geosci. 2019, 12, 46–53. [Google Scholar] [CrossRef]
- Craig, M.E.; Geyer, K.M.; Beidler, K.V.; Brzostek, E.R.; Frey, S.D.; Stuart Grandy, A.; Liang, C.; Phillips, R.P. Fast-decaying plant litter enhances soil carbon in temperate forests but not through microbial physiological traits. Nat. Commun. 2022, 13, 1229. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef]
- Jastrow, J.D. Soil aggregate formation and the accrual of particulate and mineral-associated organic matter. Soil Biol. Biochem. 1996, 28, 665–676. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Ayoubi, S.; Karchegani, P.M.; Mosaddeghi, M.R.; Honarjoo, N. Soil aggregation and organic carbon as affected by topography and land use change in western Iran. Soil Tillage Res. 2012, 121, 18–26. [Google Scholar] [CrossRef]
- He, X.; Huang, Y.; Zhang, Q.; Ye, S.; Wang, S. Distribution of organic carbon fractions in soil aggregates in Chinese fir plantations with different stand ages. Ecol. Process. 2021, 10, 49. [Google Scholar] [CrossRef]
- Wei, H.; Deng, Y.; Huang, J.; He, L.; Tang, Q.; Xiao, Y. A quantitative study of the influence of soil organic carbon and pore characteristics on the stability of aggregates of the karst peak-cluster depression area in Southwest China. J. Soil Sediment 2022, 1–19. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, S.; Liu, L.; Wu, L.; Ding, X. Combined organic amendments and mineral fertilizer application increase rice yield by improving soil structure, P availability and root growth in saline-alkaline soil. Soil Tillage Res. 2021, 212, 105060. [Google Scholar] [CrossRef]
- Zhong, X.L.; Li, J.T.; Li, X.J.; Ye, Y.C.; Liu, S.S.; Hallett, P.D.; Ogden, M.R.; Naveed, M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma 2017, 285, 323–332. [Google Scholar] [CrossRef]
- Lu, X.; Hou, E.; Guo, J.; Gilliam, F.S.; Li, J.; Tang, S.; Kuang, Y. Nitrogen addition stimulates soil aggregation and enhances carbon storage in terrestrial ecosystems of China: A meta-analysis. Glob. Change Biol. 2021, 27, 2780–2792. [Google Scholar] [CrossRef] [PubMed]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Oades, J.M.; Waters, A.G. Aggregate Hierarchy in Soils. Aust. J. Soil Res. 1991, 29, 815–828. [Google Scholar] [CrossRef]
- Totsche, K.U.; Amelung, W.; Gerzabek, M.H.; Guggenberger, G.; Klumpp, E.; Knief, C.; Lehndorff, E.; Mikutta, R.; Peth, S.; Prechtel, A.; et al. Microaggregates in soils. J. Plant Nutr. Soil Sci. 2018, 181, 104–136. [Google Scholar] [CrossRef] [Green Version]
- Abiven, S.; Menasseri, S.; Angers, D.A.; Leterme, P. Dynamics of aggregate stability and biological binding agents during decomposition of organic materials. Eur. J. Soil Sci. 2007, 58, 239–247. [Google Scholar] [CrossRef]
- Haynes, R.J.; Swift, R.S.; Stephen, R.C. Influence of mixed cropping rotations (pasture-arable) on organic matter content, water stable aggregation and clod porosity in a group of soils. Soil Tillage Res. 1991, 19, 77–87. [Google Scholar] [CrossRef]
- Luo, R.; Kuzyakov, Y.; Liu, D.; Fan, J.; Luo, J.; Lindsey, S.; He, J.-S.; Ding, W. Nutrient addition reduces carbon sequestration in a Tibetan grassland soil: Disentangling microbial and physical controls. Soil Biol. Biochem. 2020, 144, 107764. [Google Scholar] [CrossRef]
- Caravaca, F.; Barea, J.M.; Roldán, A. Synergistic influence of an arbuscular mycorrhizal fungus and organic amendment on Pistacia lentiscus L. seedlings afforested in a degraded semiarid soil. Soil Biol. Biochem. 2002, 34, 1139–1145. [Google Scholar] [CrossRef]
- Yue, K.; Fornara, D.A.; Li, W.; Ni, X.; Peng, Y.; Liao, S.; Tan, S.; Wang, D.; Wu, F.; Yang, Y. Nitrogen addition affects plant biomass allocation but not allometric relationships among different organs across the globe. J. Plant Ecol. 2020, 14, 361–371. [Google Scholar] [CrossRef]
- Rukshana, F.; Butterly, C.R.; Xu, J.M.; Baldock, J.A.; Tang, C.X. Organic anion-to-acid ratio influences pH change of soils differing in initial pH. J. Soil Sediment 2014, 14, 407–414. [Google Scholar] [CrossRef]
- Noble, A.D.; Zenneck, I.; Randall, P.J. Leaf litter ash alkalinity and neutralisation of soil acidity. Plant Soil 1996, 179, 293–302. [Google Scholar] [CrossRef]
- Du, Y.H.; Guo, P.; Liu, J.Q.; Wang, C.Y.; Yang, N.; Jiao, Z.X. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Change Biol. 2014, 20, 3222–3228. [Google Scholar] [CrossRef]
- Jardine, P.M.; Weber, N.L.; Mccarthy, J.F. Mechanisms of Dissolved Organic-Carbon Adsorption on Soil. Soil Sci. Soc. Am. J. 1989, 53, 1378–1385. [Google Scholar] [CrossRef]
- Huang, B.; Qiu, M.; Lin, J.; Chen, J.; Jiang, F.; Wang, M.-K.; Ge, H.; Huang, Y. Correlation between shear strength and soil physicochemical properties of different weathering profiles of the non-eroded and collapsing gully soils in southern China. J. Soil Sediment 2019, 19, 3832–3846. [Google Scholar] [CrossRef]
- Peng, X.; Yan, X.; Zhou, H.; Zhang, Y.Z.; Sun, H. Assessing the contributions of sesquioxides and soil organic matter to aggregation in an Ultisol under long-term fertilization. Soil Tillage Res. 2015, 146, 89–98. [Google Scholar] [CrossRef]
- Raza, S.; Miao, N.; Wang, P.; Ju, X.; Chen, Z.; Zhou, J.; Kuzyakov, Y. Dramatic loss of inorganic carbon by nitrogen-induced soil acidification in Chinese croplands. Glob. Change Biol. 2020, 26, 3738–3751. [Google Scholar] [CrossRef]
- Zamanian, K.; Zarebanadkouki, M.; Kuzyakov, Y. Nitrogen fertilization raises CO2 efflux from inorganic carbon: A global assessment. Glob. Change Biol. 2018, 24, 2810–2817. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Amelung, W.; Lehmann, J.; Kastner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Change Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef] [PubMed]
- McNally, S.R.; Beare, M.H.; Curtin, D.; Meenken, E.D.; Kelliher, F.M.; Pereira, R.C.; Shen, Q.H.; Baldock, J. Soil carbon sequestration potential of permanent pasture and continuous cropping soils in New Zealand. Glob. Change Biol. 2017, 23, 4544–4555. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, É.P. Calcium-mediated stabilisation of soil organic carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.P.W.; Scheel, T.; Mikutta, R.; van Hees, P.; Kaiser, K.; Kalbitz, K. Sorptive stabilization of organic matter by amorphous Al hydroxide. Geochim. Cosmochim. Acta 2010, 74, 1606–1619. [Google Scholar] [CrossRef] [Green Version]
- Domeignoz-Horta, L.A.; Pold, G.; Liu, X.J.A.; Frey, S.D.; Melillo, J.M.; DeAngelis, K.M. Microbial diversity drives carbon use efficiency in a model soil. Nat. Commun. 2020, 11, 3684. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Ågren, G.I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef]
- Bradford, M.A.; Wieder, W.R.; Bonan, G.B.; Fierer, N.; Raymond, P.A.; Crowther, T.W. Managing uncertainty in soil carbon feedbacks to climate change. Nat. Clim. Change 2016, 6, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Keiblinger, K.M.; Hall, E.K.; Wanek, W.; Szukics, U.; Hammerle, I.; Ellersdorfer, G.; Bock, S.; Strauss, J.; Sterflinger, K.; Richter, A.; et al. The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency. FEMS Microbiol. Ecol. 2010, 73, 430–440. [Google Scholar] [CrossRef]
- Gommers, P.J.F.; Vanschie, B.J.; Vandijken, J.P.; Kuenen, J.G. Biochemical Limits to Microbial-Growth Yields—An Analysis of Mixed Substrate Utilization. Biotechnol. Bioeng. 1988, 32, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.B.; Bezemer, T.M.; Yang, J.J.; Lu, X.T.; Li, X.Y.; Liang, W.J.; Han, X.G.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Liu, W.; Liu, L.; Yang, X.; Deng, M.; Wang, Z.; Wang, P.; Yang, S.; Li, P.; Peng, Z.; Yang, L.; et al. Long-term nitrogen input alters plant and soil bacterial, but not fungal beta diversity in a semiarid grassland. Glob. Change Biol. 2021, 27, 3939–3950. [Google Scholar] [CrossRef] [PubMed]
- Oquist, M.G.; Erhagen, B.; Haei, M.; Sparrman, T.; Ilstedt, U.; Schleucher, J.; Nilsson, M.B. The effect of temperature and substrate quality on the carbon use efficiency of saprotrophic decomposition. Plant Soil 2017, 414, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Talgre, L.; Roostalu, H.; Mäeorg, E.; Lauringson, E. Nitrogen and carbon release during decomposition of roots and shoots of leguminous green manure crops. Agron. Res. 2017, 15, 594–601. [Google Scholar]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.Z.; Li, Y.; Zhang, X.D.; Yan, Z.Q.; Wu, H.D.; Li, M.; Yan, L.; Zhang, K.R.; Wang, J.Z.; Kang, X.M. Soil pH and nutrients shape the vertical distribution of microbial communities in an alpine wetland. Sci. Total Environ. 2021, 774, 145780. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, P.; Zeng, Z. Dynamics of Bacterial Communities in a 30-Year Fertilized Paddy Field under Different Organic–Inorganic Fertilization Strategies. Agronomy 2019, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.A.; Puissant, J.; Buckeridge, K.M.; Goodall, T.; Jehmlich, N.; Chowdhury, S.; Gweon, H.S.; Peyton, J.M.; Mason, K.E.; van Agtmaal, M.; et al. Land use driven change in soil pH affects microbial carbon cycling processes. Nat. Commun. 2018, 9, 3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Q.; Huang, Y.; Wu, L.; Tian, Y.; Wang, Q.; Wang, B.; Xu, M.; Zhang, W. Long-term manuring increases microbial carbon use efficiency and mitigates priming effect via alleviated soil acidification and resource limitation. Biol. Fertil. Soils 2021, 57, 925–934. [Google Scholar] [CrossRef]
- Kinraide, T.B. Toxicity factors in acidic forest soils: Attempts to evaluate separately the toxic effects of excessive Al3+ and H+ and insufficient Ca2+ and Mg2+ upon root elongation. Eur. J. Soil Sci. 2003, 54, 323–333. [Google Scholar] [CrossRef]
- Malik, A.A.; Thomson, B.C.; Whiteley, A.S.; Bailey, M.; Griffiths, R.I. Bacterial Physiological Adaptations to Contrasting Edaphic Conditions Identified Using Landscape Scale Metagenomics. mBio 2017, 8, e00799-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, C.; Han, S.; Appanna, V.P.; Thomas, S.C.; Ulibarri, G.; Appanna, V.D. Metabolic reengineering invoked by microbial systems to decontaminate aluminum: Implications for bioremediation technologies. Biotechnol. Adv. 2013, 31, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Silver, S.; Nucifora, G.; Chu, L.; Misra, T.K. Bacterial resistance ATPases: Primary pumps for exporting toxic cations and anions. Trends Biochem. Sci. 1989, 14, 76–80. [Google Scholar] [CrossRef]
- Jones, D.L.; Cooledge, E.C.; Hoyle, F.C.; Griffiths, R.I.; Murphy, D.V. pH and exchangeable aluminum are major regulators of microbial energy flow and carbon use efficiency in soil microbial communities. Soil Biol. Biochem. 2019, 138, 107584. [Google Scholar] [CrossRef]
- Bellion, M.; Courbot, M.; Jacob, C.; Blaudez, D.; Chalot, M. Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi. FEMS Microbiol. Lett. 2006, 254, 173–181. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F. Aluminum-Nitrogen Interactions in the Soil-Plant System. Front. Plant Sci. 2018, 9, 807. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Yang, L.; Lai, X.; Yao, Q.; Chen, K. Influence of Al(III) on biofilm and its extracellular polymeric substances in sequencing batch biofilm reactors. Environ. Technol. 2019, 40, 53–59. [Google Scholar] [CrossRef]
- Malik, A.A.; Martiny, J.B.H.; Brodie, E.L.; Martiny, A.C.; Treseder, K.K.; Allison, S.D. Defining trait-based microbial strategies with consequences for soil carbon cycling under climate change. ISME J. 2020, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad Shah, J.J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, X.; Zhang, X.; Ju, W.; Duan, C.; Guo, X.; Wang, Y.; Fang, L. Soil moisture mediates microbial carbon and phosphorus metabolism during vegetation succession in a semiarid region. Soil Biol. Biochem. 2020, 147, 107814. [Google Scholar] [CrossRef]
- Sterner, R. Modelling interactions of food quality and quantity in homeostatic consumers. Freshw. Biol. 1997, 38, 473–481. [Google Scholar] [CrossRef]
- Wang, X.; Cui, Y.; Wang, Y.; Duan, C.; Niu, Y.; Sun, R.; Shen, Y.; Guo, X.; Fang, L. Ecoenzymatic stoichiometry reveals phosphorus addition alleviates microbial nutrient limitation and promotes soil carbon sequestration in agricultural ecosystems. J. Soil Sediment 2022, 22, 536–546. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Q.; Yang, F.; Lei, Y.; Zhang, Q.; Cheng, X. Afforestation impacts microbial biomass and its natural 13C and 15N abundance in soil aggregates in central China. Sci. Total Environ. 2016, 568, 52–56. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Traoré, O.Y.A.; Kiba, D.I.; Arnold, M.C.; Fliessbach, A.; Oberholzer, H.R.; Nacro, H.B.; Lompo, F.; Oberson, A.; Frossard, E.; Bünemann, E.K. Fertilization practices alter microbial nutrient limitations after alleviation of carbon limitation in a Ferric Acrisol. Biol. Fertil. Soils 2016, 52, 177–189. [Google Scholar] [CrossRef]
- Li, J.; Sang, C.P.; Yang, J.Y.; Qu, L.R.; Xia, Z.W.; Sun, H.; Jiang, P.; Wang, X.G.; He, H.B.; Wang, C. Stoichiometric imbalance and microbial community regulate microbial elements use efficiencies under nitrogen addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Du, L.; Zhu, Z.; Qi, Y.; Zou, D.; Zhang, G.; Zeng, X.; Ge, T.; Wu, J.; Xiao, Z. Effects of different stoichiometric ratios on mineralisation of root exudates and its priming effect in paddy soil. Sci. Total Environ. 2020, 743, 140808. [Google Scholar] [CrossRef]
- Manzoni, S.; Čapek, P.; Mooshammer, M.; Lindahl, B.D.; Richter, A.; Šantrůčková, H. Optimal metabolic regulation along resource stoichiometry gradients. Ecol. Lett. 2017, 20, 1182–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Zhu, Z.; Xu, X.; Liu, S.; Jones, D.L.; Kuzyakov, Y.; Shibistova, O.; Wu, J.; Ge, T. Carbon and nitrogen recycling from microbial necromass to cope with C:N stoichiometric imbalance by priming. Soil Biol. Biochem. 2020, 142, 107720. [Google Scholar] [CrossRef]
- Reid, J.P.; Adair, E.C.; Hobbie, S.E.; Reich, P.B. Biodiversity, Nitrogen Deposition, and CO2 Affect Grassland Soil Carbon Cycling but not Storage. Ecosystems 2012, 15, 580–590. [Google Scholar] [CrossRef]
- Li, Y.; Niu, S.L.; Yu, G.R. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Glob. Change Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Zhao, J.; Zhang, W.; Xiao, K.; Wang, K. Nitrogen addition aggravates microbial carbon limitation: Evidence from ecoenzymatic stoichiometry. Geoderma 2018, 329, 61–64. [Google Scholar] [CrossRef]
- Kopáček, J.; Cosby, B.J.; Evans, C.D.; Hruška, J.; Moldan, F.; Oulehle, F.; Šantrůčková, H.; Tahovská, K.; Wright, R.F. Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: Linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry 2013, 115, 33–51. [Google Scholar] [CrossRef]
- Adingo, S.; Yu, J.R.; Xuelu, L.; Li, X.; Jing, S.; Xiaong, Z. Variation of soil microbial carbon use efficiency (CUE) and its Influence mechanism in the context of global environmental change: A review. PeerJ 2021, 9, e12131. [Google Scholar] [CrossRef]
- Li, T.; Wang, R.; Cai, J.; Meng, Y.; Wang, Z.; Feng, X.; Liu, H.; Turco, R.F.; Jiang, Y. Enhanced carbon acquisition and use efficiency alleviate microbial carbon relative to nitrogen limitation under soil acidification. Ecol. Process. 2021, 10, 32. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef] [Green Version]
- Treseder, K.K.; Allen, E.B.; Egerton-Warburton, L.M.; Hart, M.M.; Klironomos, J.N.; Maherali, H.; Tedersoo, L. Arbuscular mycorrhizal fungi as mediators of ecosystem responses to nitrogen deposition: A trait-based predictive framework. J. Ecol. 2018, 106, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Vasar, M.; Andreson, R.; Davison, J.; Jairus, T.; Moora, M.; Remm, M.; Young, J.P.W.; Zobel, M.; Öpik, M. Increased sequencing depth does not increase captured diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.R.; Thornley, J.H.M. A Model of Shoot—Root Partitioning with Optimal-Growth. Ann. Bot. 1987, 60, 133–142. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; McDowell, W.H. Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests. Biogeochemistry 2011, 104, 203–225. [Google Scholar] [CrossRef] [Green Version]
- Keiluweit, M.; Bougoure, J.J.; Nico, P.S.; Pett-Ridge, J.; Weber, P.K.; Kleber, M. Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Change 2015, 5, 588–595. [Google Scholar] [CrossRef] [Green Version]
- Roller, B.R.; Schmidt, T.M. The physiology and ecological implications of efficient growth. ISME J. 2015, 9, 1481–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.L.; Hill, P.W.; Smith, A.R.; Farrell, M.; Ge, T.; Banning, N.C.; Murphy, D.V. Role of substrate supply on microbial carbon use efficiency and its role in interpreting soil microbial community-level physiological profiles (CLPP). Soil Biol. Biochem. 2018, 123, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Craine, J.M.; McLauchlan, K.K.; Schimel, J.P. Litter quality and the temperature sensitivity of decomposition. Ecology 2005, 86, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Allison, S.D. Modeling adaptation of carbon use efficiency in microbial communities. Front. Microbiol. 2014, 5, 571. [Google Scholar] [CrossRef] [Green Version]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L.; Gallo, M.E.; Lauber, C.L. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol. Appl. 2004, 14, 1172–1177. [Google Scholar] [CrossRef]
- Knicker, H. Stabilization of N-compounds in soil and organic-matter-rich sediments—What is the difference? Mar. Chem. 2004, 92, 167–195. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, A.W.; Diochon, A.; Ma, B.L.; Morrison, M.J.; Kellman, L.; Walley, F.L.; Regier, T.Z.; Chevrier, D.; Dynes, J.J.; Gregorich, E.G. Nitrogen input quality changes the biochemical composition of soil organic matter stabilized in the fine fraction: A long-term study. Biogeochemistry 2014, 117, 337–350. [Google Scholar] [CrossRef]
- Semenov, V.M.; Tulina, A.S.; Semenova, N.A.; Ivannikova, L.A. Humification and nonhumification pathways of the organic matter stabilization in soil: A review. Eurasian Soil Sci. 2013, 46, 355–368. [Google Scholar] [CrossRef]
- Bull, A.T. Inhibition of polysaccharases by melanin: Enzyme inhibition in relation to mycolysis. Arch. Biochem. Biophys. 1970, 137, 345–356. [Google Scholar] [CrossRef]
- Chen, J.; Hu, Y.; Hall, S.; Hui, D.; Li, J.; Chen, G.; Sun, L.; Zhang, D.; Deng, Q. Increased Iron-Carbon Interactions under Long-Term Acid Deposition Enhance Soil Organic Carbon Sequestration in a Tropical Forest in Southern China; Research Square: Durham, NC, USA, 2021. [Google Scholar]
- Mustafa, G.; Singh, B.; Kookana, R.S. Cadmium adsorption and desorption behaviour on goethite at low equilibrium concentrations: Effects of pH and index cations. Chemosphere 2004, 57, 1325–1333. [Google Scholar] [CrossRef]
- Kaiser, K.; Zech, W. Nitrate, sulfate, and biphosphate retention in acid forest soils affected by natural dissolved organic carbon. J. Environ. Qual. 1996, 25, 1325–1331. [Google Scholar] [CrossRef]
- Goldberg, S.; Glaubig, R.A. Effect of Saturating Cation, Ph, and Aluminum and Iron-Oxide on the Flocculation of Kaolinite and Montmorillonite. Clays Clay Miner. 1987, 35, 220–227. [Google Scholar] [CrossRef]
- Saidy, A.R.; Smernik, R.J.; Baldock, J.A.; Kaiser, K.; Sanderman, J. The sorption of organic carbon onto differing clay minerals in the presence and absence of hydrous iron oxide. Geoderma 2013, 209, 15–21. [Google Scholar] [CrossRef]
- Bergkvist, B.; Folkeson, L. Soil Acidification and Element Fluxes of a Fagus-Sylvatica Forest as Influenced by Simulated Nitrogen Deposition. Water Air Soil Pollut. 1992, 65, 111–133. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Ge, T.; Zhu, Z.; Luo, Y.; Yang, Y.; Xiao, M.; Yan, Z.; Li, Y.; Wu, J.; Kuzyakov, Y. Comparing carbon and nitrogen stocks in paddy and upland soils: Accumulation, stabilization mechanisms, and environmental drivers. Geoderma 2021, 398, 115121. [Google Scholar] [CrossRef]
- Yu, G.; Xiao, J.; Hu, S.; Polizzotto, M.L.; Zhao, F.; McGrath, S.P.; Li, H.; Ran, W.; Shen, Q. Mineral Availability as a Key Regulator of Soil Carbon Storage. Environ. Sci. Technol. 2017, 51, 4960–4969. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Tang, H.; Kang, W.; Yu, G.; Ran, W.; Hong, J.; Shen, Q. Redox interface-associated organo-mineral interactions: A mechanism for C sequestration under a rice-wheat cropping system. Soil Biol. Biochem. 2018, 120, 12–23. [Google Scholar] [CrossRef]
- Yu, G.-H.; Chi, Z.-L.; Teng, H.H.; Dong, H.-L.; Kappler, A.; Gillings, M.R.; Polizzotto, M.L.; Liu, C.-Q.; Zhu, Y.-G. Fungus-initiated catalytic reactions at hyphal-mineral interfaces drive iron redox cycling and biomineralization. Geochim. Cosmochim. Acta 2019, 260, 192–203. [Google Scholar] [CrossRef]
- Yu, G.; Sun, F.-S.; Yang, L.; He, X.; Polizzotto, M.L. Influence of biodiversity and iron availability on soil peroxide: Implications for soil carbon stabilization and storage. Land Degrad. Dev. 2019, 31, 463–472. [Google Scholar] [CrossRef]
Figure 1.
Effect of nitrogen on POC and MAOC cycle processes.

Figure 2.
Conceptual model of the dynamic responses of soil aggregates to N enrichment and the potential mechanisms involved in soil aggregation and C sequestration. The blue, red, and black boxes indicate variables that are positively, negatively, or not changed by N enrichment. Reprinted with permission from Ref. [54]. Copyright 2021 Wiley.
Figure 2.
Conceptual model of the dynamic responses of soil aggregates to N enrichment and the potential mechanisms involved in soil aggregation and C sequestration. The blue, red, and black boxes indicate variables that are positively, negatively, or not changed by N enrichment. Reprinted with permission from Ref. [54]. Copyright 2021 Wiley.

Figure 3.
Hierarchy of building units and microaggregates in soil. Reprinted with permission from Ref. [57]. Copyright 2018 Wiley.
Figure 3.
Hierarchy of building units and microaggregates in soil. Reprinted with permission from Ref. [57]. Copyright 2018 Wiley.

Figure 4.
Relationship between soil pH and microbial carbon use efficiency (upper panel) and soil pH and exchangeable Al (lower panel). Values represent means ± SEM. Different letters at the top of each panel represent significant differences between soil pH categories at the p < 0.05 level. The average number of values in each pH category is 68. The dotted line represents the soil pH value point at which there is a transition in the response of both CUE and exchangeable Al. Reprinted with permission from Ref. [98]. Copyright 2019 Elsevier.
Figure 4.
Relationship between soil pH and microbial carbon use efficiency (upper panel) and soil pH and exchangeable Al (lower panel). Values represent means ± SEM. Different letters at the top of each panel represent significant differences between soil pH categories at the p < 0.05 level. The average number of values in each pH category is 68. The dotted line represents the soil pH value point at which there is a transition in the response of both CUE and exchangeable Al. Reprinted with permission from Ref. [98]. Copyright 2019 Elsevier.

Figure 5.
Conceptual diagram for the influence of soil moisture change on processes controlling soil microbial metabolism during the secondary succession. Reprinted with permission from Ref. [104]. Copyright 2020 Elsevier.
Figure 5.
Conceptual diagram for the influence of soil moisture change on processes controlling soil microbial metabolism during the secondary succession. Reprinted with permission from Ref. [104]. Copyright 2020 Elsevier.

Figure 6.
Schematic diagram of microbial−mediated free radical catalysis reaction and its impact on soil carbon stability and storage. Reprinted with permission from Ref. [146]. Copyright 2019 Wiley.
Figure 6.
Schematic diagram of microbial−mediated free radical catalysis reaction and its impact on soil carbon stability and storage. Reprinted with permission from Ref. [146]. Copyright 2019 Wiley.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of nitrogen on the turnover process of POC and MAOC.
| SOC Types | Process | Category | Promotional Effects | Inhibitory Effects |
|---|---|---|---|---|
| POC | Plant litter | Quantity | NPP elevation due to increased N factor effectiveness. | Soil acidification triggers heavy metal toxicity leading to lower NPP. |
| Quality | N elements cause elevated organic matter content of structural complexity. | Lower C/N of plant litter. | ||
| Physical protection | Aggregates | Increasing the amount of organic cementing substances promotes the formation of aggregates. The entanglement of plant roots increases and thus promotes the formation of aggregates. Soil acidification makes Fe-Al oxides positively charged and thus promotes aggregate formation. | Soil acidification causes extensive depletion of inorganic carbon and reduces its cementation. | |
| MAOC | CUE | Microbial community structure | The dominant role of fungal communities under soil acidification conditions produced high CUE. | Elevated N efficiency shifts the microbial community structure toward bacterial dominance and negatively affects CUE. |
| Ecological Chemometrics | Microorganisms allocate more C to maintain their stoichiometric balance, resulting in higher CUE. | The increased effectiveness of N elements led to increased C limitation, inhibition of microbial growth, and decreased CUE. | ||
| Substrate quality and bioavailability | The substrate decrease leads to the increase in microbial extracellular enzyme secretion and the reduction in microbial CUE. | |||
| Mineral–organic complexes | Soil minerals | Soil acidification makes iron and aluminum oxides have positive charges, promoting the formation of mineral-organic complexes. | Oxalic acid in root exudates releases organic compounds from the protective association with minerals. | |
| Bond bridge | Organic nitrogen fertilizers promote the formation of hydroxyl radicals (HO·) and increase the appearance of intermolecular covalent bonds and mineral-organic complexes in SOC. | Soil acidification produces competition between H+ and exchangeable base cations for adsorption sites (neutral and alkaline soils). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meng, D.; Cheng, H.; Shao, Y.; Luo, M.; Xu, D.; Liu, Z.; Ma, L. Progress on the Effect of Nitrogen on Transformation of Soil Organic Carbon. Processes 2022, 10, 2425. https://doi.org/10.3390/pr10112425
AMA Style
Meng D, Cheng H, Shao Y, Luo M, Xu D, Liu Z, Ma L. Progress on the Effect of Nitrogen on Transformation of Soil Organic Carbon. Processes. 2022; 10(11):2425. https://doi.org/10.3390/pr10112425
Chicago/Turabian StyleMeng, Danyang, Hangxin Cheng, Yang Shao, Min Luo, Diandou Xu, Zhiming Liu, and Lingling Ma. 2022. "Progress on the Effect of Nitrogen on Transformation of Soil Organic Carbon" Processes 10, no. 11: 2425. https://doi.org/10.3390/pr10112425
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.