
Arduino Soft Sensor for Monitoring Schizochytrium sp. Fermentation, a Proof of Concept for the Industrial Application of Genome-Scale Metabolic Models in the Context of Pharma 4.0

Abstract
:1. Introduction
- A method for adapting GEM estimations into an online soft sensor for culture fermentation monitoring has not yet been reported.
- No research has stated if a soft sensor can be implemented directly into low-cost electronic boards instead of computers.
- No reported mathematical model has been able to describe the effect of dissolved oxygen on biomass growth and DHA accumulation of Schizochytrium sp.
2. Materials and Methods
2.1. Mathematical Model for Schizochytrium Carbon-Fed-Batch Fermentations
2.2. Kinetic Relationships on Thraustochytrid Culture Described by a GEM
2.3. Evolutionary Algorithm for Simplified Kinetic Relations
2.4. Reported Data Used for Parameter Fitting
2.5. Soft Sensor Testing on Electronic Board
3. Results
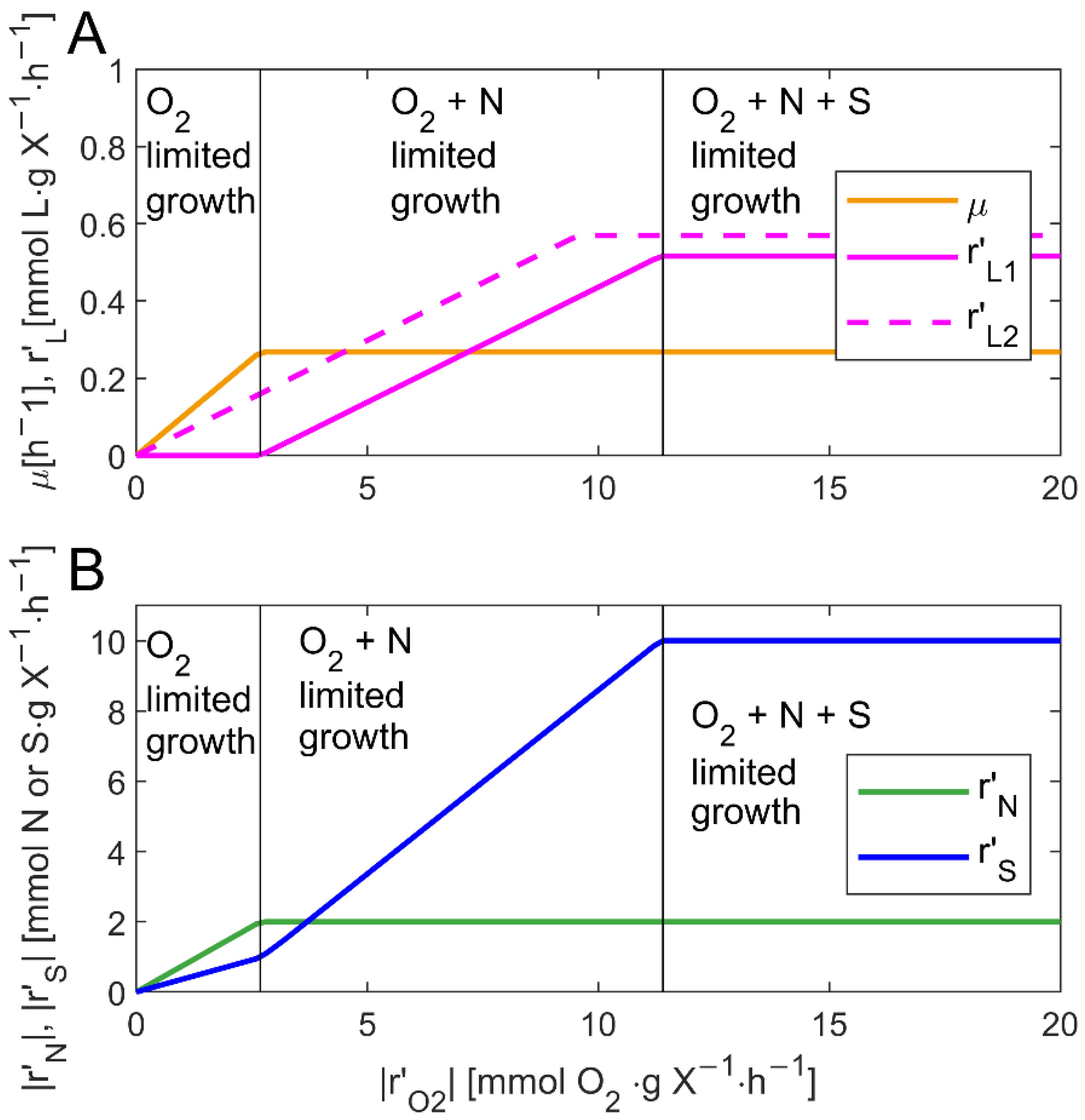
3.1. Effect of Nutrient and Oxygen Consumption on the Growth and Lipid Synthesis by Thraustochytrids Analyzed Using GEM
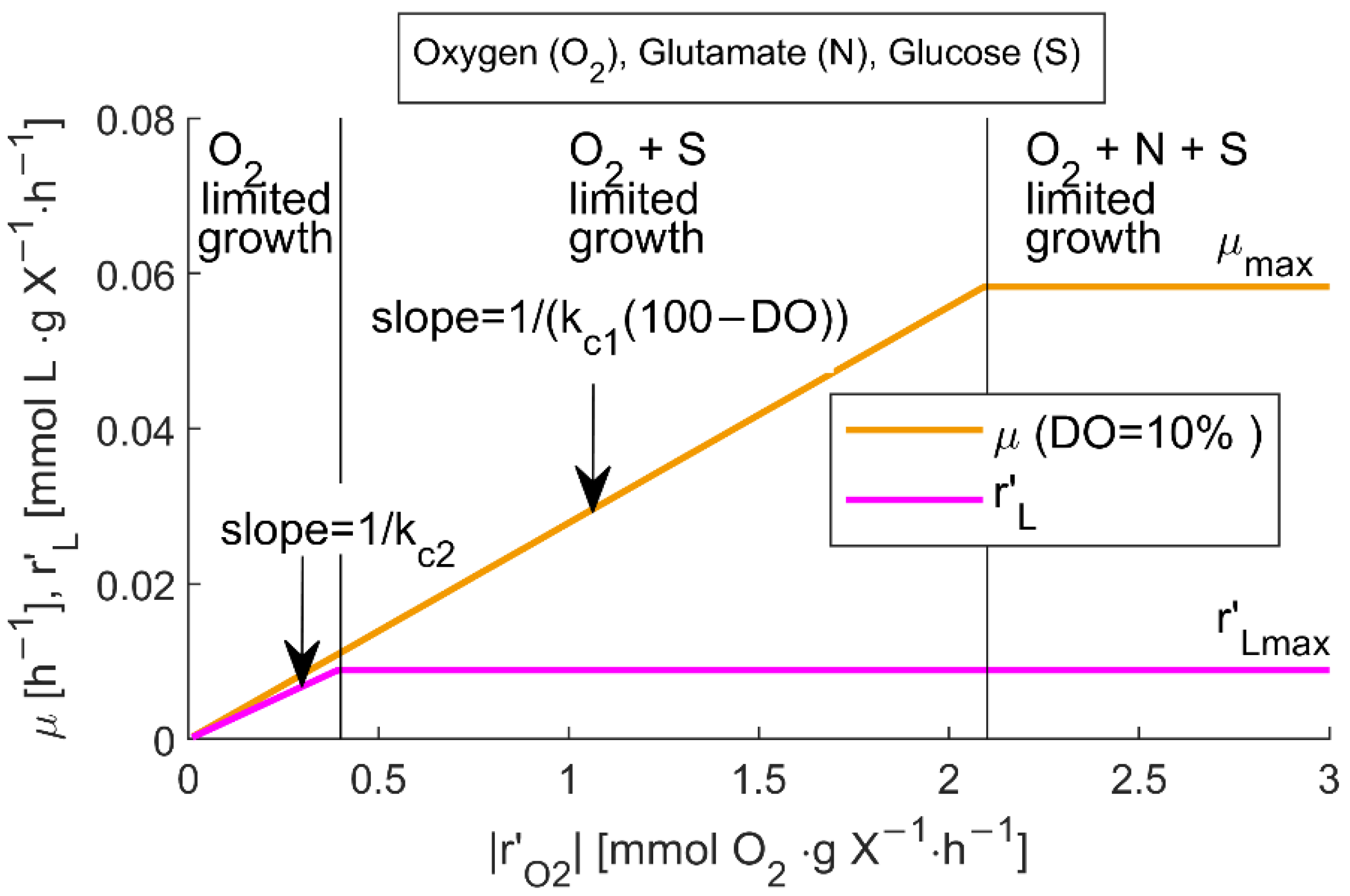
3.2. Mathematical Model for the Soft Sensor Using Simplified GEM Results
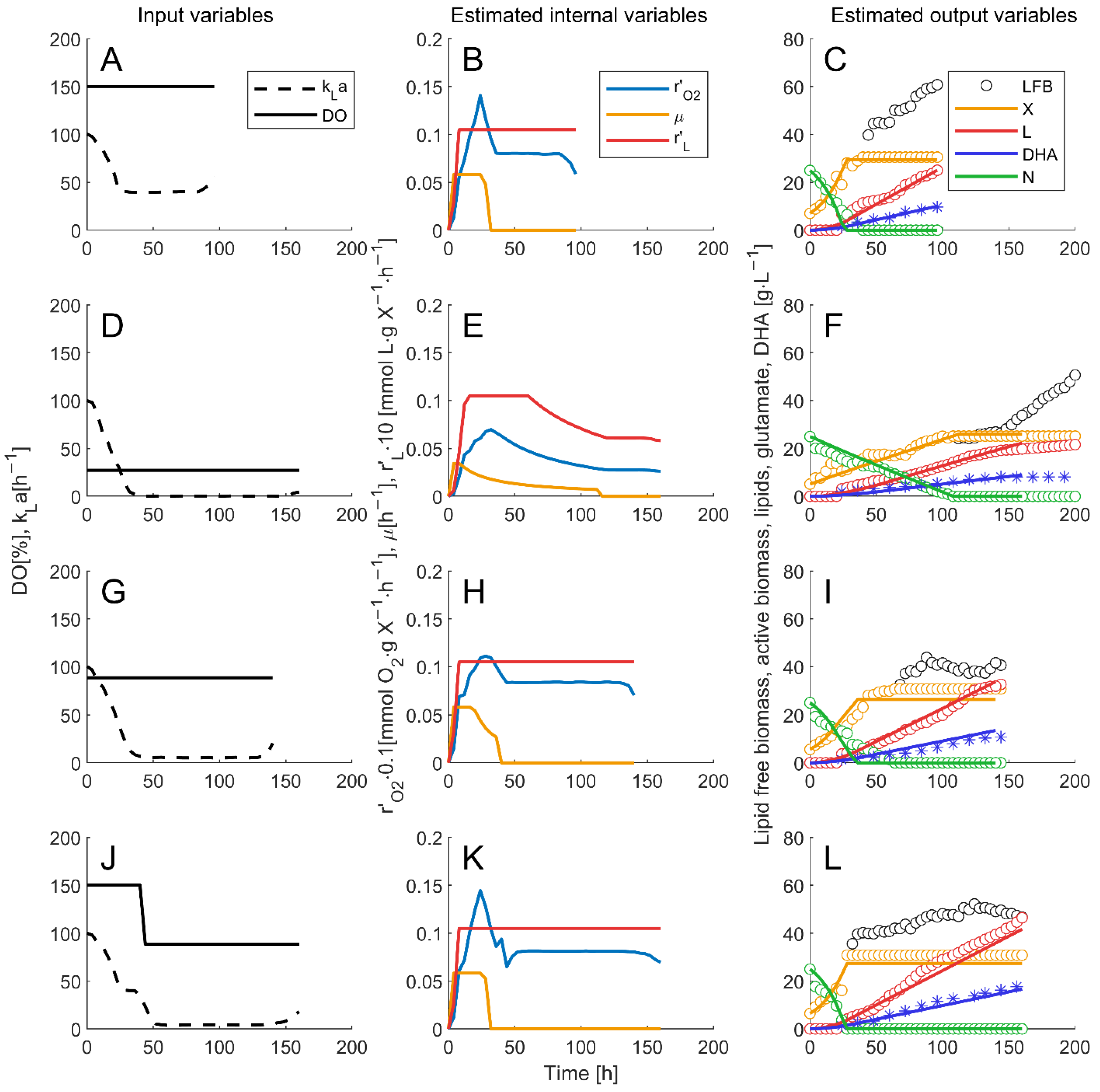
3.3. Soft Sensor Implemented in Arduino UNO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Narayanan, H.; Luna, M.F.; von Stosch, M.; Cruz Bournazou, M.N.; Polotti, G.; Morbidelli, M.; Butté, A.; Sokolov, M. Bioprocessing in the Digital Age: The Role of Process Models. Biotechnol. J. 2020, 15, 1900172. [Google Scholar] [CrossRef] [PubMed]
- Nargund, S.; Guenther, K.; Mauch, K. The Move toward Biopharma 4.0. Genet. Eng. Biotechnol. News 2019, 39, 53–55. [Google Scholar] [CrossRef]
- Alarcon, C.; Shene, C. Fermentation 4.0, a Case Study on Computer Vision, Soft Sensor, Connectivity, and Control Applied to the Fermentation of a Thraustochytrid. Comput. Ind. 2021, 128, 103431. [Google Scholar] [CrossRef]
- Tokuyama, K.; Shimodaira, Y.; Kodama, Y.; Matsui, R.; Kusunose, Y.; Fukushima, S.; Nakai, H.; Tsuji, Y.; Toya, Y.; Matsuda, F.; et al. Soft-Sensor Development for Monitoring the Lysine Fermentation Process. J. Biosci. Bioeng. 2021, 132, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Kusuda, M.; Shimizu, H.; Toya, Y. Reactor Control System in Bacterial Co-Culture Based on Fluorescent Proteins Using an Arduino-Based Home-Made Device. Biotechnol. J. 2021, 16, 2100169. [Google Scholar] [CrossRef] [PubMed]
- Wilken, S.E.; Leggieri, P.A.; Kerdman-Andrade, C.; Reilly, M.; Theodorou, M.K.; O’Malley, M.A. An Arduino Based Automatic Pressure Evaluation System to Quantify Growth of Non-Model Anaerobes in Culture. AIChE J. 2020, 66, e16540. [Google Scholar] [CrossRef]
- Curreri, F.; Patanè, L.; Xibilia, M.G. Soft Sensor Transferability: A Survey. Appl. Sci. 2021, 11, 7710. [Google Scholar] [CrossRef]
- Biechele, P.; Busse, C.; Solle, D.; Scheper, T.; Reardon, K. Sensor Systems for Bioprocess Monitoring. Eng. Life Sci. 2015, 15, 469–488. [Google Scholar] [CrossRef]
- Wang, B.; Shahzad, M.; Zhu, X.; Ur Rehman, K.; Ashfaq, M.; Abubakar, M. Soft-Sensor Modeling for l-Lysine Fermentation Process Based on Hybrid ICS-MLSSVM. Sci. Rep. 2020, 10, 11630. [Google Scholar] [CrossRef] [PubMed]
- Wechselberger, P.; Sagmeister, P.; Herwig, C. Real-Time Estimation of Biomass and Specific Growth Rate in Physiologically Variable Recombinant Fed-Batch Processes. Bioprocess Biosyst. Eng. 2013, 36, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Winckler, S.; Krueger, R.; Schnitzler, T.; Zang, W.; Fischer, R.; Biselli, M. A Sensitive Monitoring System for Mammalian Cell Cultivation Processes: A PAT Approach. Bioprocess Biosyst. Eng. 2014, 37, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, R.; Mandenius, C.F. Soft Sensor Control of Metabolic Fluxes in a Recombinant Escherichia Coli Fed-Batch Cultivation Producing Green Fluorescence Protein. Bioprocess Biosyst. Eng. 2013, 36, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Pan, F. A Soft Sensor Modelling of Biomass Concentration during Fermentation Using Accurate Incremental Online V-Support Vector Regression Learning Algorithm. Am. J. Biochem. Biotechnol. 2015, 11, 149–159. [Google Scholar] [CrossRef]
- Imtiaz, U.; Jamuar, S.S.; Sahu, J.N.; Ganesan, P.B. Bioreactor Profile Control by a Nonlinear Auto Regressive Moving Average Neuro and Two Degree of Freedom PID Controllers. J. Process Control 2014, 24, 1761–1777. [Google Scholar] [CrossRef]
- Baart, G.J.E.; Martens, D.E. Genome-Scale Metabolic Models: Reconstruction and Analysis. In Methods in Molecular Biology; Humana Press: Totova, NJ, USA, 2012; Volume 799, pp. 107–126. ISBN 9781617793455. [Google Scholar]
- Zhang, C.; Hua, Q. Applications of Genome-Scale Metabolic Models in Biotechnology and Systems Medicine. Front. Physiol. 2016, 6, 413. [Google Scholar] [CrossRef] [Green Version]
- Orth, J.D.; Thiele, I.; Palsson, B.O. What Is Flux Balance Analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef]
- Kuriya, Y.; Inoue, M.; Yamamoto, M.; Murata, M.; Araki, M. Knowledge Extraction from Literature and Enzyme Sequences Complements FBA Analysis in Metabolic Engineering. Biotechnol. J. 2021, 16, 2000443. [Google Scholar] [CrossRef]
- O’Brien, E.J.; Monk, J.M.; Palsson, B.O. Using Genome-Scale Models to Predict Biological Capabilities. Cell 2015, 161, 971–987. [Google Scholar] [CrossRef] [Green Version]
- Mahadevan, R.; Schilling, C.H. The Effects of Alternate Optimal Solutions in Constraint-Based Genome-Scale Metabolic Models. Metab. Eng. 2003, 5, 264–276. [Google Scholar] [CrossRef]
- Mahadevan, R.; Edwards, J.S.; Doyle, F.J. Dynamic Flux Balance Analysis of Diauxic Growth in Escherichia Coli. Biophys. J. 2002, 83, 1331–1340. [Google Scholar] [CrossRef]
- Kuriya, Y.; Araki, M. Dynamic Flux Balance Analysis to Evaluate the Strain Production Performance on Shikimic Acid Production in Escherichia Coli. Metabolites 2020, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, W.L.; Saha, R. Introducing an Optimization- and Explicit Runge-Kutta-Based Approach to Perform Dynamic Flux Balance Analysis. Sci. Rep. 2020, 10, 9241. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Heiner, M.; Gilbert, D. Hybrid Modelling of Biological Systems: Current Progress and Future Prospects. Brief. Bioinform. 2022, 23, bbac081. [Google Scholar] [CrossRef] [PubMed]
- Dale, J.M.; Popescu, L.; Karp, P.D. Machine Learning Methods for Metabolic Pathway Prediction. BMC Bioinform. 2010, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Pearcy, N.; Hu, Y.; Baker, M.; Maciel-Guerra, A.; Xue, N.; Wang, W.; Kaler, J.; Peng, Z.; Li, F.; Dottorini, T. Genome-Scale Metabolic Models and Machine Learning Reveal Genetic Determinants of Antibiotic Resistance in Escherichia Coli and Unravel the Underlying Metabolic Adaptation Mechanisms. mSystems 2021, 6, e00913-20. [Google Scholar] [CrossRef]
- Wu, S.G.; Wang, Y.; Jiang, W.; Oyetunde, T.; Yao, R.; Zhang, X.; Shimizu, K.; Tang, Y.J.; Bao, F.S. Rapid Prediction of Bacterial Heterotrophic Fluxomics Using Machine Learning and Constraint Programming. PLoS Comput. Biol. 2016, 12, e1004838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frioux, C.; Huet, S.; Labarthe, S.; Martinelli, J.; Sherman, D.J.; Taupin, M.; Ugalde-salas, P.; Frioux, C.; Huet, S.; Labarthe, S.; et al. Accelerating Metabolic Models Evaluation with Statistical Metamodels: Application to Salmonella Infection Models. 2022. Available online: https://hal.inrae.fr/hal-03635862 (accessed on 22 July 2022).
- Richelle, A.; David, B.; Demaegd, D.; Dewerchin, M.; Kinet, R.; Morreale, A.; Portela, R.; Zune, Q.; von Stosch, M. Towards a Widespread Adoption of Metabolic Modeling Tools in Biopharmaceutical Industry: A Process Systems Biology Engineering Perspective. npj Syst. Biol. Appl. 2020, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Xie, N.; Sen, B.; Song, Z.; Zhao, Y.; Chen, Z.; Shi, W.; Zhang, Y.; Zhang, J.; Johnson, Z.I.; Wang, G. High Phylogenetic Diversity and Abundance Pattern of Labyrinthulomycete Protists in the Coastal Waters of the Bohai Sea. Environ. Microbiol. 2018, 20, 3042–3056. [Google Scholar] [CrossRef] [PubMed]
- Finco, A.M.D.O.; Mamani, L.D.G.; Carvalho, J.C.D.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Soccol, C.R. Technological Trends and Market Perspectives for Production of Microbial Oils Rich in Omega-3. Crit. Rev. Biotechnol. 2017, 37, 656–671. [Google Scholar] [CrossRef] [PubMed]
- Bekatorou, A.; Dima, A.; Tsafrakidou, P.; Boura, K.; Lappa, K.; Kandylis, P.; Pissaridi, K.; Kanellaki, M.; Koutinas, A.A. Downstream Extraction Process Development for Recovery of Organic Acids from a Fermentation Broth. Bioresour. Technol. 2016, 220, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Yang, Y.-T. Combining Engineering Strategies and Fermentation Technology to Enhance Docosahexaenoic Acid (DHA) Production from an Indigenous Thraustochytrium sp. BM2 Strain. Biochem. Eng. J. 2018, 133, 179–185. [Google Scholar] [CrossRef]
- Kim, H.; Choi, O.; Seung, B.; Choe, W.; Sang, B. Bioresource Technology Impact of Feedstocks and Downstream Processing Technologies on the Economics of Caproic Acid Production in Fermentation by Megasphaera Elsdenii T81. Bioresour. Technol. 2020, 301, 122794. [Google Scholar] [CrossRef]
- Nazir, Y.; Shuib, S.; Kalil, M.S.; Song, Y.; Hamid, A.A. Optimization of Culture Conditions for Enhanced Growth, Lipid and Docosahexaenoic Acid (DHA) Production of Aurantiochytrium SW1 by Response Surface Methodology. Sci. Rep. 2018, 8, 8909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nor, M.; Sohedein, A.; Abd, W.; Qadr, A. Optimisation of Biomass and Lipid Production of a Tropical Thraustochytrid Aurantiochytrium sp. UMACC-T023 in Submerged-Liquid Fermentation for Large-Scale Biodiesel Production Biocatalysis and Agricultural Biotechnology Optimisation of Biomass and Lipid. Biocatal. Agric. Biotechnol. 2020, 23, 101496. [Google Scholar] [CrossRef]
- Ye, H.; He, Y.; Xie, Y.; Sen, B.; Wang, G. Fed-Batch Fermentation of Mixed Carbon Source Significantly Enhances the Production of Docosahexaenoic Acid in Thraustochytriidae sp. PKU#Mn16 by Differentially Regulating Fatty Acids Biosynthetic Pathways. Bioresour. Technol. 2020, 297, 122402. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ji, X.-J.; Ren, L.-J.; Nie, Z.-K.; Feng, Y.; Wu, W.-J.; Ouyang, P.-K.; Huang, H. Enhancement of Docosahexaenoic Acid Production by Schizochytrium sp. Using a Two-Stage Oxygen Supply Control Strategy Based on Oxygen Transfer Coefficient. Lett. Appl. Microbiol. 2011, 52, 22–27. [Google Scholar] [CrossRef]
- Qu, L.; Ren, L.J.; Huang, H. Scale-up of Docosahexaenoic Acid Production in Fed-Batch Fermentation by Schizochytrium sp. Based on Volumetric Oxygen-Transfer Coefficient. Biochem. Eng. J. 2013, 77, 82–87. [Google Scholar] [CrossRef]
- Heggeset, T.M.B.; Ertesvåg, H.; Liu, B.; Ellingsen, T.E.; Vadstein, O.; Aasen, I.M. Lipid and DHA-Production in Aurantiochytrium sp.—Responses to Nitrogen Starvation and Oxygen Limitation Revealed by Analyses of Production Kinetics and Global Transcriptomes. Sci. Rep. 2019, 9, 19470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.J.; Sun, G.N.; Ji, X.J.; Hu, X.C.; Huang, H. Compositional Shift in Lipid Fractions during Lipid Accumulation and Turnover in Schizochytrium sp. Bioresour. Technol. 2014, 157, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.S.; Ji, X.J.; Ren, L.J.; Li, G.L.; Yin, F.W.; Huang, H. Development of a Real-Time Bioprocess Monitoring Method for Docosahexaenoic Acid Production by Schizochytrium sp. Bioresour. Technol. 2016, 216, 422–427. [Google Scholar] [CrossRef]
- Wang, S.; Wan, W.; Wang, Z.; Zhang, H.; Liu, H.; Arunakumara, K.K.I.U.; Cui, Q.; Song, X. A Two-Stage Adaptive Laboratory Evolution Strategy to Enhance Docosahexaenoic Acid Synthesis in Oleaginous Thraustochytrid. Front. Nutr. 2021, 8, 795491. [Google Scholar] [CrossRef]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strøm, A.R. Accumulation of Docosahexaenoic Acid-Rich Lipid in Thraustochytrid aurantiochytrium sp. Strain T66: Effects of N and P Starvation and O2 Limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Shene, C.; Paredes, P.; Flores, L.; Leyton, A.; Asenjo, J.A.; Chisti, Y. Dynamic Flux Balance Analysis of Biomass and Lipid Production by Antarctic Thraustochytrid oblongichytrium sp. RT2316-13. Biotechnol. Bioeng. 2020, 117, 3006–3017. [Google Scholar] [CrossRef]
- Orth, J.D.; Conrad, T.M.; Na, J.; Lerman, J.A.; Nam, H.; Feist, A.M.; Palsson, B. A Comprehensive Genome-Scale Reconstruction of Escherichia coli Metabolism—2011. Mol. Syst. Biol. 2011, 7, 535. [Google Scholar] [CrossRef] [PubMed]
- De Martino, D.; Capuani, F.; Mori, M.; De Martino, A.; Marinari, E. Counting and Correcting Thermodynamically Infeasible Flux Cycles in Genome-Scale Metabolic Networks. Metabolites 2013, 3, 946–966. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.H.J.; Cai, J.; Wang, L.; Simons-Senftle, M.N.; Maranas, C.D. Standardizing Biomass Reactions and Ensuring Complete Mass Balance in Genome-Scale Metabolic Models. Bioinformatics 2017, 33, 3603–3609. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ren, L.-J.; Sun, G.-N.; Ji, X.-J.; Nie, Z.-K.; Huang, H. Batch, Fed-Batch and Repeated Fed-Batch Fermentation Processes of the Marine Thraustochytrid schizochytrium sp. for Producing Docosahexaenoic Acid. Bioprocess Biosyst. Eng. 2013, 36, 1905–1912. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Kumelj, T.; Karlsen, E.; Almaas, E. Genome-Scale Metabolic Modelling When Changes in Environmental Conditions Affect Biomass Composition. PLoS Comput. Biol. 2021, 17, e1008528. [Google Scholar] [CrossRef] [PubMed]
- Murugan, C.; Natarajan, P. Estimation of Fungal Biomass Using Multiphase Artificial Neural Network Based Dynamic Soft Sensor. J. Microbiol. Methods 2019, 159, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Barrigón, J.M.; Ramon, R.; Rocha, I.; Valero, F.; Ferreira, E.C.; Montesinos, J.L. State and Specific Growth Estimation in Heterologous Protein Production by Pichia Pastoris. AIChE J. 2012, 58, 2966–2979. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Exp | kLa (h−1) | Model Parameters * | Xerror (%) | Nerror (%) | Lerror (%) | DHAerror (%) |
|---|---|---|---|---|---|---|
| I | 150.1 | µmax = 0.0583 rLmax = 0.0089 kc1 = 0.3988 kc2 = 44.1433 kDHA = 0.3730 YX/N = 0.8053 | 1.1 | 1.7 | 1.8 | 3.3 |
| II | 27.3 | 1.2 | 1.2 | 0.8 | 2.1 | |
| III | 88.5 | 2.3 | 2.0 | 0.7 | 3.9 | |
| IV | step 150.1 to 88.5 at 40 h | 1.8 | 0.8 | 1.2 | 3.7 | |
| Average ± std | 1.1 ± 0.4 | 1.3 ± 0.4 | 1.1 ± 0.5 | 3.2 ± 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alarcon, C.; Shene, C. Arduino Soft Sensor for Monitoring Schizochytrium sp. Fermentation, a Proof of Concept for the Industrial Application of Genome-Scale Metabolic Models in the Context of Pharma 4.0. Processes 2022, 10, 2226. https://doi.org/10.3390/pr10112226
Alarcon C, Shene C. Arduino Soft Sensor for Monitoring Schizochytrium sp. Fermentation, a Proof of Concept for the Industrial Application of Genome-Scale Metabolic Models in the Context of Pharma 4.0. Processes. 2022; 10(11):2226. https://doi.org/10.3390/pr10112226
Chicago/Turabian StyleAlarcon, Claudio, and Carolina Shene. 2022. "Arduino Soft Sensor for Monitoring Schizochytrium sp. Fermentation, a Proof of Concept for the Industrial Application of Genome-Scale Metabolic Models in the Context of Pharma 4.0" Processes 10, no. 11: 2226. https://doi.org/10.3390/pr10112226